
Early- and Late-Light Embryonic Stimulation Modulates Similarly Chicks’ Ability to Filter out Distractors


1
Department of Life Sciences, University of Trieste, I-34127 Trieste, Italy
2
Institute of applied biology (IBFA), University of Caen, 14000 Caen, France
3
Center for Mind/Brain Sciences, University of Trento, I-38068 Rovereto, Italy
*
Author to whom correspondence should be addressed.
Symmetry 2017, 9(6), 84; https://doi.org/10.3390/sym9060084
Submission received: 24 March 2017
/
Revised: 2 June 2017
/
Accepted: 6 June 2017
/
Published: 8 June 2017
(This article belongs to the Special Issue Brain Asymmetry of Structure and/or Function)
{kind=link}
{kind=link}
Abstract
:Chicks (Gallus gallus) learned to run from a starting box to a target located at the end of a runway. At test, colourful and bright distractors were placed just outside the starting box. Dark incubated chicks (maintained in darkness from fertilization to hatching) stopped significantly more often, assessing more the left-side distractor than chicks hatched after late (for 42 h during the last three days before hatching) or early (for 42 h after fertilization) exposure to light. The results show that early embryonic light stimulation can modulate this particular behavioural lateralization comparably to the late application of it, though via a different route.
Keywords:
attention; functional lateralization; cerebral lateralization; embryo; light; fish; chicks; birds1. Introduction
It is now well established that environmental light stimulation interplays with a genetic cascade of events in promoting brain specialization in two different classes of vertebrates, fish and birds (reviews in [1,2,3]).
A complex chain of developmental steps leads to brain lateralization in zebrafish starting with an asymmetrical expression of a gene network that controls the development of structural left-right differences within the epithalamus, including asymmetric parapineal migration [4,5,6,7]. As a secondary consequence, in the transparent eggs of the zebrafish an early action of light prompts functional brain asymmetries including motor and sensory processing. Fries hatched from eggs exposed to the photic input during the first week after fertilization prefer to attend to conspecifics with the left eye, whereas fries whose embryonic development happened in darkness do not display the same asymmetry [8]. If the light fails to reach the embryos in two distinct moments within the first week post-fertilization, the normal development of some lateralized behaviours is either compromised or prevented. For instance, darkness during the first day results in an inversion of the reaction to a dummy predator: after normal light regimes, zebrafish avoid the predator appearing on the left side, whereas after darkness they respond more intensely to a predator coming from the right side [9]; in contrast, darkness during the third day prevents the appearance of any asymmetric response to the predator [10]. However, the role of light stimulation on brain structural asymmetry has not been conclusively clarified in zebrafish, as no effect of light has been shown on the asymmetry of molecular markers [11], and some behaviours are lateralized while some others are not, independently of the neuroanatomical asymmetries [12].
Embryonic light application influences also the neurodevelopment of cerebral lateralization in the avian brain, but via a different pathway, i.e., by the asymmetric stimulation of one eye. For more than three decades, it has been repeatedly shown that an asymmetrical embryonic positioning before hatching (due to unilateral expression of Nodal signals responsible for the body torsion [13]) allows light penetration of the egg during the final days of incubation to act selectively on one side of the chick’s head and to trigger anatomical and functional brain asymmetries via right eye stimulation [14]. In response to the asymmetric light input to the retinal cells, brain regions in the left hemisphere fed by the right eye develop earlier than their counterparts in the right hemisphere and a higher number of fibers crosses from the left side of the thalamus to the right hemisphere via the supraoptic decussation [15,16,17]. Such structural asymmetry is functionally detectable in several visually-guided behaviours, such as the advantage of the right eye in preventing pecks to not edible elements when searching for food [18]. Furthermore, chicks presenting such an asymmetry outperform chicks hatched in darkness in dual tasks [19] or when they have to combine different kinds of information to master a correct discrimination [20]. The multifaceted role of light is apparent in the fact that light exposure affects not only abilities related to the stimulated right eye but also functions of the left eye related to attack, copulation, predator detection [15], and visuospatial abilities [21]. Moreover, reversing the eye exposed to light by untwisting the embryo’s head and applying a patch to the right eye causes the pattern of asymmetries to be inverted [15,22]. Note, however, that although light exerts such an important role in the establishment of lateralization, some forms of asymmetries as those associated with unilateral eye used during sleep [23] or with the neural mechanisms of social recognition and imprinting [24,25,26] develop even if the incubation process takes place in darkness [27,28].
The depicted scenario shows a composite set of mechanisms at play in the development of brain asymmetries and the common thread to birds and fish seems to indicate that, following two different anatomical routes, light moulds a similar functional cerebral specialization in the two taxa [29]. Broadly speaking, the right hemisphere orchestrates a form of primitive avoidance and wariness while the left-hemisphere complements brain specialization with the control of routine behaviours of feeding and analysis in familiar contexts, counteracting distraction and irrelevant response to novelty credited to the right hemisphere [1,30]. Thus, apparently similar behavioural asymmetries can be generated by different neural asymmetric systems [31,32].
To check whether a different critical period for the application of the light input could be part of the asymmetric neurodevelopment of cerebral functions also in the chick, Chiandetti et al. [33] exposed eggs to light for a brief period after fertilization, when other photosensitive regions are developing but no retinal photoreceptors have been differentiated yet [34,35], and thereafter maintained in the dark. Chicks hatched under this condition performed in a comparable fashion to chicks hatched from eggs light-stimulated in the canonical time-window, i.e., during the last three days of incubation [33]. The testing condition required chicks to avoid an obstacle placed midway between the starting box and the target at the other end. In such a situation, the two light-stimulated groups showed no preference to detour the obstacle by circumventing it well as much from the left as the right side. By contrast, chicks hatched in darkness showed a pronounced bias to detour the obstacle systematically on the left side. In that case, a motoric difference between stimulated and unstimulated individuals was insufficient to explain the pattern of behaviour observed because all the chicks showed the same motor bias to run slightly toward the left side of the environment when tested without any obstacle on their way to the target (for the specificity of light effect, see reviews in [2,31]). Rather, unstimulated chicks’ selective bias emerged only when an obstacle was on the way to the target. It is possible that, when freely running, chicks previously kept in darkness are less able to sustain attention toward the target and need the right eye to view the obstacle in order to keep track of it while running and avoiding the obstacle toward the target.
Here, in the attempt to widen the comprehension of when and how light stimulation is effective in shaping visually-driven asymmetric responses, we assessed stimulated (early and lately) and unstimulated chicks and compared the performance of the three groups of animals in a further testing situation. Briefly, two days old domestic chicks (Gallus gallus) first learned to run from one end of a runway to a target located at the opposite end; then, at day 5 of age, colourful and bright distractors were placed close to the starting area and we scored whether the chicks pecked at the distractor, how many times and whether preferentially on the one positioned on the left, assuming that the novelty would have engaged mainly the left eye (right hemisphere). The task was chosen as a replication of a previous one with a change in the type of distractor that could provide an incremental knowledge about the observed phenomenon.
2. Materials and Methods
The study was carried out in compliance with the European Community and the Italian law on animal experiments by the Ministry of Health, under the authorization of the Ethical Committee of the University of Trieste (protocol number 385 pos II/9 dd 16.03.2012).
2.1. Subjects
Chicks of the Ross 308 (Aviagen) broiler strain hatched in our laboratory under controlled conditions. The eggs were collected from a local commercial hatchery immediately after fertilization and, thereafter, kept in a FIEM snc, MG 100 H incubator under controlled temperature (37.7 °C) and humidity (about 50–60%) conditions, in a darkened room so that no further incidental light could reach the eggs. Fifty eggs were incubated in complete darkness from the arrival to the laboratory and until the hatching day (Di-chicks = 38); fifty eggs were exposed to light from their arrival to the lab and for 42 h and thereafter remained in the dark (EarlyLi-chicks = 39); fifty eggs were maintained in darkness and exposed to light from day 18 and for 42 h before hatching (LateLi-chicks = 36). A 60 W incandescent light bulb or 15 LEDs (18 lumens per LED) provided homogeneous light of about 250 lux within the incubator. As reviewed in [2], high intensity and prolonged exposure to light can exert various effects (from hatchability to interlimb coordination) and this applies to LEDs too; however, the light regime adopted in our protocol is not proven to have comparable side-effects (and see [33] for an analysis of the identical running trajectories in Di-, EarlyLi-, and LateLi-chicks). Immediately after hatching, each chick was reared singly in a metal home-cage (28 cm wide × 32 cm high × 40 cm deep) illuminated by LED (12 L: 12 D cycle) and located in a separate room at 30 °C. Food and water were available ad libitum.
2.2. Apparatus
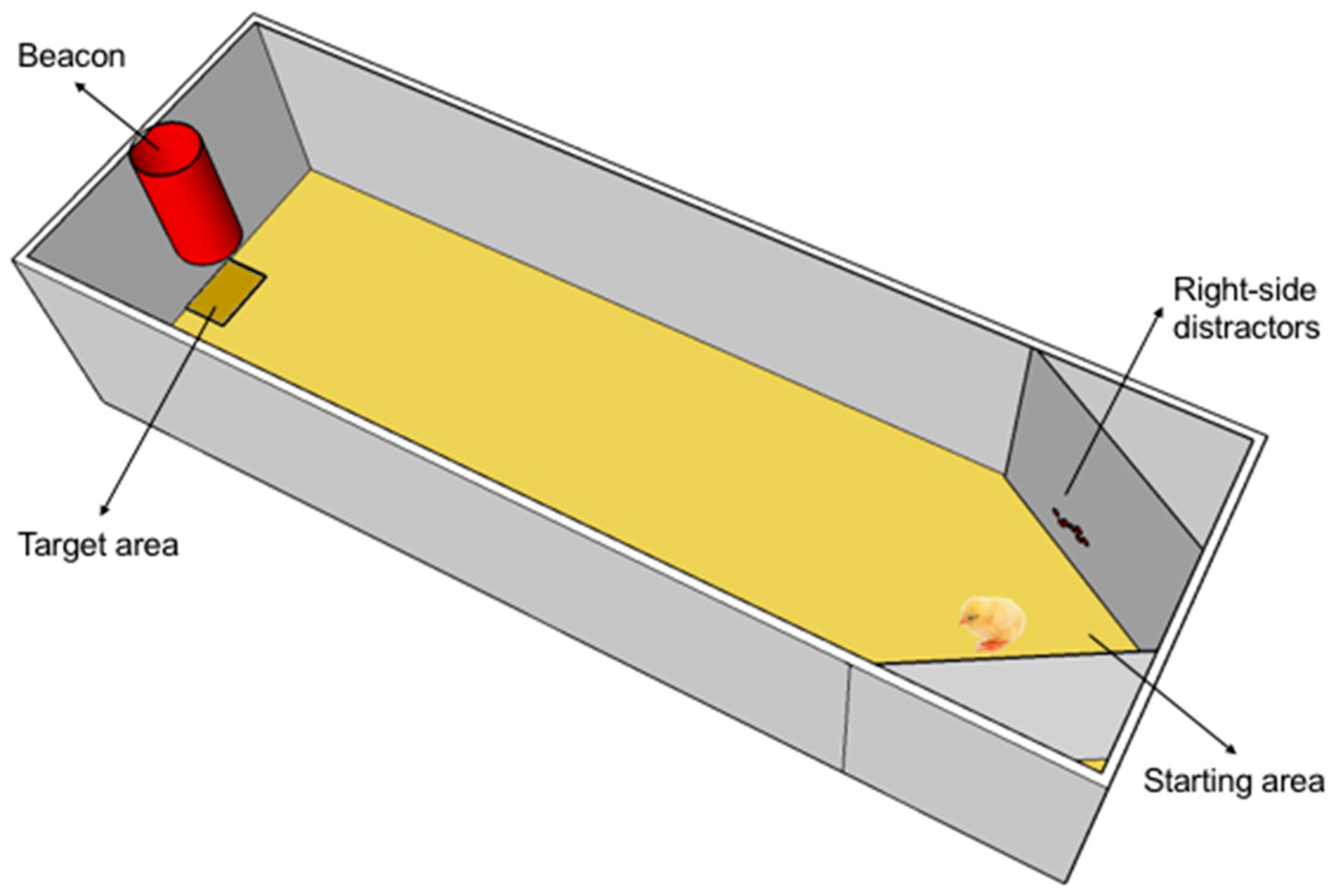
A white rectangular enclosure (40 cm wide × 50 cm high × 160 cm deep) with sawdust (5 cm in depth) on the floor served as training apparatus. A red conspicuous plastic beacon was placed at the middle of the smaller end of the apparatus and 7 cm above the floor, indicating the presence of a plastic feeder (target) exactly below it. Two lamps of 50 W centered on the top of the smaller ends provided uniform illumination to the apparatus. For the testing, two slanted walls were adjusted close to the starting point on both the left and the right (see Figure 1) and decorated with salient shiny beads placed at about chick’s head height, functioning as distractors.
In order to keep track of the chick’s movements within the apparatus, a black removable sticky paper was temporarily attached on the chick’s back. The behaviour was videorecorded from above and scored offline by an independent observer blind with respect to the hatching conditions of the animals.
2.3. Procedure
On day 2 of age, after 3 h of food deprivation, each chick was first accustomed to the training apparatus by letting it free to explore the environment for about 30 min and reach the target where some mealworm larvae (Tenebrio molitor) were placed. The next two days, each chick was placed within the apparatus at the opposite end in front of the target and left free to run toward the feeder. This procedure was repeated 20 times (10 times per day).
On day 5, each chick was given one trial as used during training to reinstate motivation and immediately after it was tested only once with the distractors. In the single testing trial, no mealworm was available under the target. This procedure was chosen to rule out any potential influence of the presence of the reward. The trial ended as soon as the chick reached the feeder. The positions of the starting point and the target were counterbalanced between subjects in order to control for any undesired asymmetry within the environment. As a dependent measure, we scored the number of pecks directed at right and left distractors.
3. Results
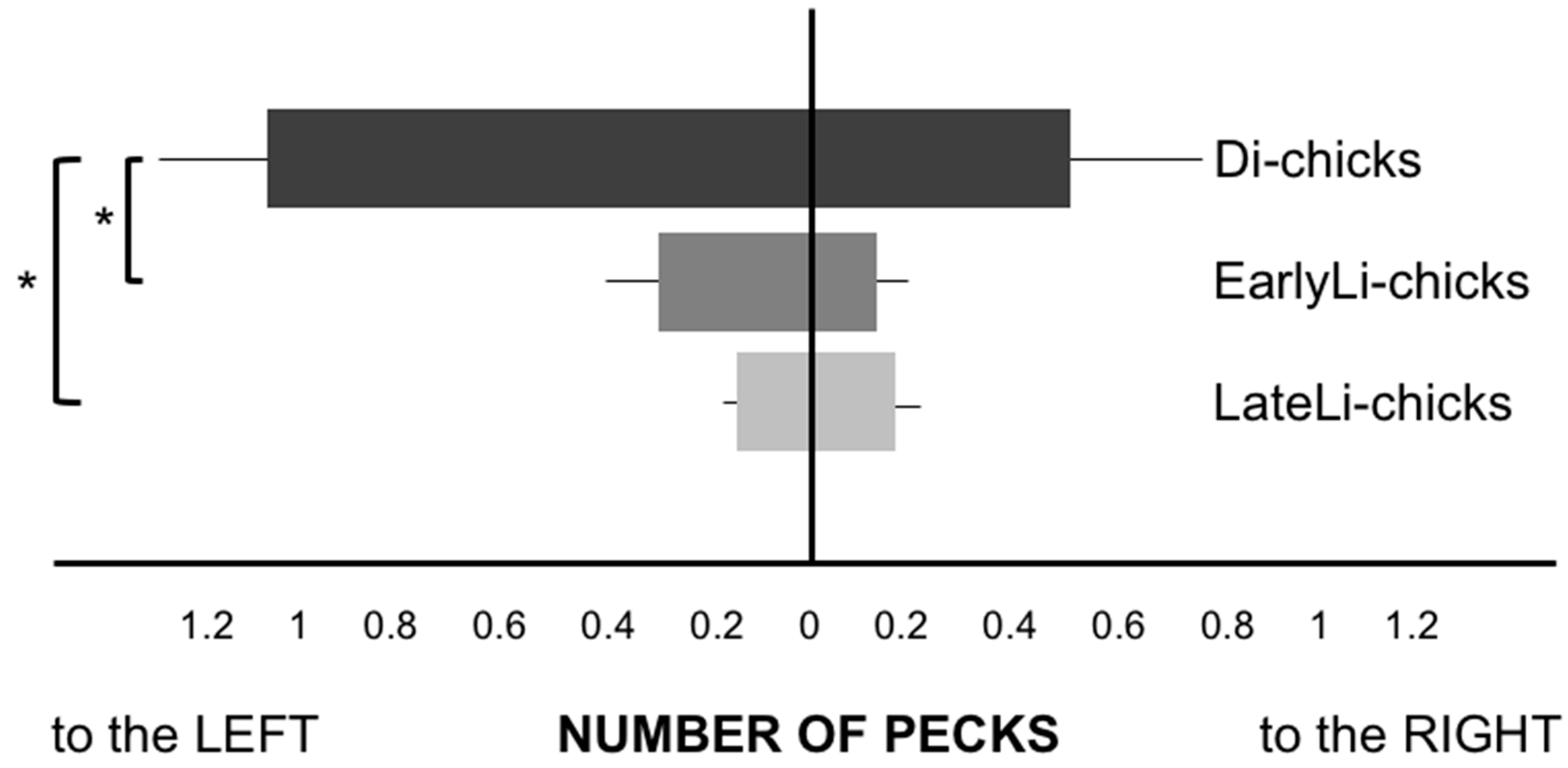
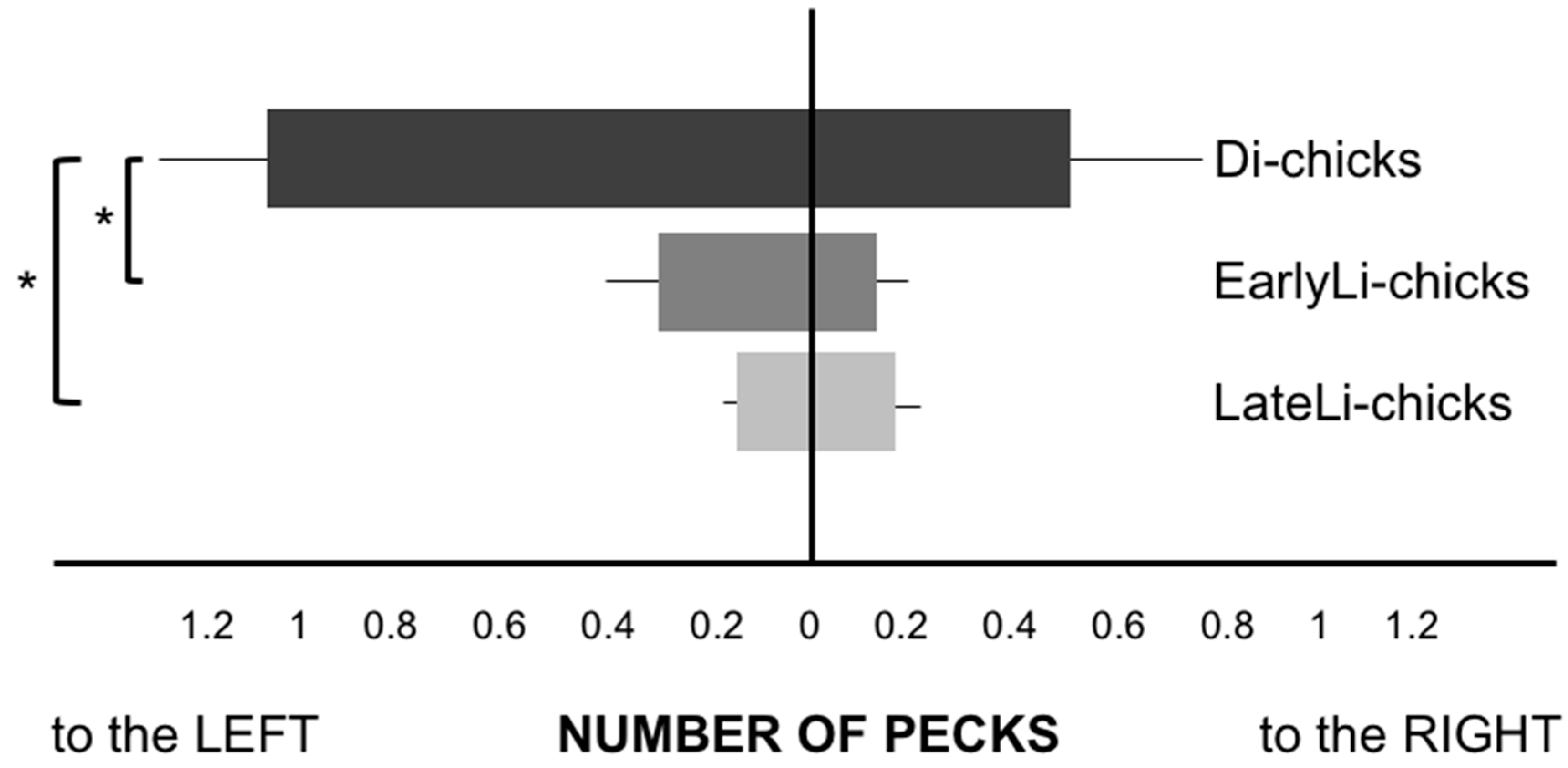
After having verified that the assumption of homogeneity of variances was not satisfied with the Levene’s test, we ran the non-parametric test Kruskal-Wallis on the overall number of pecks, which showed a significant difference between the three hatching groups (χ2(2) = 10.194, p = 0.006): Di-chicks were more distracted than the two stimulated groups on the pecks toward the left distractor (χ2(2) = 9.352, p = 0.009), but not to the right distractor (χ2(2) = 2.844, p = 0.241), as visible in Figure 2. Di-chicks pecked more at the distractor placed on the left side than both EarlyLi- (Z = −2.271, p = 0.023) and LateLi-chicks (Z = −2.588, p = 0.010), whereas no difference emerged between the two light-stimulated groups (Z = −0.368, p = 0.713, Mann-Whitney Post Hoc test).
4. Discussion
In this investigation of the time windows in which embryonic light stimulation affects the development of functional brain asymmetries, we replicated previous findings showing that chicks hatched from eggs exposed to environmental illumination for 42 h, at either an early or a late stage of embryonic development, display a comparable behaviour. Specifically, both EarlyLi- and LateLi-chicks were not distracted by the novel elements placed in proximity of the starting area. Conversely, Di-chicks, hatched from eggs maintained for the whole developmental period in complete darkness, were significantly attracted by the novel elements and could not restrain from pecking at these items before reaching the target, and especially at those placed on their left side. Note that the procedure used here matches the one used in our previous work [33], where we showed a specific effect of light stimulation on hemispheres’ functionality with no detriment of dark incubation condition on a typical motor and cognitive development (see also [36] for comparable results on pigeons).
The observed pattern of chicks’ performance confirms that embryonic application of light stimulation modulates the ability to sustain attention. EarlyLi- and LateLi-chicks ignored the novel elements and focused on the target, directly approaching it in a routine-like behaviour as learned during the familiarization trials without distractors. Both the left and the right eyes seemed equally good in targeting the goal and avoiding the salient distractor elements presented at test. By contrast, Di-chicks were strongly biased toward the distractors. The fact that the distractors placed on the left side resulted more attractive than those placed on the right side, uncovers the brain asymmetry at play in Di-chicks: the right eye is engaged in sustaining attention to the target, while the left eye mediates attention deployment to the novel and salient elements located on the left side. Our findings suggest that in Di-chicks the separation of the two hemispheres also maintains segregated the processing of the target and the distractor. While the left hemisphere would control the routine running behaviour toward the target, the right hemisphere is engaged by novelty and the chicks stop their running to assess the distractor located on the left, as they do when they monitor the predator [19,37].
In LateLi-chicks, instead, it appears that the right hemispheric involvement in response to novelty is modulated by the asymmetric embryonic light stimulation. The cross-talk between the two halves of the brain makes the left hemisphere capable of inhibitory control over the compulsory attention directed toward the novelty, similarly to the testing situation in which the right eye (left hemisphere) inhibits peck at irrelevant elements spread among grains [19].
What remains to be understood is how light induces a comparable performance in LateLi- and EarlyLi-chicks, considering that only on LateLi-chicks light acts asymmetrically on the fully-formed eye. In zebrafish, the involvement of both habenulae in the control of behaviour is shown by the fact that selective inactivation of these nuclei induces a persistent freezing response [38]; furthermore, the use of the right eye to target the food implies that the left hemisphere is engaged in sustained control, with the enrollment of the left lateral habenulae, reducing the probability of being distracted [39,40]. Despite the fact that there is no clear evidence that the role of light on the lateralized behaviour depends directly on a stimulation of the parapineal, this could account for the performance observed in both fish and EarlyLi-chicks. This hypothesis requires further investigation: indeed, in fish, other photosensitive areas than the eye participate in determining lateralization [41,42] and one may wonder whether the involvement of the same regions could be extended to explain chicks’ performance, since analogous cells are developing in the chick embryo at the early stages when we applied the light stimulation [43]. Due to the common differentiation of the diencephalic areas in birds and fish, the involvement of the ephyphysis-habenula axis could be the target for a further window in which light may be operating in chicks as well. A further complication may derive from the fact that the habenular nuclei are asymmetric in several species [44,45] and hence might be differentially stimulated by the action of light. By contrast, in birds the habenulae are assumed to be symmetric, despite one study on chicks showed that there can be individual asymmetries and males tend to present a larger right medial habenula [46]; unfortunately, there was no mention of the incubation condition in this study and hence whether it applies to our results or not is open to speculation. On the basis of atlases of different avian species, it appears that the pineal gland, projecting to the habenulae, is larger in absolute size in chicks than, for instance, in pigeons. The cytochemical characterization of the avian pineal organ demonstrates many structural, functional and biochemical analogies between the retinal and the pineal photoreceptors [47,48]. Furthermore, other brain regions involved in lateralization may have been simply overlooked in previous histological assessments of light stimulation effects. There might also be a further extra-retinal photoreceptive candidate in birds outside the pineal gland. As shown in quails, in the avian ventral thalamus and septal region there are so-called deep photoreceptors that seem to participate in the regulation of seasonal cycles of reproduction [49,50]. These further photosensitive receptors respond to light in the quail and might be activated in domestic chicks as well. Certainly, this hypothesis paves the way to further investigations addressing specifically the neural substrates enrolled by light at precocious stages of the chick’s embryonic development.
5. Conclusions
Here we documented that an early application of light during incubation modulates a particular functional asymmetry in chicks in a similar way to the well-known late stimulation. Light seems to operate on a genetically determined asymmetry by mediating a better cooperation between the two hemispheres. The asymmetrical light stimulation experience does not simply affect hemispheric specialization (like a left-hemispheric dominance of visuomotor control (discussed for instance in [32])) but also how efficiently the hemispheres can interact or cooperate [36,51].
The mechanisms responsible for the early modulation remain to be investigated, however a broader consideration on the effects of light stimulation before birth is worth discussing. Despite it is controversial whether the human foetus is reached by asymmetric light to one eye (it is attested that 2/3 of the embryos are rotated with the right eye toward the external abdominal wall in the latest stages of gestation), in principle the light reaches the intrauterine environment [52]. Indeed, at about 36–40 weeks, the foetus responds to flashes of light to the maternal abdomen with an increment in cardiac frequency and eye and body movements [53]. There are also indications that light can affect the development of cerebral lateralization in human foetuses by modulating the available hormonal levels [54]. Despite assessing this hypothesis has proven to be very difficult, seasonal anisotropy has been recently shown with respect to the distribution of handedness as related to gender: longer photoperiods experienced during the first 14–18 weeks are associated with left-handed males [55]. Hence, at present, a hormonal modulation cannot be ruled out.
Asymmetries induced by genetic factors are shaped by environmental illumination, as indicated by previous results on zebrafish and chicks, but here we showed that in an avian species this takes place in two different time-windows. If light entails two different processes in the two time-windows, a more sensitive test could reveal a specific involvement of each hemisphere. For instance, an investigation of the performance under monocular testing condition [56] could reasonably refine the enrolment of each hemisphere depending on the specific genetic-environment route, since monocular and binocular performances could differ profoundly (e.g., [57]).
Acknowledgments
Thanks are extended to Margherita Lucadello and Eliana Boschetti for supporting data collection and off-line scoring, and to the anonymous reviewers for the thoroughness with which they evaluated our manuscript. C.C. was partially funded by a UniTs-FRA2015 grant.
Author Contributions
C.C. and G.V. conceived the experiment; C.C., B.L. and E.V. designed the procedure; B.L. and C.C. performed the experiments; C.C. analyzed the data and prepared the figures; C.C. drafted the paper; C.C., G.V., E.V. and B.L. revised and approved the final version of the paper.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Rogers, L.J.; Vallortigara, G.; Andrew, R.J. Divided Brains: The Biology and Behaviour of Brain Asymmetries; Cambridge University Press: Cambridge, UK, 2013. [Google Scholar]
- Chiandetti, C. Manipulation of strength of cerebral lateralization via embryonic light stimulation in birds. In Lateralized Brain Functions. Methods in Human and Non-Human Species; Rogers, L.J., Vallortigara, G., Eds.; Humana Press: New York, NY, USA, 2017; pp. 611–631. [Google Scholar]
- Vallortigara, G.; Versace, E. Laterality at the Neural, Cognitive, and Behavioral Levels. In APA Handbook of Comparative Psychology: Vol. 1. Basic Concepts, Methods, Neural Substrate, and Behavior; Call, J., Ed.; American Psychological Association: Washington DC, USA, 2017; pp. 557–577. [Google Scholar]
- Kuan, Y.-S.; Gamse, J.T.; Schreiber, A.M.; Halpern, M.E. Selective asymmetry in a conserved forebrain to midbrain projection. J. Exp. Zool. B Mol. Dev. Evol. 2007, 308, 669–678. [Google Scholar] [CrossRef] [PubMed]
- Roussigné, M.; Blader, P.; Wilson, S.W. Breaking symmetry: The zebrafish as a model for understanding left-right asymmetry in the developing brain. Dev. Neurobiol. 2012, 72, 269–281. [Google Scholar] [CrossRef] [PubMed]
- Gamse, J.T.; Thisse, C.; Thisse, B.; Halpern, M.E. The parapineal mediates left-right asymmetry in the zebrafish diencephalon. Development 2003, 130, 1059–1068. [Google Scholar] [CrossRef] [PubMed]
- Concha, M.L.; Bianco, I.H.; Wilson, S.W. Encoding asymmetry within neural circuits. Nat. Rev. Neurosci. 2012, 13, 832–843. [Google Scholar] [CrossRef] [PubMed]
- Andrew, R.J.; Osorio, D.; Budaev, S. Light during embryonic development modulates patterns of lateralization strongly and similarly in both zebrafish and chick. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2009, 364, 983–989. [Google Scholar] [CrossRef] [PubMed]
- Budaev, S.; Andrew, R.J. Patterns of early embryonic light exposure determine behavioural asymmetries in zebrafish: A habenular hypothesis. Behav. Brain Res. 2009, 200, 91–94. [Google Scholar] [CrossRef] [PubMed]
- Budaev, S.; Andrew, R. Shyness and behavioural asymmetries in larval zebrafish (Brachydanio rerio) developed in light and dark. Behaviour 2009, 146, 1037–1052. [Google Scholar] [CrossRef]
- De Borsetti, N.H.; Dean, B.J.; Bain, E.J.; Clanton, J.A.; Taylor, R.W.; Gamse, J.T. Light and melatonin schedule neuronal differentiation in the habenular nuclei. Dev. Biol. 2011, 358, 251–261. [Google Scholar] [CrossRef] [PubMed]
- Barth, K.A.; Miklósi, A.; Watkins, J.; Bianco, I.H.; Wilson, S.W.; Andrew, R.J. Fsi zebrafish show concordant reversal of laterality of viscera, neuroanatomy, and a subset of behavioral responses. Curr. Biol. 2005, 15, 844–850. [Google Scholar] [CrossRef] [PubMed]
- Levin, M.; Johnson, R.L.; Stern, C.D.; Kuehn, M.R.; Tabin, C. A molecular pathway determining left-right asymmetry in chick embryogenesis. Cell 1995, 82, 803–814. [Google Scholar] [CrossRef]
- Rogers, L.J. Light experience and asymmetry of brain function in chickens. Nature 1982, 297, 223–225. [Google Scholar] [CrossRef] [PubMed]
- Rogers, L.J. Light input and the reversal of functional lateralization in the chicken brain. Behav. Brain Res. 1990, 38, 211–221. [Google Scholar] [CrossRef]
- Rogers, L.J.; Deng, C. Light experience and lateralization of the two visual pathways in the chick. Behav. Brain Res. 1999, 98, 277–287. [Google Scholar] [CrossRef]
- Ströckens, F.; Güntürkün, O. Cryptochrome 1b: A possible inducer of visual lateralization in pigeons? Eur. J. Neurosci. 2016, 43, 162–168. [Google Scholar] [CrossRef] [PubMed]
- Rogers, L.J.; Anson, J.M. Lateralisation of function in the chicken fore-brain. Pharmacol. Biochem. Behav. 1979, 10, 679–686. [Google Scholar] [CrossRef]
- Rogers, L.J.; Zucca, P.; Vallortigara, G. Advantages of having a lateralized brain. Proc. R. Soc. B Biol. Sci. 2004, 271, S420–S422. [Google Scholar] [CrossRef] [PubMed]
- Chiandetti, C.; Vallortigara, G. Effects of embryonic light stimulation on the ability to discriminate left from right in the domestic chick. Behav. Brain Res. 2009, 198, 240–246. [Google Scholar] [CrossRef] [PubMed]
- Chiandetti, C. Pseudoneglect and embryonic light stimulation in the avian brain. Behav. Neurosci. 2011, 125, 775–782. [Google Scholar] [CrossRef] [PubMed]
- Manns, M.; Güntürkün, O. Monocular deprivation alters the direction of functional and morphological asymmetries in the pigeon’s (Columba livia) visual system. Behav. Neurosci. 1999, 113, 1257–1266. [Google Scholar] [CrossRef] [PubMed]
- Mascetti, G.G.; Vallortigara, G. Why do birds sleep with one eye open? Light exposure of the chick embryo as a determinant of monocular sleep. Curr. Biol. 2001, 11, 971–974. [Google Scholar] [CrossRef]
- Vallortigara, G.; Andrew, R.J. Lateralization of response by chicks to change in a model partner. Anim. Behav. 1991, 41, 187–194. [Google Scholar] [CrossRef]
- Vallortigara, G.; Andrew, R.J. Olfactory lateralization in the chick. Neuropsychologia 1994, 32, 417–423. [Google Scholar] [CrossRef]
- Vallortigara, G. Right hemisphere advantage for social recognition in the chick. Neuropsychologia 1992, 30, 761–768. [Google Scholar] [CrossRef]
- Andrew, R.J.; Johnston, A.N.B.; Robins, A.; Rogers, L.J. Light experience and the development of behavioural lateralisation in chicks. II. Choice of familiar versus unfamiliar model social partner. Behav. Brain Res. 2004, 155, 67–76. [Google Scholar] [CrossRef] [PubMed]
- Johnston, A.N.B.; Rogers, L.J. Light exposure of chick embryo influences lateralized recall of imprinting memory. Behav. Neurosci. 1999, 113, 1267–1273. [Google Scholar] [CrossRef] [PubMed]
- Andrew, R.J. Origins of asymmetry in the CNS. Semin. Cell Dev. Biol. 2009, 20, 485–490. [Google Scholar] [CrossRef] [PubMed]
- MacNeilage, P.F.; Rogers, L.J.; Vallortigara, G. Origins of the left & right brain. Sci. Am. 2009, 301, 60–67. [Google Scholar] [PubMed]
- Rogers, L.J. Asymmetry of Brain and Behavior in Animals: Its Development, Function, and Human Relevance. Genesis 2014, 52, 555–571. [Google Scholar] [CrossRef] [PubMed]
- Manns, M.; Ströckens, F. Functional and structural comparison of visual lateralization in birds—Similar but still different. Front. Psychol. 2014, 5, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Chiandetti, C.; Galliussi, J.; Andrew, R.J.; Vallortigara, G. Early-light embryonic stimulation suggests a second route, via gene activation, to cerebral lateralization in vertebrates. Sci. Rep. 2013, 3, 2701. [Google Scholar] [CrossRef] [PubMed]
- Hamburger, V.; Hamilton, H. A series of normal stages in the development of the chick embryo. J. Morphol. 1951, 88, 49–92. [Google Scholar] [CrossRef] [PubMed]
- Tomonari, S.; Takagi, A.; Akamatsu, S.; Noji, S.; Ohuchi, H. A non-canonical photopigment, melanopsin, is expressed in the differentiating ganglion, horizontal, and bipolar cells of the chicken retina. Dev. Dyn. 2005, 234, 783–790. [Google Scholar] [CrossRef] [PubMed]
- Manns, M.; Römling, J. The impact of asymmetrical light input on cerebral hemispheric specialization and interhemispheric cooperation. Nat. Commun. 2012, 3, 696. [Google Scholar] [CrossRef] [PubMed]
- Rogers, L.J. The two hemispheres of the avian brain: Their differing roles in perceptual processing and the expression of behavior. J. Ornithol. 2012, 153, 61–74. [Google Scholar] [CrossRef]
- Agetsuma, M.; Aizawa, H.; Aoki, T.; Nakayama, R.; Takahoko, M.; Goto, M.; Sassa, T.; Amo, R.; Shiraki, T.; Kawakami, K.; et al. The habenula is crucial for experience-dependent modification of fear responses in zebrafish. Nat. Neurosci. 2010, 13, 1354–1356. [Google Scholar] [CrossRef] [PubMed]
- Miklósi, A.; Andrew, R.J. Right eye use associated with decision to bite in zebrafish. Behav. Brain Res. 1999, 105, 199–205. [Google Scholar] [CrossRef]
- Miklósi, A.; Andrew, R.J.; Gasparini, S. Role of right hemifield in visual control of approach to target in zebrafish. Behav. Brain Res. 2001, 122, 57–65. [Google Scholar] [CrossRef]
- Omura, Y.; Oguri, M. Early development of the pineal photoreceptors prior to the retinal differentiation in the embryonic rainbow trout, Oncorhynchus mykiss (Teleostei). Arch. Histol. Cytol. 1993, 56, 283–291. [Google Scholar] [CrossRef] [PubMed]
- Östholm, T.; Brännäs, E.; van Veen, T. The pineal organ is the first differentiated light receptor in the embryonic salmon, Salmo salar L. Cell Tissue Res. 1987, 249, 641–646. [Google Scholar] [CrossRef] [PubMed]
- Harris, J.A.; Guglielmotti, V.; Bentivoglio, M. Diencephalic asymmetries. Neurosci. Biobehav. Rev. 1996, 20, 637–643. [Google Scholar] [CrossRef]
- Braitenberg, V.; Kemali, M. Exceptions to bilateral symmetry in the epithalamus of lower vertebrates. J. Comp. Neurol. 1970, 138, 137–146. [Google Scholar] [CrossRef] [PubMed]
- Bianco, I.H.; Wilson, S.W. The habenular nuclei: A conserved asymmetric relay station in the vertebrate brain. Philos. Trans. R. Soc. B Biol. Sci. 2009, 364, 1005–1020. [Google Scholar] [CrossRef] [PubMed]
- Gurusinghe, C.J.; Ehrlich, D. Sex-dependent structural asymmetry of the medial habenular nucleus of the chicken brain. Cell. Tissue Res. 1985, 240, 149–152. [Google Scholar] [CrossRef] [PubMed]
- Fejér, Z.; Röhlich, P.; Szél, Á.; Dávid, C.; Zádori, A.; Manzano, M.J.; Vígh, B. Comparative ultrastructure and cytochemistry of the avian pineal organ. Microsc. Res. Tech. 2001, 53, 12–24. [Google Scholar] [CrossRef] [PubMed]
- Vígh, B.; Röhlich, P.; Görcs, T.; Maria, M.J.; Szél, Á.; Fejér, Z.; Vígh-Teichmann, I. The pineal organ as a folded retina: Immunocytochemical localization of opsins. Biol. Cell 1998, 90, 653–659. [Google Scholar] [PubMed]
- Nakane, Y. Intrinsic photosensitivity of a deep brain photoreceptor. Curr. Biol. 2014, 24, R596–R597. [Google Scholar] [CrossRef] [PubMed]
- Nakane, Y.; Ikegami, K.; Ono, H.; Yamamoto, N.; Yoshida, S.; Hirunagi, K.; Ebihara, S.; Kubo, Y.; Yoshimura, T. A mammalian neural tissue opsin (Opsin 5) is a deep brain photoreceptor in birds. Proc. Natl. Acad. Sci. USA 2010, 107, 15264–15268. [Google Scholar] [CrossRef] [PubMed]
- Letzner, S.; Patzke, N.; Verhaal, J.; Manns, M. Shaping a lateralized brain: Asymmetrical light experience modulates access to visual interhemispheric information in pigeons. Sci. Rep. 2014, 4, 4253. [Google Scholar] [CrossRef] [PubMed]
- Del Giudice, M. Alone in the dark? Modeling the conditions for visual experience in human fetuses. Dev. Psychobiol. 2011, 53, 214–219. [Google Scholar] [CrossRef] [PubMed]
- Kiuchi, M.; Nagata, N.; Ikeno, S.; Terakawa, N. The relationship between the response to external light stimulation and behavioral states in the human fetus: How it differs from vibroacoustic stimulation. Early Hum. Dev. 2000, 58, 153–165. [Google Scholar] [CrossRef]
- Geschwind, N.; Galaburda, A.M. Cerebral Lateralization: Biological Mechanisms, Associations and Pathology; MIT Press: Cambridge, MA, USA, 1987. [Google Scholar]
- Tran, U.S.; Stieger, S.; Voracek, M. Latent variable analysis indicates that seasonal anisotropy accounts for the higher prevalence of left-handedness in men. Cortex 2014, 57, 188–197. [Google Scholar] [CrossRef] [PubMed]
- Andrew, R.J. Neural and Behavioural Plasticity: The Use of the Domestic Chick as a Model; Oxford University Press: Oxford, UK, 1991. [Google Scholar]
- Vallortigara, G.; Regolin, L.; Zucca, P. Secondary imprinting in the domestic chick: Binocular and lateralized monocular performance. Int. J. Comp. Psychol. 2000, 13, 119–136. [Google Scholar]
Figure 1.
Schematic layout of the experimental apparatus as prepared for the test, with exemplifier distractors placed on both the left and the right side of the starting point and the red beacon signalling the presence of the plate with the mealworms (available only during training). A chick is inspecting the distractors located on the left of the starting point.
Figure 1.
Schematic layout of the experimental apparatus as prepared for the test, with exemplifier distractors placed on both the left and the right side of the starting point and the red beacon signalling the presence of the plate with the mealworms (available only during training). A chick is inspecting the distractors located on the left of the starting point.
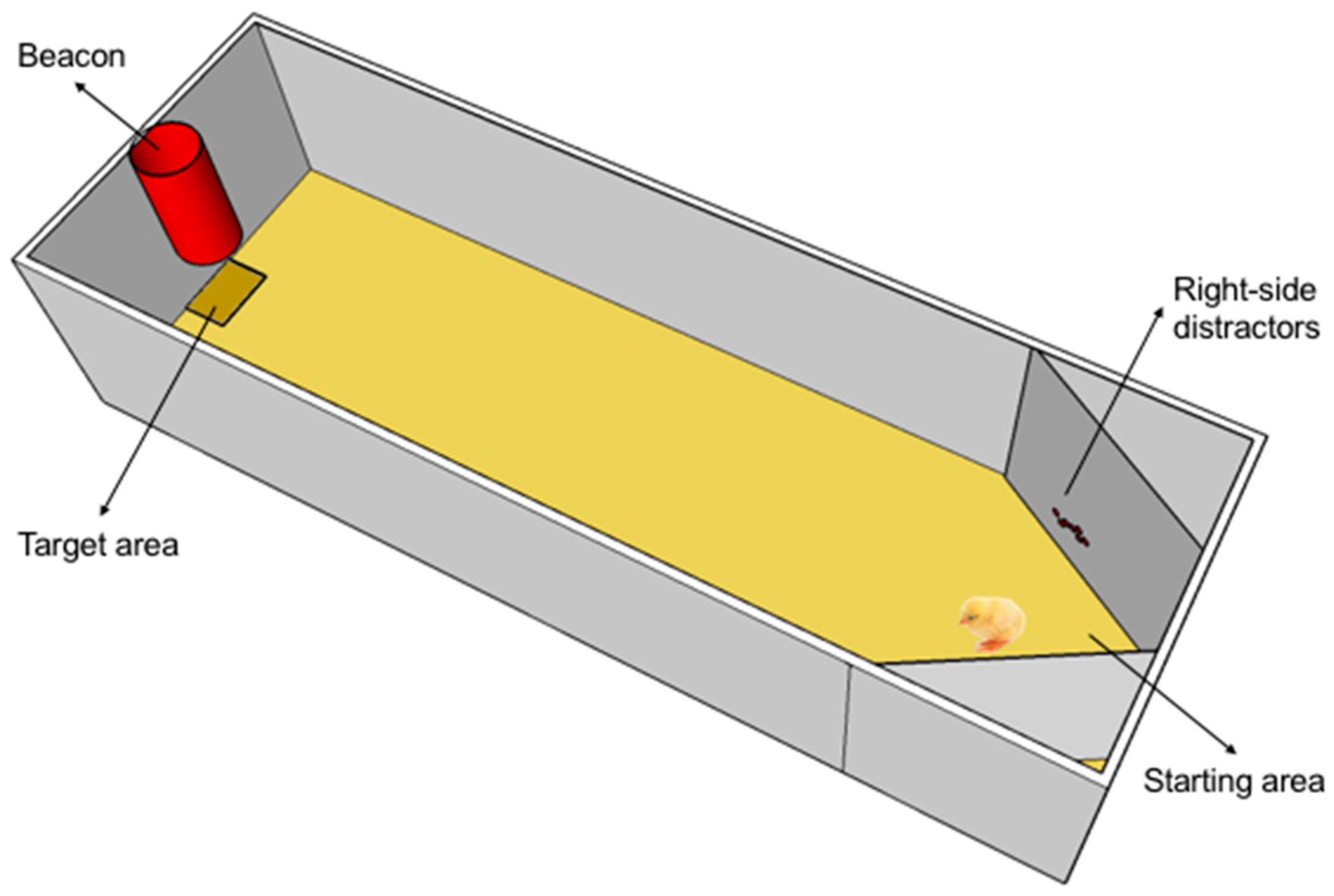
Figure 2.
Plot of the performance (average number of pecks and S.E.M.) of the three groups of chicks in the presence of the distractor (* p < 0.05).
Figure 2.
Plot of the performance (average number of pecks and S.E.M.) of the three groups of chicks in the presence of the distractor (* p < 0.05).
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Chiandetti, C.; Lemaire, B.S.; Versace, E.; Vallortigara, G. Early- and Late-Light Embryonic Stimulation Modulates Similarly Chicks’ Ability to Filter out Distractors. Symmetry 2017, 9, 84. https://doi.org/10.3390/sym9060084
AMA Style
Chiandetti C, Lemaire BS, Versace E, Vallortigara G. Early- and Late-Light Embryonic Stimulation Modulates Similarly Chicks’ Ability to Filter out Distractors. Symmetry. 2017; 9(6):84. https://doi.org/10.3390/sym9060084
Chicago/Turabian StyleChiandetti, Cinzia, Bastien S. Lemaire, Elisabetta Versace, and Giorgio Vallortigara. 2017. "Early- and Late-Light Embryonic Stimulation Modulates Similarly Chicks’ Ability to Filter out Distractors" Symmetry 9, no. 6: 84. https://doi.org/10.3390/sym9060084
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.