
Fluctuating Asymmetry of Human Populations: A Review

1
Biology Department, Berry College, Mount Berry, GA 30149, USA
2
Department of Anthropology, Istanbul University, Istanbul 34452, Turkey
*
Author to whom correspondence should be addressed.
Symmetry 2016, 8(12), 154; https://doi.org/10.3390/sym8120154
Submission received: 23 September 2016
/
Revised: 14 November 2016
/
Accepted: 12 December 2016
/
Published: 16 December 2016
(This article belongs to the Special Issue Fluctuating Asymmetry 2016)
{kind=link}
{kind=link}
{kind=link}
Abstract
:Fluctuating asymmetry, the random deviation from perfect symmetry, is a widely used population-level index of developmental instability, developmental noise, and robustness. It reflects a population’s state of adaptation and genomic coadaptation. Here, we review the literature on fluctuating asymmetry of human populations. The most widely used bilateral traits include skeletal, dental, and facial dimensions; dermatoglyphic patterns and ridge counts; and facial shape. Each trait has its advantages and disadvantages, but results are most robust when multiple traits are combined into a composite index of fluctuating asymmetry (CFA). Both environmental (diet, climate, toxins) and genetic (aneuploidy, heterozygosity, inbreeding) stressors have been linked to population-level variation in fluctuating asymmetry. In general, these stressors increase average fluctuating asymmetry. Nevertheless, there have been many conflicting results, in part because (1) fluctuating asymmetry is a weak signal in a sea of noise; and (2) studies of human fluctuating asymmetry have not always followed best practices. The most serious concerns are insensitive asymmetry indices (correlation coefficient and coefficient of indetermination), inappropriate size scaling, unrecognized mixture distributions, inappropriate corrections for directional asymmetry, failure to use composite indices, and inattention to measurement error. Consequently, it is often difficult (or impossible) to compare results across traits, and across studies.
1. Introduction
Research on the developmental instability of human populations, as estimated by fluctuating asymmetry, has blossomed only in the past 35 years. The first papers explicitly mentioning human fluctuating asymmetry were Adams and Niswander’s [1] study of fluctuating asymmetry in individuals born with cleft palate and Bailit et al.’s [2] study of four human populations experiencing different levels of environmental and genetic stress. Both of these studies followed Mather’s [3] and Van Valen’s [4] classic papers, which grew out of foundational work by Astauroff [5], Ludwig [6], and Waddington [7]. Since 1980, the number of papers focusing on human fluctuating asymmetry has grown exponentially with each decade. Originally the domain of population geneticists and evolutionary biologists studying Drosophila, Mus, and other species of plants and animals, nearly 60 percent of the more than 900 papers published in 2015 that mentioned fluctuating asymmetry in their titles, their text, or as a keyword, also mentioned humans. This spate of papers is mostly in the fields of anthropology, medicine, and evolutionary psychology. Human fluctuating asymmetry is an exciting, and often controversial, area of research. Exciting, because it involves our species, and controversial, because it sometimes makes surprising claims about our species.
Fluctuating asymmetry is a population-level measure of developmental instability, developmental noise, and robustness [3,6,7,8,9,10,11,12]. It is influenced principally by environmental and genetic stressors [12,13,14], but also arises spontaneously from random and nonlinear developmental processes [15,16,17,18], and the failures of distributed robustness [19,20,21].
There are three kinds of population asymmetries [4]: fluctuating asymmetry, directional asymmetry, and antisymmetry. Fluctuating asymmetry is the random developmental variation of a trait (or character) that is perfectly symmetrical, on average. It is estimated as the variance of the difference between right and left sides, Var(di), where di = Ri − Li and Ri is the value of a trait on the right side and Li is the value of the same trait on the left side of individual i. Fluctuating asymmetry has a symmetrical distribution of values around a mean of zero (µd = 0). Directional asymmetry, on the other hand, has a mean not equal to zero (µd ≠ 0). Handedness in humans is a classic example; most humans are right handed and show greater development of the long bones of their right arms (µd > 0). Finally, antisymmetry is a kind of asymmetry in which the mean of d is zero, or close to it, but the distribution around the mean is either platykurtic or bimodal. The claws of lobsters and crayfish are antisymmetric; 50% of the population have enlarged right claws and 50% have enlarged left claws. Few individuals have claws of the same size.
2. Measuring Fluctuating Asymmetry
Although fluctuating asymmetry is most easily, and conceptually, understood as Var(di) = Var(Ri − Li), it makes more sense in practice to use the mean absolute deviation of d as the index of fluctuating asymmetry [8,12,22,23]. The mean absolute deviation is the expectation of the absolute value of d, or E|di| = E|Ri − Li|. It is equivalent to Levene’s test for the differences between two variances. Means are more robustly compared than variances, since they are amenable to analysis of variance, regression, and other linear models [8]. They are also amenable to a variety of transformations, such as log transforms, E|log R − log L| = E|log(R/L)|, and power transforms, E [(Rλ − 1)/λ] − [(Lλ − 1)/λ].
Individual asymmetry (d) of a trait (or character) is usually estimated from measurements of linear dimensions, or counts, on right and left sides of the body. Shape asymmetry, involving either matching symmetry or object symmetry [24,25,26] can be estimated from landmarks or semi-landmarks in either two or three dimensions [25,26,27,28,29,30,31,32], as well as continuous symmetry measures [12,33,34,35]. Despite the apparent simplicity of fluctuating asymmetry, careless researchers can easily reach erroneous conclusions [8,12,22,23,36].
Not all indices of fluctuating asymmetry are equivalent. Indices based upon the correlation coefficient [37], for example, and the related coefficient of indetermination [4,38], are the least powerful [8,39]. The correlation coefficient is strongly influenced by the range of observations. A population can appear to be more asymmetric, on average, than another simply because the range of variation is smaller. Remarkably, these two indices were still in use as recently as 2011 [40], mostly in studies of human dermatoglyphic asymmetry. We recommend indices based on (1) the variance of d, such as the mean absolute deviation [8,12]; (2) the Procrustes distance of geometric mophometrics [25,26,28,41]; (3) dense surface modeling [42]; and (4) continuous symmetry measures [33,35].
More importantly, positive size scaling is rarely handled properly in studies of human fluctuating asymmetry. Failure to understand the behavior of additive and multiplicative errors can easily lead to erroneous conclusions. Multiplicative error is generated by actively growing tissue, in which old tissue gives rise to new tissue [12,43,44]. The probability distribution of a trait exhibiting multiplicative error is generally lognormal. Positive size scaling of fluctuating asymmetry, in which asymmetry increases with size (compare mouse and elephant ears), is generally a consequence of such active growth. Moreover, leptokurtic asymmetry distributions may also arise from multiplicative errors, because the probability distribution of the difference between two lognormal distributions is leptokurtic [43]. (This is why we define fluctuating asymmetry as symmetrically distributed, not normally distributed, variation around a mean of zero.) In contrast to the active growth model, inert structures, such as nails, feathers, dermatoglyphic ridge counts, and probably teeth, usually exhibit additive errors. Correction for size scaling is inappropriate for such traits, because there should not be any size scaling to correct (Figure 1). Finally, measurement error is also additive.
For traits composed of actively growing tissue, asymmetry variation is always a mixture of additive and multiplicative errors, with most of the additive part being measurement error. If measurement error is large, then correcting for positive size scaling by dividing |R − L| by (R + L) or using |log R − log L|, can easily generate even greater negative size scaling (Figure 1). This is especially problematic with teeth and dermatoglyphic ridge counts, which develop as inert structures. In any case, correction for size scaling always needs to be justified. In some subfields of human fluctuating asymmetry, researchers rarely justify the size scaling they employ. To date, no one has conducted a thorough survey of the growth models applicable to commonly used human traits.
3. Human Symmetries and Asymmetries
The external human body is bilaterally symmetrical for the most part, at least at birth. Directional asymmetries may develop later, a consequence of nervous system laterality and bone remodeling. Most of the internal organs (heart, lungs, kidneys, stomach, etc.) and brain are directionally asymmetric.
3.1. Skeletal Asymmetry
Skeletal asymmetry has a long history in studies of fluctuating asymmetry, especially in paleoanthropological studies. Bones are relatively easy to measure, and even in living subjects they can be measured non-invasively with x-rays [46]. Moreover, upper limb mass can be estimated from joystick movements [47]. Nevertheless, despite their widespread use, there are two problems associated with bone: directional asymmetry and bone remodeling.
There is considerable directional asymmetry in the human skeleton. Directional asymmetry can inflate estimates of fluctuating asymmetry, even when some indices that purport to remove it are used [48]. Long bones of the upper limbs, for example, have a right bias in most populations [49,50,51]. Furthermore, parts of the cranial skeleton can also be directionally asymmetric [32,52,53]. One can either remove the directional component [48,54] or decompose a mixture distribution into fluctuating, directional, and antisymmetric components [55]. These two approaches, however, make the implicit assumption that all individuals in a population have the same degree of directional asymmetry, which may not be true.
The second, and most problematic issue, is that bones experiencing mechanical loading are subject to remodeling throughout the life of the individual [56]. Consequently, individual asymmetry of a trait can change over time [57]. In particular, mechanical loading on one side of the body can magnify directional asymmetries. Nevertheless, bone remodeling can sometimes be advantageous, for example when one’s goal is to infer cultural transitions in the archeological record. Mechanical loading through tool use, for example, can exacerbate directional asymmetry of the upper limbs. Macintosh et al. [51], for example, have inferred that changes in the upper limb asymmetry of women from the Bronze Age to the Iron Age were caused by the introduction and use of the ard and plow in agriculture.
Bone is an actively growing tissue; consequently, errors should be largely multiplicative [43,44]. If additive measurement error is high, the resulting mixture distribution can create problems, such as inverse size scaling with certain transformations, such as |log R − log L|. One should always check that asymmetry does not increase (or decrease) with size. Moreover, size scaling needs to be reevaluated after every transformation.
According to Hallgrimsson [57], skeletal asymmetry (even after scaling for size) increases throughout the life of the individual. He attributed this to asymmetrical remodeling, locally variable growth, or undirected remodeling. (See also Palestis and Trivers [58].) Whatever the cause of the increasing asymmetry, age should probably be included as a covariate in all studies of fluctuating asymmetry.
3.2. Dental Asymmetry
Fluctuating asymmetry of teeth was pioneered by Leigh Van Valen [4], who studied the lengths and widths of upper teeth of extinct horses (Griphippus gratus = Pseudhipparion gratum Leidy 1869) and the upper and lower cheek teeth of white-footed mice (Peromyscus leucopus). In human populations, dental asymmetry has been especially popular with paleoanthropologists [59,60]. Bone is easily damaged during fossilization or burial, but teeth are resistant to deformation and fracture [61]. Nevertheless, paleoanthropologists can reliably assign teeth to right and left sides of the same individual only if the dental arch is intact (or partly intact).
For collections of fossilized human remains or skeletal populations from cemeteries, measurements can be made on the actual teeth. For samples of living individuals, however, measurements are generally made on dental casts.
Humans have 20 primary teeth and 32 permanent teeth. Although they are subject to wear, teeth are not subject to remodeling, like bone. Various measurements can be made on teeth, including buccolingual diameter of the lower first molars [1] and mesiodistal diameter of the upper and lower incisors [62] and of all the permanent teeth (excluding third molars, [2]).
Because teeth are inert structures, one would expect errors (natural variation) to be mostly additive [43,44]. Consequently, there should be little or no positive size scaling. In other words, fluctuating asymmetry should not increase with tooth size, unless it is doing so for a reason other than active growth. However, Garn [63,64] reported that tooth asymmetry of mesiodistal diameter increased with size across tooth types (incisors, canines, premolars, molars), but that asymmetry of buccolingual diameters did not. Ideally, this analysis needs to be done within a category of tooth. Barrett et al. [65] and Sciulli [66], for example, found no significant size scaling when they plotted |R − L| against (R + L) for pairs of individual kinds of teeth (M1, M2, P1, etc.). Most studies of dental asymmetry up to this time have corrected for size scaling (usually without justifying the correction). Indeed, both Barrett et al. [65] and Sciulli [66] went ahead and “corrected” for size-scaling anyway. Consequently, much of the literature on dental asymmetry may be flawed. Nevertheless, researchers should not simply assume that errors are additive (or multiplicative) when working with teeth. It always pays to plot |R − L| against (R + L); there should be no relationship between the two unless something biologically interesting is going on.
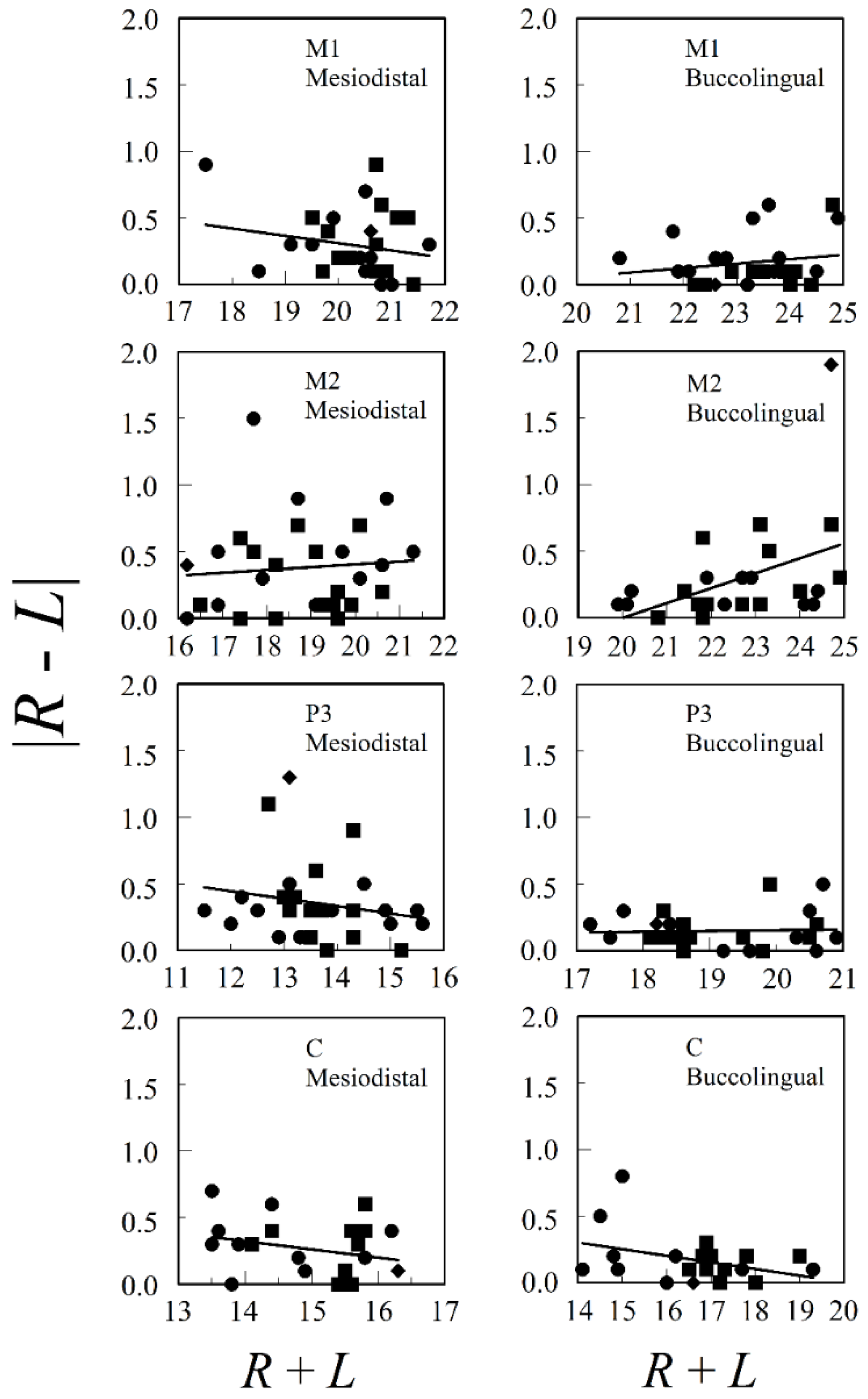
Figure 2 presents the relationships between both mesiodistal and buccolingual asymmetry and adult tooth size for M1, M2, P3, and C from the upper jaws of both males and females from a prehistoric village, Ban Chiang, in Northeast Thailand. Database E.4 was accessed from tDAR [67], the Digital Archeological Database. The only significant regression was for buccolingual M2 (F1, 26 = 6.001, p = 0.021, r2 = 0.188) and this was influenced mostly by a single outlier. There are no consistent positive relationships between asymmetry and size in the data, whether we looked at males and females separately or pooled. The mean correlation (±standard error) between |R − L| and R + L for the eight combinations of tooth (M1, M2, P3, or C) and tooth dimension (mediodistal or buccolingual) in the pooled sexes was , with n = 22–30.
In addition to linear measurements, landmark methods of geometric morphometrics have recently been used to study shape asymmetry of the dental arch [68].
3.3. Facial Asymmetry
Facial asymmetry is widely used in studies of sexual selection, under the assumption that mate choice is partly based upon a potential mate’s facial symmetry. Linear measurements, such as deviations of eye, ear, nostril, and cheek horizontal midpoints from the facial mid-line [69] are common, as are two- and three-dimensional imaging [30,70] and landmark methods for shape asymmetry (geometric morphometrics) [28,41].
3.4. Dermatoglyphic Asymmetry
Dermatoglyphic literally means “skin carving” [71]. It refers to the friction ridges on the fingers, palms, and feet [72,73,74]. These ridges form during weeks 10–16 of fetal development [75,76] and are not subject to environmental alterations after birth. Consequently, dermatoglyphic asymmetry can be used to measure developmental instability during a precise period of fetal development [77,78]. This is a significant advantage over many other traits, such as bones and soft tissues, because it allows one to infer precisely when development has been perturbed. For example, King et al. [79,80] found that mothers of children who experienced a natural disaster (an ice storm) during the critical period of gestation gave birth to children with greater dermatoglyphic asymmetry than those gestating earlier or later.
Dermatoglyphic variables are either qualitative (patterns formed by dermal ridges and creases) or quantitative (counts of ridges or measurements of angles) [38]. The most common traits used for right–left asymmetry are finger and palmar ridge counts, palmar atd angles [1,81], ridge patterns on the fingertips, and palmar flexion crease patterns. Finger ridge counts are made between the core of a pattern and the nearest triradius. Arches are assigned a ridge count of zero, because they lack a triradius.
There are at least two main ways of treating finger ridge counts. They can be treated finger by finger (i.e., homologous fingers are treated as separate traits) or all five fingers on each hand can be taken together, as a single trait. The simplest approach is to take the difference between the total ridge counts on each hand. For example,
where RI and LI through RV and LV are thumb through little finger on right and left hands, respectively. This approach, however, ignores the symmetry of homologous fingers. One can also take the absolute differences between each homologous finger and add them together, dividing by the total number of fingers to get an average ridge count difference.
|dc1| = |RTRC − LTRC| = |(RI + RII + RIII + RIV + RV) − (LI + LII + LIII + LIV + LV)|
|dc2| = (|RI − LI| + |RII − LII| + |RIII − LIII| + |RIV − LIV| + |RV − LV|)/5.
We recommend the second approach, because it incorporates more information than the first one.
Palmar ridge counts are made between the a, b, c, and d triradii. The a triradius is on the palm at the base of the index finger; the b triradius is at the base of the middle finger, and so on. There are potentially three sets of counts, the a–b, b–c, and c–d palmar ridge counts on right and left sides. Because c and d triradii are sometimes missing, however, the a–b ridge count is the one most commonly encountered [77,82,83]. It has the advantage over fingerprint ridge counts in that zero counts do not normally occur.
For atd angles, the measure of individual asymmetry (d) is straightforward, d = R − L or |d| = |R − L|. The atd angle changes with age, however, as the size and shape of the hand increases [84]. Consequently, it is important to compare individuals of similar ages.
Wang et al. [85] have introduced a new approach to dermatoglyphic asymmetry that resembles landmark methods of geometric morphometrics, but without landmarks. It compares the ridge orientation fields of homologous fingers on right and left hands. Digital images of homologous fingers are superimposed by their contours and then the right and left orientation fields are compared. The authors found this approach to be more sensitive than traditional approaches to right–left asymmetry in a sample of schizophrenic patients.
For ridge patterns (arches, radial loops, ulnar loops, and whorls), pattern discordance is used [38,86,87]. For example, if right and left index fingers both have the same ridge pattern (e.g., whorls on both fingers), then the patterns are concordant. If the ridge patterns differ (e.g., whorl on the right index and arch on the left index), then the patterns are discordant. The number of discordant fingers ranges from 0, if all fingers are concordant, to 5, if all fingers are discordant. Radial and ulnar loops can be treated as being either concordant or discordant. Most studies, however, have lumped radial and ulnar loops together as concordant.
In addition to ridge pattern discordance, Holt [88] introduced the finger-to-finger diversity of ridge counts—S2, the sum of squares of deviations of the ten separate finger ridge-counts from their mean: , where the qi are the ridge-counts on individual digits q1 through q10 and Q is the total ridge count (Σq). Later, Holt [89] settled on , where S is the standard deviation of q1 through q10; this index of diversity has a more symmetrical distribution. In contrast to Holt’s diversity of ridge counts, Micle and Kobyliansky [90] introduced a ridge count diversity index based on Shannon’s diversity index.
Because dermatoglyphic ridges are formed during a narrow window of time, one would not expect them to exhibit multiplicative error and positive size scaling. Holt’s [45] raw data show no evidence of size scaling; indeed, the slope of the regression was slightly negative (Figure 1). When one divides |R − L| by (R + L), strong negative size scaling is generated. Groups with smaller total ridge count will now appear to be more asymmetric. If this is a general pattern, then Karmaker et al.’s [91] conclusion that West Bengali women have greater fluctuating dermatoglyphic asymmetry than the men may be an artifact of inappropriate size scaling, because women usually have smaller ridge counts than men.
Statistical analyses of dermatoglyphic patterns are often seriously flawed. The most serious issue is that individual fingers are not statistically independent of one another. One cannot, for example, take a sample of 25 individuals and assume the sample size is now n = 250 fingers (with an order of magnitude greater degrees of freedom). The unit of observation is the individual. While we have not seen this in studies of fluctuating dermatoglyphic asymmetry, we have seen fluctuating asymmetry correlated with finger ridge patterns that were inflated in such a way. Consequently, claims that a greater frequency of, say, ulnar loops is associated with a particular physical anomaly, such as oral cleft, needs to be reexamined with more realistic degrees of freedom (see Mathew et al. [92] for just one example). Similarly, tallying the numbers of discordant fingers over all homologous pairs inflates the degrees of freedom five-fold (see Mellor [38]).
Many analyses of dermatoglyphic asymmetry have other problems, especially with the choice of asymmetry index. At the dawn of the modern interest in fluctuating asymmetry, two classic papers took different approaches. Holt [45], following the example of Bonnevie [93], used the correlation coefficient (r) as an index of quantitative value (i.e., fluctuating asymmetry) of finger ridge counts. Mather [3], on the other hand, elected to use the variance of L − R as an index of developmental instability [48]. Nine years later, Van Valen [4] used an offshoot of the correlation coefficient (r), 1 − r2, the coefficient of indetermination, as an estimate of the unshared variance (i.e., the random variation). Most early researchers working with humans, including Bailit et al. [2], Micle and Koblyliansky [90], Mellor [38], and others [94,95,96] followed Holt’s [45] and Van Valen’s [4] lead (but see [77,82,83,97,98]). Evolutionary biologists, on the other hand, favored some variant of Mather’s [3] approach and soon found justification for it. Angus [39] argued that the correlation coefficient (r) is overly influenced by the range of R and L. Even when Var(R − L) is large, the correlation of R and L will be small if Var(R + L) is small. Palmer and Strobeck [8], in an influential paper, concluded that the correlation of R on L behaves differently from all other indices and never achieves their statistical power. Nevertheless, both the correlation coefficient and coefficient of indetermination were still in wide use five years ago, especially by those studying human dermatoglyphic asymmetry (see [40,95,96,99]).
3.5. Brain Laterality
Human brains, and the brains of our close hominin relatives, exhibit several directional asymmetries. Broca’s area, within the inferior frontal gyrus, for example, is consistently larger on the left side and is associated with language [101]. There are also directional brain asymmetries in the planum temporale. While these asymmetries are not fluctuating asymmetries, extreme directional asymmetries associated with brain laterality have been correlated with fluctuating asymmetries in other traits, such as a composite index of ear length and width, atd angle, and the widths of wrist, hand, foot, and ankle [102,103], and finger length [104].
3.6. Asymmetry of Soft Tissues
Various soft tissues, such as breasts [105,106], ears [29], and feet [107] have been subject to studies of fluctuating asymmetry. With any soft tissues, those overlying bone may reflect asymmetry of the bone [108]. This is true of wrist and ankle diameters, as well as facial features. Unless the soft tissue itself is of interest, these traits may not be the best choice, though they are often the most convenient choice. It is difficult to measure soft tissue with precision, and measurement error is high, especially compared to right–left differences. Moreover, soft tissue asymmetry may be temporally variable. Breast asymmetry, for example, varies during the menstrual cycle in women [106]. Asymmetry is greater at the beginning and end of the cycle.
3.7. Fractal Anatomy and Physiology
Fractals have symmetry of scale. Examples of human fractal anatomy and physiology include the branching fractals of blood, lymph, and tracheal and bronchial vessels, time-series fractals of heart beat and electroencephalographic records of brain-wave activity, cranial suture joints, and small-world neural networks. Loss of fractal scaling under stress often manifests as a decrease in the fractal dimension, an index of the complexity of a fractal pattern. Various fractals have been studied in plants and animals [16,17,109,110,111], but nothing has been done on humans beyond anecdotal observations on human retinas and cranial sutures. D. C. Freeman (Wayne State University, pers. comm.) observed that the fractal dimension of the vasculature of human retinas is reduced among hyper-stressed, myopic, diabetic, and senile humans [17]. He also observed that the fractal dimension of the cranial sutures of Down syndrome individuals are reduced, so much so that the cranial bones often do not fuse. Beyond this, little work has been done on human fractal structures, from the perspective of developmental instability.
3.8. Behavioral Patterns
Temporal patterns of behavior have their own symmetry. Behavioral patterns occur sequentially, in time. They can be characterized as either a kind of translatory symmetry in time, or as a fractal dimension. Phase locking musicians (drummers) and dancers are examples. This is an area of research open for exploration.
4. Sex Differences
Fluctuating asymmetry may differ between males and females. Females, for example, may be more resilient against environmental and genetic stresses, because they carry two X-chromosomes. Deleterious recessive alleles on the X-chromosome are more likely to be buffered, at least in a fraction of the cells, in females than in males. Thus, females may develop in a more stable way. If true, most men should be expected to exhibit greater fluctuating asymmetry than women. Kuswandari and Nishino [112], for example, studied dental asymmetry of young boys and girls. Dentition of males was more asymmetric than those of females. There are, however, numerous contradictory findings in the literature on this issue [113,114,115,116,117,118,119,120,121]. Furthermore, random X-inactivation can generate mosaic patterns in females for heterozygous loci on the X-chromosome; this could conceivably increase asymmetry for some traits.
Among Jamaican children, girls are more asymmetric than boys for a composite index of asymmetry [120]. Moreover, Hadza women are more asymmetric than men [115]. In contrast, Wilson and Manning [121] found no significant differences between English boys and girls for 11 traits. Guatelli-Steinberg and colleagues [116] investigated 469 dental casts taken from persons of the Gullah population living on St. James Island, South Carolina, during the 1950s. They found that, female mandibular canines exhibited significantly greater fluctuating asymmetry than those of males. Nevertheless, there were no significant sex differences in the asymmetry of the maxillary canines. Moreover, they found that deciduous teeth were not significantly less asymmetric than their permanent successors. Other studies also found little or no difference in odontometric asymmetry in comparisons of males and females [2,122]. Finally, Albert and Greene [113] compared two skeleton groups from Kulubnarti, Sudan, dating back to the early and late Christian era; they found no significant differences in asymmetry between the sexes.
Does facial symmetry differ between males and females? The most comprehensive study of facial asymmetry in males and females was performed by Özener and Fink [123], who analyzed frontal photos of 503 high school students aged 17–18 years who were either living in slum areas or wealthy neighborhoods of Ankara, Turkey. Males and females showed no differences in a composite index of asymmetry in the group living in relatively good conditions, while men living in poor conditions had significantly greater asymmetry than females. Finally, Simmons et al. [119] found that male faces were significantly more asymmetric in a mixed-ethnic sample.
Early sex hormone levels may play a crucial role in the development of body morphology [124,125] and also facial asymmetries [126]. Testosterone released at high levels, for example, results in low immunocompetence. Given that men are less resilient against environmental stress because of their testosterone levels, a high concentration of testosterone is expected to increase fluctuating asymmetry. Fink and colleagues [126] found a significant correlation between prenatal testosterone level (low second-to-fourth digit ratio) and high facial asymmetry in males, but not females. These findings, however, are controversial [127].
As we have shown, there are numerous controversial findings in the literature regarding sex differences. Some of the variation among studies may arise because of (1) different methods; (2) poor statistical power; (3) use of different characters; and (4) unexplained variation. Even if all methodological conditions necessary for the analysis of fluctuating asymmetry are provided, it may be unreasonable to expect differences between sexes in a population not exposed to significant developmental stress. Sex differences in fluctuating asymmetry probably become evident as stress increases [128]. On the other hand, living conditions of females in some traditional patriarchal societies may be worse than those of men, which may explain increased asymmetry in women. Finally, the differences in dental asymmetry may be an artifact of inappropriate size scaling; females have smaller teeth on average than men.
5. Stressors
Stress dissipates energy away from growth and production [12]. Common stressors include inadequate diet, disease and parasitism, environmental toxins, extreme climate, social competition, emotional response to natural or man-made disasters, inbreeding depression, breakdown of genomic coadaptation, homozygosity, mutation, and chromosomal aberrations (polyploidy, aneuploidy, translocation, deletion, duplication, etc.). Perturbations, on the other hand, are energy or mass communicated from the environment. A perturbation can be as small as the kinetic energy of Brownian motion within a cell or can be as large as physical trauma experienced during fetal development. Stress and perturbation, acting together, can compromise developmental stability, possibly causing an increase in fluctuating asymmetry.
Stressors operate through the hypothalamic–pituitary–adrenal axis [127], which acts in part as a link between the nervous system and the immune system. Under normally stressful conditions, corticotropin releasing hormone stimulates the release of adrenocorticotropic hormone, which stimulates the adrenal gland to release cortisol (a corticoid hormone). Cortisol suppresses the immune system and aids the metabolism of lipids, proteins, and carbohydrates. It also provides a negative feedback to both the hypothalamus and the pituitary. Acute or chronic stress, however, elevates cortisol levels. Extended prenatal exposure to maternal cortisol early in development is associated with cognitive impairment [129]. Furthermore, children born during a natural disaster (chronic stress) have both elevated cortisol levels and greater dermatoglyphic asymmetry, supporting this hypothesis [79,80].
Stressors known, or suspected of, causing increased asymmetry fall into two categories: environmental and genetic.
5.1. Environmental Stressors
5.1.1. Dietary Inadequacy, Growth, and Socioeconomic Status
Socioeconomic living conditions shape the human body. People living in low socioeconomic conditions are known to have a worse diet and to experience more infectious diseases. Consequently, growth is known to differ between individuals living in lower and upper socio-economic conditions [130].
Kieser et al. [131] studied the Lengua Indians of the Gran Chaco region of Paraguay. Historically, the Lengua were subsistence hunters and gatherers prior to the advent of missionary settlements in the Chaco. There was a high rate of dental asymmetry in older members of the population, who had been exposed to widespread malnutrition and disease. More symmetric younger individuals, in contrast, had had access to western medicine.
Livshits et al. [132] show how socioeconomic status affects body symmetry. They studied physical asymmetry of infants born at different gestational ages. In general, preterm infants were the most asymmetric. They also studied the relationship between the mother’s education and health status, and the infant’s body asymmetry. There was a negative relationship between asymmetry and maternal health (cardiovascular only), but not between asymmetry and the mother’s education.
On the other hand, a few studies have examined socioeconomic groups having different growth and nutrition. For example, Little et al. [133] studied chronically malnourished children in southern Mexico. Well-nourished children were used as the control group. Contrary to expectations, body asymmetry was lower in the malnourished children. A similar observation was seen in villages on the east coast of the Dominican Republic. Flinn and colleagues [134] compared fluctuating asymmetry of adopted and biological children from several families. The biological children had better weight and length increases compared to the foster children in all age groups between 0 and 20 years. Fluctuating asymmetry of nine bilateral traits, however, was lower in the adopted children [134].
What is the relationship between physical growth potential and developmental stability? How can one explain the surprising results of Little and Flinn? Wells et al. [135] studied 172 boys, aged 9 years. The study included estimates of fetal growth rate, early infant growth rate, and total post-natal growth rate. No relationship was found between birth weight and height at 9 years. Faster post-natal growth rate, however—but not fetal growth rate—was associated with increased asymmetry at the end of 9 years.
Several comprehensive studies of nutrition, social status, and fluctuating asymmetry have been conducted in the city of Ankara, Turkey [118,123,128,136]. Özener [118] studied the effects of heavy labor and poor socioeconomic conditions on fluctuating asymmetry in three groups of young males: (1) those from lower socioeconomic classes employed in heavy industry; (2) those from the same socioeconomic classes who were not laborers; and (3) non-laborers from the higher socioeconomic classes. Ten bilateral measures were taken from 309 young adults. The two lower socioeconomic groups had significantly greater fluctuating asymmetry. In addition, biomechanical pressures associated with heavy labor appeared to increase the directional asymmetry of the upper extremities [137]. Moreover, the directional asymmetry increased with the number of years of heavy labor [118]. In another study, tall young men from Ankara living under good conditions were found to be more symmetric [118,136].
Poor conditions also contribute to facial asymmetry. Özener and Fink [123], as mentioned previously, compared high school students from schools in city slums with those from schools in higher socioeconomic areas. The group from the lower socioeconomic level had a higher composite index of fluctuating asymmetry. Consequently, poor living conditions influence developmental instability in humans, manifested in their facial asymmetry [123]. Findings from Turkey indicate that persons living in good conditions exhibit both greater facial and morphological symmetry.
5.1.2. Climate
Inhospitable climate has long been associated with increased asymmetry in human populations [2]. Nevertheless, it is difficult to disentangle the effects of climate, diet, disease, and inbreeding in most anthropological studies. Recently, Tomaszewska et al. [138] suggested that symmetry of the orbital opening may be related to climate, with fluctuating asymmetry transitioning to directional asymmetry as one travels from warmer to colder climates. Nevertheless, the degrees of freedom in this study are inflated by the pooling of 846 skulls from several populations across Europe into three categories—warm, temperate, and cold. The degrees of freedom are inflated by not including populations nested within climatic category as an effect.
5.1.3. Environmental Toxicology
There is a large literature on the effects of toxins, such as heavy metals, chlorinated hydrocarbons, pesticides, air pollution, and sewage on the developmental stability of plants [139,140,141], invertebrates [142,143], fishes [144,145], amphibians [146], reptiles [147], birds [148], and mammals [149]. These select references represent a small fraction of the literature on environmental toxicology and fluctuating asymmetry. See Beasley et al. [150] for a wide-ranging meta-analysis of anthropogenically-induced asymmetry in animals. Yet we have been unable to find any comparable human studies. Humans, of course, are exposed to the same environmental toxins [151]. Environmental toxicology and fluctuating asymmetry of human populations should be a research priority.
5.1.4. Drugs
Despite an absence of studies on environmental toxins and fluctuating asymmetry, there have been a handful of studies on maternal drug use. Maternal alcohol consumption, for example, is associated with increased dental [152], dermatoglyphic [153], and facial [154] asymmetry of children. In contrast, smoking of tobacco by both parents was not associated with increased dental asymmetry in a sample of 199 school children [155], but a combination of smoking and obesity was associated with increased dental asymmetry in a much larger sample of 440 school children [156]. This underscores the possibility that different stressors can interact with one another.
5.1.5. Natural Disasters
King et al. [79,80] discovered stress-related phenomena in children who were in utero during the Quebec Ice Storm of 1998. In addition to elevated dermatoglyphic asymmetry, children displayed elevated levels of cortisol, a marker of stress. Two additional studies of chronic stress and natural disasters are in progress [79]: the Iowa Flood Study and the Queensland Flood Study.
5.2. Genetic Stressors
Potential genetic stressors include chromosomal anomalies (aneuploidy, polyploidy, unbalanced translocations), point mutations, deleterious recessive alleles exposed by inbreeding, reduced heterozygosity, and reduced genomic coadaptation caused by outbreeding. Genetic subsidies include heterosis and overdominance associated with increased heterozygosity.
5.2.1. Aneuploidy
Aneuploidy (monosomy and trisomy) entails the loss or gain of individual chromosomes. Monosomies, such as Turner syndrome (45, X), involve the loss of a single chromosome. Trisomies, such as Down syndrome (47, +21), Edward’s syndrome (47, +18), Patau syndrome (47, +13), Triple X syndrome (47, XXX), Klinefelter syndrome (47, XXY), and XYY syndrome (47, XYY), entail the addition of a single chromosome. The chromosomal imbalance provoked by the loss or gain of a single chromosome upsets the gene dosage (i.e., the evolved balance of gene products), which then disturbs development [157,158,159,160].
One of the first genetic stressors to be associated with fluctuating asymmetry was Down syndrome, which is a trisomy involving the 21st chromosome. Down syndrome is accompanied by deceleration of growth and differentiation [161] as well as intellectual disability and modified craniofacial features [158,162,163,164]. Garn et al. [165], Barden [166], and Townsend [167] showed that dental asymmetry was greater in Down syndrome individuals than corresponding controls. However, two of these older studies have statistical problems. Barden [166], for example, used right–left correlation coefficients as the index of fluctuating asymmetry, so the significant results could easily have been caused by a small range of sizes in the Down syndrome sample but not in the control sample. Furthermore, Townsend [167] scaled the right–left asymmetry values by tooth size, without justifying the transformation. Given that others have found no relationship between tooth asymmetry and tooth size [65,66], the elevated asymmetry of the Down syndrome sample could have been easily magnified by the transformation; Down syndrome individuals have smaller teeth and scaling by tooth size would have increased the relative asymmetry of smaller teeth. Despite these potential problems, the conclusions of both Barden and Townsend are consistent with Garn et al.’s [165] earlier paper. His team used a root-mean squared index of asymmetry, based on R − L, and no corrections for tooth size. They found that the dental asymmetry of the Down syndrome individuals was nearly twice that of a control population. More recently, Starbuck et al. [168] reported 140%–160% greater facial asymmetry in Down syndrome individuals than in normally developing siblings and sibling pairs. Nevertheless, there have been well-designed studies that have found no increase in fluctuating asymmetry with Down syndrome. Palmar dermatoglyphic asymmetry, for example, was not significantly greater in a Down syndrome sample [169]. Furthermore, Bots et al. [170] were unable to demonstrate increased asymmetry of the long bones (ulna, radius, femur, tibia, fibula, digit 2, and digit 4) in a sample of deceased fetuses having Down syndrome.
There have been few studies of fluctuating asymmetry in other aneuploidies. Nevertheless, a pooled sample of deceased fetuses having trisomy 13 (n = 10), trisomy 18 (n = 24), Turner syndrome (n = 9), and triploidy (n = 10) had 1.5 times greater asymmetry than either controls or Down syndrome individuals [170]. All three of these other trisomies are more severe in the sense that they have greater spontaneous abortion rates during pregnancy than Down syndrome [171].
5.2.2. Polyploidy
Polyploidy involves the duplication of entire haploid sets of chromosomes. The normal adult complement of chromosomes is diploid (two complete haploid sets). Triploidy, a kind of polyploidy, involves three haploid sets of chromosomes, while tetraploidy involves four sets, and so on. For human embryos and fetuses, polyploidy is invariably fatal—100% of polyploid fetuses are spontaneously aborted [171]. Bots et al. [170], studying long bone asymmetry of deceased fetuses, included 10 triploid individuals in their group of highly asymmetric individuals. Mean asymmetry of these 10 individuals was similar to that for Trisomy 18 and Turner syndrome, and was greater than that of either controls or Down syndrome individuals.
5.2.3. Unbalanced Translocation
Translocations involve the rearrangement of parts of non-homologous chromosomes. Unbalanced translocations, where one or more chromosome segments are missing or duplicated, would be most likely to compromise developmental stability. Hereditary Down syndrome, {46, t(14;21)} or {45, t(21;21)(q10;q10)}, is one such translocation that duplicates the long arm of chromosome 21. To our knowledge, however, there have been no studies of fluctuating asymmetry associated with particular chromosomal translocations in humans.
5.2.4. Point Mutation
Although a few point mutations have been linked to increased asymmetry in Drosophila [172], Mus [173], and other species [174], there are no clear and unambiguous examples in humans that we are aware of. Single-gene knockouts or over-expression studies cannot be done with humans. Furthermore, the necessary search of quantitative trait loci (QTL) affecting asymmetry has not been done with humans, though there are studies of such QTLs in mouse models [173,175,176].
5.2.5. Inbreeding and Heterozygosity
Inbreeding exposes deleterious recessive alleles and reduces heterozygosity [177]. Moreover, overdominance (heterozygote superiority) at a few critical loci (e.g., the major histocompatibility complex) may contribute to inbreeding depression if those key loci are in the homozygous state. Several studies have established that the detrimental fitness effects of inbreeding are most evident when the environment is also stressful [178,179,180]. This implies a statistical interaction between genetic and environmental stressors. Under benign conditions, then, inbred individuals may perform nearly as well as outbred ones. Meagher et al. [179], for example, raised inbred and outbred wild-caught house mice (Mus musculus) in both large, semi-natural enclosures and in the laboratory. The reduction in fitness of male mice under semi-natural conditions was 81% (57% overall for males and females), whereas the reduction in litter size in the laboratory was only 11%. The main stress in the semi-natural enclosures was male–male competition.
The link between heterozygosity and mortality in humans is significant, but small; every standard deviation increase in heterozygosity above the mean is associated with a 1.57% decrease in mortality [181]. Nevertheless, both detrimental and beneficial impacts of human inbreeding have been described [182,183]. Consequently, it may be difficult to establish absolute rules for inbreeding and developmental instability.
Inbreeding has been linked to developmental instability in a number of plant [184] and animal [3,109,185,186,187,188,189] species, though contrary results are common [188,190,191,192,193,194]. As with mice raised under controlled conditions, inbred bird populations have greater fluctuating asymmetry when there is the additional stress of forest fragmentation [195]. Results from human studies have been mixed as well, perhaps because inbreeding is interacting with environmental stress in some populations, but not in others.
Several researchers, for example, have reported high fluctuating asymmetry in small, inbred, human populations [131,196,197,198,199], but few of these studies have controlled for other variables (environment, diet, disease, parasite load, etc.). At first, children of first-cousin marriages in Sivas, Turkey, appeared to fit this same pattern. Children of first cousins had greater asymmetry than those of unrelated parents, but differences in education, not inbreeding, accounted for the difference [200]. Level of inbreeding was confounded with level of education, which was in turn confounded with local smoking rates. Less educated parents were more likely to smoke cigarettes and engage in first-cousin marriages. Smoking, or other differences in life style, may explain the results better than inbreeding.
Inbred individuals are often from the lowest socioeconomic classes. This was the case with our study of inbreeding and fluctuating asymmetry in Sivas, Turkey [200]. Education is a surrogate measure of socioeconomic class. Consequently, the less well-educated may be exposed to additional environmental stresses, and a composite measure of fluctuating asymmetry correlated strongly with education, especially males and inbred females. In another study that incorporates both socioeconomic status and inbreeding, Schaefer et al. [201] compared dental arch asymmetries of an isolated population from Hvar, in the Eastern Adriatic, to a population from Zagreb, on the mainland of Croatia. Hvar is rural and the residents are mostly farmers and fishermen. Zagreb, in contrast, is a large city and the capital of Croatia. Its residents have the highest purchasing power and gross domestic product per capita in Croatia. Fluctuating asymmetry was greater in the Hvar population than in the Zagreb population, presumably because of greater environmental stress. Even an outbred subsample from Hvar had greater dental asymmetry than the outbred sample from Zagreb. Access to medical care was much less on the island, though other stressors may also be involved. Moreover, fluctuating asymmetry increased with level of inbreeding (outbred, low endogamy, high endogamy) on Hvar. Unfortunately, the Zagreb sample was not subdivided by level of inbreeding, so as to make for a clear comparison. Future studies of inbreeding and fluctuating asymmetry need to clearly control for socioeconomic status.
5.2.6. Outbreeding Depression, Heterosis, and Genomic Coadaptation
Outbreeding between genetically distinct populations can lead to (1) the breakup of genomic coadaptation [202] or (2) the restoration of higher levels of heterozygosity. If outbreeding has negative fitness consequences, it is referred to as outbreeding depression [203]. If it has positive fitness consequences (e.g., if both populations were highly inbred before hybridization), it is referred to as hybrid vigor or heterosis. In most cases, outbreeding depression occurs when populations are separated at the species or subspecies levels, having been isolated for thousands of generations, though several studies have observed outbreeding depression within a single species of copepods in neighboring tidal pools [204,205,206] and among breeding stocks of salmon [207,208,209]. Heterosis is expected when small, inbred populations meet and hybridize.
Virtually no research that we are aware of has been done on the fitness effects (positive or negative) of outbreeding between human populations. It is a missed opportunity. An obvious suggestion would be to extend the research of Lewis [210] and Little et al. [211], who reported that individuals of mixed-ethnicity are considered more attractive than those of relatively “pure” ethnicity. Is part of the attraction due to increased facial symmetry?
6. Phylogenetic Patterns of Fluctuating Asymmetry in the Hominidae and Other Primates
Fluctuating asymmetry has been studied in several primates other than Homo sapiens [212,213,214,215,216,217]. Much of the interest, though, has been with morphological integration of the primate skull [216] and limbs [212]. Nevertheless, Manning and Chamberlain [214] studied dental asymmetry of endangered gorillas (Gorilla gorilla), following reduction in their populations over several decades.
Population declines of gorillas are associated with increased dental asymmetry, presumably because of inbreeding in smaller populations. Fluctuating asymmetry of male canines, for example, increased between 1911 and 1986, a period when gorilla populations were declining from hunting and habitat loss [214]. The authors, however, used relative fluctuating asymmetry, despite an absence of positive size scaling. Indeed, according to the authors, the females even showed negative size scaling of |R − L| for their canine asymmetries. Consequently, it is difficult to say how the use of relative asymmetry has influenced this study, because the average size of gorillas (and their teeth) did not decrease during the period.
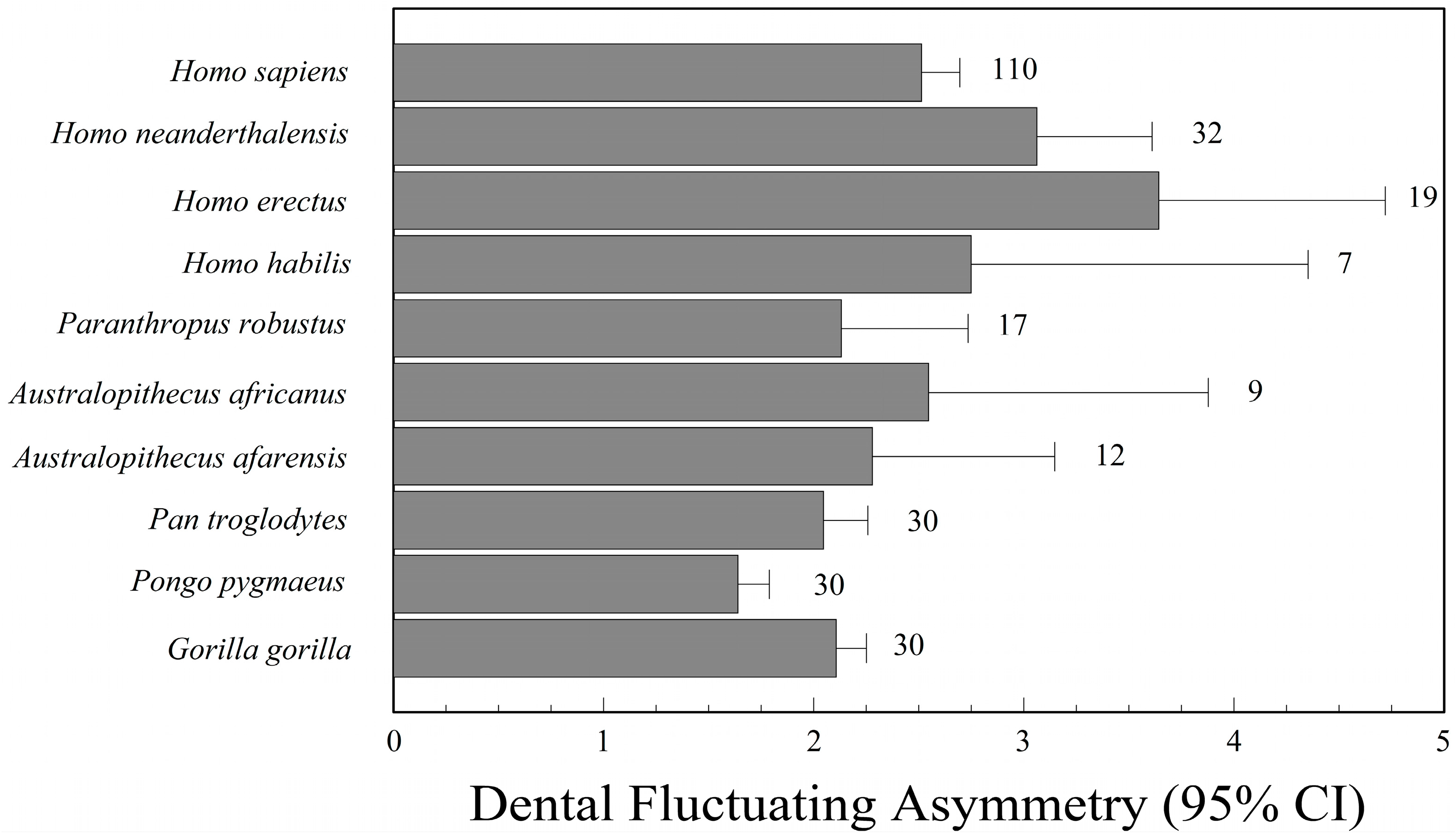
There has been keen interest in phylogenetic patterns of fluctuating asymmetry among living and extinct hominids, including extinct species in the genus Homo, as well as Australopithecus, Paranthropus, Pan, Gorilla, and Pongo. Extinct human species that have been studied include H. habilis [218,219], H. erectus [220,221], H. floresiensis [222,223], and H. neanderthalensis [220,221,224], though sample sizes are often too small to say much, especially for H. floresiensis (n = 1).
In broad phylogenetic comparisons, Kieser and Groeneveld [221] concluded that H. neanderthalensis had greater dental asymmetry than H. erectus and Australopithecus sp., while Frederick and Gallup [220] suggested there was broad overlap in dental asymmetry among H. habilis, H. erectus, H. neanderthalensis, and H. sapiens, but that all four species of Homo had greater asymmetry than Pan, Gorilla, and Pongo. All of these studies of dental asymmetry used relative indices of fluctuating asymmetry. Consequently, it is possible that the perceived increase in fluctuating asymmetry in the genus Homo is an artifact of a reduction in tooth size, especially the canines, compared to Pan, Gorilla, and Pongo. Finally, Babb and colleagues [222,225] concluded that the single specimen (LB1) of H. floresiensis from Liang Bua (Flores) was within the range of normal cranial asymmetry for the genus Homo. One cannot say more with a single individual.
7. Human Populations
7.1. Fossil Hominids
Although the literature is small, there is some information regarding fluctuating asymmetry of fossil hominids. Most of our information is based on the observation of teeth. Homo neanderthalensis is the species for which we have the greatest knowledge. Suarez [224], for example, studied 36 Neanderthal skeletons from various regions; these individuals had greater dental asymmetry (mesiodistal and buccolingual crown dimensions) than a population of H. sapiens from 20th century Ohio. According to Suarez, prevalence of inbreeding is probably the main cause of high fluctuating asymmetry in Neanderthals. Recent evidence of low genetic variation in Neanderthals is consistent with this hypothesis [226,227]. Doyle and Johnson [228], however, compared dental asymmetry of Pueblo and Eskimo populations, in which inbreeding was not widespread, to that of Neanderthals. They found no significant differences between the groups and concluded that environmental stress is responsible for the observed asymmetry in Neanderthals, rather than inbreeding. Kieser and Groeneveld [221] showed that H. erectus and H. neanderthalensis had greater dental asymmetry than Australopithecus. They also found that both H. erectus and H. neanderthalensis were developmentally unstable in mesiodistal dimensions of the maxillary teeth. Finally, H. neanderthalensis had significantly greater mesiodistal asymmetry than H. erectus. On the other hand, the methods used in fluctuating asymmetry studies before the 1990s were quite unsophisticated and based on correlation indices [2,63,221,229,230]. Barrett et al. [65], in contrast, used more current methods suggested by Palmer and Strobeck [8,231]. These methods include the analysis of measurement error, analysis of size scaling, the use of two-way mixed-model ANOVA, and the appraisal of directional asymmetry and antisymmetry. With these methods, they confirmed that Neanderthals had higher dental asymmetry than recent and prehistoric populations of H. sapiens. Unfortunately, they scaled right and left tooth sizes by their average, in essence correcting for positive size scaling where it was probably not present. It is unclear how using an index of relative fluctuating asymmetry has affected their conclusions.
Frederick and Gallup [220] studied 296 specimens, including several great apes and various species of Australopithecus, Paranthropus, and Homo. Once again, H. erectus and H. neanderthalensis had the greatest dental asymmetry (Figure 3). As with Barrett’s study, some of the variation in this data, however, may be an artifact of correcting for tooth size.
Fluctuating asymmetry of fossil hominids can also be conducted on postcranial bones. However, because these skeletal materials may be deformed during fossilization, they are not as useful as dental remains. Nevertheless, skeletal remains do reveal aspects of asymmetry. Homo erectus, KNM-WT 15000, sometimes known as Nariokotome Boy, from Nariokotome, Kenya, is a case in point. Greater development of the clavicular region on which the right deltoid muscle of the skeleton is attached, and a longer right ulna, suggest right handedness [232]. Until recently, this skeleton was thought to exhibit skeletal deformities and scoliosis—signs of developmental instability. Recent research, however, suggests that Nariokotome Boy was relatively normal [233].
Directional asymmetry, especially of the upper extremities, can indicate bone remodeling and reveal aspects of an individual’s lifestyle [232]. The hunter-gatherer lifestyle, for example, can increase asymmetry of the upper extremities; this is observed more markedly in Neanderthals [232,234]. In particular, abnormal thickening of the long bone’s diaphyseal size and narrowing of the joint space observed in this species suggests that the hunter-gatherer lifestyle has a very high biomechanical effect especially on the upper extremities.
This situation is repeated in human skeletons of the Northern European Mesolithic Age, presumably because of similar lifestyles. Constandse-Westermann and Newell [235], for example, found that diaphyseal sizes of the right upper extremities of individuals were quite large and the difference was higher among women. The Neolithic Age, which represents a transition from a hunter-gatherer lifestyle to agriculture, shows decreased upper extremity asymmetry in agricultural communities. Undoubtedly, the radical changes in lifestyle should be considered responsible for this decrease. Although agricultural production requires intense physical activity, equal levels of biomechanical stress caused by activity in the bilateral extremities is the main cause of the mentioned decrease in directional asymmetry. The conclusion to be drawn here is that increased asymmetry of the upper extremity does not always follow a parallel course to the increase in biomechanical stress. Although differences in asymmetry observed between the sexes in pre-agricultural societies has gradually decreased with the transition to agriculture, indicating that women and men have been involved in similar activities, this does not mean that biomechanical stresses on the two sexes have decreased [49,234].
7.2. Prehistoric and Historic Skeletal Samples
With the end of the Pleistocene glacial epoch, the establishment of agricultural societies, adopting a sedentary lifestyle on the fertile plains of the Middle East, is associated with numerous lifestyle diseases. Dental caries, dental hypoplasia, and growth retardation, as well as many infectious diseases have been the most important problems for agricultural societies in Neolithic communities [236].
Sciulli [66] compared dental asymmetry of pre-historic hunter-gatherers with those of settled agricultural societies in the Ohio Valley of North America. The hunter-gatherers were of the Late Archaic period (3200–2700 Before Present (BP)) and the agriculturists were of the Late Prehistoric period (950–300 BP). He found that directional asymmetry and fluctuating asymmetry were prominent in the Late Archaic and Late Prehistoric groups, but that there were no significant differences in dental asymmetry between these two societies. This finding may imply that farming of corn has not increased fluctuating asymmetry, and consequently not increased stress in the Late Prehistoric period. Similarly, Hoover and Matsumura [237] could not find significant differences in asymmetry among 13 different prehistoric communities of Japan’s Jamon Period, despite their different lifestyles. They used three bilateral traits from the nasal and orbital regions of the cranium and also collected hypoplasia data on the maxillary and mandibular anterior dentitions. Unfortunately, the study examined only 49 individuals collected from 13 different archaeological sites. This is an insufficient sample size to detect differences if they exist [237].
Albert and Greene [113] compared a temporal series of skeletal groups from Kulubnarti, Sudan, and dated from the early to late Christian periods. The authors reported more intense environmental stress during the early period, and demonstrated that the long-bone asymmetry was also greater in this group. That study, however, revealed no significant differences between males and females.
In addition to studies of the post-cranial skeleton, studies have also successfully used the skull, with its complex structure [114,238]. DeLeon [238] examined 60 skulls belonging to two Christian medieval communities, Christian Kulubnarti (Sudanese, Nubia), one early and one late-period. She used three-dimensional coordinates from landmarks on the skull to compare cranial asymmetry of the two samples. The crania from the Early Christian cemetery were more asymmetric than those from the Late Christian cemetery, which was consistent with Albert and Greene [113]. In a similar study, Bigoni and colleagues [114] studied 129 skulls from a high socioeconomic class at Mikulčice, a medieval settlement in the Czech Republic. Surprisingly, the females from this group had higher fluctuating asymmetry than those from a lower socioeconomic group in a nearby cemetery. The asymmetry of these females was similar to that of a modern low socioeconomic collection.
We have previously mentioned that bilateral asymmetries observed in skeletal groups are also caused by biomechanical stress. Consequently, fluctuating asymmetry may reveal the level of environmental and genetic stress, while directional asymmetry may indicate mechanical stress experienced by the populations. Biomechanical stress mostly causes increases in bone length, cortical thickness, and bone weight. Moreover, it manifests itself as enlargement of the joint surface in parallel with increased cartilaginous tissue. The effect of biomechanical stress on the joint surface is considered to have lower phenotypic plasticity compared with other regions of the bone [239,240]. According to some researchers, these parts of the bone are less influenced by mechanical stress, which demonstrates that development of these areas are rather controlled by genetic factors. In other words, these areas have lower phenotypic plasticity [239]. Some specialists maintain that intense mechanical stress in the joints causes deterioration of the surface of the joint rather than increasing the amount of tissue in the joint, leading to arthritis and other joint diseases [241,242]. Even small mechanical stresses caused by handedness may lead to deviation from symmetry in the joint surface. In a study of 80 skeletons from archaeological excavations in Missouri, Plochocki [243] studied joint spaces of the humerus, radius, femur, and tibia. In that study, measures of the joint surfaces in the upper extremities showed higher values on the right side. The measure that has shown the highest directional asymmetry in the upper extremities is the height of the humeral head. In a similar study by Čuk et al. [244] on a collection of 42 skeletons, the humerus was especially seen to have a high degree of asymmetry.
In addition to long bones, asymmetry studies have also been performed on clavicles and sacra [245,246]. In one of these studies, Mays et al. [245] examined the right and left clavicles of 136 skeletons from a small medieval town in Wharram Percy, England. Five measurements were taken on each clavicle; the left clavicles were longer [245]. The researchers proposed that growth was inhibited on the dominant right side by lateral differences in mechanical loading. Increased compression of the right shoulder girdle hinders longitudinal growth on that side. In a similar study, Plochocki [246] took three bilateral measures on each sacrum and found an inverse correlation between the dominant hand and these measures. In other words, the left side of the sacrum was larger in individuals who were strongly right handed [246].
8. Evolutionary Biology
A great deal of recent interest in evolutionary biology has focused on developmental stability as a component of Darwinian fitness. This naturally has led to fluctuating asymmetry as a surrogate measure of fitness. Nevertheless, because fitness is a characteristic of individuals, not populations, we are not really talking about fluctuating asymmetry any longer, but individual asymmetry.
Even if developmental instability is related to fitness, it is a weak association, because fluctuating asymmetry is a weak measure of individual developmental instability [247,248]. A significant association between fluctuating asymmetry and fitness components has been demonstrated in plants [249], but plants have the advantage of a modular design [250]. One can potentially sample hundreds of leaves from a single individual, and one can even sample multiple ramets from a single clonal colony, or genet. With animals, there are only two sides of a bilaterally symmetrical trait. One partial solution is to study several traits and combine them in a composite index of fluctuating asymmetry [251]. Composite indices of fluctuating asymmetry work best when the traits are developmentally independent of one another, but are nevertheless responsive to perturbations.
With this in mind, it is no surprise that studies of mate selection, a component of fitness, and developmental instability, as measured by fluctuating asymmetry, have proved so controversial. The hypothesis that symmetrical mates might have an advantage, because of “good genes” or an absence of disease or parasites, was a good one for its time [252]. An early meta-analysis by Møller and Thornhill [253], for example, on 146 data sets gleaned from 65 studies on a wide assortment of animal species claimed a moderately significant effect of individual symmetry on mate selection. However, a re-analysis of their data by Palmer [254] suggested that variation in asymmetry actually accounted for less than 6% of the variation in attractiveness (but see Thornhill et al.’s reply [255]). Moreover, the associations between facial symmetry and attractiveness ratings were often contradictory. Van Dongen [256] suggested that there was a relationship, but that it was weaker than had been portrayed and that the associations were stronger when the asymmetry was visible (i.e., facial traits) than when it was not (i.e., wrist, ankle, etc.). Van Dongen and Gangestad [257] and Van Dongen [256] conducted meta-analyses of published papers and concluded that the literature did not support an association between facial symmetry and attractiveness. Moreover, a second meta-analysis by Van Dongen [258] failed to find an association between developmental instability, as measured by fluctuating asymmetry, and perceived masculinity or femininity (but see [259]). Finally, an additional study by Van Dongen [260] also failed to find an association between attractiveness and facial symmetry.
Other aspects of attractiveness, such as voice quality [261,262,263,264] and skin condition [265] have been linked to fluctuating asymmetry. A meta-analysis of all five studies of voice quality and facial asymmetry in men and women supports a robust negative association between the two; individuals with more attractive voices have more symmetrical faces. The effect sizes, however, for an association between fluctuating asymmetry and voice quality are not large (r = −0.23 for men and r = 0.29 for women) [264]. Moreover, the association between skin condition (color homogeneity, texture, etc.) and fluctuating asymmetry is also small. According to Jones et al. [265], skin condition has a strong effect on male facial attractiveness (rs = 0.70), but the correlation between facial symmetry and skin condition is much smaller (rs = 0.23). Consequently, the association between facial attractiveness and symmetry may be confounded by skin condition, which is correlated with both variables.
Fluctuating asymmetry may not actually be the best indicator of reproductive fitness, simply because the modal state is still perfect symmetry, even for individuals of low developmental stability. Symmetry transitions from fluctuating asymmetry to directional asymmetry make more sense as an honest indicator of reproductive fitness and mate suitability. If individuals having low developmental stability also exhibit directional asymmetry for a sexually selected trait, then asymmetric individuals are more likely to be poor mate choices.
Fitness is always relative to other individuals in a population. Nevertheless, one may still be able to follow trends and associations and it may still be possible to estimate a population’s mean fitness.
9. Medicine
Fluctuating asymmetry has been especially problematic as an indicator of health and disease. Studies of overall health and disease have not been well supported. Van Dongen and Gangestad [257], for example, conducted a meta-analysis of nearly 100 studies addressing health and fluctuating asymmetry. They looked at studies focusing on infectious diseases, genetic defects, fetal anomalies, psychological disorders, attractiveness, and reproductive outcomes, finding only a weak effect size (Pearson r = 0.2) and evidence of publication bias. When publication bias was taken into account, the effect size decreased to r = 0.1. They concluded that “more work is needed to provide a full understanding” (Van Dongen and Gangestad [257], (p. 397)).
The most strongly supported studies have focused on genetic defects, such as cleft lip/cleft palate [266]. Adams and Niswander [1], for example, first described elevated dermatoglyphic and dental asymmetry in individuals with cleft lip/cleft palate. This is a classic, but equivocal paper, because the methods are so poorly described. In a more recent large study, Neiswanger et al. [267] did not find significant differences in ridge counts and atd angle asymmetries among 500 individuals having cleft-lip/palate, 421 of their unaffected relatives, and 66 unrelated controls. Only in families with a history of cleft-lip/palate was asymmetry (pattern discordance) greater than that of the controls. Recently, Miller et al. [70] identified three genes associated with facial asymmetry (actually directional asymmetry) in the relatives of nonsyndromic cleft lip with or without cleft palate; these genes are LEFTY1, LEFTY2, and SNAI1.
Unsurprisingly, individuals having facial paralysis have greater facial asymmetry than those without it [268]. This is one of the few noncontroversial studies.
There has been much research on limb asymmetry and the performance of athletes (both hominid and murid). In a long-term laboratory experiment involving mice, Garland and Freeman [269] selected for running endurance. The high-runner lines had reduced directional asymmetry of their hind limb bones. In general, the lower limbs of humans have lower asymmetry than the upper limbs, presumably for mechanical efficiency [120,270]. Nevertheless, directional asymmetry in the upper limbs may be amplified by behavioral laterality and bone remodeling.
Trivers et al. [270] found that knee asymmetry in Jamaican children influences sprint speed later in life. Moreover, elite athletes have more symmetrical knees, and possibly ankles, than a matched control group [271]. Moreover, Oxford [272] found that lower limb asymmetry (mostly long bones) does not influence performance of athletes. He did not examine knee asymmetry.
10. Psychology
Because psychological disorders, such as schizophrenia, psychosis, and autism spectrum disorder have their genesis during prenatal development, dermatoglyphic patterns and asymmetry have become the trait of choice for detecting developmental disturbances that might influence the nervous system [275]. Dermatoglyphic patterns form during weeks 10–16 of fetal development, just as the population of neurons in the brain is beginning to expand and differentiate [276].
Schizophrenia is a mental disorder with complex, polygenic roots, which it shares with bipolar disorder, intellectual disability, major depressive disorder, and autism spectrum disorders [277]. Several studies have linked fluctuating asymmetry with schizophrenia. Markow and Wandler [87], for example, have argued that the increased dermatoglyphic asymmetry (both fluctuating asymmetry and pattern discordance) observed in schizophrenic patients is evidence of schizophrenia’s polygenic origins. Mellor [38] and Reilly et al. [278] both reached similar conclusions, but their choice of indices is problematic. Mellor [38] had used the coefficient of indetermination, which—in addition to having low statistical power—could bias the asymmetry estimates in either direction, depending on the range of ridge counts in the samples. Furthermore, for pattern discordance, he counted each finger pair as an independent observation, thus inflating the degrees of freedom fivefold and increasing the chance of a type I error. Finally, Reilly et al. [278] corrected for size scaling when it was unnecessary (and without justification). The schizophrenia patients had smaller a–b and finger ridge counts, so it is likely that any difference between patients and controls was inflated by using an index of relative asymmetry.
Psychosis is accompanied by hallucinations, delusions, and thought disorders. It shares phenotypic and genetic features with schizophrenia, so it is only natural that it may be associated with developmental disorders during the prenatal period. Mittal et al. [275] reported significant elevation in fingerprint ridge count asymmetry in a small sample of students (n = 16) who had reported occasional non-clinical psychoses. The control group was much larger (n = 205). The effect size, however, was moderate (Cohen’s d = 0.44), by Cohen’s [279] criteria. More recently, the same research team [97,98] has shown that a larger sample of adolescents at risk for psychosis also have greater dermatoglyphic asymmetry, with a similar effect size (Cohen’s d = 0.48). The authors suggest that the results of both studies support the Diathesis-Stress Model in which prenatal insults disturb the structure and function of the hippocampal region, which regulates the hypothalamic–pituitary–adrenal axis. Then, psychosocial stress acting on the poorly regulated hypothalamic–pituitary–adrenal axis, increases cortisol secretions, even at rest, triggering further damage and psychoses.
Autism spectrum disorder is a collection of neurological disorders, including autistic disorder, Asperger syndrome, and Pervasive Developmental Disorder Not Otherwise Specified. Like schizophrenia, the autism spectrum has a complex genetic basis [280,281] and one might expect developmental disturbances similar to those that accompany schizophrenia. To the best of our knowledge, there have been no complete analyses of fluctuating asymmetry in autistic children, but pattern discordance is present in individuals having mild autism and average IQ [86]. There were no significant differences in atd angle asymmetry, and ridge count asymmetry was not examined.
Some researchers have claimed that intelligence is influenced by developmental disturbances [282]. Consequently, fluctuating asymmetry should be correlated with intelligence. Furlow et al. [283], for example, conducted two replicate studies of college students and found significant negative correlations between intelligence (culture neutral CFIT) and a composite index of fluctuating asymmetry. The traits examined were linear measurements of feet, ankles, fingers, wrists, elbows, and ears. The correlation coefficients were a modest r = −0.21 and r = −0.24. Prokosh et al. [284] found an even stronger negative relationship (r = −0.39) between the same traits and a general factor of mental ability (g), as measured by the g-loaded cognitive tests. Other studies from different research groups, however, were unable to confirm a negative correlation between asymmetry of these same traits and general intelligence, even with much larger sample sizes [54]. A meta-analysis [285] of 14 samples, both published and unpublished, suggests a significant, but lower, correlation between r = −0.12 and r = −0.20. The authors were concerned about evidence of possible publication bias (unpublished data sets had smaller correlations than published ones). Recent research suggests that dermatoglyphic asymmetries are related to hippocampal dependent cognitive function [97].
Handedness is related to brain laterality. Yeo et al. [103,286] have argued that handedness is critically influenced, not by a few gene loci, but by developmental instability. They called this the Developmental Instability Model of Handedness. Under this model, deviations from right-handedness are a consequence of disturbance early in development. Recent genomic studies [287] have failed to find a major gene for left handedness, providing some additional support for their model.
A recent trend has emerged in evolutionary psychology, in which researchers obtain a new tool, or measure, and apply it without regard for theory. Without theory to guide predictions, any finding based upon the new measure becomes interesting only to those who use it. Research on fluctuating asymmetry and behavioral patterns, for example, has occupied several researchers despite many failures to replicate findings [288,289,290,291]. The relationship between fluctuating asymmetry and aggressiveness is a case in point.
Early studies of aggressive behavior and fluctuating asymmetry revealed a negative relationship between body asymmetry and aggressive behavior. According to Manning and Wood [292], physically aggressive boys, but not girls, are more symmetrical, on average. Testosterone and/or cortisol effects on growth and behavior may explain the correlation.
Soon after Manning and Wood’s [292] paper, Furlow et al. [289] argued that asymmetry of ten body traits (e.g., ankle width, ear length, etc.) was negatively correlated with the number of fights among male university students, but not females, in the three previous years. Unlike Manning and Wood [292], who only examined asymmetry of one trait, the 10 traits they used were combined in a composite index of fluctuating asymmetry, but without standardizing each trait in the composite by its mean fluctuating asymmetry. Doing so emphasizes the most asymmetric of the 10 traits [251]. Fluctuating asymmetry was also associated with the initiation of fights by males. Consequently, human aggression is probably not a compensatory response by subdominant males, but is associated instead with alpha males maintaining their dominance.
Benderlioglu et al. [288] then attempted to replicate the findings of Manning and Wood [292] and Furlow et al. [289]. They studied 11 bilateral traits in 100 (average age = 20.1; 51 male, 49 female) college students and also combined their asymmetries (the average of two replicates) in an unstandardized composite index. In contrast to the two previous studies, however, they concluded that asymmetrical individuals were more aggressive. They also suggested that common prenatal insults, such as maternal alcohol and tobacco abuse, may influence both agonistic behavior and neurological functioning, providing further insight into the association between asymmetry and aggression.
Özener et al. [290] studied asymmetry (four bilateral traits) and aggression in a large non-western sample (100 male, 102 female) in Sivas (Central Anatolian Region), Turkey. They used the Buss and Perry Aggressiveness Questionnaire to establish self-reported aggressiveness, and found no relationship between a standardized index of composite asymmetry and any component of aggressiveness for either sex.
Further studies continued to confuse the issue. Some examined facial symmetry and aggressiveness. According to Muñoz-Reyes et al. [293], facial asymmetry is negatively associated with anger in older adolescent males, while it is negatively associated with hostility in females. Moreover, Holtzman et al. [294] argued that aggression and neuroticism are positively related to both body and facial symmetry. Özener et al. [291] concluded that facial symmetry is an unreliable predictor of self-reported aggressiveness of healthy young people in Ankara, Turkey.
As seen from these seven studies, there are three conflicting results. Manning and Wood [292], Furlow et al. [289], Holtzman et al. [294], and Muñoz-Reyes et al. [293] concluded that asymmetrical body and face is associated with lower aggressiveness. They hypothesize that aggressiveness reflects Darwinian fitness and is consequently related to lower fluctuating asymmetry. In contrast, Benderlioglu et al. [288] argued that asymmetrical individuals are more aggressive. The third group of findings are from studies of Turkish students. Özener and his colleagues [290,291] have not found any relationship between aggressive behavior and fluctuating asymmetry.
As Özener et al. [290] have stated, differences among studies may arise as a result of differences in (1) the traits studied; (2) the statistical power of the tests, which is a function of sample size and the effect size to be detected; and (3) the methods of measuring aggressiveness. Fluctuating asymmetry can be measured from a variety of traits (dermatoglyphic, skeletal, dental, morphological, etc.). The choice of traits and analysis influences the reliability of the predicted relationships. Moreover, there are a variety of methods of measuring aggressiveness. Finally, most studies were conducted on western populations. Only two studies [290,291] have been done in a non-western society. The negative association between aggressiveness and facial and bodily symmetry may not be universal. Therefore, aggression and asymmetry should be examined in several cultures.
11. Discussion
Fluctuating asymmetry of human populations has been linked to a wide variety of environmental and genetic stressors. Nevertheless, much of the research, especially the older research, does not measure up to current best practices, as outlined by Palmer and colleagues [8,22,23,36,231], Leung et al. [251], Van Dongen et al. [55], and Graham et al. [12]. The use of correlation coefficients, for example, and the closely related coefficient of indetermination, is widespread in the older literature. Both indices are highly susceptible to the range of R and L [39] and are far less sensitive than alternative indices [8]. Moreover, differences in correlation coefficients between two or more populations can easily be the spurious result of restriction in the range of R and L for one of the populations. Consequently, the probability of a Type I error (i.e., false positive result when the null hypothesis is true) may be much greater than 0.05 when the correlation coefficient is the index of asymmetry.
Because fluctuating asymmetry is such a small component of total phenotypic variation, it is important to have large sample sizes and to estimate, and account for, measurement error. Measurement error inflates estimates of fluctuating asymmetry. Consequently, replicate measurements are required to estimate measurement error. One can estimate measurement error from a subsample, but it is better to do replicate measurements for the entire sample. Besides improving the accuracy of one’s measurements, the best way to eliminate measurement error is to average replicate measurements. Each round of averaging reduces measurement error by 50%. Using the averages will become important when dealing with size scaling. Unfortunately, measurement error is only rarely reported in the human fluctuating asymmetry literature.
Size scaling needs to be addressed in every study of fluctuating asymmetry, both before and after any transformations to correct it. Positive size scaling occurs when asymmetry |R − L| increases with size (R + L). This needs to be examined within a particular population of interest, not in a pooled sample, and the range of variation needs to be great enough to detect size scaling if it really exists. Moreover, it is important to graphically examine the relationship between |R − L| and (R + L). Blindly correcting for positive size scaling can quickly get one into trouble, as we demonstrated in Figure 1, where an unnecessary transformation of fingerprint ridge counts caused asymmetry to decrease with the total ridge count. As with measurement error, very few studies of human fluctuating asymmetry report whether size scaling exists or not.
To understand why a transformation for positive size scaling might fail, fluctuating asymmetry researchers need an appreciation for multiplicative and additive error. A mixture distribution of the two can easily undermine an analysis of fluctuating asymmetry. Additive error, for example, is characteristic of measurement error and the natural variation of inert tissues, such as teeth, fingernails, and dermatoglyphic ridge counts. Multiplicative error, on the other hand, is characteristic of actively growing tissues, such as bone and soft tissues. Additive measurement error and multiplicative growth interact to generate a mixture distribution that will respond poorly to size scaling. If |log R − log L| or |R − L|/(R + L) decrease with size (R + L), it is likely that additive measurement error is a significant part of the asymmetry variation. The solution to this problem may be as simple as averaging replicate measurements until measurement error is nil.
To reduce size scaling, we prefer |log R − log L| to |R − L|/(R + L) or |R − L|/((R + L)/2). It addresses multiplicative error directly, and |log R − log L| has better statistical properties [43,57]. If a mixture distribution is encountered and replicate measurements have not been done, one can use a Box–Cox power transformation, [(Rλ − 1)/λ] − [(Lλ − 1)/λ], as a last resort to reduce size scaling [12].
Directional asymmetry needs to be addressed as well, mainly because it can inflate estimates of fluctuating asymmetry. Most of the human skeleton exhibits directional asymmetry. As with size scaling, directional asymmetry needs to be addressed graphically. For example, does directional asymmetry increase allometrically with size? Prescriptions for dealing with directional asymmetry can be found in Graham et al. [48] and Van Dongen et al. [55].
In addition to those concerns that are unique to fluctuating asymmetry, statisticians have expressed concern regarding statistical abuses in all of the sciences. Studies of fluctuating asymmetry are especially vulnerable to these abuses, which include p-hacking ([295], but see [296]), overuse of one-tailed tests [297,298], confirmation bias [299], lack of effect-size estimates and confidence intervals [295], poor experimental design, small sample sizes, low statistical power, and lack of replication.
p-hacking, the testing of statistical hypotheses until a significant effect is found, is especially problematic, because fluctuating asymmetry is, almost by definition, characterized by “small effects hidden in noisy data” [295]. If a researcher examines enough traits, at least 5% of them will be statistically significant at p < 0.05, even when the null hypothesis is true. For example, if one examines antimeric asymmetry in a full set of permanent teeth (32 total), there are 16 different variables and 16 different tests to address, unless one takes a multivariate approach or combines the 16 different variables into a composite index of fluctuating asymmetry [251].
True experiments often cannot be conducted on humans, and so anthropologists, medical researchers, and psychologists are often forced to adopt a comparative approach. Virtually all studies of human fluctuating asymmetry adopt the comparative approach and spurious results are always possible if researchers have overlooked an explanatory variable. In our study of the children of first-cousin marriages, for example, we could have wrongly concluded that inbreeding had had a significant effect on the children’s fluctuating asymmetry if we had not also included level of education as an explanatory variable. Of course, it is unlikely that education itself influences the developmental stability of one’s children, but other factors (smoking, diet, etc.) correlated with level of education may influence children in utero.
Furthermore, with respect to many of the claims that have been made for human fluctuating asymmetry, such as associations between asymmetry and general intelligence, we agree with Carl Sagan’s dictum “extraordinary claims require extraordinary proofs”.
12. Conclusions
Studies of human developmental instability and fluctuating asymmetry are extremely popular. Many studies, such as the association between schizophrenia and developmental instability, have produced robustly repeatable results. Others, such as the attempts to establish an association between developmental stability and general intelligence, have been less successful and consequently more contentious.
As we have pointed out repeatedly in this review, studies of human fluctuating asymmetry have not always followed best practices. Consequently, it has been difficult, perhaps impossible, to compare results from different studies. While it is natural that traits and measurements should vary, it is the lack of reliability introduced by less-than-ideal indices and statistical approaches that undermines most research.
We encourage researchers to take more care with how fluctuating asymmetry is treated in human studies. At the moment, many studies (even classic ones) are of questionable value, because they used insensitive indices, or used unnecessary transformations, or used the wrong transformations, or did not deal with directional asymmetry or measurement error. We recommend that every study of fluctuating asymmetry take the following precautions: (1) employ a large enough sample; (2) use a sensitive index of asymmetry; (3) take replicate measurements; (4) estimate measurement error as a fraction of total variation and asymmetry variation; (5) graphically test for size scaling, (6) use an appropriate transformation for size scaling if it exists; (7) estimate directional asymmetry; (8) evaluate whether measurement error is contributing to a mixture distribution of additive and multiplicative error; (9) measure asymmetry of several traits and either combine them in a composite index of fluctuating asymmetry or use a multivariate approach; and (10) include all correlated factors that could influence developmental instability in the design of the study.
Acknowledgments
We are grateful to Cathy Chamberlin-Graham, who helped with the references. J.H. Graham is supported by Berry College.
Author Contributions
John H. Graham and Bariş Özener wrote the paper.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| ANOVA | Analysis of Variance |
| BP | Before Present |
| C | Canine Tooth |
| CFA | Composite Index of Fluctuating Asymmetry |
| CFIT | Culture Fair Intelligence Test |
| E | Expectation |
| M1 | First Molar |
| LB1 | Liang Bua 1 specimen of Homo floresiensis |
| QTL | Quantitative Trait Locus |
| M2 | Second Molar |
| KNM-WT 15000 | Specimen Number of Nariokotome Boy |
| P3 | Third Premolar |
| Var | Variance |
References
- Adams, M.S.; Niswander, J.D. Developmental ‘noise’ and a congenital malformation. Genet. Res. 1967, 10, 313–317. [Google Scholar] [PubMed]
- Bailit, H.L.; Workman, P.L.; Niswander, J.D.; MacLean, C.J. Dental asymmetry as an indicator of genetic and environmental conditions in human populations. Hum. Biol. 1970, 42, 626–638. [Google Scholar] [PubMed]
- Mather, K. Genetical control of stability in development. Heredity 1953, 7, 297–336. [Google Scholar] [CrossRef]
- Van Valen, L. A study of fluctuating asymmetry. Evolution 1962, 16, 125–142. [Google Scholar] [CrossRef]
- Astauroff, B.L. Analyse der erblichen Störungsfälle der bilateralen Symmetrie. Zeitschrift für induktive Abstammungs Vererbungslehre 1930, 55, 183–262. [Google Scholar] [CrossRef]
- Ludwig, W. Das Rechts-Links Problem im Tierreich und heim Menschen; Springer-Verlag: Berlin, Germany, 1932; p. 496. [Google Scholar]
- Waddington, C.H. The Strategy of the Genes. A Discussion of Some Aspects of Theoretical Biology. With an Appendix by H. Kacser; George Allen Unwin: London, UK, 1957; p. ix-262. [Google Scholar]
- Palmer, A.R.; Strobeck, C. Fluctuating asymmetry: Measurement, analysis, patterns. Annu. Rev. Ecol. Syst. 1986, 17, 391–421. [Google Scholar] [CrossRef]
- Zakharov, V.M. Population phenogenetics: Analysis of developmental stability in natural populations. Acta Zool. Fenn. 1992, 191, 7–30. [Google Scholar]
- Van Dongen, S. Fluctuating asymmetry and developmental instability in evolutionary biology: Past, present and future. J. Evol. Biol. 2006, 19, 1727–1743. [Google Scholar] [CrossRef] [PubMed]
- Polak, M. Developmental Instability: Causes and Consequences; Oxford University Press: Oxford, UK, 2003; p. 488. [Google Scholar]
- Graham, J.H.; Raz, S.; Hel-Or, H.; Nevo, E. Fluctuating asymmetry: Methods, theory, and applications. Symmetry 2010, 2, 466–540. [Google Scholar] [CrossRef]
- Parsons, P.A. Fluctuating asymmetry: An epigenetic measure of stress. Biol. Rev. 1990, 65, 131–145. [Google Scholar] [CrossRef] [PubMed]
- Parsons, P.A. Fluctuating asymmetry: A biological monitor of environmental and genomic stress. Heredity 1992, 68, 361–364. [Google Scholar] [CrossRef] [PubMed]
- Graham, J.H.; Emlen, J.M.; Freeman, D.C. Nonlinear dynamics and developmental instability. In Developmental Instability: Causes and Consequences; Polak, M., Ed.; Oxford University Press: New York, NY, USA, 2003; pp. 35–49. [Google Scholar]
- Graham, J.H.; Freeman, D.C.; Emlen, J.M. Developmental stability: A sensitive indicator of populations under stress. In Environmental Toxicology and Risk Assessment; Landis, W.G., Hughes, J.S., Lewis, M.A., Eds.; ASTM International: Philadelphia, PA, USA, 1993; Volume STP1179, pp. 136–158. [Google Scholar]
- Emlen, J.M.; Freeman, D.C.; Graham, J.H. Nonlinear growth dynamics and the origin of fluctuating asymmetry. Genetica 1993, 89, 77–96. [Google Scholar] [CrossRef]
- Klingenberg, C.P.; Nijhout, H.F. Genetics of fluctuating asymmetry: A developmental model of developmental instability. Evolution 1999, 53, 358–375. [Google Scholar] [CrossRef]
- Graham, J.H.; Robb, D.T.; Poe, A.R. Random phenotypic variation of yeast (Saccharomyces cerevisiae) single-gene knockouts fits a double pareto-lognormal distribution. PLoS ONE 2012, 7, e48964. [Google Scholar] [CrossRef] [PubMed]
- Masel, J.; Siegal, M.L. Robustness: Mechanisms and consequences. Trends Genet. 2009, 25, 395–403. [Google Scholar] [CrossRef] [PubMed]
- Siegal, M.L.; Leu, J.-Y. On the nature and evolutionary impact of phenotypic robustness mechanisms. Annu. Rev. Ecol. Evol. Syst. 2014, 45, 496–517. [Google Scholar] [CrossRef] [PubMed]
- Palmer, A.R.; Strobeck, C. Fluctuating asymmetry as a measure of developmental stability: Implications of non-normal distributions and power of statistical tests. Acta Zool. Fenn. 1992, 191, 57–72. [Google Scholar]
- Palmer, A.R.; Strobeck, C. Fluctuating asymmetry analyses revisited. In Developmental Instability: Causes and Consequences; Polak, M., Ed.; Oxford University Press: Oxford, UK, 2003; pp. 279–319. [Google Scholar]
- Mardia, K.V.; Bookstein, F.L.; Moreton, I.J. Statistical assessment of bilateral symmetry of shapes. Biometrika 2000, 87, 285–300. [Google Scholar] [CrossRef]
- Klingenberg, C.P.; Barluenga, M.; Meyer, A. Shape analysis of symmetric structures: Quantifying variation among individuals and asymmetry. Evolution 2002, 56, 1909–1920. [Google Scholar] [CrossRef] [PubMed]
- Klingenberg, C.P. Analyzing fluctuating asymmetry with geometric morphometrics: Concepts, methods, and applications. Symmetry 2015, 7, 843–934. [Google Scholar] [CrossRef]
- Bookstein, F.L. Landmark methods for forms without landmarks: Morphometrics of group differences in outline shape. Med. Image Anal. 1997, 1, 225–243. [Google Scholar] [CrossRef]
- Klingenberg, C.P.; McIntyre, G.S. Geometric morphometrics of developmental instability: Analyzing patterns of fluctuating asymmetry with Procrustes methods. Evolution 1998, 52, 1363–1375. [Google Scholar] [CrossRef]
- Claes, P.; Reijniers, J.; Shriver, M.D.; Snyders, J.; Suetens, P.; Nielandt, J.; De Tré, G.; Vandermeulen, D. An investigation of matching symmetry in the human pinnae with possible implications for 3D ear recognition and sound localization. J. Anat. 2015, 226, 60–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Claes, P.; Walters, M.; Shriver, M.D.; Puts, D.; Gibson, G.; Clement, J.; Baynam, G.; Verbeke, G.; Vandermeulen, D.; Suetens, P. Sexual dimorphism in multiple aspects of 3D facial symmetry and asymmetry defined by spatially dense geometric morphometrics. J. Anat. 2012, 221, 97–114. [Google Scholar] [CrossRef] [PubMed]
- Schlager, S. Soft-tissue Reconstruction of the Human Nose. Ph.D. Thesis, Albert-Ludwigs-Universität, Freiburg, Germany, 2013. [Google Scholar]
- Schlager, S.; Rüdell, A. Analysis of the human osseous nasal shape—Population differences and sexual dimorphism. Am. J. Phys. Anthropol. 2015, 157, 571–581. [Google Scholar] [CrossRef] [PubMed]
- Zabrodsky, H.; Peleg, S.; Avnir, D. Symmetry as a continuous feature. IEEE Trans. Pattern Anal. 1995, 17, 1154–1166. [Google Scholar] [CrossRef]
- Graham, J.H.; Whitesell, M.J.; Fleming, M.I.; Hel-Or, H.; Nevo, E.; Raz, S. Fluctuating asymmetry of plant leaves: Batch processing with LAMINA and Continuous Symmetry Measures. Symmetry 2015, 7, 255–268. [Google Scholar] [CrossRef]
- Zabrodsky, H.; Peleg, S.; Avnir, D. Continuous symmetry measures. J. Am. Chem. Soc. 1992, 114, 7843–7851. [Google Scholar] [CrossRef]
- Palmer, A.R. Fluctuating asymmetry analyses: A primer. In Developmental Instability: Its Origins and Evolutionary Implications; Markow, T.A., Ed.; Kluwer: Dordrecht, The Netherlands, 1994; pp. 335–364. [Google Scholar]
- Holt, S.B. The correlations between ridge-counts on different fingers estimated from a population sample. Ann. Hum. Genet. 1959, 23, 459–460. [Google Scholar] [CrossRef] [PubMed]
- Mellor, C.S. Dermatoglyphic evidence of fluctuating asymmetry in schizophrenia. Br. J. Psychiat. 1992, 160, 467–472. [Google Scholar] [CrossRef]
- Angus, R.A. Quantifying fluctuating asymmetry—Not all methods are equivalent. Growth 1982, 46, 337–342. [Google Scholar]
- Shakibaei, F.; Asadollahi, G.A.; Tabibi, A.P. Dermatoglyphics in patients with schizophrenia. J. Res. Med. Sci. 2011, 16, 1055–1061. [Google Scholar] [PubMed]
- Bookstein, F.L. Morphometric Tools for Landmark Data; Cambridge University Press: Cambridge, UK, 1997; p. 436. [Google Scholar]
- Hopman, S.M.J.; Merks, J.H.M.; Suttie, M.; Hennekam, R.; Hammond, P. 3D morphometry aids facial analysis of individuals with a childhood cancer. Am. J. Med. Genet. A 2016, 9999A, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Graham, J.H.; Shimizu, K.; Emlen, J.M.; Freeman, D.; Merkel, J. Growth models and the expected distribution of fluctuating asymmetry. Biol. J. Linn. Soc. 2003, 80, 57–65. [Google Scholar] [CrossRef]
- Mosimann, J.E.; Campbell, G. Applications in biology: Simple growth models. In Lognormal Distributions: Theory and Applications; Crow, E.L., Shimizu, K., Eds.; Marcel Dekker: New York, NY, USA, 1988; pp. 287–302. [Google Scholar]
- Holt, S.B. Genetics of dermal ridges: Bilateral asymmetry in finger ridge-counts. Ann. Eugenic. 1952, 17, 211–231. [Google Scholar] [CrossRef]
- Livshits, G.; Yakovenko, K.; Kletselman, L.; Karasik, D.; Kobyliansky, E. Fluctuating asymmetry and morphometric variation of hand bones. Am. J. Phys. Anthropol. 1998, 107, 125–136. [Google Scholar] [CrossRef]
- Gutnik, B.; Skurvydas, A.; Zuoza, A.; Zuoziene, I.; Mickevičnienė, D.; Alekrinskis, A.; Nash, D. Evaluation of bilateral asymmetry between upper limb masses in right-handed young adults of both sexes. Percept. Mot. Skills 2015, 120, 804–815. [Google Scholar] [CrossRef] [PubMed]
- Graham, J.H.; Emlen, J.M.; Freeman, D.C.; Leamy, L.J.; Kieser, J.A. Directional asymmetry and the measurement of developmental instability. Biol. J. Linn. Soc. 1998, 64, 1–16. [Google Scholar] [CrossRef]
- Auerbach, B.M.; Ruff, C.B. Limb bone bilateral asymmetry: Variability and commonality among modern humans. J. Hum. Evol. 2006, 50, 203–218. [Google Scholar] [CrossRef] [PubMed]
- Latimer, H.B.; Lowrance, E.W. Bilateral asymmetry in weight and in length of human bones. Anat. Rec. 1965, 152, 217–224. [Google Scholar] [CrossRef] [PubMed]
- Macintosh, A.A.; Pinhasi, R.; Stock, J.T. Divergence in male and female manipulative behaviors with the intensification of metallurgy in Central Europe. PLoS ONE 2014, 9, e112116. [Google Scholar] [CrossRef] [PubMed]
- Hartman, C.; Holton, N.; Miller, S.; Yokley, T.; Marshall, S.; Srinivasan, S.; Southard, T. Nasal septal deviation and facial skeletal asymmetries. Anat. Rec. 2016, 299, 295–306. [Google Scholar] [CrossRef] [PubMed]
- Kanat, A.; Yazar, U.; Ozdemir, B.; Coskun, Z.O.; Erdivanli, O. Frontal sinus asymmetry: Is it an effect of cranial asymmetry? X-ray analysis of 469 normal adult human frontal sinus. J. Neurosci. Rural Pract. 2015, 6, 511–514. [Google Scholar] [CrossRef] [PubMed]
- Johnson, W.; Segal, N.L.; Bouchard, T.J. Fluctuating asymmetry and general intelligence: No genetic or phenotypic association. Intelligence 2008, 36, 279–288. [Google Scholar] [CrossRef]
- Van Dongen, S.; Lens, L.; Molenberghs, G. Mixture analysis of asymmetry: Modelling directional asymmetry, antisymmetry and heterogeneity in fluctuating asymmetry. Ecol. Lett. 1999, 2, 387–396. [Google Scholar]
- Proff, P.; Römer, P. The molecular mechanism behind bone remodelling: A review. Clin. Oral Investig. 2009, 13, 355–362. [Google Scholar] [CrossRef] [PubMed]
- Hallgrímsson, B. Ontogenetic patterning of skeletal fluctuating asymmetry in rhesus macaques and humans: Evolutionary and developmental implications. Int. J. Primatol. 1999, 20, 121–151. [Google Scholar] [CrossRef]
- Palestis, B.G.; Trivers, R. A longitudinal study of changes in fluctuating asymmetry with age in Jamaican youth. Symmetry 2016, 8. [Google Scholar] [CrossRef]
- Guatelli-Steinberg, D. What Teeth Reveal about Human Evolution; Cambridge University Press: Cambridge, UK, 2016; p. 294. [Google Scholar]
- Gawlikowska-Sroka, A.; Dąbrowski, P.; Szczurowski, J.; Staniowski, T. Analysis of interaction between nutritional and developmental instability in mediaeval population in Wrocław. Anthropol. Rev. 2013, 76, 51–62. [Google Scholar] [CrossRef]
- Lawn, B.R.; Lee, J.J.-W.; Chai, H. Teeth: Among nature’s most durable biocomposites. Annu. Rev. Mater. Res. 2010, 40, 55–75. [Google Scholar] [CrossRef]
- Niswander, J.D.; Chung, C.S. The effects of inbreeding on tooth size in Japanese children. Am. J. Hum. Genet. 1965, 17, 390–398. [Google Scholar] [PubMed]
- Garn, S.M.; Lewis, A.B.; Kerewsky, R.S. The meaning of bilateral asymmetry in the permanent dentition. Angle Orthod. 1966, 36, 55–62. [Google Scholar] [PubMed]
- Garn, S.M.; Lewis, A.B.; Kerewsky, R.S. Buccolingual size asymmetry and its developmental meaning. Angle Orthod. 1967, 37, 186–193. [Google Scholar] [PubMed]
- Barrett, C.K.; Guatelli-Steinberg, D.; Sciulli, P.W. Revisiting dental fluctuating asymmetry in neandertals and modern humans. Am. J. Phys. Anthropol. 2012, 149, 193–204. [Google Scholar] [CrossRef] [PubMed]
- Sciulli, P.W. Dental asymmetry in a Late Archaic and late prehistoric skeletal sample of the Ohio Valley Area. Dent. Anthropol. 2002, 16, 33–44. [Google Scholar]
- Pietrusewsky, M.; Douglas, M.T. Database E.4 Individual Records of Measurements of Permanent Dentitions in Adolescents and Adults. Available online: https://core.tdar.org/dataset/376929/database-e4-individual-records-of-measurements-of-permanent-dentitions-in-adolescents-and-adults (accessed on 9 November 2016).
- Varner, T.B. Landmark-Based Approach to Examining Changes in Arch Shape: A Longitudinal Study. Master’s Thesis, University of Iowa, Iowa City, IA, USA, 2015. [Google Scholar]
- Grammer, K.; Thornhill, R. Human (Homo sapiens) facial attractiveness and sexual selection: The role of symmetry and averageness. J. Comp. Psychol. 1994, 108, 233–242. [Google Scholar] [CrossRef] [PubMed]
- Miller, S.F.; Weinberg, S.M.; Nidey, N.L.; Defay, D.K.; Marazita, M.L.; Wehby, G.L.; Moreno Uribe, L.M. Exploratory genotype–phenotype correlations of facial form and asymmetry in unaffected relatives of children with non-syndromic cleft lip and/or palate. J. Anat. 2014, 224, 688–709. [Google Scholar] [CrossRef] [PubMed]
- Cummins, H.; Midlo, C. Palmar and plantar epidermal ridge configurations (dermatoglyphics) in European-Americans. Am. J. Phys. Anthropol. 1926, 9, 471–502. [Google Scholar] [CrossRef]
- Cummins, H.; Midlo, C. Finger Prints, Palms and Soles: An Introduction to Dermatoglyphics; Dover Publications: New York, NY, USA, 1961; p. 319. [Google Scholar]
- Galton, F. Finger Prints; Macmillan and Company: London, UK, 1892; p. 256. [Google Scholar]
- Loesch, D.Z. Quantitative Dermatoglyphics: Classification, Genetics, and Pathology; Oxford University Press: New York, NY, USA, 1983; p. 450. [Google Scholar]
- Babler, W.J. Embryologic development of epidermal ridges and their configurations. Birth Defects Orig. Artic. Ser. 1991, 27, 95–112. [Google Scholar] [PubMed]
- Kücken, M.; Newell, A.C. Fingerprint formation. J. Theor. Biol. 2005, 235, 71–83. [Google Scholar] [CrossRef] [PubMed]
- Arrieta, M.I.; Criado, B.; Martinez, B.; Lobato, M.N.; Gil, A.; Lostao, C.M. Fluctuating dermatoglyphic asymmetry: Genetic and prenatal influences. Ann. Hum. Biol. 1993, 20, 557–563. [Google Scholar] [CrossRef] [PubMed]
- Rose, R.J.; Reed, T.; Bogle, A. Asymmetry of a–b ridge count and behavioral discordance of monozygotic twins. Behav. Genet. 1987, 17, 125–140. [Google Scholar] [CrossRef] [PubMed]
- King, S.; Dancause, K.; Turcotte-Tremblay, A.-M.; Veru, F.; Laplante, D.P. Using natural disasters to study the effects of prenatal maternal stress on child health and development. Birth Defects Res. C 2012, 96, 273–288. [Google Scholar] [CrossRef] [PubMed]
- King, S.; Mancini-Marïe, A.; Brunet, A.; Walker, E.; Meaney, M.J.; Laplante, D.P. Prenatal maternal stress from a natural disaster predicts dermatoglyphic asymmetry in humans. Dev. Psychopathol. 2009, 21, 343–353. [Google Scholar] [CrossRef] [PubMed]
- Penrose, L.S. The distal triradius t on the hands of parents and sibs of mongol imbeciles. Ann. Hum. Genet. 1954, 19, 10–38. [Google Scholar] [CrossRef] [PubMed]
- Jantz, R.L.; Brehme, H. Directional and fluctuating asymmetry in the palmar interdigital ridge-counts. Anthropol. Anz. 1993, 51, 59–67. [Google Scholar] [PubMed]
- Jantz, R.L.; Webb, R.S. Dermatoglyphic asymmetry as a measure of canalization. Ann. Hum. Biol. 1980, 7, 489–493. [Google Scholar] [CrossRef] [PubMed]
- Wijerathne, B.T.B.; Meier, R.J.; Agampodi, T.C.; Agampodi, S.B. Dermatoglyphics in hypertension: A review. J. Physiol. Anthropol. 2015, 34. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.-F.; Lin, C.-L.; Yen, C.-W.; Chang, Y.-H.; Chen, T.-Y.; Su, K.-P.; Nagurka, M.L. Determining the association between dermatoglyphics and schizophrenia by using fingerprint asymmetry measures. Int. J. Pattern Recogn. 2008, 22, 601–616. [Google Scholar] [CrossRef]
- De Bruin, E.I.; Graham, J.H.; Louwerse, A.; Huizink, A.C. Mild dermatoglyphic deviations in adolescents with autism spectrum disorders and average intellectual abilities as compared to typically developing boys. Autism Res. Treat. 2014, 2014, 968134. [Google Scholar] [CrossRef] [PubMed]
- Markow, T.A.; Wandler, K. Fluctuating dermatoglyphic asymmetry and the genetics of liability to schizophrenia. Psychiatry Res. 1986, 19, 323–328. [Google Scholar] [CrossRef]
- Holt, S.B. Genetics of dermal ridges: The relation between total ridge-count and the variability of counts from finger to finger. Ann. Hum. Genet. 1958, 22, 323–339. [Google Scholar] [CrossRef] [PubMed]
- Holt, S.B. Genetics of dermal ridges: Familial correlations for (S/√10), a measurement of the diversity of ridge-counts from finger to finger. Ann. Hum. Genet. 1960, 24, 253–269. [Google Scholar] [CrossRef] [PubMed]
- Micle, S.; Kobyliansky, E. Dermatoglyphic sexual dimorphism in Israelis: Principal components and discriminant analyses applied to quantitative traits. Hum. Biol. 1986, 58, 485–498. [Google Scholar] [PubMed]
- Karmakar, B.; Yakovenko, K.; Kobyliansky, E. Sexual dimorphism: Asymmetry and diversity of 38 dermatoglyphic traits in five endogamous populations of West Bengal, India. Coll. Antropol. 2001, 25, 167–187. [Google Scholar] [PubMed]
- Mathew, L.; Hegde, A.M.; Rai, K. Dermatoglyphic peculiarities in children with oral clefts. J. Indian Soc. Ped. Prev. Dent. 2005, 23, 179–182. [Google Scholar]
- Bonnevie, K. Studies on papillary patterns of human fingers. J. Genet. 1924, 15, 1–111. [Google Scholar] [CrossRef]
- Ponnudurai, R.; Menon, M.S.; Muthu, M. Dermatoglyphic fluctuating asymmetry and symmetry in familial and non familial schizophrenia. Indian J. Psychiatry 1997, 39, 205–211. [Google Scholar] [PubMed]
- Ravindranath, R.; Joseph, A.M.; Bosco, S.I.; Rajangam, S.; Balasubramanyam, V. Fluctuating asymmetry in dermatoglyphics of non-insulin-dependent diabetes mellitus in Bangalore-based population. Indian J. Hum. Genet. 2005, 11, 149–153. [Google Scholar] [CrossRef]
- Saha, S.; Loesch, D.; Chant, D.; Welham, J.; El-Saadi, O.; Fañanás, L.; Mowry, B.; McGrath, J. Directional and fluctuating asymmetry in finger and ab ridge counts in psychosis: A case-control study. BMC Psychiatry 2003, 3, 1–9. [Google Scholar] [CrossRef]
- Russak, O. Dermatoglyphic Asymmetries, Symptoms, and Cognitive Function in Adolescents at Ultrahigh-risk for Psychotic Disorders. Bachelor’s Thesis, University of Colorado, Boulder, CO, USA, 2015. [Google Scholar]
- Russak, O.D.F.; Ives, L.; Mittal, V.A.; Dean, D.J. Fluctuating dermatoglyphic asymmetries in youth at ultrahigh-risk for psychotic disorders. Schizophr. Res. 2016, 170, 301–303. [Google Scholar] [CrossRef] [PubMed]
- Sivkov, S.T.; Akabaliev, V.H.; Akabalieva, K.V. Fluctuating asymmetry in dermatoglyphic traits in schizophrenic patients. Folia Med. 2007, 49, 5–10. [Google Scholar]
- Acree, M.A. Is there a gender difference in fingerprint ridge density? Forensic Sci. Int. 1999, 102, 35–44. [Google Scholar] [CrossRef]
- Cantalupo, C.; Hopkins, W.D. Asymmetric Broca’s area in great apes. Nature 2001, 414, 505. [Google Scholar] [CrossRef] [PubMed]
- Thoma, R.J.; Yeo, R.A.; Gangestad, S.W.; Lewine, J.D.; Davis, J.T. Fluctuating asymmetry and the human brain. Laterality 2002, 7, 45–58. [Google Scholar] [CrossRef] [PubMed]
- Yeo, R.A.; Gangestad, S.W.; Thoma, R.; Shaw, P.; Repa, K. Developmental instability and cerebral lateralization. Neuropsychology 1997, 11, 552–561. [Google Scholar] [CrossRef] [PubMed]
- Schulter, G.; Papousek, I. Believing in paranormal phenomena: Relations to asymmetry of body and brain. Cortex 2008, 44, 1326–1335. [Google Scholar] [CrossRef] [PubMed]
- Manning, J.T.; Scutt, D.; Whitehouse, G.H.; Leinster, S.J. Breast asymmetry and phenotypic quality in women. Evol. Hum. Behav. 1997, 18, 223–236. [Google Scholar] [CrossRef]
- Manning, J.T.; Scutt, D.; Whitehouse, G.H.; Leinster, S.J.; Walton, J.M. Asymmetry and the menstrual cycle in women. Ethol. Sociobiol. 1996, 17, 129–143. [Google Scholar] [CrossRef]
- Domjanic, J.; Fieder, M.; Seidler, H.; Mitteroecker, P. Geometric morphometric footprint analysis of young women. J. Foot Ankle Res. 2013, 6, 27. [Google Scholar] [CrossRef] [PubMed]
- Schlager, S. Sliding semi-landmarks on symmetric structures in three dimensions. Am. J. Phys. Anthropol. 2012, 147, 261. [Google Scholar]
- Alados, C.L.; Escós, J.; Emlen, J.M. Fluctuating asymmetry and fractal dimension of the sagittal suture as indicators of inbreeding depression in dama and dorcas gazelles. Can. J. Zool. 1995, 73, 1967–1974. [Google Scholar] [CrossRef]
- Escos, J.M.; Alados, C.L.; Emlen, J.M. Fractal structures and fractal functions as disease indicators. Oikos 1995, 74, 310–314. [Google Scholar] [CrossRef]
- Graham, J.H.; Emlen, J.M.; Freeman, D.C. Developmental stability and its applications in ecotoxicology. Ecotoxicology 1993, 2, 175–184. [Google Scholar] [CrossRef] [PubMed]
- Kuswandari, S.; Nishino, M. The mesiodistal crown diameters of primary dentition in Indonesian Javanese children. Arch. Oral Biol. 2004, 49, 217–222. [Google Scholar] [CrossRef] [PubMed]
- Albert, A.M.; Greene, D.L. Bilateral asymmetry in skeletal growth and maturation as an indicator of environmental stress. Am. J. Phys. Anthropol. 1999, 110, 341–349. [Google Scholar] [CrossRef]
- Bigoni, L.; Krajíček, V.; Sládek, V.; Velemínský, P.; Velemínská, J. Skull shape asymmetry and the socioeconomic structure of an early medieval central European society. Am. J. Phys. Anthropol. 2013, 150, 349–364. [Google Scholar] [CrossRef] [PubMed]
- Gray, P.B.; Marlowe, F. Fluctuating asymmetry of a foraging population: The Hadza of Tanzania. Ann. Hum. Biol. 2002, 29, 495–501. [Google Scholar] [CrossRef] [PubMed]
- Guatelli-Steinberg, D.; Sciulli, P.W.; Edgar, H.H.J. Dental fluctuating asymmetry in the Gullah: Tests of hypotheses regarding developmental stability in deciduous vs permanent and male vs female teeth. Am. J. Phys. Anthropol. 2006, 129, 427–434. [Google Scholar] [CrossRef] [PubMed]
- Livshits, G.; Smouse, P.E. Multivariate fluctuating asymmetry in Israeli adults. Hum. Biol. 1993, 65, 547–578. [Google Scholar] [PubMed]
- Özener, B. Fluctuating and directional asymmetry in young human males: Effect of heavy working condition and socioeconomic status. Am. J. Phys. Anthropol. 2010, 143, 112–120. [Google Scholar] [CrossRef] [PubMed]
- Simmons, L.W.; Rhodes, G.; Peters, M.; Koehler, N. Are human preferences for facial symmetry focused on signals of developmental instability? Behav. Ecol. 2004, 15, 864–871. [Google Scholar] [CrossRef]
- Trivers, R.; Manning, J.T.; Thornhill, R.; Singh, D.; McGuire, M. Jamaican symmetry project: Long-term study of fluctuating asymmetry in rural Jamaican children. Hum. Biol. 1999, 417–430. [Google Scholar]
- Wilson, J.M.; Manning, J.T. Fluctuating asymmetry and age in children: Evolutionary implications for the control of developmental stability. J. Hum. Evol. 1996, 30, 529–537. [Google Scholar] [CrossRef]
- Noss, J.F.; Scott, G.R.; Potter, R.H.Y.; Dahlberg, A.A. Fluctuating asymmetry in molar dimensions and discrete morphological traits in Pima Indians. Am. J. Phys. Anthropol. 1983, 61, 437–445. [Google Scholar] [CrossRef] [PubMed]
- Özener, B.; Fink, B. Facial symmetry in young girls and boys from a slum and a control area of Ankara, Turkey. Evol. Hum. Behav. 2010, 31, 436–441. [Google Scholar] [CrossRef]
- Ertuğrul, B. Sexually dimorphic human body fat distribution and second-to-fourth digit ratio. Eurasian J. Anthropol. 2013, 3, 54–62. [Google Scholar]
- Ertuğrul, B.; Otağ, İ. Cinsiyete özgü vücut morfolojisi ile elin ikinci ve dördüncü parmak uzunluk oranları arasındaki ilişki. İnsanbilim Dergisi 2012, 1, 94–107. [Google Scholar]
- Fink, B.; Manning, J.T.; Neave, N.; Grammer, K. Second to fourth digit ratio and facial asymmetry. Evol. Hum. Behav. 2004, 25, 125–132. [Google Scholar] [CrossRef]
- Benderlioglu, Z. Fluctuating asymmetry and steroid hormones: A review. Symmetry 2010, 2, 541–553. [Google Scholar] [CrossRef]
- Özener, B. Brief communication: Facial fluctuating asymmetry as a marker of sex differences of the response to phenotypic stresses. Am. J. Phys. Anthropol. 2010, 143, 321–324. [Google Scholar] [CrossRef] [PubMed]
- Davis, E.P.; Sandman, C.A. The timing of prenatal exposure to maternal cortisol and psychosocial stress is associated with human infant cognitive development. Child Dev. 2010, 81, 131–148. [Google Scholar] [CrossRef] [PubMed]
- Bogin, B. Patterns of Human Growth; Cambridge University Press: Cambridge, UK, 1999; Volume 23, p. 472. [Google Scholar]
- Kieser, J.A.; Groeneveld, H.T.; Preston, C.B. Fluctuating odontometric asymmetry in the Lengua Indians of Paraguay. Ann. Hum. Biol. 1986, 13, 489–498. [Google Scholar] [CrossRef] [PubMed]
- Livshits, G.; Davidi, L.; Kobyliansky, E.; Ben-Amitai, D.; Levi, Y.; Merlob, P.; Optiz, J.M.; Reynolds, J.F. Decreased developmental stability as assessed by fluctuating asymmetry of morphometric traits in preterm infants. Am. J. Med. Genet. 1988, 29, 793–805. [Google Scholar] [CrossRef] [PubMed]
- Little, B.B.; Buschang, P.H.; Malina, R.M. Anthropometric asymmetry in chronically undernourished children from Southern Mexico. Ann. Hum. Biol. 2002, 29, 526–537. [Google Scholar] [CrossRef] [PubMed]
- Flinn, M.V.; Leone, D.V.; Quinlan, R.J. Growth and fluctuating asymmetry of stepchildren. Evol. Hum. Behav. 1999, 20, 465–479. [Google Scholar] [CrossRef]
- Wells, J.C.; Hallal, P.C.; Manning, J.T.; Victora, C.G. A trade-off between early growth rate and fluctuating asymmetry in Brazilian boys. Ann. Hum. Biol. 2006, 33, 112–124. [Google Scholar] [CrossRef] [PubMed]
- Özener, B.; Ertuğrul, B. Relationship between shortness of final body height and fluctuating asymmetry in Turkish young males. Ann. Hum. Biol. 2011, 38, 34–38. [Google Scholar] [CrossRef] [PubMed]
- Özener, B. Biomechanical pressures and upper extremity asymmetry: A study on young laborers. Coll. Antropol. 2007, 31, 693–699. [Google Scholar] [PubMed]
- Tomaszewska, A.; Kwiatkowska, B.; Jankauskas, R. Is the area of the orbital opening in humans related to climate? Am. J. Hum. Biol. 2015, 27, 845–850. [Google Scholar] [CrossRef] [PubMed]
- Freeman, D.C.; Graham, J.H.; Emlen, J.M. Developmental stability in plants: Symmetries, stress and epigenesis. Genetica 1993, 89, 97–119. [Google Scholar] [CrossRef]
- Wuytack, T.; Samson, R.; Wuyts, K.; Adriaenssens, S.; Kardel, F.; Verheyen, K. Do Leaf Characteristics of White Willow (Salix alba L.), Northern Red Oak (Quercus rubra L.), and Scots Pine (Pinus sylvestris L.) Respond Differently to Ambient Air Pollution and Other Environmental Stressors? Water Air Soil Pollut. 2013, 224. [Google Scholar] [CrossRef]
- Wuytack, T.; Wuyts, K.; Van Dongen, S.; Baeten, L.; Kardel, F.; Verheyen, K.; Samson, R. The effect of air pollution and other environmental stressors on leaf fluctuating asymmetry and specific leaf area of Salix alba L. Environ. Poll. 2011, 159, 2405–2411. [Google Scholar] [CrossRef] [PubMed]
- Polak, M.; Opoka, R.; Cartwright, I.L. Response of fluctuating asymmetry to arsenic toxicity: Support for the developmental selection hypothesis. Environ. Poll. 2002, 118, 19–28. [Google Scholar] [CrossRef]
- Savriama, Y.; Stige, L.C.; Gerber, S.; Pérez, T.; Alibert, P.; David, B. Impact of sewage pollution on two species of sea urchins in the Mediterranean Sea (Cortiou, France): Radial asymmetry as a bioindicator of stress. Ecol. Indic. 2015, 54, 39–47. [Google Scholar] [CrossRef]
- Allenbach, D.M. Fluctuating asymmetry and exogenous stress in fishes: A review. Rev. Fish Biol. Fish. 2011, 21, 355–376. [Google Scholar] [CrossRef]
- Lajus, D.; Yurtseva, A.; Birch, G.; Booth, D.J. Fluctuating asymmetry as a pollution monitor: The Australian estuarine smooth toadfish Tetractenos glaber (Teleostei: Tetraodontidae). Mar. Pollut. Bull. 2015, 101, 758–767. [Google Scholar] [CrossRef] [PubMed]
- Costa, R.N.; Nomura, F. Measuring the impacts of Roundup Original® on fluctuating asymmetry and mortality in a Neotropical tadpole. Hydrobiologia 2016, 765, 85–96. [Google Scholar] [CrossRef]
- Lazić, M.M.; Kaliontzopoulou, A.; Carretero, M.A.; Crnobrnja-Isailović, J. Lizards from urban areas are more asymmetric: Using fluctuating asymmetry to evaluate environmental disturbance. PLoS ONE 2013, 8, e84190. [Google Scholar] [CrossRef] [PubMed]
- Eeva, T.; Tanhuanpää, S.; Råbergh, C.; Airaksinen, S.; Nikinmaa, M.; Lehikoinen, E. Biomarkers and fluctuating asymmetry as indicators of pollution-induced stress in two hole-nesting passerines. Funct. Ecol. 2000, 14, 235–243. [Google Scholar] [CrossRef]
- Sánchez-Chardi, A.; García-Pando, M.; López-Fuster, M.J. Chronic exposure to environmental stressors induces fluctuating asymmetry in shrews inhabiting protected Mediterranean sites. Chemosphere 2013, 93, 916–923. [Google Scholar] [CrossRef] [PubMed]
- Beasley, D.A.E.; Bonisoli-Alquati, A.; Mousseau, T.A. The use of fluctuating asymmetry as a measure of environmentally induced developmental instability: A meta-analysis. Ecol. Indic. 2013, 30, 218–226. [Google Scholar] [CrossRef]
- Philp, R.B. Ecosystems and Human Health: Toxicology and Environmental Hazards; CRC Press: Boca Raton, FL, USA, 2015; p. 440. [Google Scholar]
- Kieser, J.A. Fluctuating odontometric asymmetry and maternal alcohol consumption. Ann. Hum. Biol. 1992, 19, 513–520. [Google Scholar] [CrossRef] [PubMed]
- Wilber, E.; Newell-Morris, L.; Streissguth, A.P. Dermatoglyphic asymmetry in fetal alcohol syndrome. Neonatology 1993, 64, 1–6. [Google Scholar] [CrossRef]
- Klingenberg, C.P.; Wetherill, L.; Rogers, J.; Moore, E.; Ward, R.; Autti-Rämö, I.; Fagerlund, Å.; Jacobson, S.W.; Robinson, L.K.; Hoyme, H.E. Prenatal alcohol exposure alters the patterns of facial asymmetry. Alcohol 2010, 44, 649–657. [Google Scholar] [CrossRef] [PubMed]
- Kieser, J.A.; Groeneveld, H.T. Effects of prenatal exposure to tobacco smoke on developmental stability in children. J. Craniofac. Genet. Dev. Biol. 1994, 14, 43–47. [Google Scholar] [PubMed]
- Kieser, J.A.; Groeneveld, H.T.; Da Silva, P.C.F. Dental asymmetry, maternal obesity, and smoking. Am. J. Phys. Anthropol. 1997, 102, 133–139. [Google Scholar] [CrossRef]
- Epstein, C.J. The Consequences of Chromosome Imbalance: Principles, Mechanisms, and Models; Cambridge University Press: Cambridge, UK, 2007; Volume 18, p. 510. [Google Scholar]
- Reeves, R.H.; Baxter, L.L.; Richtsmeier, J.T. Too much of a good thing: Mechanisms of gene action in Down syndrome. Trends Genet. 2001, 17, 83–88. [Google Scholar] [CrossRef]
- Shapiro, B.L. Development of human autosomal aneuploid phenotypes (with an emphasis on Down syndrome). Acta Zool. Fenn. 1992, 191, 97–105. [Google Scholar]
- Shapiro, B.L.; Hermann, J.; Opitz, J.M. Down syndrome—A disruption of homeostasis. Am. J. Med. Genet. 1983, 14, 241–269. [Google Scholar] [CrossRef] [PubMed]
- Benda, C.E. Down’s Syndrome: Mongolism and Its Management; Grune & Stratton: New York, NY, USA, 1969; p. 296. [Google Scholar]
- Antonarakis, S.E.; Lyle, R.; Dermitzakis, E.T.; Reymond, A.; Deutsch, S. Chromosome 21 and Down syndrome: From genomics to pathophysiology. Nat. Rev. Genet. 2004, 5, 725–738. [Google Scholar] [CrossRef] [PubMed]
- Korenberg, J.R.; Chen, X.N.; Schipper, R.; Sun, Z.; Gonsky, R.; Gerwehr, S.; Carpenter, N.; Daumer, C.; Dignan, P.; Disteche, C. Down syndrome phenotypes: The consequences of chromosomal imbalance. Proc. Natl. Acad. Sci. USA 1994, 91, 4997–5001. [Google Scholar] [CrossRef] [PubMed]
- Roizen, N.J.; Patterson, D. Down’s syndrome. Lancet 2003, 361, 1281–1289. [Google Scholar] [CrossRef]
- Garn, S.M.; Cohen, M.M.; Geciauskas, M.A. Increased crown-size asymmetry in trisomy G. J. Dent. Res. 1970, 49, 465. [Google Scholar] [CrossRef] [PubMed]
- Barden, H.S. Fluctuating dental asymmetry: A measure of developmental instability in Down syndrome. Am. J. Phys. Anthropol. 1980, 52, 169–173. [Google Scholar] [CrossRef] [PubMed]
- Townsend, G.C. Fluctuating dental asymmetry in Down’s syndrome. Aust. Dent. J. 1983, 28, 39–44. [Google Scholar] [CrossRef] [PubMed]
- Starbuck, J.M.; Cole, T.M.; Reeves, R.H.; Richtsmeier, J.T. Trisomy 21 and facial developmental instability. Am. J. Phys. Anthropol. 2013, 151, 49–57. [Google Scholar] [CrossRef] [PubMed]
- Plato, C.C.; Cereghino, J.J.; Steinberg, F.S. Palmar dermatoglyphics of Down’s syndrome: Revisited. Pediatr. Res. 1973, 7, 111–118. [Google Scholar] [CrossRef] [PubMed]
- Bots, J.; ten Broek, C.M.A.; Belien, J.A.M.; Bugiani, M.; Galis, F.; Van Dongen, S. Higher limb asymmetry in deceased human fetuses and infants with aneuploidy. Sci. Rep. 2014, 4, 3703. [Google Scholar] [CrossRef] [PubMed]
- Hartl, D.L.; Ruvolo, M. Genetics; Jones & Bartlett Publishers: Sudbury, MA, USA, 2011; p. 804. [Google Scholar]
- Debat, V.; Bloyer, S.; Faradji, F.; Gidaszewski, N.; Navarro, N.; Orozco-terWengel, P.; Ribeiro, V.; Schlötterer, C.; Deutsch, J.S.; Peronnet, F. Developmental stability: A major role for Cyclin G in Drosophila melanogaster. PLoS Genet. 2011, 7, e1002314. [Google Scholar] [CrossRef] [PubMed]
- Leamy, L.J.; Workman, M.S.; Routman, E.J.; Cheverud, J.M. An epistatic genetic basis for fluctuating asymmetry of tooth size and shape in mice. Heredity 2005, 94, 316–325. [Google Scholar] [CrossRef] [PubMed]
- Leamy, L.J.; Klingenberg, C.P. The genetics and evolution of fluctuating asymmetry. Annu. Rev. Ecol. Evol. Syst. 2005, 36, 1–21. [Google Scholar] [CrossRef]
- Leamy, L.J.; Routman, E.J.; Cheverud, J.M. Quantitative trait loci for fluctuating asymmetry of discrete skeletal characters in mice. Heredity 1998, 80, 509–518. [Google Scholar] [CrossRef] [PubMed]
- Leamy, L.J.; Routman, E.J.; Cheverud, J.M. An epistatic genetic basis for fluctuating asymmetry of mandible size in mice. Evolution 2002, 56, 642–653. [Google Scholar] [CrossRef] [PubMed]
- Charlesworth, D.; Willis, J.H. The genetics of inbreeding depression. Nat. Rev. Genet. 2009, 10, 783–796. [Google Scholar] [CrossRef] [PubMed]
- Keller, L.F.; Waller, D.M. Inbreeding effects in wild populations. Trends Ecol. Evol. 2002, 17, 230–241. [Google Scholar] [CrossRef]
- Meagher, S.; Penn, D.J.; Potts, W.K. Male–male competition magnifies inbreeding depression in wild house mice. Proc. Natl. Acad. Sci. USA 2000, 97, 3324–3329. [Google Scholar] [CrossRef] [PubMed]
- Miller, P.S. Is inbreeding depression more severe in a stressful environment? Zoo Biol. 1994, 13, 195–208. [Google Scholar] [CrossRef]
- Bihlmeyer, N.A.; Brody, J.A.; Smith, A.V.; Lunetta, K.L.; Nalls, M.; Smith, J.A.; Tanaka, T.; Davies, G.; Yu, L.; Mirza, S.S. Genetic diversity is a predictor of mortality in humans. BMC Genet. 2014, 15, 159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bittles, A.H. Time to get real: Investigating potential beneficial genetic aspects of consanguinity. Public Health Genom. 2011, 14, 169–171. [Google Scholar] [CrossRef] [PubMed]
- Bittles, A.H.; Black, M.L. The impact of consanguinity on neonatal and infant health. Early Hum. Dev. 2010, 86, 737–741. [Google Scholar] [CrossRef] [PubMed]
- Waldmann, P. The effect of inbreeding on fluctuating asymmetry in Scabiosa canescens (Dipsacaceae). Evol. Ecol. 2001, 15, 117–127. [Google Scholar] [CrossRef]
- Beardmore, J.A. Developmental stability in constant and fluctuating temperatures. Heredity 1960, 14, 411–422. [Google Scholar] [CrossRef]
- Carter, A.J.R.; Weier, T.M.; Houle, D. The effect of inbreeding on fluctuating asymmetry of wing veins in two laboratory strains of Drosophila melanogaster. Heredity 2009, 102, 563–572. [Google Scholar] [CrossRef] [PubMed]
- Gomendio, M.; Cassinello, J.; Roldan, E.R.S. A comparative study of ejaculate traits in three endangered ungulates with different levels of inbreeding: Fluctuating asymmetry as an indicator of reproductive and genetic stress. Proc. R. Soc. Lond. B. Biol. Sci. 2000, 267, 875–882. [Google Scholar] [CrossRef] [PubMed]
- Leamy, L. Morphometric studies in inbred and hybrid house mice. V. Directional and fluctuating asymmetry. Am. Nat. 1984, 123, 579–593. [Google Scholar] [CrossRef]
- Van Noordwijk, A.J.; Scharloo, W. Inbreeding in an island population of the great tit. Evolution 1981, 35, 674–688. [Google Scholar] [CrossRef]
- Fowler, K.; Whitlock, M.C. Fluctuating asymmetry does not increase with moderate inbreeding in Drosophila melanogaster. Heredity 1994, 73, 373–376. [Google Scholar] [CrossRef] [PubMed]
- Gilligan, D.M.; Woodworth, L.M.; Montgomery, M.E.; Nurthen, R.K.; Briscoe, D.A.; Frankham, R. Can fluctuating asymmetry be used to detect inbreeding and loss of genetic diversity in endangered populations? Anim. Conserv. 2000, 3, 97–104. [Google Scholar] [CrossRef]
- Hosken, D.J.; Blanckenhorn, W.U.; Ward, P.I. Developmental stability in yellow dung flies (Scathophaga stercoraria): Fluctuating asymmetry, heterozygosity and environmental stress. J. Evol. Biol. 2000, 13, 919–926. [Google Scholar] [CrossRef]
- Keller, L.; Passera, L. Incest avoidance, fluctuating asymmetry, and the consequences of inbreeding in Iridomyrmex humilis, an ant with multiple queen colonies. Behav. Ecol. Sociobiol. 1993, 33, 191–199. [Google Scholar] [CrossRef]
- Kruuk, L.E.B.; Slate, J.; Pemberton, J.M.; Clutton-Brock, T.H. Fluctuating asymmetry in a secondary sexual trait: No associations with individual fitness, environmental stress or inbreeding, and no heritability. J. Evol. Biol. 2003, 16, 101–113. [Google Scholar] [CrossRef] [PubMed]
- Lens, L.; Van Dongen, S.; Galbusera, P.; Schenck, T.; Matthysen, E.; Van de Casteele, T. Developmental instability and inbreeding in natural bird populations exposed to different levels of habitat disturbance. J. Evol. Biol. 2000, 13, 889–896. [Google Scholar] [CrossRef]
- Ben-David, Y.; Hershkovitz, I.; Rubin, D.; Moscona, D.; Ring, B. Inbreeding effects on tooth size, eruption age, and dental directional and fluctuating asymmetry among South Sinai Bedouins. In Structure, Function and Evolution of Teeth; Smith, P., Tchernov, E., Eds.; Freund Publishing House, Ltd.: London, UK, 1992; pp. 361–389. [Google Scholar]
- Livshits, G.; Kobyliansky, E. Fluctuating asymmetry as a possible measure of developmental homeostasis in humans: A review. Hum. Biol. 1991, 63, 441–466. [Google Scholar] [PubMed]
- Markow, T.A.; Martin, J.F. Inbreeding and developmental stability in a small human population. Ann. Hum. Biol. 1993, 20, 389–394. [Google Scholar] [CrossRef] [PubMed]
- Özener, B. Effect of inbreeding depression on growth and fluctuating asymmetry in Turkish young males. Am. J. Hum. Biol. 2010, 22, 557–562. [Google Scholar] [CrossRef] [PubMed]
- Özener, B.; Graham, J.H. Growth and fluctuating asymmetry of human newborns: Influence of inbreeding and parental education. Am. J. Phys. Anthropol. 2014, 153, 45–51. [Google Scholar] [CrossRef] [PubMed]
- Schaefer, K.; Lauc, T.; Mitteroecker, P.; Gunz, P.; Bookstein, F.L. Dental arch asymmetry in an isolated Adriatic community. Am. J. Phys. Anthropol. 2006, 129, 132–142. [Google Scholar] [CrossRef] [PubMed]
- Graham, J.H.; Felley, J.D. Genomic coadaptation and developmental stability within introgressed populations of Enneacanthus gloriosus and E. obesus (Pisces, Centrarchidae). Evolution 1985, 39, 104–114. [Google Scholar]
- Lynch, M. The genetic interpretation of inbreeding depression and outbreeding depression. Evolution 1991, 45, 622–629. [Google Scholar] [CrossRef]
- Brown, A.F. Outbreeding depression as a cost of dispersal in the harpacticoid copepod, Tigriopus californicus. Biol. Bull. 1991, 181, 123–126. [Google Scholar] [CrossRef]
- Edmands, S. Heterosis and outbreeding depression in interpopulation crosses spanning a wide range of divergence. Evolution 1999, 1757–1768. [Google Scholar] [CrossRef]
- Burton, R.S. Hybrid breakdown in developmental time in the copepod Tigriopus californicus. Evolution 1990, 44, 1814–1822. [Google Scholar] [CrossRef]
- Emlen, J.M. Heterosis and outbreeding depression: A multi-locus model and an application to salmon production. Fish. Res. 1991, 12, 187–212. [Google Scholar] [CrossRef]
- Gharrett, A.J.; Smoker, W.W.; Reisenbichler, R.R.; Taylor, S.G. Outbreeding depression in hybrids between odd- and even-broodyear pink salmon. Aquaculture 1999, 173, 117–129. [Google Scholar] [CrossRef]
- Gilk, S.E.; Wang, I.A.; Hoover, C.L.; Smoker, W.W.; Taylor, S.G.; Gray, A.K.; Gharrett, A. Outbreeding depression in hybrids between spatially separated pink salmon, Oncorhynchus gorbuscha, populations: Marine survival, homing ability, and variability in family size. Environ. Biol. Fishes 2004, 69, 287–297. [Google Scholar] [CrossRef]
- Lewis, M.B. Why are mixed-race people perceived as more attractive? Perception 2010, 39, 136–138. [Google Scholar] [CrossRef] [PubMed]
- Little, A.C.; Hockings, K.J.; Apicella, C.L.; Sousa, C. Mixed-ethnicity face shape and attractiveness in humans. Perception 2012, 41, 1486–1496. [Google Scholar] [CrossRef] [PubMed]
- Hallgrimsson, B.; Willmore, K.; Hall, B.K. Canalization, developmental stability, and morphological integration in primate limbs. Yearb. Phys. Anthropol. 2002, 45, 131–158. [Google Scholar] [CrossRef]
- Manning, J.T.; Chamberlain, A.T. Fluctuating asymmetry, sexual selection and canine teeth in primates. Proc. R. Soc. Lond. B. Biol. Sci. 1993, 251, 83–87. [Google Scholar] [CrossRef] [PubMed]
- Manning, J.T.; Chamberlain, A.T. Fluctuating asymmetry in gorilla canines: A sensitive indicator of environmental stress. Proc. R. Soc. Lond. B. Biol. Sci. 1994, 255, 189–193. [Google Scholar] [CrossRef] [PubMed]
- Sarringhaus, L.A.; Stock, J.T.; Marchant, L.F.; McGrew, W.C. Bilateral asymmetry in the limb bones of the chimpanzee (Pan troglodytes). Am. J. Phys. Anthropol. 2005, 128, 840–845. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.; Harvati, K.; Hublin, J.-J.; Klingenberg, C.P. Morphological evolution through integration: A quantitative study of cranial integration in Homo, Pan, Gorilla and Pongo. J. Hum. Evol. 2012, 62, 155–164. [Google Scholar] [CrossRef] [PubMed]
- Willmore, K.E.; Klingenberg, C.P.; Hallgrímsson, B. The relationship between fluctuating asymmetry and environmental variance in rhesus macaque skulls. Evolution 2005, 59, 898–909. [Google Scholar] [CrossRef] [PubMed]
- Kegley, A.D.T.; Kuykendall, K.L. Dental asymmetry in South African australopithecines: A preliminary analysis of odontometric and morphological components. J. Hum. Evol. 2002, 42, A18–A19. [Google Scholar]
- Kegley, A.D.T.; Hemingway, J. Surveying stress and developmental stability in Homo habilis via fluctuating odontometric asymmetry: Assessing generic patterns and australopithecine differentiation. In Voyages in Science: Essays by South African Anatomists in Honour of Phillip V. Tobias’ 80th Birthday; Štrkalj, G., Pather, N., Kramer, B., Eds.; Content Solutions: Pretoria, South Africa, 2005; pp. 35–49. [Google Scholar]
- Frederick, M.J.; Gallup, G.G., Jr. Fluctuating dental asymmetry in great apes, fossil hominins, and modern humans: Implications for changing stressors during human evolution. Acta Psychol. Sin. 2007, 39, 489–494. [Google Scholar]
- Kieser, J.A.; Groeneveld, H.T. The assessment of fluctuating odontometric asymmetry from incomplete hominid fossil data. Anthropol. Anz. 1986, 44, 175–182. [Google Scholar] [PubMed]
- Baab, K.L.; McNulty, K.P. Size, shape, and asymmetry in fossil hominins: The status of the LB1 cranium based on 3D morphometric analyses. J. Hum. Evol. 2009, 57, 608–622. [Google Scholar] [CrossRef] [PubMed]
- Henneberg, M.; Eckhardt, R.B.; Chavanaves, S.; Hsü, K.J. Evolved developmental homeostasis disturbed in LB1 from Flores, Indonesia, denotes Down syndrome and not diagnostic traits of the invalid species Homo floresiensis. Proc. Natl. Acad. Sci. USA 2014, 111, 11967–11972. [Google Scholar] [CrossRef] [PubMed]
- Suarez, B.K. Neandertal dental asymmetry and the probable mutation effect. Am. J. Phys. Anthropol. 1974, 41, 411–416. [Google Scholar] [CrossRef] [PubMed]
- Baab, K.L.; Brown, P.; Falk, D.; Richtsmeier, J.T.; Hildebolt, C.F.; Smith, K.; Jungers, W. A critical evaluation of the Down syndrome diagnosis for LB1, type specimen of Homo floresiensis. PLoS ONE 2016, 11, e0155731. [Google Scholar] [CrossRef] [PubMed]
- Ríos, L.; Rosas, A.; Estalrrich, A.; García-Tabernero, A.; Bastir, M.; Huguet, R.; Pastor, F.; Sanchís-Gimeno, J.A.; de la Rasilla, M. Possible further evidence of low genetic diversity in the El Sidrón (Asturias, Spain) Neandertal group: Congenital clefts of the atlas. PLoS ONE 2015, 10, e0136550. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Quinto, F.; Lalueza-Fox, C. Almost 20 years of Neanderthal palaeogenetics: Adaptation, admixture, diversity, demography and extinction. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 2015, 370. [Google Scholar] [CrossRef] [PubMed]
- Doyle, W.J.; Johnston, O. On the meaning of increased fluctuating dental asymmetry: A cross populational study. Am. J. Phys. Anthropol. 1977, 46, 127–134. [Google Scholar] [CrossRef] [PubMed]
- Dibennardo, R.; Bailit, H.L. Stress and dental asymmetry in a population of Japanese children. Am. J. Phys. Anthropol. 1978, 48, 89–94. [Google Scholar] [CrossRef] [PubMed]
- Perzigian, A.J. Fluctuating dental asymmetry: Variation among skeletal populations. Am. J. Phys. Anthropol. 1977, 47, 81–88. [Google Scholar] [CrossRef] [PubMed]
- Palmer, A.R.; Strobeck, C. Fluctuating asymmetry and developmental stability: Heritability of observable variation vs. heritability of inferred cause. J. Evol. Biol. 1997, 10, 39–49. [Google Scholar] [CrossRef]
- Steele, J. Handedness in past human populations: Skeletal markers. Laterality 2000, 5, 193–220. [Google Scholar] [PubMed]
- Schiess, R.; Haeusler, M. No skeletal dysplasia in the Nariokotome boy KNM-WT 15000 (Homo erectus)—A reassessment of congenital pathologies of the vertebral column. Am. J. Phys. Anthropol. 2013, 150, 365–374. [Google Scholar] [CrossRef] [PubMed]
- Trinkaus, E.; Churchill, S.E.; Ruff, C.B. Postcranial robusticity in Homo. II: Humeral bilateral asymmetry and bone plasticity. Am. J. Phys. Anthropol. 1994, 93, 1–34. [Google Scholar] [CrossRef] [PubMed]
- Constandse-Westermann, T.S.; Newell, R.R. Limb lateralization and social stratification in western European Mesolithic societies. In People and Culture in Change; Hershkovitz, I., Ed.; British Archeological Reports International Series: Oxford, UK, 1989; pp. 405–433. [Google Scholar]
- Cohen, M.N.; Armelagos, G.J. Paleopathology at the Origins of Agriculture; University Press of Florida: Gainesville, FL, USA, 1984; p. 615. [Google Scholar]
- Hoover, K.C.; Matsumura, H. Temporal variation and interaction between nutritional and developmental instability in prehistoric Japanese populations. Am. J. Phys. Anthropol. 2008, 137, 469–478. [Google Scholar] [CrossRef] [PubMed]
- DeLeon, V.B. Fluctuating asymmetry and stress in a medieval Nubian population. Am. J. Phys. Anthropol. 2007, 132, 520–534. [Google Scholar] [CrossRef] [PubMed]
- Lieberman, D.E.; Devlin, M.J.; Pearson, O.M. Articular area responses to mechanical loading: Effects of exercise, age, and skeletal location. Am. J. Phys. Anthropol. 2001, 116, 266–277. [Google Scholar] [CrossRef] [PubMed]
- Pearson, O.M.; Lieberman, D.E. The aging of Wolff’s “law”: Ontogeny and responses to mechanical loading in cortical bone. Yearb. Phys. Anthropol. 2004, 47, 63–99. [Google Scholar] [CrossRef] [PubMed]
- Frost, H.M. Joint anatomy, design, and arthroses: Insights of the Utah paradigm. Anat. Record 1999, 255, 162–174. [Google Scholar] [CrossRef]
- Hamrick, M.W. A chondral modeling theory revisited. J. Theor. Biol. 1999, 201, 201–208. [Google Scholar] [CrossRef] [PubMed]
- Plochocki, J.H. Bilateral variation in limb articular surface dimensions. Am. J. Hum. Biol. 2004, 16, 328–333. [Google Scholar] [CrossRef] [PubMed]
- Ĉuk, T.; Leben-Seljak, P.; Štefanĉiĉ, M. Lateral asymmetry of human long bones. Var. Evol. 2001, 9, 19–32. [Google Scholar]
- Mays, S.; Steele, J.; Ford, M. Directional asymmetry in the human clavicle. Int. J. Osteoarchaeol. 1999, 9, 18–28. [Google Scholar] [CrossRef]
- Plochocki, J.H. Directional bilateral asymmetry in human sacral morphology. Int. J. Osteoarchaeol. 2002, 12, 349–355. [Google Scholar] [CrossRef]
- Whitlock, M. The heritability of fluctuating asymmetry and the genetic control of developmental stability. Proc. R. Soc. Lond. B. Biol. Sci. 1996, 263, 849–853. [Google Scholar] [CrossRef] [PubMed]
- Whitlock, M. The repeatability of fluctuating asymmetry: A revision and extension. Proc. R. Soc. Lond. B. Biol. Sci. 1998, 265, 1429–1431. [Google Scholar] [CrossRef]
- Tracy, M.; Freeman, D.C.; Duda, J.J.; Miglia, K.J.; Graham, J.H.; Hough, R.A. Developmental instability: An appropriate indicator of plant fitness components. In Developmental Instability: Causes and Consequences; Polak, M., Ed.; Oxford University Press: New York, NY, USA, 2003; pp. 196–212. [Google Scholar]
- Cowart, N.M.; Graham, J.H. Within- and among-individual variation in fluctuating asymmetry of leaves in the fig (Ficus carica L.). Int. J. Plant Sci. 1999, 160, 116–121. [Google Scholar] [CrossRef]
- Leung, B.; Forbes, M.R.; Houle, D. Fluctuating asymmetry as a bioindicator of stress: Comparing efficacy of analyses involving multiple traits. Am. Nat. 2000, 155, 101–115. [Google Scholar] [CrossRef] [PubMed]
- Thornhill, R.; Gangestad, S.W. Facial attractiveness. Trends Cogn. Sci. 1999, 3, 452–460. [Google Scholar] [CrossRef]
- Møller, A.P.; Thornhill, R. Bilateral symmetry and sexual selection: A meta-analysis. Am. Nat. 1998, 151, 174–192. [Google Scholar] [CrossRef] [PubMed]
- Palmer, A.R. Detecting publication bias in meta-analyses: A case study of fluctuating asymmetry and sexual selection. Am. Nat. 1999, 154, 220–233. [Google Scholar] [CrossRef]
- Thornhill, R.; Møller, A.P.; Gangestad, S.W. The biological significance of fluctuating asymmetry and sexual selection: A reply to Palmer. Am. Nat. 1999, 154, 234–241. [Google Scholar] [CrossRef]
- Van Dongen, S. Associations between asymmetry and human attractiveness: Possible direct effects of asymmetry and signatures of publication bias. Ann. Hum. Biol. 2011, 38, 317–323. [Google Scholar] [CrossRef] [PubMed]
- Van Dongen, S.; Gangestad, S.W. Human fluctuating asymmetry in relation to health and quality: A meta-analysis. Evol. Hum. Behav. 2011, 32, 380–398. [Google Scholar] [CrossRef]
- Van Dongen, S. Fluctuating asymmetry and masculinity/femininity in humans: A meta-analysis. Arch. Sex. Behav. 2012, 41, 1453–1460. [Google Scholar] [CrossRef] [PubMed]
- Sim, K. The relationship between sex-typical body shape and quality indicators. J. Soc. Evol. Cult. Psychol. 2013, 7, 97–120. [Google Scholar] [CrossRef]
- Van Dongen, S. Associations among facial masculinity, physical strength, fluctuating asymmetry and attractiveness in young men and women. Ann. Hum. Biol. 2014, 41, 205–213. [Google Scholar] [CrossRef] [PubMed]
- Hughes, S.M.; Harrison, M.A.; Gallup, G.G. The sound of symmetry: Voice as a marker of developmental instability. Evol. Hum. Behav. 2002, 23, 173–180. [Google Scholar] [CrossRef]
- Hughes, S.M.; Pastizzo, M.J.; Gallup, G.G., Jr. The sound of symmetry revisited: Subjective and objective analyses of voice. J. Nonverbal Behav. 2008, 32, 93–108. [Google Scholar] [CrossRef]
- Abend, P.; Pflüger, L.S.; Koppensteiner, M.; Coquerelle, M.; Grammer, K. The sound of female shape: A redundant signal of vocal and facial attractiveness. Evol. Hum. Behav. 2015, 36, 174–181. [Google Scholar] [CrossRef]
- Hill, A.K.; Cárdenas, R.A.; Wheatley, J.R.; Welling, L.L.M.; Burriss, R.P.; Claes, P.; Apicella, C.L.; McDaniel, M.A.; Little, A.C.; Shriver, M.D.; et al. Are there vocal cues to human developmental stability? Relationships between facial fluctuating asymmetry and voice attractiveness. Evol. Hum. Behav. 2016. [Google Scholar] [CrossRef]
- Jones, B.C.; Little, A.C.; Feinberg, D.R.; Penton-Voak, I.S.; Tiddeman, B.P.; Perrett, D.I. The relationship between shape symmetry and perceived skin condition in male facial attractiveness. Evol. Hum. Behav. 2004, 25, 24–30. [Google Scholar] [CrossRef]
- Weinberg, S.M.; Neiswanger, K.; Martin, R.A.; Mooney, M.P.; Kane, A.A.; Wenger, S.L.; Losee, J.; Deleyiannis, F.; Ma, L.; De Salamanca, J.E. The Pittsburgh Oral-Facial Cleft study: Expanding the cleft phenotype. Background and justification. Cleft Palate Cranio-Fac. J. 2006, 43, 7–20. [Google Scholar] [CrossRef] [PubMed]
- Neiswanger, K.; Cooper, M.E.; Weinberg, S.M.; Flodman, P.; Keglovits, A.B.; Liu, Y.; Hu, D.-N.; Melnick, M.; Spence, M.A.; Marazita, M.L. Cleft lip with or without cleft palate and dermatoglyphic asymmetry: Evaluation of a Chinese population. Orthod. Craniofac. Res. 2002, 5, 140–146. [Google Scholar] [CrossRef] [PubMed]
- Flores-Mondragón, G.; Paredes-Espinoza, M.A.; Hernández-Campos, N.A.; Sánchez-Chapul, L.; Paniagua-Pérez, R.; Martínez-Canseco, C.; Renán-León, S.; Araujo-Monsalvo, V.M.; Perea-Paz, J.M.; Flores-Jacinto, A. Facial anthropometry: A tool for quantitative evaluation in patients with peripheral facial paralysis. Int. J. Sci. Eng. Res. 2015, 6, 1657–1660. [Google Scholar]
- Garland, T.; Freeman, P.W. Selective breeding for high endurance running increases hindlimb symmetry. Evolution 2005, 59, 1851–1854. [Google Scholar] [CrossRef] [PubMed]
- Trivers, R.; Palestis, B.G.; Manning, J.T. The symmetry of children’s knees is linked to their adult sprinting speed and their willingness to sprint in a long-term Jamaican study. PLoS ONE 2013, 8, e72244. [Google Scholar] [CrossRef] [PubMed]
- Trivers, R.; Fink, B.; Russell, M.; McCarty, K.; James, B.; Palestis, B.G. Lower body symmetry and running performance in elite Jamaican track and field athletes. PLoS ONE 2014, 9, e113106. [Google Scholar] [CrossRef] [PubMed]
- Oxford, M. The Effect of Muscle Imbalance on Running Performance in Collegiate Level Athletes. Master’s Thesis, University of Windsor, Windsor, ON, Canada, 2013. [Google Scholar]
- Al-Eisa, E.; Egan, D.; Wassersug, R. Fluctuating asymmetry and low back pain. Evol. Hum. Behav. 2004, 25, 31–37. [Google Scholar] [CrossRef]
- Goldberg, C.J.; Fogarty, E.E.; Moore, D.P.; Dowling, F.E. Fluctuating asymmetry and vertebral malformation: A study of palmar dermatoglyphics in congenital spinal deformities. Spine 1997, 22, 775–779. [Google Scholar] [CrossRef] [PubMed]
- Mittal, V.A.; Dean, D.J.; Pelletier, A. Dermatoglyphic asymmetries and fronto-striatal dysfunction in young adults reporting non-clinical psychosis. Acta Psychiatr. Scand. 2012, 126, 290–297. [Google Scholar] [CrossRef] [PubMed]
- Stiles, J.; Jernigan, T.L. The basics of brain development. Neuropsychol. Rev. 2010, 20, 327–348. [Google Scholar] [CrossRef] [PubMed]
- Kavanagh, D.H.; Tansey, K.E.; O’Donovan, M.C.; Owen, M.J. Schizophrenia genetics: Emerging themes for a complex disorder. Mol. Psychiatry 2015, 20, 72–76. [Google Scholar] [CrossRef] [PubMed]
- Reilly, J.L.; Murphy, P.T.; Byrne, M.; Larkin, C.; Gill, M.; O’Callaghan, E.; Lane, A. Dermatoglyphic fluctuating asymmetry and atypical handedness in schizophrenia. Schizophr. Res. 2001, 50, 159–168. [Google Scholar] [CrossRef]
- Cohen, J. Statistical Power Analysis for the Behavior Sciences; Lawrance Eribaum Association: Hillsdale, NJ, USA, 1988; p. 487. [Google Scholar]
- Abrahams, B.S.; Geschwind, D.H. Connecting genes to brain in the autism spectrum disorders. Arch. Neurol. 2010, 67, 395–399. [Google Scholar] [CrossRef] [PubMed]
- Geschwind, D.H. Genetics of autism spectrum disorders. Trends Cognit. Sci. 2011, 15, 409–416. [Google Scholar] [CrossRef] [PubMed]
- Bates, T.C. Fluctuating asymmetry and intelligence. Intelligence 2007, 35, 41–46. [Google Scholar] [CrossRef]
- Furlow, F.B.; Armijo–Prewitt, T.; Gangestad, S.W.; Thornhill, R. Fluctuating asymmetry and psychometric intelligence. Proc. R. Soc. Lond. B. Biol. Sci. 1997, 264, 823–829. [Google Scholar] [CrossRef] [PubMed]
- Prokosch, M.D.; Yeo, R.A.; Miller, G.F. Intelligence tests with higher g-loadings show higher correlations with body symmetry: Evidence for a general fitness factor mediated by developmental stability. Intelligence 2005, 33, 203–213. [Google Scholar] [CrossRef]
- Banks, G.C.; Batchelor, J.H.; McDaniel, M.A. Smarter people are (a bit) more symmetrical: A meta-analysis of the relationship between intelligence and fluctuating asymmetry. Intelligence 2010, 38, 393–401. [Google Scholar] [CrossRef]
- Yeo, R.A.; Gangestad, S.W.; Daniel, W.F. Hand preference and developmental instability. Psychobiology 1993, 21, 161–168. [Google Scholar]
- Somers, M.; Ophoff, R.A.; Aukes, M.F.; Cantor, R.M.; Boks, M.P.; Dauwan, M.; de Visser, K.L.; Kahn, R.S.; Sommer, I.E. Linkage analysis in a Dutch population isolate shows no major gene for left-handedness or atypical language lateralization. J. Neurosci. 2015, 35, 8730–8736. [Google Scholar] [CrossRef] [PubMed]
- Benderlioglu, Z.; Sciulli, P.W.; Nelson, R.J. Fluctuating asymmetry predicts human reactive aggression. Am. J. Hum. Biol. 2004, 16, 458–469. [Google Scholar] [CrossRef] [PubMed]
- Furlow, B.; Gangestad, S.W.; Armijo-Prewitt, T. Developmental stability and human violence. Proc. R. Soc. Lond. B. Biol. Sci. 1998, 265, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Özener, B.; Atamtürk, D.; Duyar, İ. Self-reported aggressiveness is not related to developmental instability. Am. J. Hum. Biol. 2015, 27, 867–870. [Google Scholar] [CrossRef] [PubMed]
- Özener, B.; Pelin, C.; Kürkçüoğlu, A.; Zağyapan, R. Facial symmetry is not a reliable cue of aggressive behavior. Eurasian J. Anthropol. 2012, 3, 32–40. [Google Scholar]
- Manning, J.T.; Wood, D. Fluctuating asymmetry and aggression in boys. Hum. Nat. 1998, 9, 53–65. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-Reyes, J.A.; Gil-Burmann, C.; Fink, B.; Turiegano, E. Facial asymmetry and aggression in Spanish adolescents. Personal. Individ. Differ. 2012, 53, 857–861. [Google Scholar] [CrossRef]
- Holtzman, N.S.; Augustine, A.A.; Senne, A.L. Are pro-social or socially aversive people more physically symmetrical? Symmetry in relation to over 200 personality variables. J. Res. Personal. 2011, 45, 687–691. [Google Scholar] [CrossRef]
- Nuzzo, R. Statistical errors. Nature 2014, 506, 150–152. [Google Scholar] [CrossRef] [PubMed]
- Head, M.L.; Holman, L.; Lanfear, R.; Kahn, A.T.; Jennions, M.D. The extent and consequences of p-hacking in science. PLoS Biol. 2015, 13, e1002106. [Google Scholar] [CrossRef] [PubMed]
- Hurlbert, S.H.; Lombardi, C.M. Final collapse of the Neyman-Pearson decision theoretic framework and rise of the neoFisherian. Ann. Zool. Fenn. 2009, 46, 311–349. [Google Scholar] [CrossRef]
- Lombardi, C.M.; Hurlbert, S.H. Misprescription and misuse of one-tailed tests. Austral Ecol. 2009, 34, 447–468. [Google Scholar] [CrossRef]
- Kozlov, M.V.; Zvereva, E.L. Confirmation bias in studies of fluctuating asymmetry. Ecol. Indic. 2015, 57, 293–297. [Google Scholar] [CrossRef]
Figure 1.
Relative indices of individual asymmetry can generate negative size scaling when the trait, dermatoglyphic ridge counts in this case, exhibits an additive, not multiplicative, error. Data is from Holt’s [45]—50 male and 50 female parents—Table 1 (left and right-hand ridge counts of 50 families). (a) Absolute asymmetry of finger ridge count regressed on total ridge count (F1, 98 = 0.571, p > 0.45); (b) Relative absolute asymmetry of finger ridge count regressed on total ridge count (F1, 98 = 41.023, p < 0.00001). Filled circles represent individual parents.
Figure 1.
Relative indices of individual asymmetry can generate negative size scaling when the trait, dermatoglyphic ridge counts in this case, exhibits an additive, not multiplicative, error. Data is from Holt’s [45]—50 male and 50 female parents—Table 1 (left and right-hand ridge counts of 50 families). (a) Absolute asymmetry of finger ridge count regressed on total ridge count (F1, 98 = 0.571, p > 0.45); (b) Relative absolute asymmetry of finger ridge count regressed on total ridge count (F1, 98 = 41.023, p < 0.00001). Filled circles represent individual parents.
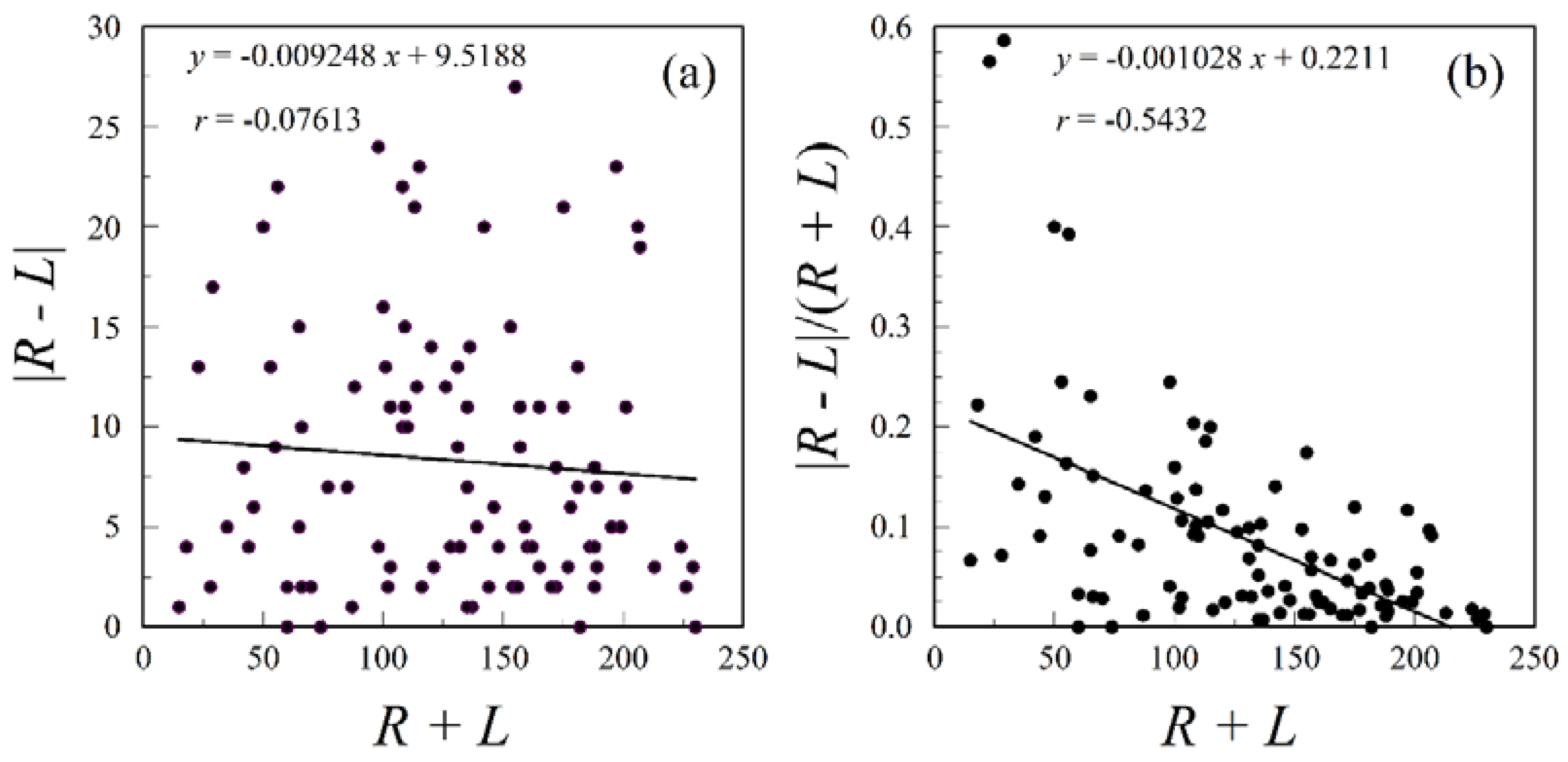
Figure 2.
Regressions of |R − L| on R + L for mesiodistal and buccolingual tooth diameters (mm) of M1, M2, P3, and C from the upper jaws of individuals from Ban Chiang, Thailand. Points represent individuals in the sample of upper jaws. Females are filled circles; males are filled squares; unidentified sexes are diamonds. Data from Database E.4 [67].
Figure 2.
Regressions of |R − L| on R + L for mesiodistal and buccolingual tooth diameters (mm) of M1, M2, P3, and C from the upper jaws of individuals from Ban Chiang, Thailand. Points represent individuals in the sample of upper jaws. Females are filled circles; males are filled squares; unidentified sexes are diamonds. Data from Database E.4 [67].
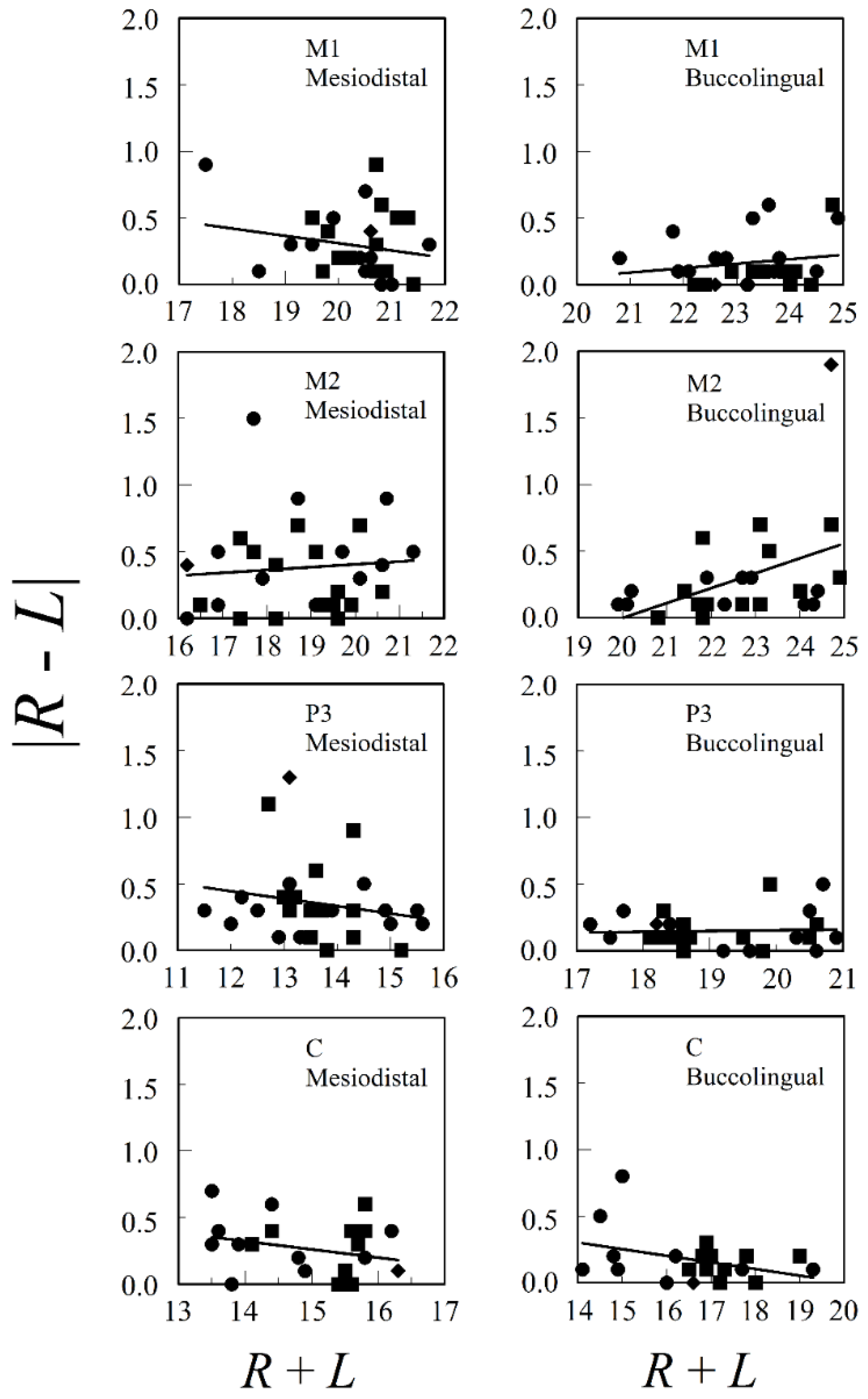
Figure 3.
Dental fluctuating asymmetry in hominids and humans (from Frederick and Gallup [220]). Mean, 95% confidence interval, and sample size.
Figure 3.
Dental fluctuating asymmetry in hominids and humans (from Frederick and Gallup [220]). Mean, 95% confidence interval, and sample size.
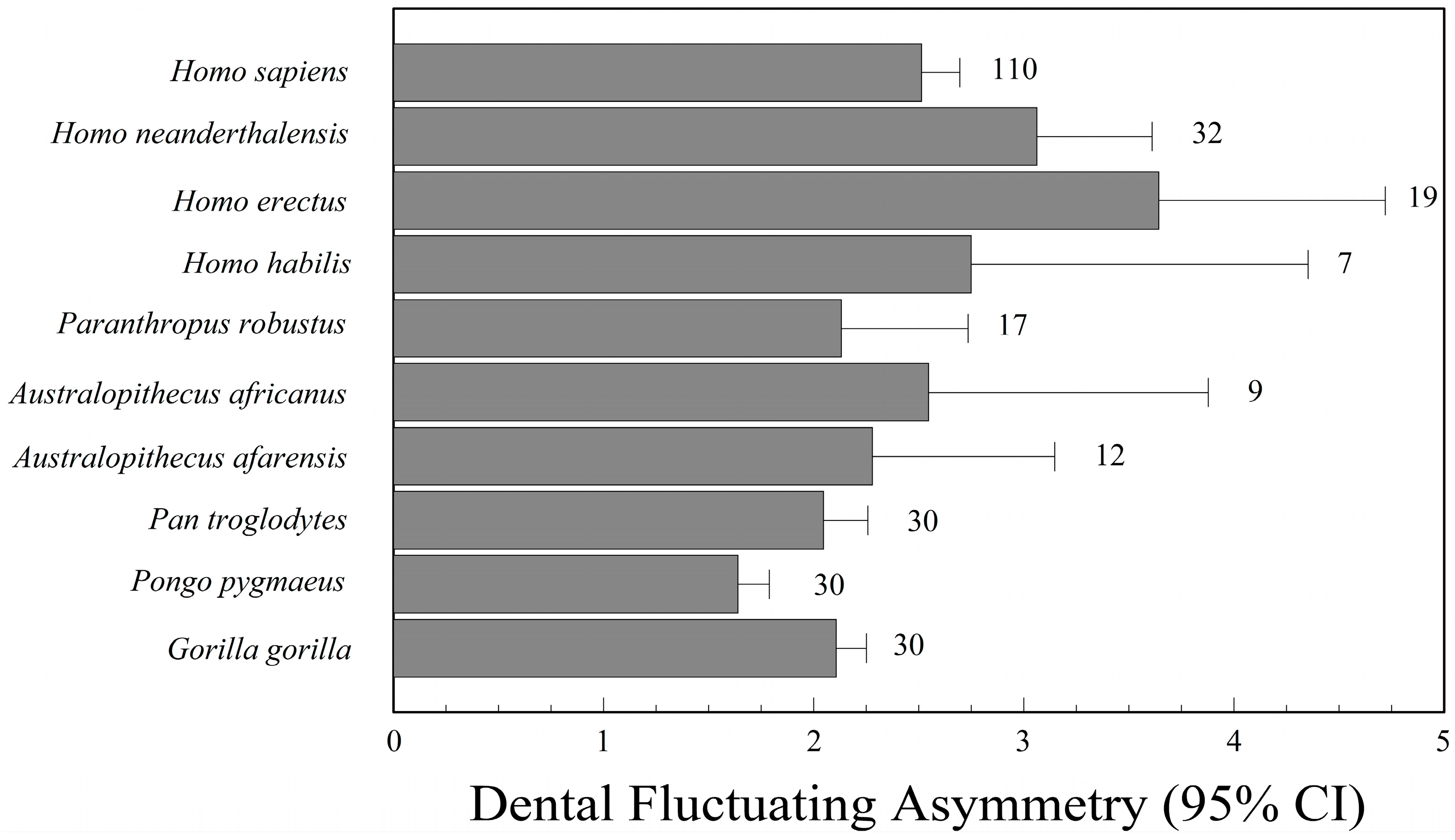
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Graham, J.H.; Özener, B. Fluctuating Asymmetry of Human Populations: A Review. Symmetry 2016, 8, 154. https://doi.org/10.3390/sym8120154
AMA Style
Graham JH, Özener B. Fluctuating Asymmetry of Human Populations: A Review. Symmetry. 2016; 8(12):154. https://doi.org/10.3390/sym8120154
Chicago/Turabian StyleGraham, John H., and Bariş Özener. 2016. "Fluctuating Asymmetry of Human Populations: A Review" Symmetry 8, no. 12: 154. https://doi.org/10.3390/sym8120154
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.