


Characterization of Cross-Species Transmission of Drosophila melanogaster Nora Virus
by
 ,
,
Ella G. Buhlke
1,2,†,
Alexis M. Hobbs
2,†,
Sunanda Rajput
2,
Blase Rokusek
2,
Darby J. Carlson
2,
Chelle Gillan
1 and
Kimberly A. Carlson
2,* 1
Central City Senior High School, 1510 28th Street, Central City, NE 68826, USA
2
Department of Biology, University of Nebraska at Kearney, 2401 11th Ave, Kearney, NE 68849, USA
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Life 2022, 12(11), 1913; https://doi.org/10.3390/life12111913
Submission received: 21 October 2022
/
Revised: 7 November 2022
/
Accepted: 15 November 2022
/
Published: 17 November 2022
(This article belongs to the Collection Feature Papers in Microbiology)
Abstract
:Drosophila melanogaster Nora virus (DmNV) is a novel picorna-like virus first characterized in 2006. Since then, Nora virus has been detected in several non-Drosophila species, including insects in the Orders Hymenoptera, Lepidoptera, Coleoptera, and Orthoptera. The objective of this study was to determine if DmNV could infect individuals of other species of invertebrates besides D. melanogaster. The presence of DmNV in native invertebrates and commercially available stocks was determined. Laboratory-reared D. yakuba, D. mercatorum, Gryllodes sigillatus, Tenebrio molitor, Galleria mellonella, and Musca domestica were intentionally infected with DmNV. In addition, native invertebrates were collected and D. melanogaster stocks were purchased and screened for DmNV presence using reverse transcription-polymerase chain reaction (RT-PCR) before being intentionally infected for study. All Drosophila species and other invertebrates, except M. domestica, that were intentionally infected with DmNV ended up scoring positive for the virus via RT-PCR. DmNV infection was also detected in three native invertebrates (Spilosoma virginica, Diplopoda, and Odontotaenius disjunctus) and all commercially available stocks tested. These findings suggest that DmNV readily infects individuals of other species of invertebrates, while also appearing to be an endemic virus in both wild and laboratory D. melanogaster populations. The detection of DmNV in commercially available stocks presents a cautionary message for scientists using these stocks in studies of virology and immunology.
1. Introduction
Drosophila melanogaster Nora virus (DmNV) is a positive-sense single-stranded RNA virus that is spread via the fecal-oral route. DmNV has genetic similarity to viruses of the Picornaviridae and Iflaviridae families, specifically regarding the helicase and RNA polymerase regions, respectively. The DmNV genome has 4 open reading frames (ORFs), with ORF2 encoding a helicase-protease-replicase cassette, characteristic of picornaviruses [1]. Meanwhile, ORF1 encodes an RNAi inhibitor [2], and ORF4 encodes a polyprotein that, after proteolytic cleavage gives rise to 3 mature capsid proteins. Finally, ORF3 appears to encode a capsid-stabilizing protein [3,4,5]. The structure of the virion resembles an icosahedrally symmetric particle, characteristic of viruses within the order Picornavirales [6].
While DmNV is picorna-like, it does not appear to fall neatly within the Picornaviridae family [1]. In fact, since the characterization of DmNV, several similar picorna-like insect viruses have been discovered. These are the Nasonia vitripennis virus 3 in Nasonia parasitoid wasps [7], an unnamed virus related to Nora virus in Haematobia irritans [8], the Spodoptera exigua Nora virus [9], Agrotis ipsilon Nora virus [10], Apis mellifera Nora virus [11] and Helicoverpa armigera Nora virus. These viruses are all genetically similar to the DmNV and appear to cluster together in a previously unknown family of insect viruses distinct from Picornaviridae [12]. Given the similarity of these picorna-like viruses, it is reasonable to postulate that cross-infection of DmNV outside of D. melanogaster is possible.
Finally, the only overt pathogenic effects of DmNV are minimal decreases in survivability and a locomotor deficiency that is apparent through geotaxis assays with large sample sizes [13]. However, while there may be little outward symptomology, DmNV does elicit changes to its host on a gene expression level. Previously, differential gene expression was found in 58 genes in DmNV-infected Canton S wild type flies early in the course of adult infection [14]. Expanding on these findings, differential expression of many genes over the course of DmNV infection, sampling flies on days 2, 10, 20, and 30 after infection, was also shown [15]. This is a rather troubling reality for D. melanogaster research, as unrecognized DmNV infection in laboratory stocks could, in theory, confound experimental results. For this reason, stocks ordered from a stock center were tested to investigate the prevalence of DmNV in laboratory-reared D. melanogaster.
The purpose of this study was to investigate the ability of DmNV to infect individuals of other species of Drosophila, and other insects, including Gryllodes sigillatus (crickets), Tenebrio molitor (mealworms), Galleria mellonella (wax moths), and Musca domestica (house flies) in the laboratory setting via an established infection protocol. Further, to investigate the potential for DmNV cross-species infection in the wild, insects were collected and tested for the presence of DmNV infection via RNA extraction and reverse transcription-polymerase chain reaction (RT-PCR) using primers specific for DmNV ORF1. Finally, the presence of DmNV infection in laboratory stocks of D. melanogaster from a stock center was performed to raise awareness of the prevalence of DmNV in D. melanogaster populations within the research laboratory.
2. Materials and Methods
2.1. Fly Husbandry and Dechorionation
Witi RelE23 (a kind gift from Dan Hultmark from Umeå, Sweden) D. melanogaster were maintained at 25 °C on standard cornmeal, molasses and torula yeast medium with diurnal light. Flies were either infected fecal-orally to establish Nora virus-infected (NV+) stocks or maintained uninfected for further analysis. Once adequately established, stocks were expanded for fly collection by transferring flies into new bottles. The Witi RelE23 stocks were checked weekly via RT-PCR for NV infection via Section 2.8 to verify that they were productively infected with DmNV.
Stock bottles were routinely checked to verify uninfected status. If a stock became NV+, embryos were collected on apple juice agar plates and dechorionated in 2.7% hypochlorite for 2–5 min. The dechorionated embryos were washed with 1X Drosophila Ringer’s solution (3 mM CaCl2·H2O, 182 mM KCl, 46 mM NaCl, 10 mM Tris base, pH adjusted to 7.2, and was sterilized by autoclaving), placed on fresh food, and reared under standard conditions. Once the adults had developed, they were subjected to testing for NV infection via Section 2.8.
2.2. Infection of Individuals from Different Drosophila Species
Drosophila yakuba and D. mercatorum were ordered from the National Drosophila Species Stock Center. D. yakuba originated from Nairobi, Kenya, D. mercatorum originated from Sao Paulo, Brazil, and both were listed as wild-type for their species. The infection protocol used in this study has been well established [14,16]. For the infection protocol, five NV+ D. melanogaster males were reared on Formula 4-24® Instant Blue Drosophila Medium (Carolina Biological Supply Company, Burlington, NC, USA). After 96 h, the NV+ D. melanogaster males were removed and replaced with 5 Nora virus negative (NV−) males and 5 NV− females of each species: D. yakuba, D. mercatorum, and D. melanogaster. Flies were collected after 5 days and stored at −80 °C. Three biological replicates with ten flies each were tested in duplicate for NV infection via Section 2.8.
2.3. Infection of M. domestica and G. mellonella
Ten NV+ D. melanogaster males were placed into 10 vials with Formula 4-24® Instant Drosophila Medium (Carolina Biological Supply Company). The males were allowed to defecate on the food for 96 h to ensure adequate transfer of Nora virus to the surface of the food. After 96 h, 10 M. domestica pupa (Carolina Biological Supply Company) and 10 G. mellonella larva (Carolina Biological Supply Company) were added to individual vials and allowed to eclose at room temperature. The mature M. domestica and G. mellonella remained on the infected food for 7 days to allow for a productive infection. Collected M. domestica and G. mellonella were stored at −80 °C. Three biological replicates with ten M. domestica or G. mellonella each were tested in duplicate for NV infection via Section 2.8.
2.4. Infection of G. sigillatus
Ten NV+ D. melanogaster were placed into vials containing a 1:1 combination of Formula 4-24® Instant Drosophila Medium (Carolina Biological Supply Company) and Instant Dry Cricket medium (Carolina Biological Supply Company). The males were allowed to defecate on the food for 96 h to ensure adequate transfer of Nora virus to the surface of the food. After 96 h, 5 nymph G. sigillatus (Carolina Biological Supply Company) were added to pre-prepared bug containers with sterile sand and the infected food was placed inside. The G. sigillatus remained on the infected food for 7 days. Collected G. sigillatus were stored at −80 °C. Three biological replicates with ten G. sigillatus each were tested in duplicate for NV infection via Section 2.8.
2.5. Infection of T. molitor
Ten small plastic insect containers were prepared with soil and potato pieces. Five T. molitor (Carolina Biological Supply Company) were placed into each container. Every third day, the T. molitor were fed 30 NV+ D. melanogaster for 14 days. After 14 days, the T. molitor were collected and stored at −80 °C. Three biological replicates with ten T. molitor each were tested in duplicate for NV infection via Section 2.8.
2.6. Collection of Native Invertebrates
Native insects were randomly collected from counties Hamilton, Butler, Buffalo, Merrick, Hall, and Polk in Nebraska. Invertebrates were placed in sealable baggies and placed on ice. Once collected, insects were stored at −80 °C. Native insects were individually tested in duplicate for NV infection via Section 2.8.
2.7. Commercially Available D. melanogaster Stocks
Drosophila stocks were purchased from the Bloomington Drosophila Stock Center (BDSC) to investigate the prevalence of DmNV in commercially available/laboratory-reared D. melanogaster. Canton Special wild-type (CS+; stock #64,349), Oregon-R wild-type (Ore-R+; Stock #5), vestigial (vg; Stock #432), and apterous (ap; stock #4189) D. melanogaster stocks were purchased. Upon receipt of the stocks, they were quarantined in a separate room isolated from the other stocks maintained for research purposes that were known to be infected with Nora virus. Ten emergents from each stock vial were collected within 24 h of eclosion and tested in duplicate for Nora virus infection per Section 2.8.
2.8. RNA Extraction and RT-PCR Analysis of Nora Virus
Total RNA extraction was performed using TRIzol® per manufacturer’s instructions (ThermoFisher Scientific, Waltham, MA, USA). Each sample was quantitated using a NanoDropTM ONE spectrophotometer (ThermoFisher Scientific) to assess RNA purity (260/280 ≈ 2.0) and concentration. Samples were analyzed for the presence of Nora virus using Nora ORF1 55–844 (Forward 5′-TGGTAGTACGCAGGTTGTGGGAAA-3′; Reverse 5′-AAGTCATGCTGGCTTCTCAAC-3′) primers and qScript XLT 1-Step RT-PCR (Quantabio, Beverly, MA) according to manufacturer’s instructions. The positive controls were an RNA extraction that previously tested positive for Nora virus. Reactions using 250 ng of total RNA were set-up under the following conditions for Nora virus: 50 °C for 30 min, 94 °C for 2 min, (94 °C for 30 s, 55 °C for 30 s, 68 °C for 1 min) for 30 cycles, 68 °C for 5 min, and hold at 4 °C. Samples were analyzed on a 1.0% agarose gel in a TAE buffer solution at 50 V for 3 h. A positive reaction yielded a product at approximately 790 bp for DmNV [13,14,15]. DmNV positive PCR products were submitted for sequencing verification by preparing 20 ng of the purified PCR product along with 25 pM of forward primer for the target gene. These samples were sent to the University of Nebraska Medical Center (UNMC) Genomics Core Facility, for traditional Sanger sequencing via Genewiz (Azenta, Chelmsford, MA, USA). The resulting sequence files were uploaded into the NCBI Nucleotide BLAST Program (https://blast.ncbi.nlm.nih.gov/Blast.cgi; accessed on 1 June 2022) to determine sequence identity. All sequenced products were identified as DmNV glycoprotein 1 (gp1; ORF1).
3. Results
3.1. Validation of Nora virus Infection Using RT-PCR
The presence of DmNV was analyzed by RT-PCR using gene specific primers for ORF1. A 790 bp product for ORF1 verified DmNV infection (Figure 1A, lanes 2 & 3; Supplemental Table S1). Stocks that were reared to be infected with DmNV demonstrated a 790 bp product confirming infection. Uninfected stocks were also tested for the presence of DmNV and were found to be negative for the DmNV ORF1 RT-PCR product in D. melanogaster (Figure 1A, lanes 4 & 5). All Drosophila species and laboratory reared insects that were to be subjected to the DmNV infection protocol were tested before being used and all were found to be negative for the presence of DmNV infection.
3.2. Nora Virus Is Transmitted across Drosophila Species
DmNV infection was found in D. melanogaster (Figure 1A, lanes 2 & 3; Supplemental Table S1), as expected, and in both D. yakuba and D. mercatorum (Figure 1B; Supplemental Table S1). Drosophila yakuba showed a higher incidence of infection with all 3 biological replicates appearing positive (Figure 1B, lanes 3–5; Supplemental Table S1). In contrast, D. meractorum had 2 out of 3 biological replicates appear positive (Figure 1B, lanes 6–8; Supplemental Table S1).
3.3. Nora Virus Is Transmitted to Individuals of Select Laboratory Reared Insect Species
To determine whether DmNV could infect individuals of species outside of the Drosophila genus, four commercially available insects were tested. All laboratory reared insects that were to be subjected to the DmNV infection protocol were tested before being used and all were found to be negative for the presence of DmNV infection. M. domestica (house fly) was selected because it is a non-Drosophilidae Dipteran species. G. sigillatus (house cricket), G. mellonella (wax moth), and T. molitor (mealworm) were selected because they are commercially available, easy to rear, and easily adaptable for the infection protocol. M. domestica were not able to be infected with DmNV (Figure 2, lanes 2–4; Supplemental Table S1), as demonstrated by the absence of a 790 bp product. G. sigillatus showed that 2 out of 3 biological replicates became infected (Figure 2, lanes 5–7; Supplemental Table S1). G. mellonella exhibited infection in all 3 biological replicates (Figure 2, lanes 8–10; Supplemental Table S1). T. molitor demonstrated a positive product in 2 out of 3 biological replicates (Figure 2, Lanes 11–13; Supplemental Table S1). In toto, these results demonstrate that DmNV can successfully cross-infect other individuals of species of insects.
3.4. Nora Virus Is Present in Individuals of Native Nebraska Invertebrate Species
In order to determine whether DmNV is present in native non-laboratory reared invertebrates, select invertebrates were collected from multiple counties in Central Nebraska and tested for DmNV. Thirty-two different individuals from native invertebrate species from 8 Nebraska counties were tested. Using RT-PCR, DmNV infection was found in 3 individuals from native species in two counties, Merrick and Hall, across Central Nebraska (Figure 3). These individuals include Spilosoma virginica, Diplopoda, and Odontotaenius disjunctus (Table 1).
3.5. Nora Virus Is Present in Commercially Available Drosophila Stocks
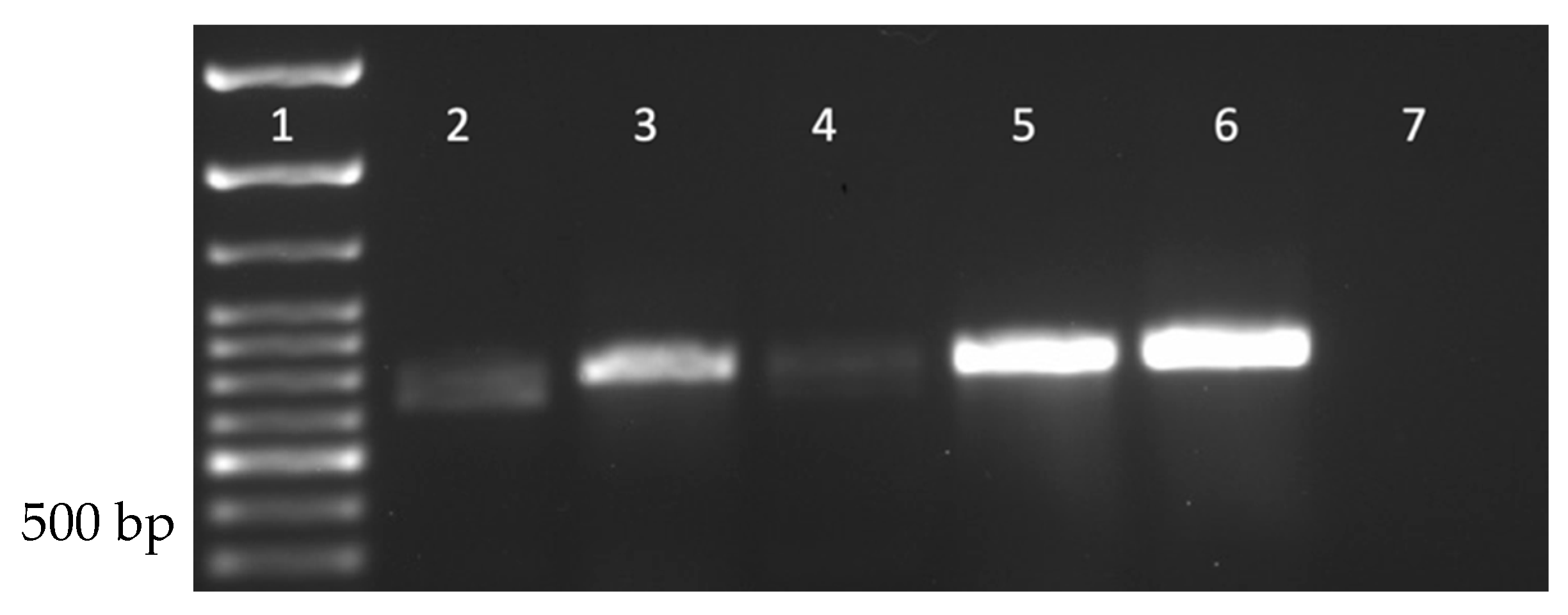
Drosophila stocks were purchased to investigate the prevalence of DmNV in commercially available/laboratory-reared D. melanogaster. CS+, Ore-R+, vg, and ap D. melanogaster stocks were purchased because they are among the most used stocks for genetic studies (CS+ and OreR+) and/or are currently being used in the laboratory (CS+, OreR+, vg, and ap). Using RT-PCR, DmNV infection was found in all stocks purchased (Figure 4).
4. Discussion
The current study provides evidence that DmNV may be circulating not only within D. melanogaster populations, but also other Drosophila species, and other lab insects, as well as native insects. DmNV was thought to have a narrow host range [1], but the data presented here provide support for a broader host range. Initially, DmNV was found to not infect D. yakuba [1]. Interestingly, D. yakuba showed infection in all biological replicates, whereas D. mercatorum showed infection in only 2 of the biological replicates (Figure 1B). This could be because DmNV is not well adapted to the D. mercatorum species, as it is more distantly related to D. melanogaster, both genetically and geographically, than D. yakuba is [17]. Persistently infected flies produce DmNV at a rate on the order of 109 viral genomes per fly and 107–1010 viral genomes per fly per 5 h in the feces [16], therefore intentional infection may account for the differences between our results and earlier published results [1] with D. yakuba. The objective of this project was the detection of presence or absence of DmNV infection and not the quantification of viral load. Therefore, we were not able to determine if the viral load was higher in the individuals from the different species.
Not only was DmNV able to infect individuals of other Drosophila species, it was also able to infect laboratory-reared insects. DmNV was able to infect G. mellonella (Order: Lepidoptera –moth), G. sigillatus (Order: Orthoptera–cricket), and T. molitor (Order: Coleoptera–beetle) (Figure 2), but not M. domestica (Order: Diptera, Family: Muscidae). The viral connections between some of these species and D. melanogaster have been shown in past studies with different viruses. For example, G. sigillatus is susceptible to Cricket Paralysis virus (CrPV) [18], which is also known to infect D. melanogaster, and G. mellonella [19]. In addition, Invertebrate iridescent virus 6 (IIV-6) infects many insects including D. melanogaster, G. mellonella [20], and T. molitor [21], whereas G. sigillatus can be infected by a related iridovirus, Cricket iridovirus (CrIV) [22]. Additionally, new picorna-like Nora viruses have been found in non-Drosophilidae Dipterans, such as H. irritans (Order: Diptera, Family: Muscidae) [8], S. exigua (Order: Lepidoptera) [9], A. ipsilon (Order: Lepidoptera) [10], A. mellifera (Order: Hymenoptera) [11] and H. armigera (Order: Lepidoptera) [11]. Furthermore, it was also found that Nora virus is present in 3 individuals from different native invertebrate species within central Nebraska (Table 1). These invertebrate species represented include members of the Class Insecta, such as S. virginica from Order Lepidoptera and O. disjunctus from the Order Coleoptera, as well a non-Insecta class species, Diplopoda (Class: Diplopoda). This indicates that invertebrates in the Orders Lepidoptera and Coleoptera, may share similar virus receptors to Drosophila, specifically when considering DmNV. This also demonstrates the breadth of organisms that can be infected by DmNV, both in the laboratory and in nature. These data corroborate the hypothesis that DmNV may represent the first member of a widespread family of viruses [23]. Interestingly, M. domestica was not able to be infected in the laboratory setting (Figure 2), nor was DmNV detected in the native sample collected (Table 1). Upon an extensive review of the literature, there was no indication that M. domestica could be cross-infected with other Drosophila viruses (Drosophila C virus; DCV, CrPV, Sigma virus, Drosophila X virus, or IIV-6). Therefore, not being able to infect individuals of this species with DmNV is not surprising.
Due to the ability of DmNV to not only cross-infect individuals of other species of Drosophila but also other invertebrates both artificially and naturally, the prevalence of infection in commercially available stocks was tested. RT-PCR of DmNV ORF1 of these stocks indicates that DmNV is present within all 4 stocks purchased (Figure 4). This is not surprising since DmNV has been previously detected in both laboratory-reared and wild-caught samples [1,24], as well as a contaminant in solutions and other laboratory preparations [16].
As noted earlier, the only observed phenotype with DmNV infection is a geotaxis defect detectable when large sample sizes are analyzed via a geotaxis assay [13]. What is of concern is the effect of DmNV on the innate immune system of Drosophila species. After four days of persistent infection, DmNV infected Canton S flies were found to have genes involved in the Toll and immune deficient (Imd) pathways, Janus Kinase Signal Transducer and Activator of Transcription (Jak-Stat) interactions, as well as gut-specific innate immune responses differentially regulated [14]. In another DmNV infection experiment, w118 flies were persistently infected and next generation sequencing performed at 2, 10, 20, and 30 days post-infection. The results indicated an increase in immune related gene expression over time with vago and vir-1 (virus induced RNA 1) identified as candidate markers of DmNV infection [15], similar to what occurs with DCV infection of D. melanogaster [25,26]. This immune gene activation becomes a problem when researchers do not know that their stocks are infected with DmNV. One possible outcome is viral interference, where one virus competitively suppresses replication of another coinfecting virus. Another outcome is that coinfections modulate virus virulence and cell death, which alters severity of infection. Lastly, immunity to the primary infection reduces the immune response to the secondary infection (reviewed in [27]). All of these outcomes are unfavorable to any laboratory investigation involving D. melanogaster, especially those examining gene regulation or innate immunity. The data presented suggests that DmNV has the ability to cross-infect not only individuals of other Drosophila species but other organisms, and unknown contamination of stocks from either stock centers or other laboratories. It is important to quarantine stocks upon receipt and subsequently test for infection before carrying out experiments. Fortunately, since DmNV is horizontally transferred, infected stocks can be easily cured by dechorionation of embryos and transferring these to fresh food.
5. Conclusions
In conclusion, this study demonstrates that the host range of DmNV is perhaps larger than previously expected. Namely, DmNV has the ability to infect not only Drosophila species but other invertebrates as well. Further examination of DmNV or DmNV-like viruses in these native invertebrate species needs to be performed. In addition, the receptor that gives DmNV access to host cells needs to be elucidated. This would give a better understanding as to how DmNV is able to infect other organisms besides D. melanogaster. Lastly, this investigation offers a cautionary tale for those who perform research in the realm of virology and immunology using stocks from a stock center or donated from other laboratories, as DmNV infection appears to be endemic to many laboratory-reared stocks.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/life12111913/s1, Table S1: Nora virus infection of Drosophila species and other laboratory-reared insects. Each biological replicate consists of 10 individuals that have been pooled together.
Author Contributions
Conceptualization, E.G.B., A.M.H., C.G., D.J.C. and K.A.C.; methodology, E.G.B., A.M.H., C.G., D.J.C. and K.A.C.; software, C.G., K.A.C.; validation, E.G.B., A.M.H., S.R., B.R., D.J.C. and K.A.C.; formal analysis, E.G.B., A.M.H., S.R., B.R., D.J.C. and K.A.C.; investigation, E.G.B., A.M.H., S.R., B.R., D.J.C. and K.A.C.; resources, C.G. and K.A.C.; data curation, K.A.C.; writing—original draft preparation, E.G.B., A.H, S.R., B.R. and K.A.C.; writing—review and editing, C.G. and D.J.C.; visualization, E.G.B., A.M.H. and K.A.C.; supervision, C.G. and K.A.C.; project administration, C.G. and K.A.C.; funding acquisition, C.G. and K.A.C. All authors have read and agreed to the published version of the manuscript.
Funding
The project described was supported by grants from the National Institute for General Medical Science (NIGMS; GM103427) a component of the National Institutes of Health (NIH), and Carlson is also supported by a grant from the National Institute of General Medical Sciences, 1U54GM115458. The UNMC Genomics Core receives partial support from the NIGMS INBRE-P20GM103427-19 grant as well as The Fred & Pamela Buffett Cancer Center Support Grant-P30 CA036727. This publication’s contents are the sole responsibility of the authors and do not necessarily represent the official views of the NIH or NIGMS.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
Hultmark and Ekström for their gift of the Witi RelE23 DmNV infected stocks. Kevin Cook of the Blooming Drosophila Stock Center for discussion.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Habayeb, M.S.; Ekengren, S.K.; Hultmark, D. Nora virus, a present virus in Drosophila, defines a new picorna-like virus family. J. Gen. Virol. 2006, 87, 3045–3051. [Google Scholar] [CrossRef]
- van Mierlo, J.T.; Bronkhorst, A.W.; Overheul, G.J.; Sadanandan, S.A.; Ekström, J.-O.; Heestermans, M.; Hultmark, D.; Antoniewski, C.; van Rij, R.P. Convergent evolution of argonaute-2 slicer antagonism in two distinct insect RNA viruses. PLoS Pathog. 2012, 8, e1002872. [Google Scholar] [CrossRef] [PubMed]
- Ekström, J.-O.; Habayeb, M.S.; Srivastava, V.; Kieselbach, T.; Wingsle, G.; Hultmark, D. Drosophila Nora virus capsid proteins differ from those of other picorna-like viruses. Virus Res. 2011, 160, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Licking-Murray, K.D.; Carlson, D.J.; Sowle, R.; Carlson, K.A. In vitro assembly and evaluation of Nora virus VLPs. Acta Virol. 2021, 65, 381–389. [Google Scholar] [CrossRef]
- Sadanandan, S.A.; Ekström, J.-O.; Jonna, V.R.; Hofer, A.; Hultmark, D. VP3 is crucial for the stability of Nora virus virions. Virus Res. 2016, 223, 20–27. [Google Scholar] [CrossRef] [Green Version]
- Laurinmäki, P.; Shakeel, S.; Ekström, J.-O.; Mohammadi, P.; Hultmark, D.; Butcher, S.J. Structure of Nora virus at 2.7 Å resolution and implications of receptor binding, capsid stability and taxonomy. Sci. Rep. 2020, 10, 19675. [Google Scholar] [CrossRef]
- Oliveira, D.C.S.G.; Hunter, W.B.; Ng, J.; Desjardins, C.A.; Dang, P.M.; Werren, J.H. Data mining cDNAs reveals three new single stranded RNA viruses in Nasonia (Hymenoptera: Pteromalidae). Insect Mol. Biol. 2010, 19, 99–107. [Google Scholar] [CrossRef] [Green Version]
- Torres, L.; Almazán, C.; Ayllón, N.; Galindo, R.C.; Rosario-Cruz, R.; Quiroz-Romero, H.; Gortazar, C.; De La Fuente, J. Identification of microorgnisms in partially fed female horn flies, Haematobia irritans. Parasitol. Res. 2012, 111, 1391–1395. [Google Scholar] [CrossRef]
- Jakubowska, A.K.; D’Angiolo, M.; González-Martínez, R.M.; Millán-Leiva, A.; Carballo, A.; Murillo, R.; Caballero, P.; Herrero, S. Simultaneous occurrence of covert infections with small RNA viruses in the lepidopteran Spodoptera exigua. J. Invertebr. Pathol. 2014, 121, 56–63. [Google Scholar] [CrossRef]
- Li, T.; Guan, R.; Wu, Y.; Chen, S.; Yuan, G.; Miao, X.; Li, H. The novel Agrotis ipsilon Nora virus confers deleterious effects to the fitness of Spodoptera frugiperda (Lepidoptera: Noctuidae). Front. Microbiol. 2021, 12, 727202. [Google Scholar] [CrossRef]
- Remnant, E.J.; Shi, M.; Buchmann, G.; Blacquiere, T.; Holmes, E.C.; Beekman, M.; Ashe, A. A diverse range of novel RNA viruses in geographically distinct honey bee populations. J. Virol. 2017, 91, e00158-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, X.; Xu, P.; Yuan, H.; Graham, R.I.; Wilson, K.; Wu, K. Discovery and characterization of a novel picorna-like RNA virus in the cotton bollworm Helocoverpa armigera. J. Invertebr. Pathol. 2019, 160, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rogers, A.; Towery, L.; McCown, A.; Carlson, K.A. Impaired geotaxis as a novel phenotype of Nora virus infection of Drosophila melanogaster. Scientifica 2020, 2020, 1804510. [Google Scholar] [CrossRef]
- Cordes, E.J.; Licking-Murray, K.D.; Carlson, K.A. Differential gene expression related to Nora virus infection of Drosophila melanogaster. Virus Res. 2013, 175, 95–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopez, W.; Page, A.M.; Carlson, D.J.; Ericson, B.L.; Cserhati, M.F.; Guda, C.G.; Carlson, K.A. Analysis of immune-related genes during Nora virus infection of Drosophila melanogaster using next generation sequencing. AIMS Microbiol. 2018, 4, 123–139. [Google Scholar] [CrossRef] [PubMed]
- Habayeb, M.S.; Cantera, R.; Casanova, G.; Ekström, J.-O.; Albright, S.; Hultmark, D. The Drosophila Nora virus is an enteric virus, transmitted via feces. J. Invertebr. Pathol. 2009, 101, 29–33. [Google Scholar] [CrossRef]
- Jezovit, J.A.; Levine, J.D.; Schneider, J. Phylogeny, environment and sexual communication across the Drosophila genus. J. Exp. Biol. 2017, 220, 42–52. [Google Scholar] [CrossRef] [Green Version]
- Gryllodes sigillatus Draft Assessment Report. Available online: https://www.awe.gov.au/sites/default/files/env/consultations/03754e31-19da-4746-ab08-c2bf575da692/files/gryllodes-sigillatus-draft-assessment-report.pdf (accessed on 21 June 2022).
- Moore, N.F.; Kearns, A.; Pullin, J.S. Characterization of Cricket paralysis virus-induced polypeptides in Drosophila cells. J. Virol. 1980, 33, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Constantino, M.; Christian, P.; Marina, C.F.; Williams, T. A comparison of techniques for detecting Invertebrate iridescent virus 6. J. Virol. Meth. 2001, 98, 109–118. [Google Scholar] [CrossRef]
- Gencer, D.; Yesilyurt, A.; Gullu, M.; Nalcacioglu, R. Insecticidal activities of wild type and recombinant invertebrate iridescent viruses on five common pests. Turk. Entomol. Derg. 2020, 44, 365–373. [Google Scholar] [CrossRef]
- Duffield, K.R.; Hunt, J.; Sadd, B.M.; Sakaluk, S.K.; Oppert, B.; Rosario, K.; Behle, R.W.; Ramirez, J.L. Active and covert infections of Cricket iridovirus and Acheta domesticus densovirus in reared Gryllodes sigillatus crickets. Front. Microbiol. 2021, 12, 780796. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Cherry, S. Viruses and antiviral immunity in Drosophila. Dev. Comp. Immunol. 2014, 42, 67–84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munn, B.; Ericson, B.; Carlson, D.J.; Carlson, K.A. Detecting the presence of Nora virus in Drosophila utilizing single fly RT-PCR. Bioscene J. Coll. Biol. Teach. 2015, 41, 40–44. [Google Scholar]
- Deddouche, S.; Matt, N.; Budd, A.; Mueller, S.; Kemp, C.; Galiana-Arnoux, D.; Dostert, C.; Antoniewski, C.; Homan, J.A.; Imler, J.-L. The DExD/H-box helicase Dicer-2 mediates the induction of antiviral activity in Drosophila. Nat. Immunol. 2008, 9, 1425–1432. [Google Scholar] [CrossRef] [PubMed]
- Dostert, C.; Jouanguy, E.; Irving, P.; Troxler, L.; Galiana-Arnoux, D.; Hetru, C.; Humann, J.A.; Imler, J.-L. The Jak-STAT signaling pathway is required but not sufficient for the antiviral response of Drosophila. Nat. Immunol. 2005, 6, 946–953. [Google Scholar] [CrossRef]
- Kumar, N.; Sharma, S.; Barua, S.; Tripathi, B.N.; Rouse, B.T. Virological and immunological outcomes of coinfections. Clin. Microbiol. Rev. 2018, 31, e00111–e00117. [Google Scholar] [CrossRef]
Figure 1.
Nora virus presence in individuals from different Drosophila species (A) D. melanogaster. Lane 1: 100 bp ladder; Lanes 2 & 3: NV+ D. melanogaster; Lanes 4 & 5: NV− D. melanogaster. (B) D. yakuba and D. mercatorum. Lane 1: 100 bp ladder; Lane 2: NV+ D. melanogaster (positive control); Lanes 3–5: D. yakuba; Lanes 6–8: D. meractorum; Lane 9: water (negative control). Each lane represents a separate biological replicate and a product of approximately 790 bp is indicative of DmNV infection.
Figure 1.
Nora virus presence in individuals from different Drosophila species (A) D. melanogaster. Lane 1: 100 bp ladder; Lanes 2 & 3: NV+ D. melanogaster; Lanes 4 & 5: NV− D. melanogaster. (B) D. yakuba and D. mercatorum. Lane 1: 100 bp ladder; Lane 2: NV+ D. melanogaster (positive control); Lanes 3–5: D. yakuba; Lanes 6–8: D. meractorum; Lane 9: water (negative control). Each lane represents a separate biological replicate and a product of approximately 790 bp is indicative of DmNV infection.
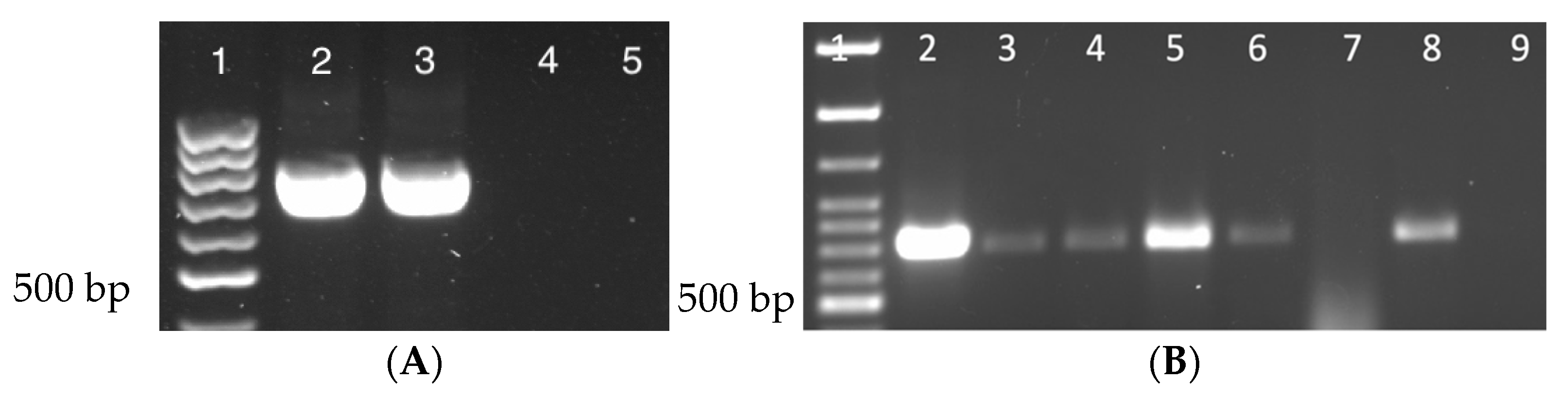
Figure 2.
Nora virus presence within laboratory-reared insects. Lane 1: 100 bp ladder; Lanes 2–4: Musca domestica; Lanes 5–7: G. sigillatus; Lanes 8–10: G. mellonella; Lanes 11–13: T. molitor; Lane 14: D. melanogaster (positive control); Lane 15: water (negative control). Each lane represents a separate biological replicate and a product of approximately 790 bp is indicative of DmNV infection.
Figure 2.
Nora virus presence within laboratory-reared insects. Lane 1: 100 bp ladder; Lanes 2–4: Musca domestica; Lanes 5–7: G. sigillatus; Lanes 8–10: G. mellonella; Lanes 11–13: T. molitor; Lane 14: D. melanogaster (positive control); Lane 15: water (negative control). Each lane represents a separate biological replicate and a product of approximately 790 bp is indicative of DmNV infection.
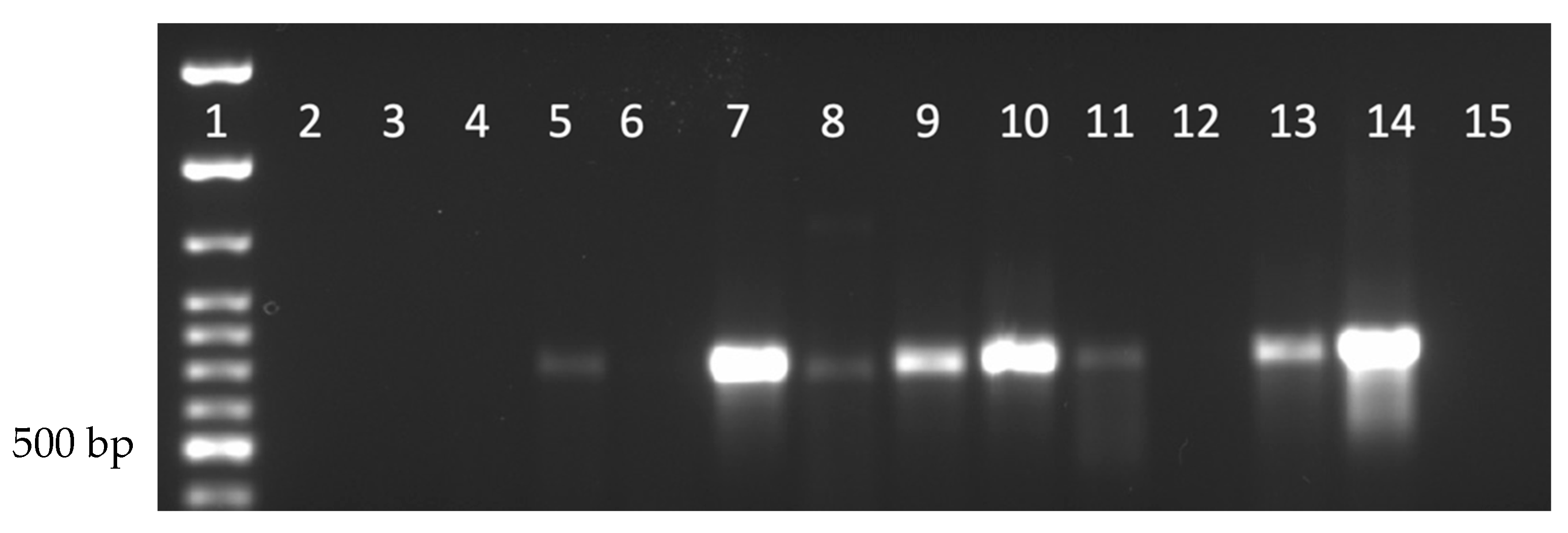
Figure 3.
Collection locations in Nebraska of native invertebrates. Counties sampled included A. Boone, B. Polk, C. Merrick, D. Butler, E. Buffalo, F. Hall, G. Hamilton, and H. Clay.
Figure 3.
Collection locations in Nebraska of native invertebrates. Counties sampled included A. Boone, B. Polk, C. Merrick, D. Butler, E. Buffalo, F. Hall, G. Hamilton, and H. Clay.
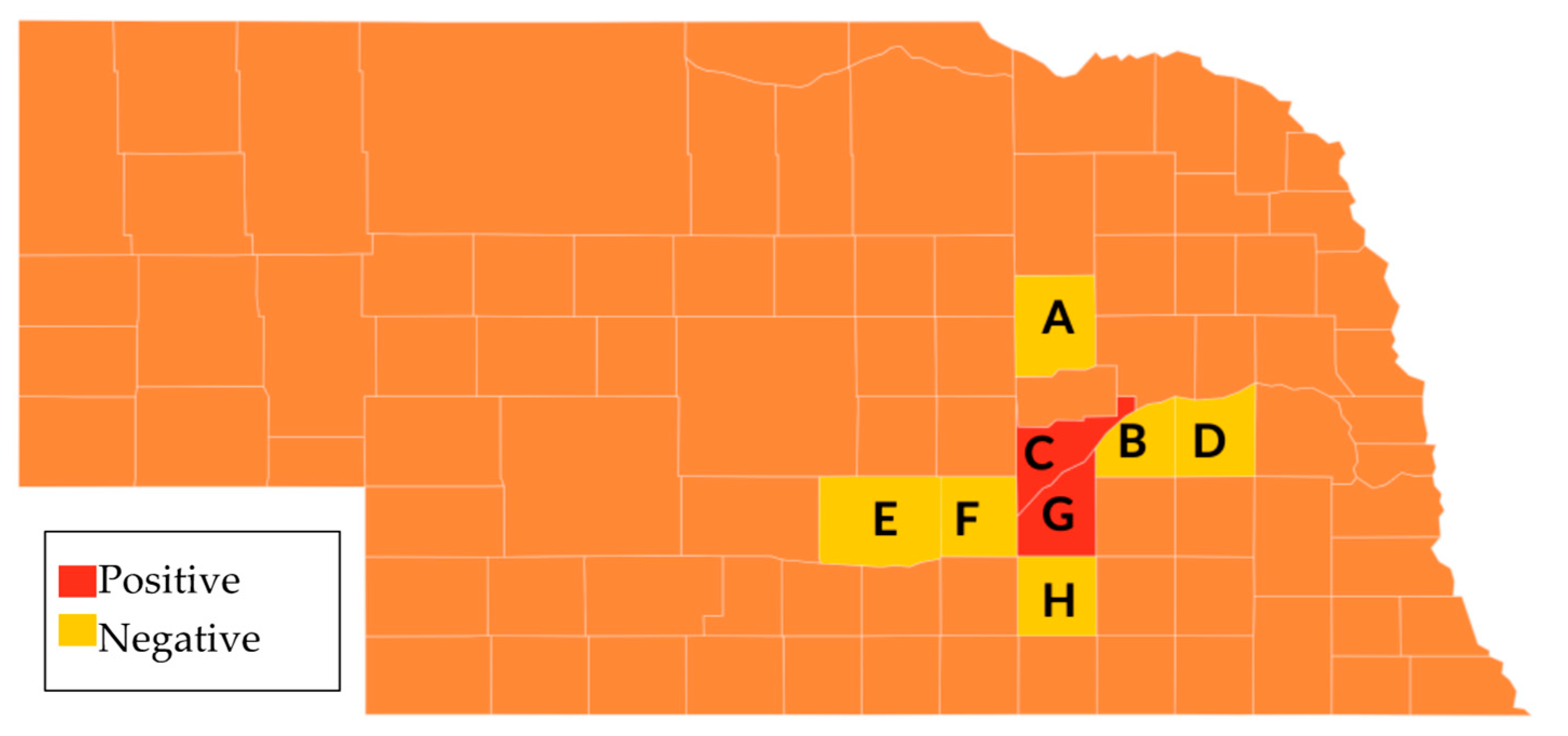
Figure 4.
Nora virus is present within Drosophila stocks commercially available from the BDSC. Lane 1: 100 bp ladder; Lane 2: NV+ Ore-R+ D. melanogaster; Lane 3: NV+ vg D. melanogaster; Lane 4: NV+ ap D. melanogaster; Lane 5: NV+ CS+ D. melanogaster; Lane 6: known NV+ D. melanogaster (positive control); Lane 7: water (negative control). A product of approximately 790 bp is indicative of DmNV infection.
Figure 4.
Nora virus is present within Drosophila stocks commercially available from the BDSC. Lane 1: 100 bp ladder; Lane 2: NV+ Ore-R+ D. melanogaster; Lane 3: NV+ vg D. melanogaster; Lane 4: NV+ ap D. melanogaster; Lane 5: NV+ CS+ D. melanogaster; Lane 6: known NV+ D. melanogaster (positive control); Lane 7: water (negative control). A product of approximately 790 bp is indicative of DmNV infection.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Individuals of native Nebraska invertebrate species collected to determine Nora virus infection status.
Table 1.
Individuals of native Nebraska invertebrate species collected to determine Nora virus infection status.
| Scientific Name | Common Name | County | Result |
|---|---|---|---|
| Camponotus | Carpenter ant | Hamilton | Negative |
| Anax junius | Green darner | Merrick | Negative |
| Harmonia axyridis | Asian lady beetle | Merrick | Negative |
| Mantodea | Mantis | Merrick | Negative |
| Polistes fuscatus | Northern paper wasp | Butler | Negative |
| Helcystogramma badia | N/A | Merrick | Negative |
| Leucoma salicis | White satin moth | Merrick | Negative |
| Acheta domesticus | House cricket | Merrick | Negative |
| Apis mellifera | Western honey bee | Merrick | Negative |
| Spilosoma virginica | Yellow woolly bear | Merrick | Positive |
| Musca domestica | House fly | Merrick | Negative |
| Melanoplus femurrubrum | Red-legged grasshopper | Merrick | Negative |
| Phoberia atomaris | Common oak moth | Merrick | Negative |
| Hyles lineata | White-lined sphinx | Merrick | Negative |
| Ceratomia amyntor | Elm sphinx | Merrick | Negative |
| Pterophoridae | Plume moth | Merrick | Negative |
| Diplopoda | Millipede | Merrick | Positive |
| Boisea trivittata | Boxelder bug | Merrick | Negative |
| Teleogryllus commodus | Black field cricket | Boone | Negative |
| Dermaptera | Earwig | Merrick | Negative |
| Chilopoda | Centipede | Merrick | Negative |
| Badumna longinqua | Grey house spider | Polk | Negative |
| Odontotaenius disjunctus | Horned passalus beetle | Hamilton | Positive |
| Scudderia furcata | Fork-tailed bush katydid | Hall | Negative |
| Pholidoptera griseoaptera | Dark bush-cricket | Merrick | Negative |
| Tenodera aridifolia sinensis | Chinese mantis | Merrick | Negative |
| Spodoptera ornithogalli | Yellow-striped armyworm | Merrick | Negative |
| Haematopis grataria | Chickweed geometer | Merrick | Negative |
| Armadillidiidae | Pill bug | Merrick | Negative |
| Badumna insignis | Black house spider | Buffalo | Negative |
| Brachypnoea | Leaf beetle | Merrick | Negative |
| Manduca | Hawkmoth | Merrick | Negative |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Buhlke, E.G.; Hobbs, A.M.; Rajput, S.; Rokusek, B.; Carlson, D.J.; Gillan, C.; Carlson, K.A. Characterization of Cross-Species Transmission of Drosophila melanogaster Nora Virus. Life 2022, 12, 1913. https://doi.org/10.3390/life12111913
AMA Style
Buhlke EG, Hobbs AM, Rajput S, Rokusek B, Carlson DJ, Gillan C, Carlson KA. Characterization of Cross-Species Transmission of Drosophila melanogaster Nora Virus. Life. 2022; 12(11):1913. https://doi.org/10.3390/life12111913
Chicago/Turabian StyleBuhlke, Ella G., Alexis M. Hobbs, Sunanda Rajput, Blase Rokusek, Darby J. Carlson, Chelle Gillan, and Kimberly A. Carlson. 2022. "Characterization of Cross-Species Transmission of Drosophila melanogaster Nora Virus" Life 12, no. 11: 1913. https://doi.org/10.3390/life12111913
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.