
Origins and Early Evolution of the tRNA Molecule

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
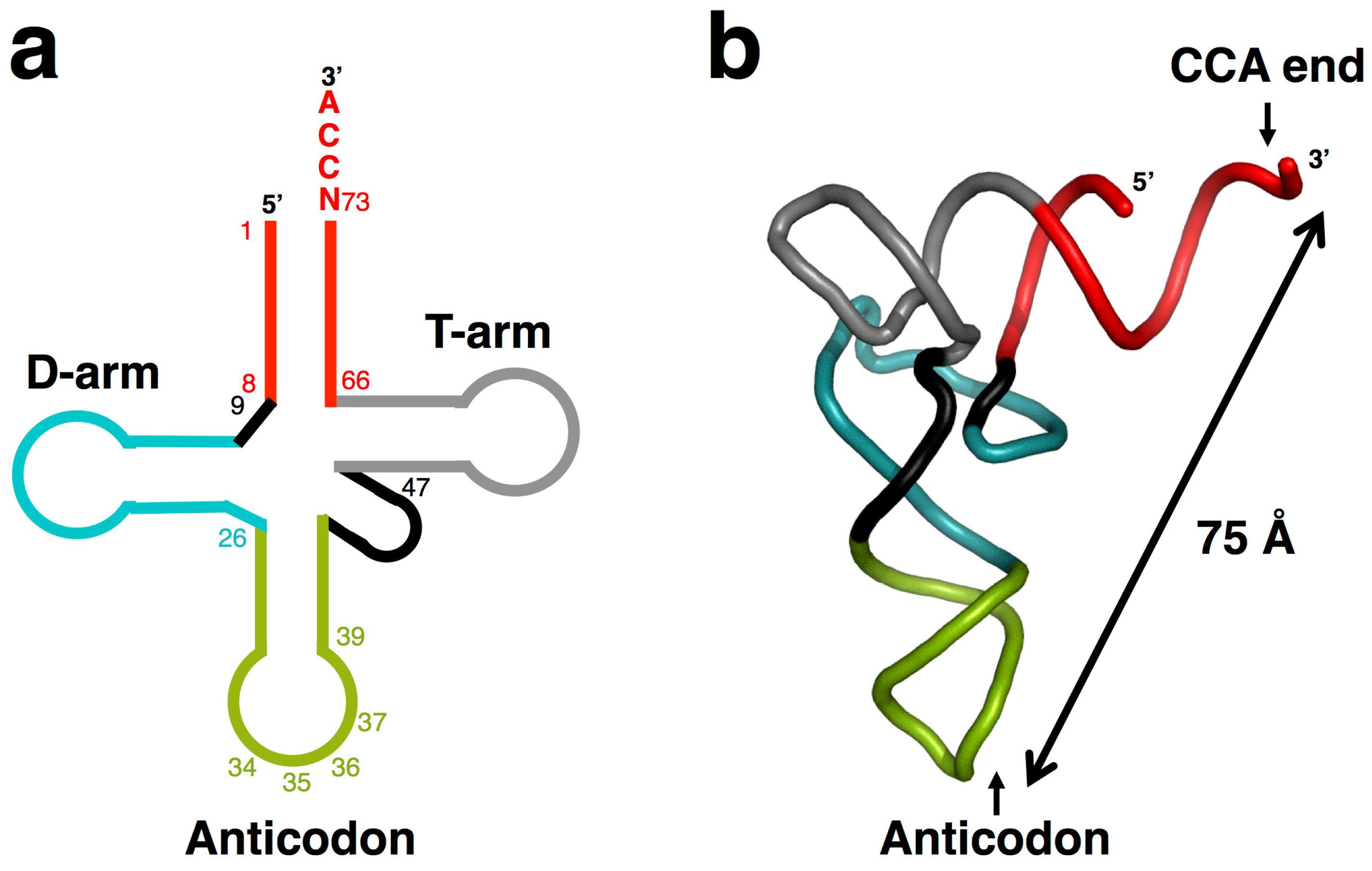
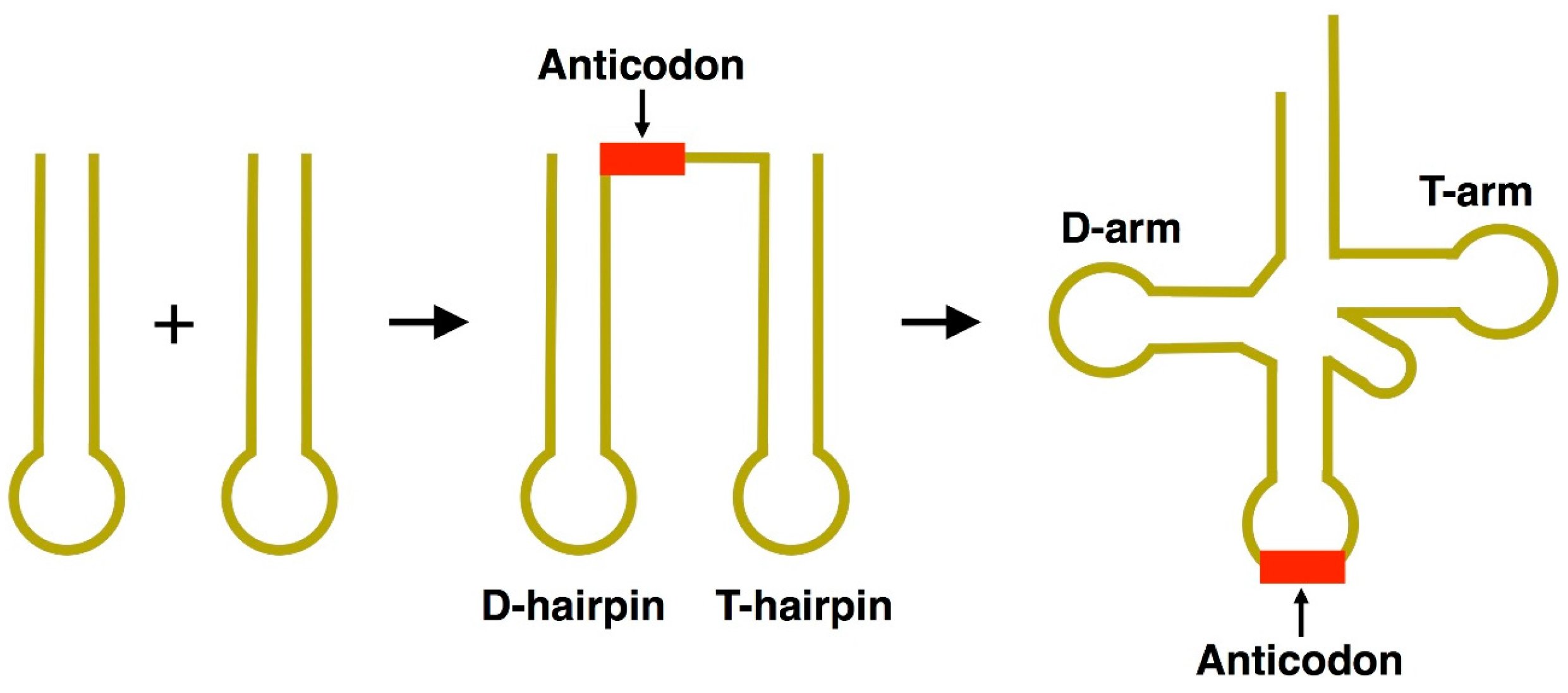
:1. Origins of tRNA

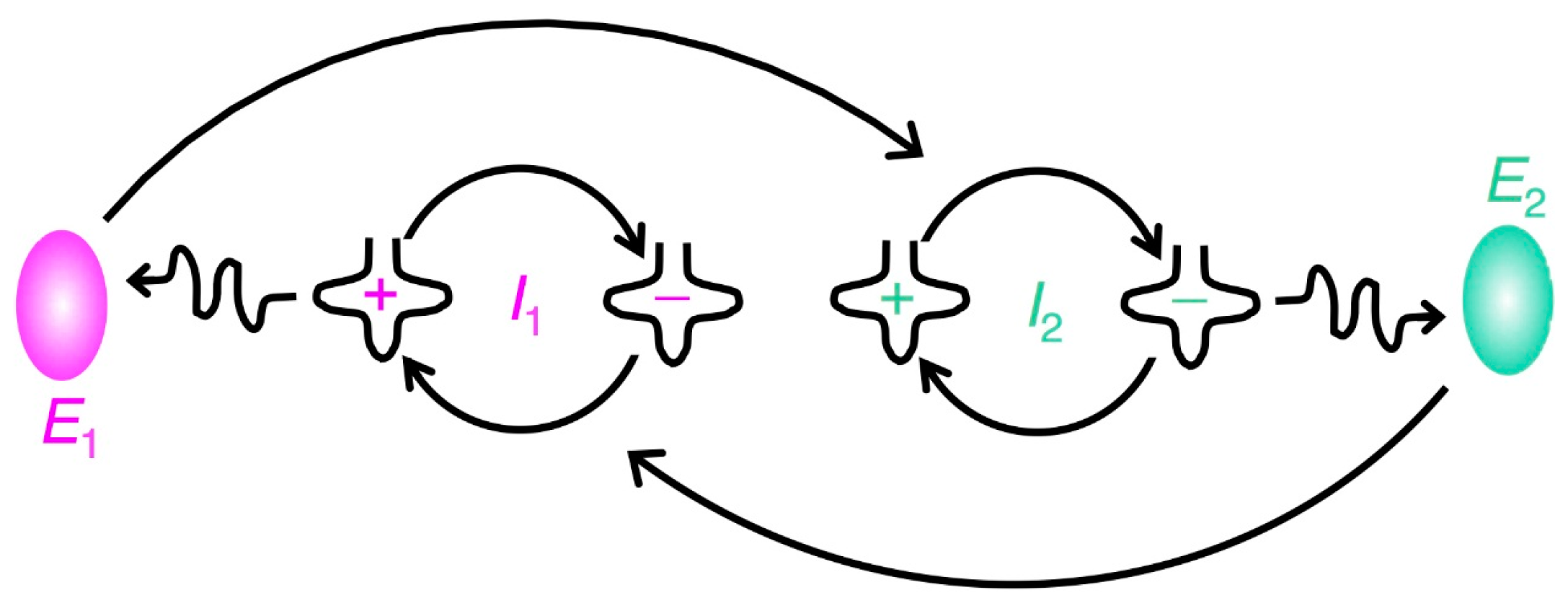
2. Emergence of a Self-Replicating Unit

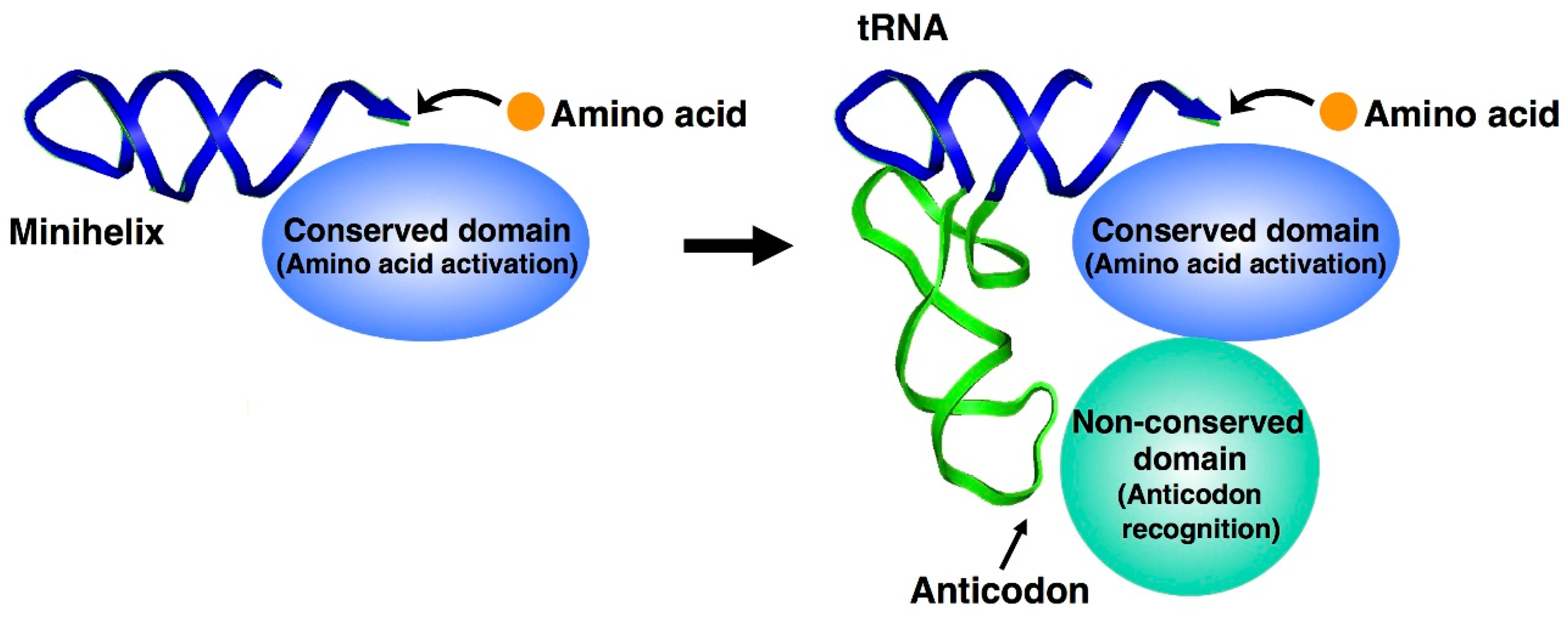
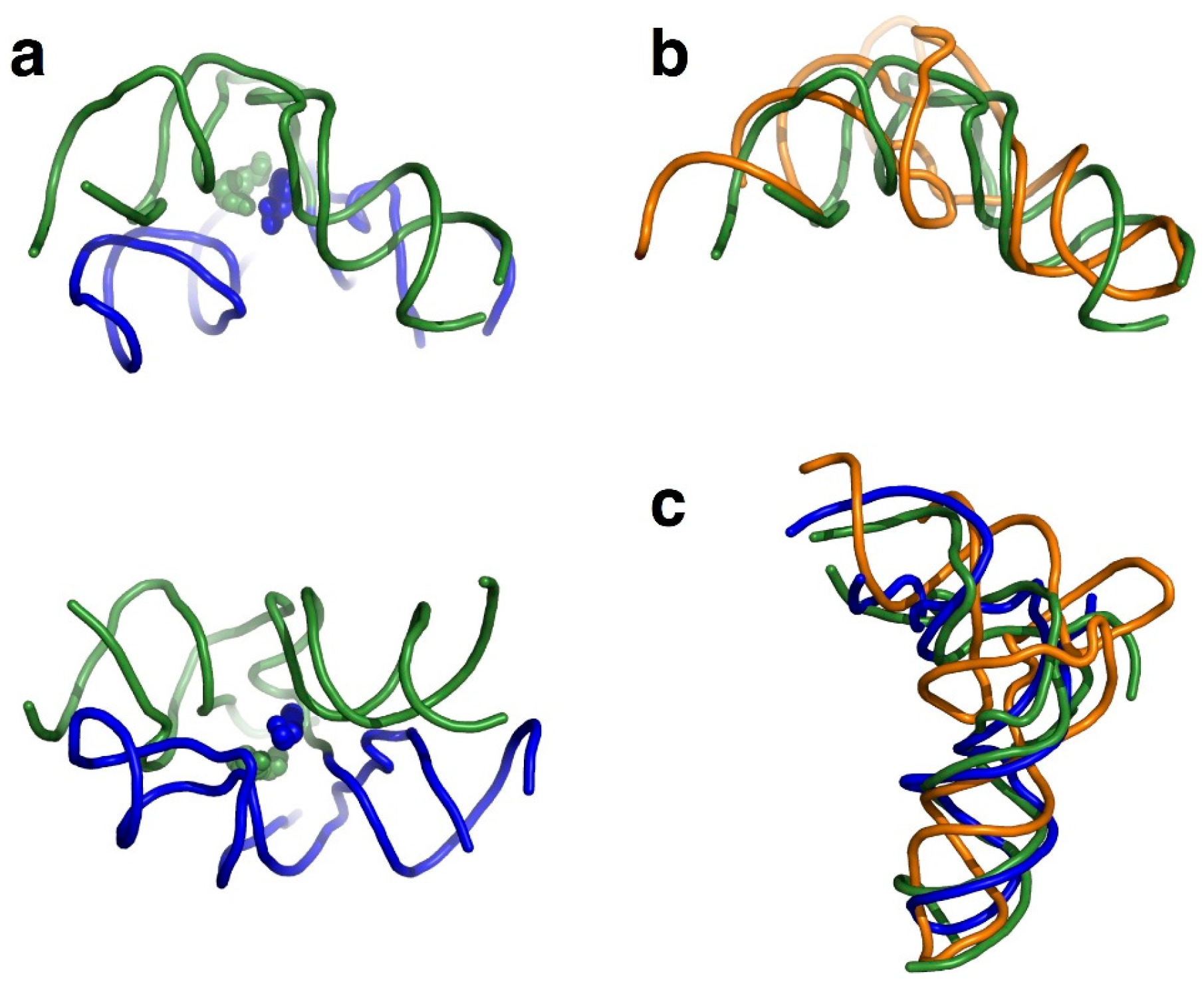
3. Features of tRNA Aminoacylation and Evolutionary Significance

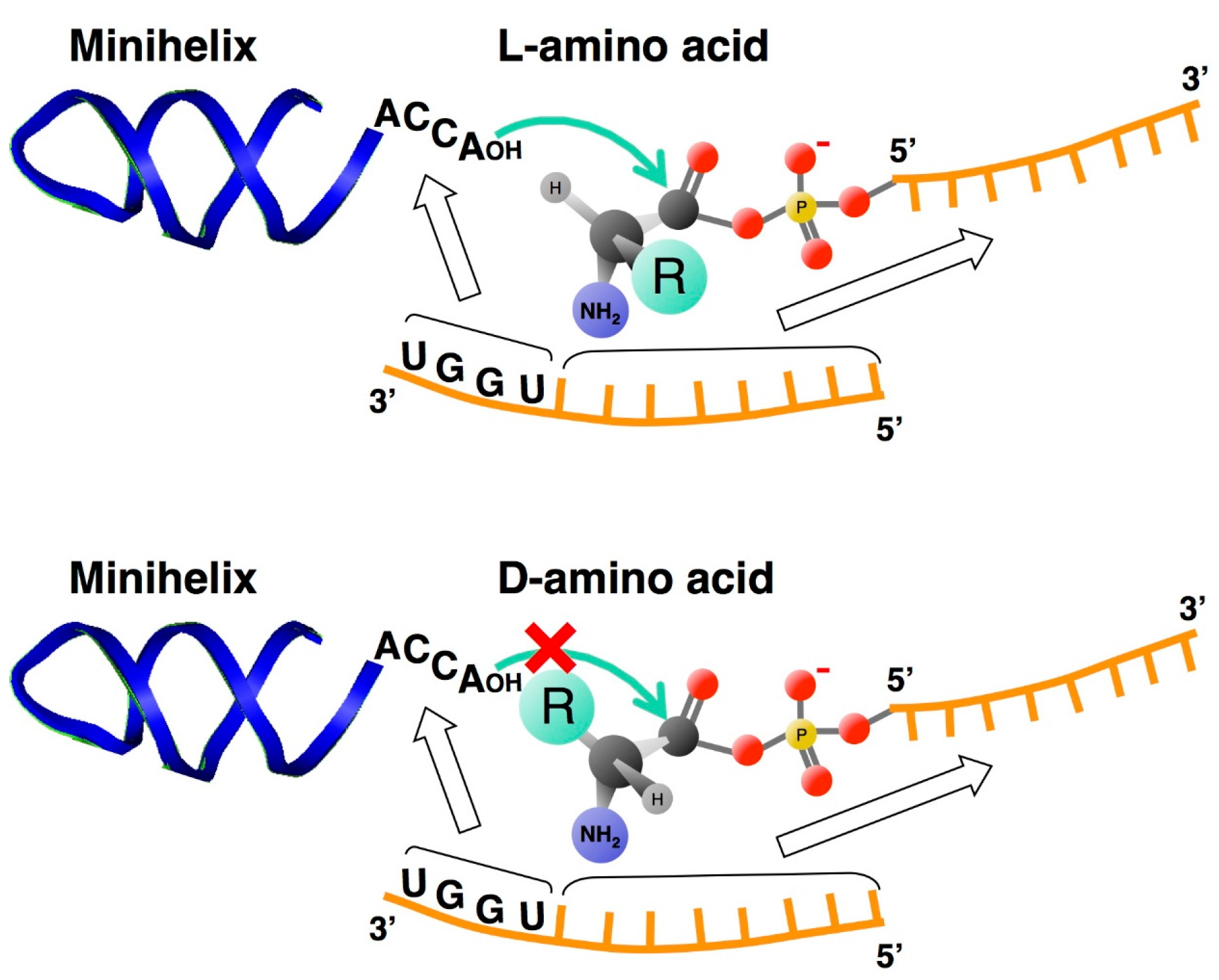
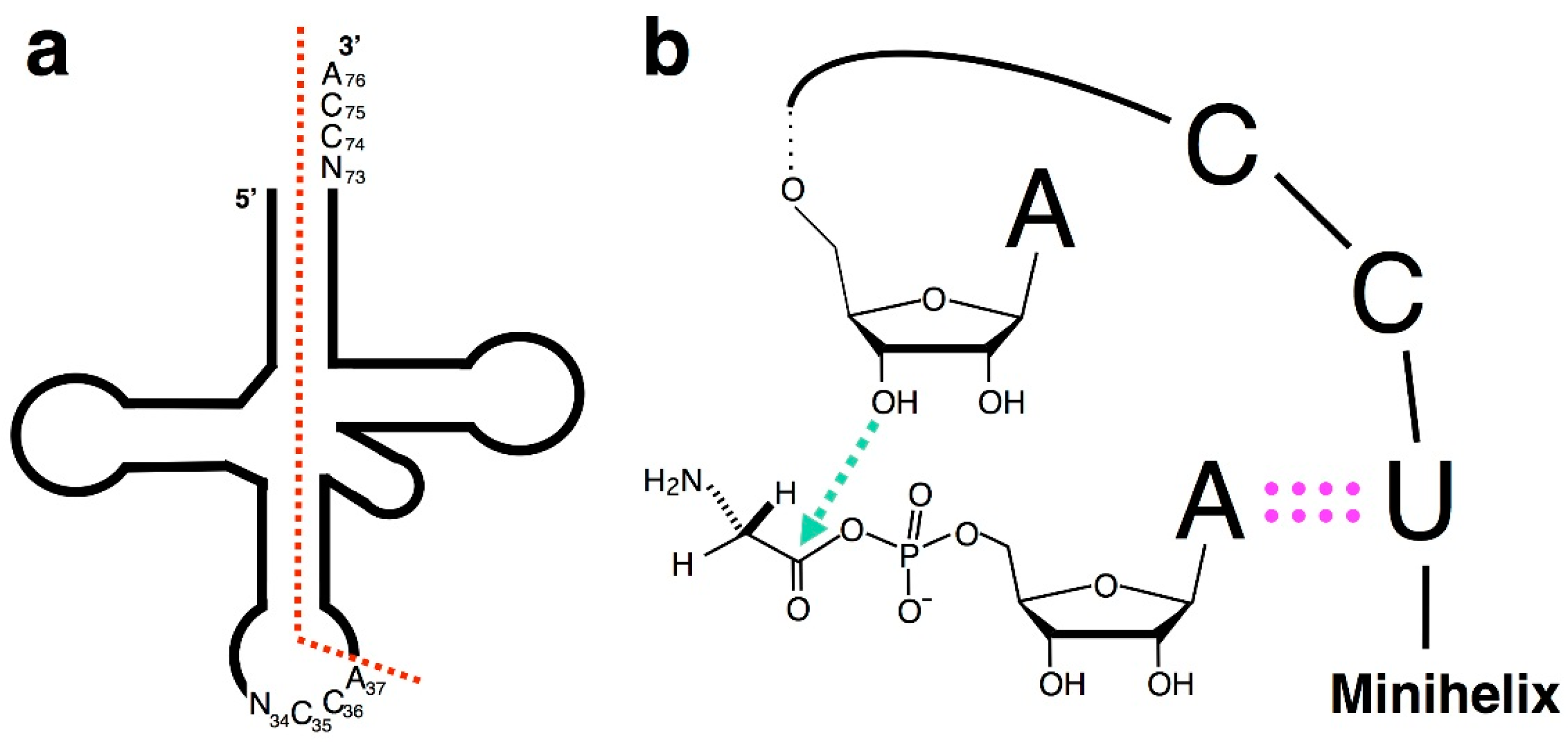
4. Chiral-Selective Primordial tRNA Aminoacylation

5. The CCA Sequence of tRNAs and the Origin of the Genetic Code


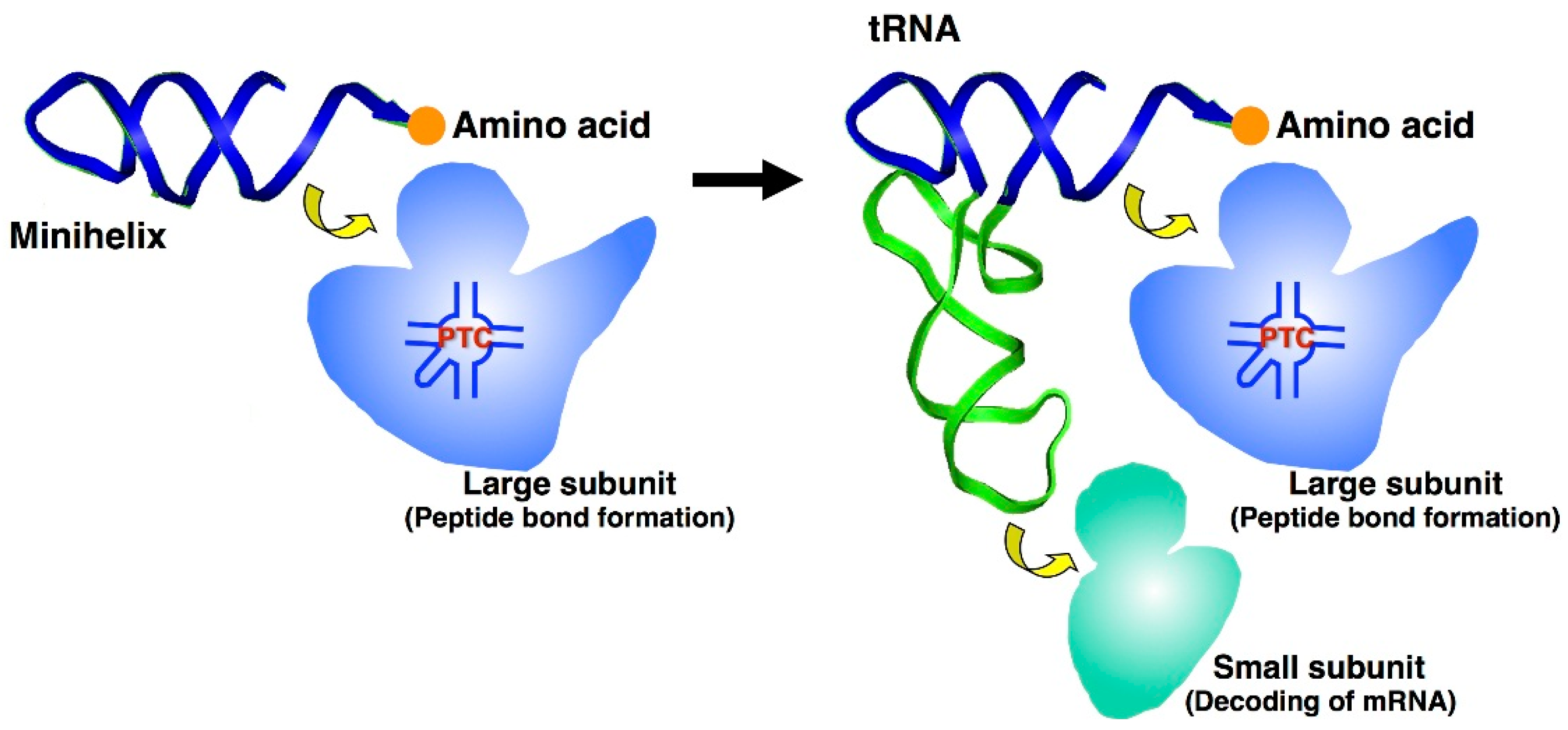
6. Origins of the Ribosome PTC and tRNA

7. Evolution of the Genetic Code and tRNA

8. Concluding Remarks
Acknowledgments
Conflicts of Interest
References
- Crick, F.H.C. The genetic code—Yesterday, today, and tomorrow. Cold Spring Harb. Quant. Biol. 1966, 31, 1–9. [Google Scholar] [CrossRef]
- Hoagland, M.B.; Stephenson, M.L.; Scott, J.F.; Hecht, L.I.; Zamecnik, P.C. A soluble ribonucleic acid intermediate in protein synthesis. J. Biol. Chem. 1958, 231, 241–257. [Google Scholar] [PubMed]
- Crick, F.H.C. On degenerate templates and the adapter hypothesis. 1955. Available online: https://profiles.nlm.nih.gov/ps/access/SCBBGF.pdf (accessed on 27 November 2015). [Google Scholar]
- Couzon, J. Small RNAs make big splash. Science 2002, 298, 2296–2297. [Google Scholar] [CrossRef] [PubMed]
- Jühling, F.; Mörl, M.; Hartmann, R.K.; Sprinzl, M.; Stadler, P.F.; Pütz, J. tRNAdb 2009: Compilation of tRNA sequences and tRNA genes. Nucleic Acids Res. 2009, 37, D159–D162. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.H.; Suddath, F.L.; Quigley, G.J.; McPherson, A.; Sussman, J.L.; Wang, A.H.; Seeman, N.C.; Rich, A. Three-dimensional tertiary structure of yeast phenylalanine transfer RNA. Science 1974, 185, 435–440. [Google Scholar] [CrossRef] [PubMed]
- Robertus, J.D.; Ladner, J.E.; Finch, J.T.; Rhodes, D.; Brown, R.S.; Clark, B.F.; Klug, A. Structure of yeast phenylalanine tRNA at 3 Å resolution. Nature 1974, 250, 546–551. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, W. Origin of life: The RNA world. Nature 1986, 319, 618. [Google Scholar] [CrossRef]
- Kruger, K.; Grabowski, P.J.; Zaug, A.J.; Sands, J.; Gottschling, D.E.; Cech, T.R. Self-splicing RNA: Autoexcision and autocyclization of the ribosomal RNA intervening sequence of Tetrahymena. Cell 1982, 31, 147–157. [Google Scholar] [CrossRef]
- Guerrier-Takada, C.; Gardiner, K.; Marsh, T.; Pace, N.; Altman, S. The RNA moiety of ribonuclease P is the catalytic subunit of the enzyme. Cell 1983, 35, 849–857. [Google Scholar] [CrossRef]
- Eigen, M. Selforganization of matter and the evolution of biological macromolecules. Naturwissenschaften 1971, 58, 465–523. [Google Scholar] [CrossRef] [PubMed]
- Eigen, M.; Schuster, P. Hypercycle. A principle of natural self-organization. Part A: Emergence of the hypercycle. Naturwissenschaften 1977, 64, 541–565. [Google Scholar] [CrossRef] [PubMed]
- Schimmel, P.; Giegé, R.; Moras, D.; Yokoyama, S. An operational RNA code for amino acids and possible relationship to genetic code. Proc. Natl. Acad. Sci. USA 1993, 90, 8763–8768. [Google Scholar] [CrossRef] [PubMed]
- Schimmel, P.; Ribas de Pouplana, L. Transfer RNA: From minihelix to genetic code. Cell 1995, 81, 983–986. [Google Scholar] [CrossRef]
- Francklyn, C.; Shi, J.-P.; Schimmel, P. Overlapping nucleotide determinants for specific aminoacylation of RNA microhelices. Science 1992, 255, 1121–1125. [Google Scholar] [CrossRef] [PubMed]
- Frugier, M.; Florentz, C.; Giegé, R. Efficient aminoacylation of resected RNA helices by class II aspartyl-tRNA synthetase dependent on a single nucleotide. EMBO J. 1994, 13, 2218–2226. [Google Scholar] [PubMed]
- Saks, M.E.; Sampson, J.R. Variant minihelix RNAs reveal sequence-specific recognition of the helical tRNASer acceptor stem by E. coli seryl-tRNA synthetase. EMBO J. 1996, 15, 2843–2849. [Google Scholar] [PubMed]
- Schimmel, P. Aminoacyl tRNA synthetases: General scheme of structure-function relationships in the polypeptides and recognition of transfer RNAs. Annu. Rev. Biochem. 1987, 56, 125–158. [Google Scholar] [CrossRef] [PubMed]
- Eriani, G.; Delarue, M.; Poch, O.; Gangloff, J.; Moras, D. Partition of tRNA synthetases into two classes based on mutually exclusive sets of sequence motifs. Nature 1990, 347, 203–206. [Google Scholar] [CrossRef] [PubMed]
- Arnez, J.G.; Moras, D. Structural and functional considerations of the aminoacylation reaction. Trends Biochem. Sci. 1997, 22, 211–216. [Google Scholar] [CrossRef]
- Shimizu, M.; Asahara, H.; Tamura, K.; Hasegawa, T.; Himeno, H. The role of anticodon bases and the discriminator nucleotide in the recognition of some E. coli tRNAs by their aminoacyl-tRNA synthetases. J. Mol. Evol. 1992, 35, 436–443. [Google Scholar] [CrossRef] [PubMed]
- Moras, D. Structural and functional relationships between aminoacyl-tRNA synthetases. Trends Biochem. Sci. 1992, 17, 159–164. [Google Scholar] [CrossRef]
- Ribas de Pouplana, L.; Schimmel, P. Aminoacyl-tRNA synthetases: Potential markers of genetic code development. Trends Biochem. Sci. 2001, 26, 591–596. [Google Scholar] [CrossRef]
- Ribas de Pouplana, L.; Schimmel, P. Two classes of tRNA synthetases suggested by sterically compatible dockings on tRNA acceptor stem. Cell 2001, 104, 191–193. [Google Scholar] [CrossRef]
- Rodin, S.N.; Ohno, S. Four primordial modes of tRNA-synthetase recognition, determined by the (G,C) operational code. Proc. Natl. Acad. Sci. USA 1997, 94, 5183–5188. [Google Scholar] [CrossRef] [PubMed]
- Rodin, A.S.; Szathmáry, E.; Rodin, S.N. One ancestor for two codes viewed from the perspective of two complementary modes of tRNA aminoacylation. Biol. Direct 2009, 4, 4. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Rodriguez, L.; Erdogan, O.; Jimenez-Rodriguez, M.; Gonzalez-Rivera, K.; Williams, T.; Li, L.; Weinreb, V.; Collier, M.; Chandrasekaran, S.N.; Ambroggio, X.; et al. Functional Class I and II amino acid-activating enzymes can be coded by opposite strands of the same gene. J. Biol. Chem. 2015, 290, 19710–19725. [Google Scholar] [CrossRef] [PubMed]
- Carpenter, F.H. The free energy in hydrolytic reactions: The non-ionized compound convention. J. Am. Chem. Soc. 1960, 82, 1111–1122. [Google Scholar] [CrossRef]
- Paecht-Horowitz, M.; Katchalsky, A. Synthesis of amino acyl-adenylates under prebiotic conditions. J. Mol. Evol. 1973, 2, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Schimmel, P. Chiral-selective aminoacylation of an RNA minihelix. Science 2004, 305, 1253. [Google Scholar] [CrossRef] [PubMed]
- Bürgi, H.B.; Dunitz, J.D.; Lehn, J.M.; Wipff, G. Stereochemistry of reaction paths at carbonyl centre. Tetrahedron 1974, 30, 1563–1572. [Google Scholar]
- Tamura, K.; Schimmel, P.R. Chiral-selective aminoacylation of an RNA minihelix: Mechanistic features and chiral suppression. Proc. Natl. Acad. Sci. USA 2006, 103, 13750–13752. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K. Origin of amino acid homochirality: Relationship with the RNA world and origin of tRNA aminoacylation. BioSystems 2008, 92, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K. Molecular handedness of life: Significance of RNA aminoacylation. J. Biosci. 2009, 34, 991–994. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K. Molecular basis for chiral selection in RNA aminoacylation. Int. J. Mol. Sci. 2011, 12, 4745–4757. [Google Scholar] [CrossRef] [PubMed]
- Bolli, M.; Micura, R.; Eschenmoser, A. Pyranosyl-RNA: Chiroselective self-assembly of base sequences by ligative oligomerization of tetranucleotide-2′,3′-cyclophosphates (with a commentary concerning the origin of biomolecular homochirality). Chem. Biol. 1997, 4, 309–320. [Google Scholar] [CrossRef]
- Weiner, A.M.; Maizels, N. tRNA-like structures tag the 3′ ends of genomic RNA molecules for replication: Implications for the origin of protein synthesis. Proc. Natl. Acad. Sci. USA 1987, 84, 7383–7387. [Google Scholar] [CrossRef] [PubMed]
- Akins, R.A.; Kelley, R.L.; Lambowitz, A.M. Characterization of mutant mitochondrial plasmids of Neurospora spp. that have incorporated tRNAs by reverse transcription. Mol. Cell. Biol. 1989, 9, 678–691. [Google Scholar] [CrossRef] [PubMed]
- Saville, B.J.; Collins, R.A. A site-specific self-cleavage reaction performed by a novel RNA in Neurospora mitochondria. Cell 1990, 61, 685–696. [Google Scholar] [CrossRef]
- Blackburn, E.H. Structure and function of telomeres. Nature 1991, 350, 569–573. [Google Scholar] [CrossRef] [PubMed]
- Freier, S.M.; Alkema, D.; Sinclair, A.; Neilson, T.; Turner, D.H. Contributions of dangling end stacking and terminal base-pair formation to the stabilities of XGGCCp, XCCGGp, XGGCCYp, and XCCGGYp helixes. Biochemistry 1985, 24, 4533–4539. [Google Scholar] [CrossRef] [PubMed]
- SantaLucia, J., Jr.; Kierzek, R.; Turner, D.H. Effects of GA mismatches on the structure and thermodynamics of RNA internal loops. Biochemistry 1990, 29, 8813–8819. [Google Scholar] [CrossRef] [PubMed]
- Limmer, S.; Hofmann, H.P.; Ott, G.; Sprinzl, M. The 3′-terminal end (NCCA) of tRNA determines the structure and stability of the aminoacyl acceptor stem. Proc. Natl. Acad. Sci. USA 1993, 90, 6199–6202. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K. Ribosome evolution: Emergence of peptide synthesis machinery. J. Biosci. 2011, 36, 921–928. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K. RNA evolution conjectured from tRNA and riboswitches. Hypothesis 2015, 13, e3. [Google Scholar] [CrossRef]
- Tamura, K. The role of the CCA sequence of tRNA in the peptidyl transfer reaction. FEBS Lett. 1994, 353, 173–176. [Google Scholar] [CrossRef]
- Nissen, P.; Hansen, J.; Ban, N.; Moore, P.B.; Steitz, T.A. The structural basis of ribosome activity in peptide bond synthesis. Science 2000, 289, 920–930. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Nameki, N.; Hasegawa, T.; Shimizu, M.; Himeno, H. Role of the CCA terminal sequence of tRNAVal in aminoacylation with valyl-tRNA synthetase. J. Biol. Chem. 1994, 269, 22173–22177. [Google Scholar] [PubMed]
- Bernhardt, H.S.; Tate, W.P. Evidence from glycine transfer RNA of a frozen accident at the dawn of the genetic code. Biol. Direct 2008, 3, 53. [Google Scholar] [CrossRef] [PubMed]
- Bernhardt, H.S.; Tate, W.P. The transition from noncoded to coded protein synthesis: Did coding mRNAs arise from stability-enhancing binding partners to tRNA? Biol. Direct 2010, 5, 16. [Google Scholar] [CrossRef] [PubMed]
- Puglisi, E.V.; Puglisi, J.D.; Williamson, J.R.; RajBhandary, U.L. NMR analysis of tRNA acceptor stem microhelices: Discriminator base change affects tRNA conformation at the 3′ end. Proc. Natl. Acad. Sci. USA 1994, 91, 11467–11471. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K. Beyond the frozen accident: Glycine assignment in the genetic code. J. Mol. Evol. 2015, 81, 69–71. [Google Scholar] [CrossRef] [PubMed]
- Kvenvolden, K.; Lawless, J.; Pering, K.; Peterson, E.; Flores, J.; Ponnamperuma, C.; Kaplan, I.R.; Moore, C. Evidence for extraterrestrial amino-acids and hydrocarbons in the Murchison meteorite. Nature 1970, 228, 923–926. [Google Scholar] [CrossRef] [PubMed]
- Miller, S.L. A production of amino acids under possible primitive earth conditions. Science 1953, 117, 528–529. [Google Scholar] [CrossRef] [PubMed]
- Wong, J.T. A co-evolution theory of the genetic code. Proc. Natl. Acad. Sci. USA 1975, 72, 1909–1912. [Google Scholar] [CrossRef] [PubMed]
- Wong, J.T. Coevolution theory of the genetic code at age thirty. BioEssays 2005, 27, 416–425. [Google Scholar] [CrossRef] [PubMed]
- Kagawa, Y.; Nojima, H.; Nukiwa, N.; Ishizuka, M.; Nakajima, T.; Yasuhara, T.; Tanaka, T.; Oshima, T. High guanine plus cytosine content in the third letter of codons of an extreme thermophile. DNA sequence of the isopropylmalate dehydrogenase of Thermus thermophilus. J. Biol. Chem. 1984, 259, 2956–2960. [Google Scholar] [PubMed]
- Vieille, C.; Zeikus, G.J. Hyperthermophilic enzymes: Sources, uses, and molecular mechanisms for thermostability. Microbiol. Mol. Biol. Rev. 2001, 65, 1–43. [Google Scholar] [CrossRef] [PubMed]
- Chakravarty, S.; Varadarajan, R. Elucidation of factors responsible for enhanced thermal stability of proteins: A structural genomics based study. Biochemistry 2002, 41, 8152–8161. [Google Scholar] [CrossRef] [PubMed]
- Ohnuma, M.; Ganbe, T.; Terui, Y.; Niitsu, M.; Sato, T.; Tanaka, N.; Tamakoshi, M.; Samejima, K.; Kumasaka, T.; Oshima, T. Crystal structures and enzymatic properties of a triamine/agmatine aminopropyltransferase from Thermus thermophiles. J. Mol. Biol. 2011, 408, 971–986. [Google Scholar] [CrossRef] [PubMed]
- Jaenicke, R.; Bohm, G. The stability of proteins in extreme environments. Curr. Opin. Struct. Biol. 1998, 8, 738–748. [Google Scholar] [CrossRef]
- Terasaka, N.; Hayashi, G.; Katoh, T.; Suga, H. An orthogonal ribosome-tRNA pair via engineering of the peptidyl transferase center. Nat. Chem. Biol. 2014, 10, 555–557. [Google Scholar] [CrossRef] [PubMed]
- Hopfield, J.J. Origin of the genetic code: A testable hypothesis based on tRNA structure, sequence, and kinetic proofreading. Proc. Natl. Acad. Sci. USA 1978, 75, 4334–4338. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, M. Molecular basis for the genetic code. J. Mol. Evol. 1982, 18, 297–303. [Google Scholar] [CrossRef] [PubMed]
- Lacey, J.C., Jr.; Mullins, D.W., Jr. Experimental studies related to the origin of the genetic code and the process of protein synthesis. Orig. Life 1983, 13, 3–42. [Google Scholar] [CrossRef] [PubMed]
- Yarus, M. Amino acids as RNA ligands: A direct-RNA-template theory for the code’s origin. J. Mol. Evol. 1998, 47, 109–117. [Google Scholar] [CrossRef] [PubMed]
- Fontecilla-Camps, J.C. The stereochemical basis of the genetic code and the (mostly) autotrophic origin of life. Life 2014, 4, 1013–1025. [Google Scholar] [CrossRef] [PubMed]
- Komatsu, R.; Sawada, R.; Umehara, T.; Tamura, K. Proline might have been the first amino acid in the primitive genetic code. J. Mol. Evol. 2014, 78, 310–312. [Google Scholar] [CrossRef] [PubMed]
- Ban, N.; Nissen, P.; Hansen, J.; Moore, P.B.; Steitz, T.A. The complete atomic structure of the large ribosomal subunit at 2.4 Å resolution. Science 2000, 289, 905–920. [Google Scholar] [CrossRef] [PubMed]
- Noller, H.F.; Hoffarth, V.; Zimniak, L. Unusual resistance of peptidyl transferase to protein extraction procedures. Science 1992, 256, 1416–1419. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Schimmel, P. Oligonucleotide-directed peptide synthesis in a ribosome- and ribozyme-free system. Proc. Natl. Acad. Sci. USA 2001, 98, 1393–1397. [Google Scholar] [CrossRef] [PubMed]
- Agmon, I.; Bashan, A.; Zarivach, R.; Yonath, A. Symmetry at the active site of the ribosome: Structural and functional implications. Biol. Chem. 2005, 386, 833–844. [Google Scholar] [CrossRef] [PubMed]
- Agmon, I. The dimeric proto-ribosome: Structural details and possible implications on the origin of life. Int. J. Mol. Sci. 2009, 10, 2921–2934. [Google Scholar] [CrossRef] [PubMed]
- Moore, P.B. Ribosomes. Curr. Opin. Struct. Biol. 1991, 1, 258–263. [Google Scholar] [CrossRef]
- Ito, K.; Ebihara, K.; Uno, M.; Nakamura, Y. Conserved motifs in prokaryotic and eukaryotic polypeptide release factors: tRNA-protein mimicry hypothesis. Proc. Natl. Acad. Sci. USA 1996, 93, 5443–5448. [Google Scholar] [CrossRef] [PubMed]
- Ito, K.; Uno, M.; Nakamura, Y. A tripeptide “anticodon” deciphers stop codons in messenger RNA. Nature 2000, 403, 680–684. [Google Scholar] [CrossRef] [PubMed]
- Rodin, S.; Rodin, A.; Ohno, S. The presence of codon-anticodon pairs in the acceptor stem of tRNAs. Proc. Natl. Acad. Sci. USA 1996, 93, 4537–4542. [Google Scholar] [CrossRef] [PubMed]
- Hou, Y.M.; Schimmel, P. A simple structural feature is a major determinant of the identity of a transfer RNA. Nature 1988, 333, 140–145. [Google Scholar] [CrossRef] [PubMed]
- McClain, W.H.; Foss, K. Changing the identity of a tRNA by introducing a G-U wobble pair near the 3′ acceptor end. Science 1988, 240, 793–796. [Google Scholar] [CrossRef] [PubMed]
- de Duve, C. Transfer RNAs: The second genetic code. Nature 1988, 333, 117–118. [Google Scholar] [CrossRef] [PubMed]
- Di Giulio, M. On the origin of the transfer RNA molecule. J. Theor. Biol. 1992, 159, 199–214. [Google Scholar] [CrossRef]
- Tanaka, T.; Kikuchi, Y. Origin of the cloverleaf shape of transfer RNA-the double-hairpin model: Implication for the role of tRNA intro and the long extra loop. Viva Orig. 2001, 29, 134–142. [Google Scholar]
- Nagaswamy, U.; Fox, G.E. RNA ligation and the origin of tRNA. Orig. Life Evol. Biosph. 2003, 33, 199–209. [Google Scholar] [CrossRef] [PubMed]
- Widmann, J.; Di Giulio, M.; Yarus, M.; Knight, R. tRNA creation by hairpin duplication. J. Mol. Evol. 2005, 61, 524–530. [Google Scholar] [CrossRef] [PubMed]
- Crick, F.H.C. The origin of the genetic code. J. Mol. Biol. 1968, 38, 367–379. [Google Scholar] [CrossRef]
- Bernhardt, H.S.; Patrick, W.M. Genetic code evolution started with the incorporation of glycine, followed by other small hydrophilic amino acids. J. Mol. Evol. 2014, 78, 307–309. [Google Scholar] [CrossRef] [PubMed]
- Orgel, L.E. β-Turns and the evolution of protein synthesis. In The organization and Expression of the Eukaryotic Genome, Proceedings of the International Symposium, Teheran, Iran, 3–6 May 1976; Bradbury, E.M., Javaherian, K., Eds.; Academic Press: London, UK, 1997; pp. 499–504. [Google Scholar]
- Jurka, J.; Smith, T.F. β-Turn-driven early evolution: The genetic code and biosynthetic pathways. J. Mol. Evol. 1987, 25, 15–19. [Google Scholar] [CrossRef] [PubMed]
- Wilmot, C.M.; Thornton, J.M. Analysis and prediction of the different types of β-turn in proteins. J. Mol. Biol. 1988, 203, 221–232. [Google Scholar] [CrossRef]
- Huppert, J.L.; Bugaut, A.; Kumari, S.; Balasubramanian, S. G-Quadruplexes: The beginning and end of UTRs. Nucleic Acids Res. 2008, 36, 6260–6268. [Google Scholar] [CrossRef] [PubMed]
- Umehara, T.; Kitagawa, T.; Nakazawa, Y.; Yoshino, H.; Nemoto, R.; Tamura, K. RNA tetraplex as a primordial peptide synthesis scaffold. BioSystems 2012, 109, 145–150. [Google Scholar] [CrossRef] [PubMed]
- Pauling, L. The probability of errors in the process of synthesis of protein molecules. In Arbeiten aus dem Gebiet der Naturstoffchemie, Festschrift Arthur Stroll; Birkhäuser-Verlag: Basel, Switzerland, 1957; pp. 597–602. [Google Scholar]
- Freist, W.; Pardowitz, I.; Cramer, F. Isoleucyl-tRNA synthetase from bakers’ yeast: Multistep proofreading in discrimination between isoleucine and valine with modulated accuracy, a scheme for molecular recognition by energy dissipation. Biochemistry 1985, 24, 7014–7023. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tamura, K. Origins and Early Evolution of the tRNA Molecule. Life 2015, 5, 1687-1699. https://doi.org/10.3390/life5041687
Tamura K. Origins and Early Evolution of the tRNA Molecule. Life. 2015; 5(4):1687-1699. https://doi.org/10.3390/life5041687
Chicago/Turabian StyleTamura, Koji. 2015. "Origins and Early Evolution of the tRNA Molecule" Life 5, no. 4: 1687-1699. https://doi.org/10.3390/life5041687