Comparison between Effects of Retroactivity and Resource Competition upon Change in Downstream Reporter Genes of Synthetic Genetic Circuits

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Mathematical Modeling of Chemical Reactions, Simulation and Stability Analysis
2.2. Bacterial Strains and Construction of Plasmids
2.3. Reporter Assay
2.4. Microscopy Experiments
2.5. Imaging Process
3. Results and Discussion
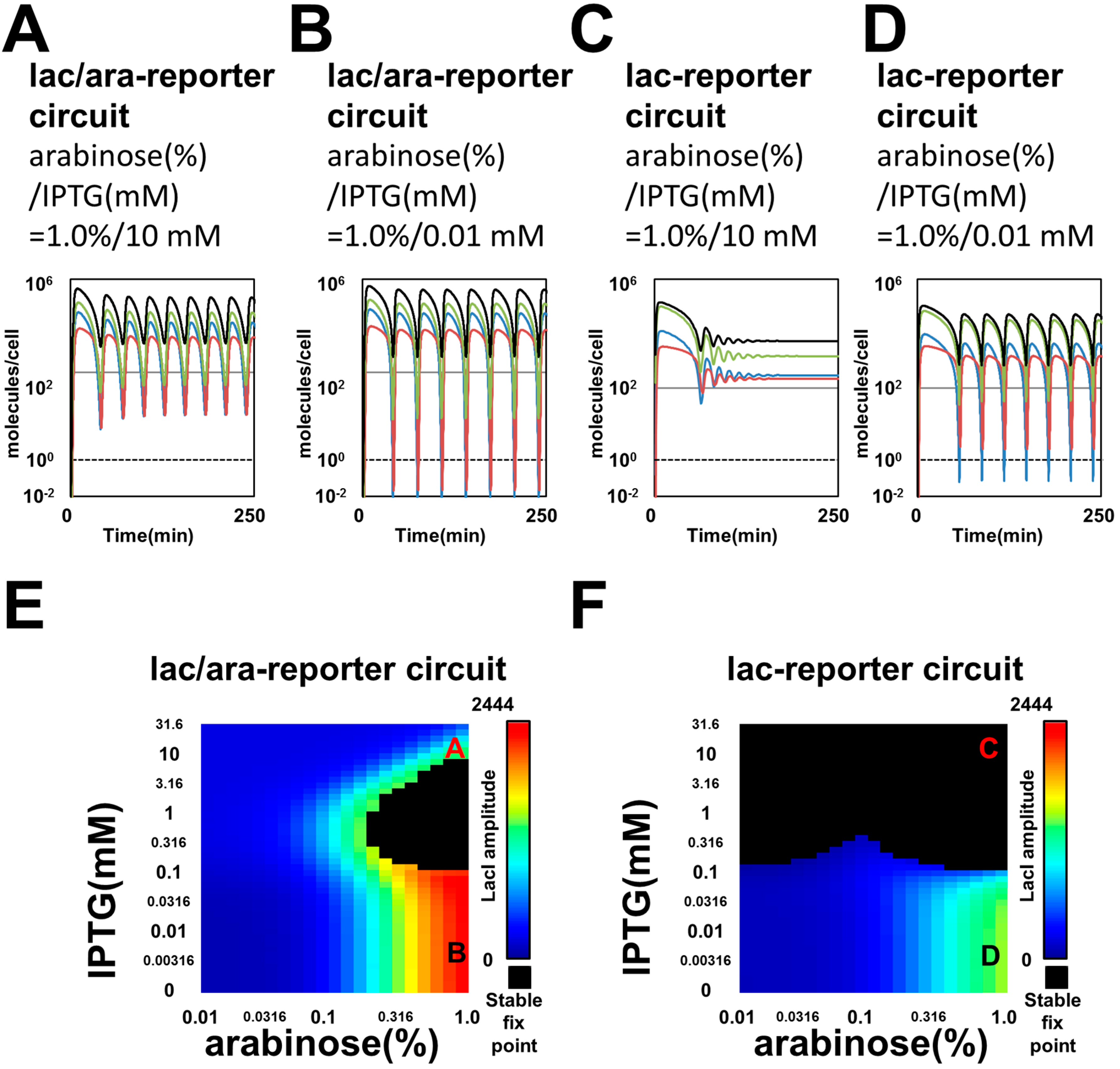
3.1. Mathematical Modeling and Simulation Suggested Perturbation of Oscillation Dynamics by Competitions for Regulatory Protein and Protease
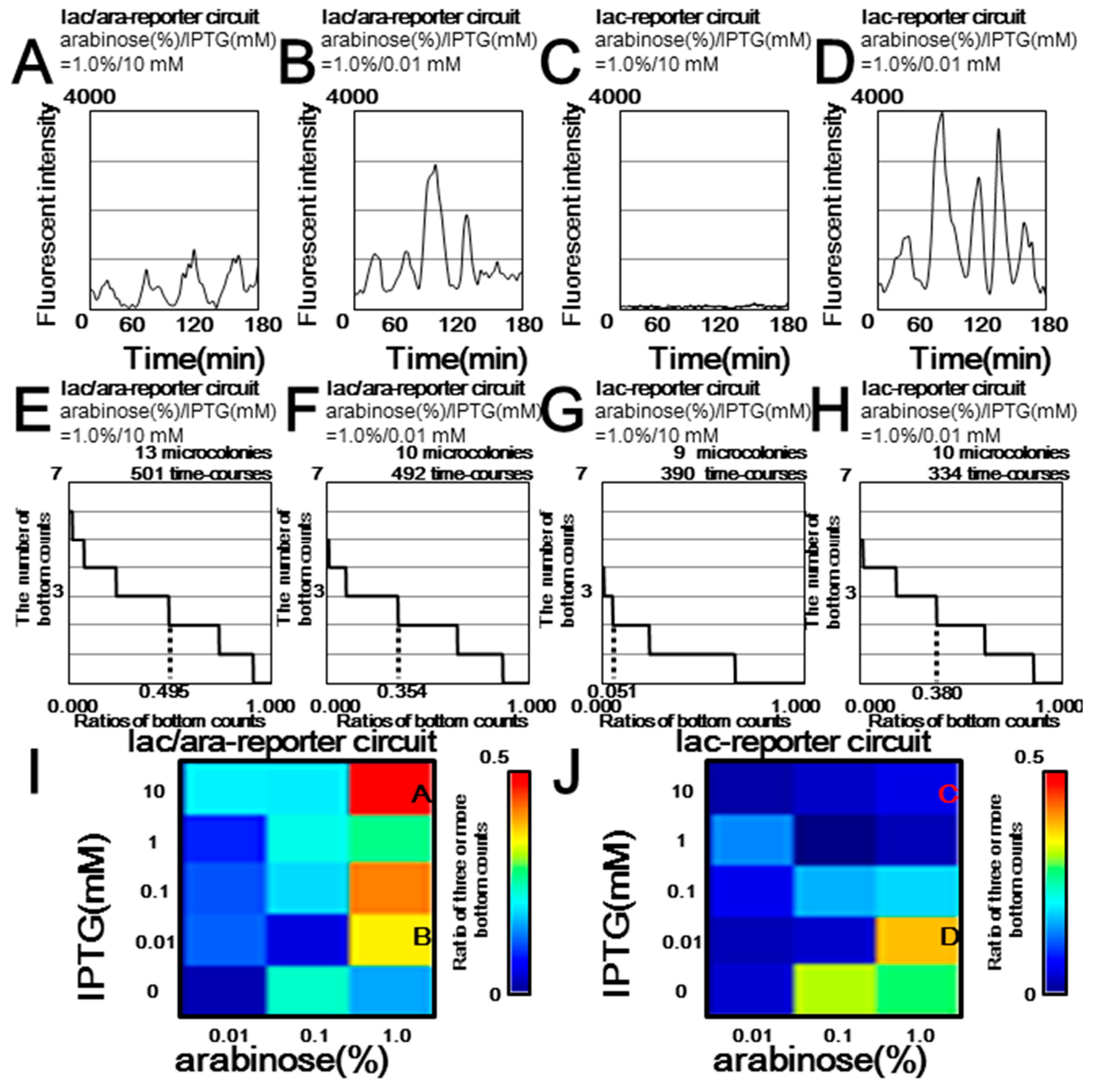
3.2. Microscopy Experiments Showed that Downstream Reporter Genes Changed the Oscillation Dynamics of Upstream Regulatory Genes
3.3. Comparison of Retroactivity and Protease Sharing with Upstream Gene Expression Dynamics Even for the Same Set of Protein-Binding Sites in a Downstream Component Having Differences in the Allocation
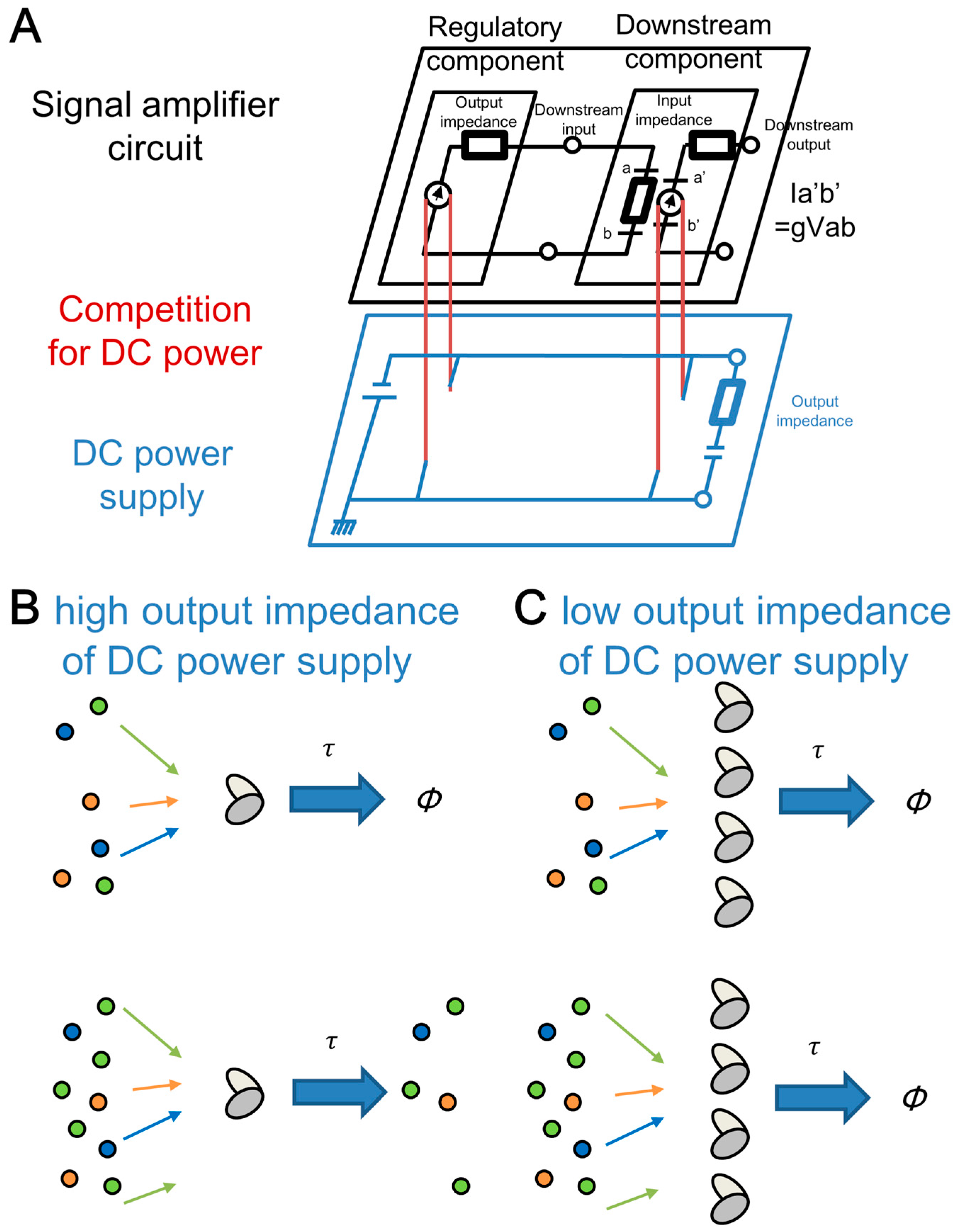
3.4. Relationship of the Effects of the Molecular Competitions between Electric Circuit and Genetic Circuit
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter Name | Parameter Value | Description | Units | Source |
|---|---|---|---|---|
| ba, br, bd1 | 0.36 | transcription rate of lac/ara promoter | min−1 | [11] |
| bd2 | 7.2 (=αbd1) | transcription rate of lac promoter | min−1 | This study |
| α | 20 | coefficient of active promoter | [11] | |
| k-a, k-r | 1.8 | Protein and its protein-binding site dissociation rate | min−1 | [11] |
| ta, tr, td | 90 | translation rate | min−1 | [11] |
| da, dr, dd | 0.54 | mRNA degradation rate | min−1 | [11] |
| kfa, kfr, kfd | 0.9 | folding rate | min−1 | [11] |
| kda, kdr, kt | 0.018 | multimerization rate | min−1molecules−1 | [11] |
| k-da, k-dr, k-t | 0.00018 | multimer dissociation rate | min−1 | [11] |
| kl | 1.08 | loop forming rate | min−1 | [31] |
| k-l | 0.036 | loop unforming rate | min−1 | [31] |
| kul | 0.90 | loop dissociation rate | min−1 | [34] |
| γ | 1080 | maximum degradation rate | molecules · min−1 | [11] |
| ce | 0.1 | dissociation constant | molecules | [11] |
| λ | 2.5 | coefficient of degradation rate of AraC and downstream free molecule | [11] | |
| ε | 0.2 | coefficient of degradation rate in loop promoter | [11] | |
| ωa, ωr, ωd | 1 | coefficient of tagged degradation rate | This study | |
| Na | 50 | copy number of activator gene | [11,40] | |
| Nr | 25 | copy number of repressor gene | [11,40] | |
| Nd | 50 | copy number of downstream gene | [12,40] | |
| Ndecoy | 50 | copy number of downstream decoy site | [40] | |
| 0.2 | maximum affinity of LacI for the protein-binding site | molecules−1 | [11] | |
| 0.01 | minimum affinity of LacI for the protein-binding site | molecules−1 | [11] | |
| [IPTG] | 0–10 | IPTG concentration | mM | [11] |
| kr1 | 0.035 | correction constant of IPTG | mM | [11] |
| kr2 | 1.8 | correction constant of arabinose | mM | [11] |
| b1 | 2 | Hill coefficient of inhibition by LacI | [11] | |
| 1 | maximum affinity of AraC for the promoter | molecules−1 | [11] | |
| 0 | minimum affinity of AraC for the promoter | molecules−1 | [11] | |
| [ara] | 0–2 | arabinose concentration | % (w/v) | [11] |
| ka1 | 2.5 | correction constant of arabinose | % (w/v) | [11] |
| c1 | 2 | Hill coefficient of activation by AraC | [11] | |
| b2 | 2 | Hill coefficient of inhibition by LacI | [11] |
Appendix B. Detailed Construction of Plasmids
| Strain or Plasmid | Relevant Genotype and/or Property | Source |
|---|---|---|
| E. coli strains | ||
| JS006 | MG1655 ∆ araC ∆ lacI KanS | [11] |
| JS00611 | JS006 strain with pJS167 and pJS169 plasmids | [11] |
| JSDT10611 | JS006 strain with pJSDT267 and pJS169 plasmids | This study |
| Plasmids | ||
| pJS167 | ara-lac oscillator activator/Plac/ara-gfp vector, ColE1, Kanr | [11] |
| pJSDT168 | PBAD/ara + Plac/ara-gfp vector, p15A, Cmr | This study |
| pJS169 | ara-lac oscillator repressor vector, p15A, Ampr | [11] |
| pJSDT170 | promoter deleted PBAD/ara + Plac/ara-gfp vector, p15A, Cmr | This study |
| pJSDT171 | promoter deleted PBAD/ara + Plac/ara-gfp vector, p15A, Kanr | This study |
| pPlac-gfp_partial | lac promoter and gfp N-terminal partial region in pIDTSMART vector, pUC, Ampr | This study |
| pPlac-gfp | Plac-gfp in pIDTSMART vector, pUC, Ampr | This study |
| pJSDT267 | ara-lac oscillator activator/Plac-gfp vector, ColE1, Kanr | This study |
| pJSDT268 | PBAD/ara + Plac-gfp vector, p15A, Cmr | This study |
| pJSDT270 | promoter deleted PBAD/ara + Plac-gfp vector, p15A, Cmr | This study |
| pJSDT271 | promoter deleted PBAD/ara + Plac-gfp vector, p15A, Kanr | This study |
| PBAD/ara | BioBrick BBa_K808000 on pSB3C5 BioBrick vector, p15A, Cmr | [48,49,50] |
| pSB-GFP | promoter-less GFP on pSB3K3 BioBrick vector, p15A, Kanr | [47] |
| Ptet-gfp | BioBrick BBa_I13522 on pSB6A1 BioBrick vector, pMB1, Ampr | [48,49] |
References
- Montminy, M.R.; Sevarino, K.A.; Wagner, J.A.; Mandel, G.; Goodman, R.H. Identification of a cyclic-amp-responsive element within the rat somatostatin gene. Proc. Natl. Acad. Sci. USA 1986, 83, 6682–6686. [Google Scholar] [CrossRef]
- Selden, R.F.; Howie, K.B.; Rowe, M.E.; Goodman, H.M.; Moore, D.D. Human growth hormone as a reporter gene in regulation studies employing transient gene expression. Mol. Cell. Biol. 1986, 6, 3173–3179. [Google Scholar] [CrossRef]
- Nordeen, S.K. Luciferase reporter gene vectors for analysis of promoters and enhancers. Biotechniques 1988, 6, 454–458. [Google Scholar] [PubMed]
- Gorman, C.M.; Moffat, L.F.; Howard, B.H. Recombinant genomes which express chloramphenicol acetyltransferase in mammalian cells. Mol. Cell. Biol. 1982, 2, 1044–1051. [Google Scholar] [CrossRef] [PubMed]
- Chalfie, M.; Tu, Y.; Euskirchen, G.; Ward, W.W.; Prasher, D.C. Green fluorescent protein as a marker for gene expression. Science 1994, 263, 802–805. [Google Scholar] [CrossRef] [PubMed]
- Beal, J. Bridging the gap: A roadmap to breaking the biological design barrier. Front. Bioeng. Biotechnol. 2014, 2, 87. [Google Scholar] [CrossRef]
- Gardner, T.S.; Cantor, C.R.; Collins, J.J. Construction of a genetic toggle switch in Escherichia coli. Nature 2000, 403, 339–342. [Google Scholar] [CrossRef]
- Elowitz, M.B.; Leibler, S. A synthetic oscillatory network of transcriptional regulators. Nature 2000, 403, 335–338. [Google Scholar] [CrossRef] [PubMed]
- Sekine, R.; Yamamura, M.; Ayukawa, S.; Ishimatsu, K.; Akama, S.; Takinoue, M.; Hagiya, M.; Kiga, D. Tunable synthetic phenotypic diversification on waddington’s landscape through autonomous signaling. Proc. Natl. Acad. Sci. USA 2011, 108, 17969–17973. [Google Scholar] [CrossRef]
- Ishimatsu, K.; Hata, T.; Mochizuki, A.; Sekine, R.; Yamamura, M.; Kiga, D. General applicability of synthetic gene-overexpression for cell-type ratio control via reprogramming. ACS Synth. Biol. 2014, 3, 638–644. [Google Scholar] [CrossRef]
- Stricker, J.; Cookson, S.; Bennett, M.R.; Mather, W.H.; Tsimring, L.S.; Hasty, J. A fast, robust and tunable synthetic gene oscillator. Nature 2008, 456, 516–519. [Google Scholar] [CrossRef] [PubMed]
- Moriya, T.; Yamamura, M.; Kiga, D. Effects of downstream genes on synthetic genetic circuits. BMC Syst. Biol. 2014, 8 (Suppl. 4), S4. [Google Scholar] [CrossRef]
- Del Vecchio, D.; Ninfa, A.J.; Sontag, E.D. Modular cell biology: Retroactivity and insulation. Mol. Syst. Biol. 2008, 4, 161. [Google Scholar] [CrossRef] [PubMed]
- Gyorgy, A.; Del Vecchio, D. Modular composition of gene transcription networks. PLoS Comput. Biol. 2014, 10, e1003486. [Google Scholar] [CrossRef] [PubMed]
- Jayanthi, S.; Nilgiriwala, K.S.; Del Vecchio, D. Retroactivity controls the temporal dynamics of gene transcription. ACS Synth. Biol. 2013, 2, 431–441. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.H.; Sauro, H.M. Measuring retroactivity from noise in gene regulatory networks. Biophys. J. 2011, 100, 1167–1177. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.; Scherrer, S.; Buetti-Dinh, A.; Ratna, P.; Pizzolato, J.; Jaquet, V.; Becskei, A. Stochastic signalling rewires the interaction map of a multiple feedback network during yeast evolution. Nat. Commun. 2012, 3, 682. [Google Scholar] [CrossRef] [Green Version]
- Burger, A.; Walczak, A.M.; Wolynes, P.G. Influence of decoys on the noise and dynamics of gene expression. Phys. Rev. E Stat. Nonlinear Soft. Matter Phys. 2012, 86, 041920. [Google Scholar] [CrossRef]
- Lee, T.H.; Maheshri, N. A regulatory role for repeated decoy transcription factor binding sites in target gene expression. Mol. Syst. Biol. 2012, 8, 576. [Google Scholar] [CrossRef]
- Franco, E.; Friedrichs, E.; Kim, J.; Jungmann, R.; Murray, R.; Winfree, E.; Simmel, F.C. Timing molecular motion and production with a synthetic transcriptional clock. Proc. Natl. Acad. Sci. USA 2011, 108, E784–E793. [Google Scholar] [CrossRef] [Green Version]
- Brewster, R.C.; Weinert, F.M.; Garcia, H.G.; Song, D.; Rydenfelt, M.; Phillips, R. The transcription factor titration effect dictates level of gene expression. Cell 2014, 156, 1312–1323. [Google Scholar] [CrossRef] [PubMed]
- Kumar, N.S.; Vecchio, D.D. Loading as a Design Parameter for Genetic Circuits. In Proceedings of the 2016 American Control Conference (ACC), Boston, MA, USA, 6–8 July 2016; pp. 7358–7364. [Google Scholar]
- Saez-Rodriguez, J.; Kremling, A.; Gilles, E.D. Dissecting the puzzle of life: Modularization of signal transduction networks. Comp. Chem. Eng. 2005, 29, 619–629. [Google Scholar] [CrossRef]
- Prindle, A.; Selimkhanov, J.; Li, H.; Razinkov, I.; Tsimring, L.S.; Hasty, J. Rapid and tunable post-translational coupling of genetic circuits. Nature 2014, 508, 387–391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mather, W.H.; Cookson, N.A.; Hasty, J.; Tsimring, L.S.; Williams, R.J. Correlation resonance generated by coupled enzymatic processing. Biophys. J. 2010, 99, 3172–3181. [Google Scholar] [CrossRef] [PubMed]
- Cookson, N.A.; Mather, W.H.; Danino, T.; Mondragon-Palomino, O.; Williams, R.J.; Tsimring, L.S.; Hasty, J. Queueing up for enzymatic processing: Correlated signaling through coupled degradation. Mol. Syst. Biol. 2011, 7, 561. [Google Scholar] [CrossRef] [PubMed]
- Potvin-Trottier, L.; Lord, N.D.; Vinnicombe, G.; Paulsson, J. Synchronous long-term oscillations in a synthetic gene circuit. Nature 2016, 538, 514–517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rondelez, Y. Competition for catalytic resources alters biological network dynamics. Phys. Rev. Lett. 2012, 108, 018102. [Google Scholar] [CrossRef]
- Hasty, J.; Dolnik, M.; Rottschafer, V.; Collins, J.J. Synthetic gene network for entraining and amplifying cellular oscillations. Phys. Rev. Lett. 2002, 88, 148101. [Google Scholar] [CrossRef]
- Smolen, P.; Baxter, D.A.; Byrne, J.H. Frequency selectivity, multistability, and oscillations emerge from models of genetic regulatory systems. Am. J. Physiol. 1998, 274, C531–C542. [Google Scholar] [CrossRef] [PubMed]
- Wong, O.K.; Guthold, M.; Erie, D.A.; Gelles, J. Interconvertible lac repressor-DNA loops revealed by single-molecule experiments. PLoS Biol. 2008, 6, e232. [Google Scholar] [CrossRef]
- Vanzi, F.; Broggio, C.; Sacconi, L.; Pavone, F.S. Lac repressor hinge flexibility and DNA looping: Single molecule kinetics by tethered particle motion. Nucleic Acids Res. 2006, 34, 3409–3420. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.J.; Johnson, S.; Mulligan, P.; Spakowitz, A.J.; Phillips, R. Modulation of DNA loop lifetimes by the free energy of loop formation. Proc. Natl. Acad. Sci. USA 2014, 111, 17396–17401. [Google Scholar] [CrossRef] [Green Version]
- Le, T.T.; Kim, H.D. Probing the elastic limit of DNA bending. Nucleic Acids Res. 2014, 42, 10786–10794. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.K.; Chou, H.H.; Pfleger, B.F.; Newman, J.D.; Yoshikuni, Y.; Keasling, J.D. Directed evolution of arac for improved compatibility of arabinose- and lactose-inducible promoters. Appl. Environ. Microbiol. 2007, 73, 5711–5715. [Google Scholar] [CrossRef]
- Graham, D.; McRuer, D.T. Analysis of Nonlinear Control Systems. Wiley: Hoboken, NJ, USA, 1961. [Google Scholar]
- Rasband, W. ImageJ; Version 1.49; U. S. National Institutes of Health: Bethesda, MA, USA, 2015; pp. 1997–2018.
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. Nih image to imagej: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Meijering, E.; Dzyubachyk, O.; Smal, I. Methods for cell and particle tracking. Methods Enzymol. 2012, 504, 183–200. [Google Scholar]
- Lutz, R.; Bujard, H. Independent and tight regulation of transcriptional units in Escherichia coli via the lacr/o, the tetr/o and arac/i1-i2 regulatory elements. Nucleic Acids Res. 1997, 25, 1203–1210. [Google Scholar] [CrossRef] [PubMed]
- Gillespie, D.T. Exact stochastic simulation of coupled chemical-reactions. J. Phys. Chem. 1977, 81, 2340–2361. [Google Scholar] [CrossRef]
- Elowitz, M.B.; Levine, A.J.; Siggia, E.D.; Swain, P.S. Stochastic gene expression in a single cell. Science 2002, 297, 1183–1186. [Google Scholar] [CrossRef] [PubMed]
- Jayanthi, S.; Del Vecchio, D. Tuning genetic clocks employing DNA binding sites. PLoS ONE 2012, 7, e41019. [Google Scholar] [CrossRef]
- Kim, K.H.; Sauro, H.M. Fan-out in gene regulatory networks. J. Biol. Eng. 2010, 4, 16. [Google Scholar] [CrossRef] [PubMed]
- Tyler, J.; Shiu, A.; Walton, J. Revisiting a synthetic intracellular regulatory network that exhibits oscillations. arXiv, 2018; arXiv:1808.00595. [Google Scholar]
- Page, K.M.; Perez-Carrasco, R. Degradation rate uniformity determines success of oscillations in repressive feedback regulatory networks. J. Royal Soc. Interface 2018, 15, 20180157. [Google Scholar] [CrossRef] [Green Version]
- Ayukawa, S.; Kobayashi, A.; Nakashima, Y.; Takagi, H.; Hamada, S.; Uchiyama, M.; Yugi, K.; Murata, S.; Sakakibara, Y.; Hagiya, M.; et al. Construction of a genetic and gate under a new standard for assembly of genetic parts. BMC Genom. 2010, 11 (Suppl. 4), S16. [Google Scholar] [CrossRef]
- Miyada, C.G.; Horwitz, A.H.; Cass, L.G.; Timko, J.; Wilcox, G. DNA sequence of the arac regulatory gene from Escherichia coli b/r. Nucleic Acids Res. 1980, 8, 5267–5274. [Google Scholar] [CrossRef]
- Shetty, R.P.; Endy, D.; Knight, T.F., Jr. Engineering biobrick vectors from biobrick parts. J. Biol. Eng. 2008, 2, 5. [Google Scholar] [CrossRef] [PubMed]
- Wallace, R.G.; Lee, N.; Fowler, A.V. The arac gene of Escherichia coli: Transcriptional and translational start-points and complete nucleotide sequence. Gene 1980, 12, 179–190. [Google Scholar] [CrossRef]








© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moriya, T.; Yamaoka, T.; Wakayama, Y.; Ayukawa, S.; Zhang, Z.; Yamamura, M.; Wakao, S.; Kiga, D. Comparison between Effects of Retroactivity and Resource Competition upon Change in Downstream Reporter Genes of Synthetic Genetic Circuits. Life 2019, 9, 30. https://doi.org/10.3390/life9010030
Moriya T, Yamaoka T, Wakayama Y, Ayukawa S, Zhang Z, Yamamura M, Wakao S, Kiga D. Comparison between Effects of Retroactivity and Resource Competition upon Change in Downstream Reporter Genes of Synthetic Genetic Circuits. Life. 2019; 9(1):30. https://doi.org/10.3390/life9010030
Chicago/Turabian StyleMoriya, Takefumi, Tomohiro Yamaoka, Yuki Wakayama, Shotaro Ayukawa, Zicong Zhang, Masayuki Yamamura, Shinji Wakao, and Daisuke Kiga. 2019. "Comparison between Effects of Retroactivity and Resource Competition upon Change in Downstream Reporter Genes of Synthetic Genetic Circuits" Life 9, no. 1: 30. https://doi.org/10.3390/life9010030