Increased E2F1 mRNA and miR-17-5p Expression Is Correlated to Invasiveness and Proliferation of Pituitary Neuroendocrine Tumours

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Samples
2.2. Clinical, Radiological, and Pathological Data of the Patients
2.3. Nucleic Acid Extraction
2.4. mRNA and miRNA Expression Analysis
2.5. Statistical Analysis
3. Results
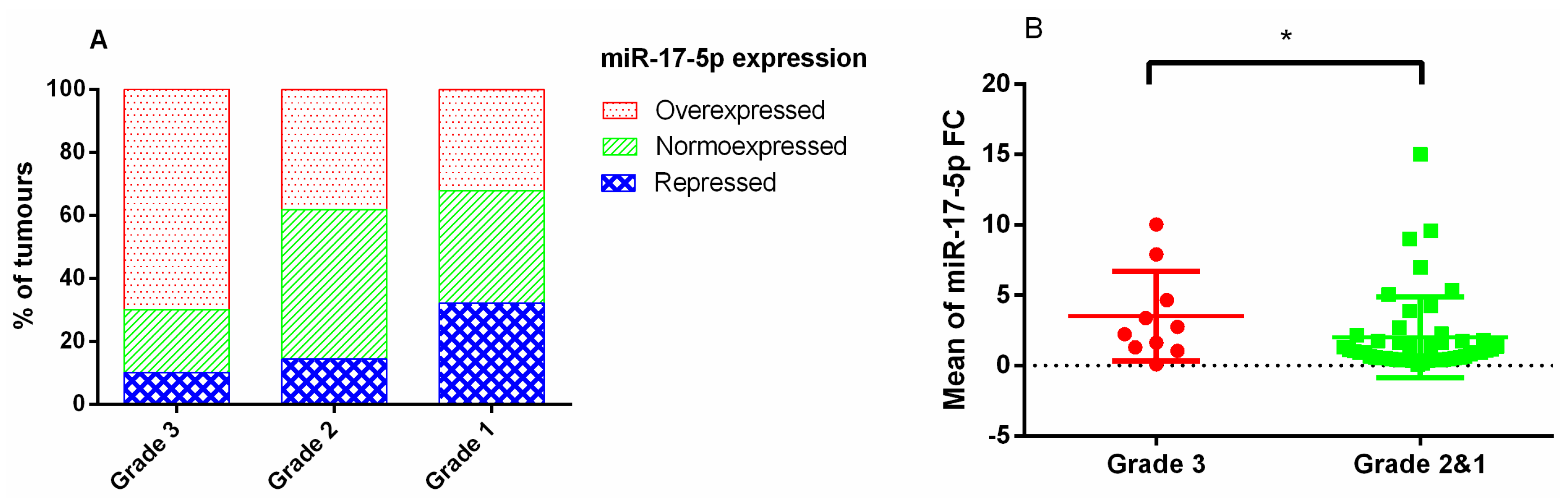
3.1. Relationships between Genes (E2F1/MYC) and miRNAs Studied and PitNET Proliferation
3.2. Relationships between Genes (E2F1/MYC) and miRNAs with the Invasiveness of the Tumours by Magnetic Resonance Imaging
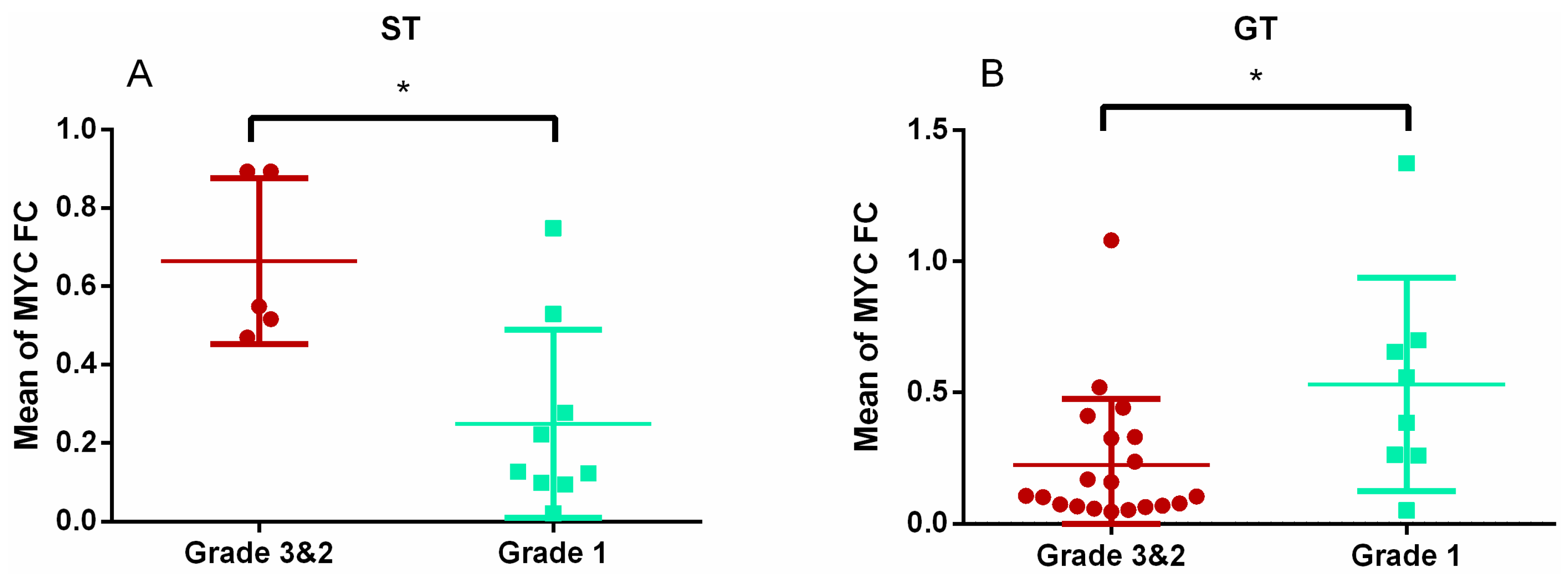
3.3. Expression Profiles of E2F1, MYC Genes, and Pri-miR-17-92 by Subtypes
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| CI | Confidence interval |
| CT | Corticotroph tumour |
| FC | Fold change |
| fCT | Functioning corticotroph tumour |
| GT | Gonadotroph |
| HOF | Human oesophageal fibroblast |
| IHC | Immunohistochemical |
| IQR | Interquartile range |
| MTD | Maximum tumour diameter |
| MRI | Magnetic resonance image |
| OR | Odds ratio |
| qRT-PCR | Quantitative real time polymerase chain reaction |
| ROC | Receiver operator curve |
| RQ | Relative quantification |
| RT-PCR | Real time polymerase chain reaction |
| sCT | Silent corticotroph tumour |
| ST | Somatotroph tumour |
References
- Bell, L.A.; Ryan, K.M. Life and death decisions by E2F-1. Cell Death Differ. 2004, 11, 137–142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olson, M.V.; Johnson, D.G.; Jiang, H.; Xu, J.; Alonso, M.M.; Aldape, K.D.; Fuller, G.N.; Bekele, B.N.; Yung, W.K.A.; Gomez-Manzano, C.; et al. Transgenic E2F1 expression in the mouse brain induces a human-like bimodal pattern of tumors. Cancer Res. 2007, 67, 4005–4009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knoll, S.; Emmrich, S.; Pützer, B.M. The E2F1-miRNA cancer progression network. In MicroRNA Cancer Regulation; Springer: Dordrecht, The Netherland, 2013; pp. 135–147. [Google Scholar] [CrossRef]
- Emmrich, S.; Pützer, B.M. Checks and balances: E2F-microRNA crosstalk in cancer control. Cell Cycle 2010, 9, 2555–2567. [Google Scholar] [CrossRef]
- Massimi, I.; Guerrieri, F.; Petroni, M.; Veschi, V.; Truffa, S.; Screpanti, I.; Frati, S.; Levrero, M.; Gulino, A.; Giannini, G. The HMGA1 protoncogene frequently deregulated in cancer is a transcriptional target of E2F1. Mol. Carcinog. 2013, 52, 526–534. [Google Scholar] [CrossRef]
- Zhou, C.; Wawrowsky, K.; Bannykh, S.; Gutman, S.; Melmed, S. E2F1 induces pituitary tumor transforming gene (PTTG1) expression in human pituitary tumors. Mol. Endocrinol. 2009, 23, 2000–2012. [Google Scholar] [CrossRef] [Green Version]
- Dong, P.; Maddali, M.V.; Srimani, J.K.; Thélot, F.; Nevins, J.R.; Mathey-Prevot, B.; You, L. Division of labour between Myc and G1 cyclins in cell cycle commitment and pace control. Nat. Commun. 2014, 5, 4750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, C.; Han, J.; Dong, D.; Wang, N. MicroRNA-152 suppresses human osteosarcoma cell proliferation and invasion by targeting E2F transcription factor 3. Oncol. Res. 2018, 26, 765–773. [Google Scholar] [CrossRef] [PubMed]
- Sippl, C.; Teping, F.; Ketter, R.; Braun, L.; Tremmel, L.; Schulz-Schaeffer, W.; Oertel, J.; Urbschat, S. The Influence of Distinct Regulatory miRNAs of the p15/p16/RB1/E2F Pathway on the Clinical Progression of Glioblastoma Multiforme. World Neurosurg. 2019, 132, e900–e908. [Google Scholar] [CrossRef]
- Guo, H.; Zhang, L. MicroRNA-30a suppresses papillary thyroid cancer cell proliferation, migration and invasion by directly targeting E2F7. Exp. Ther. Med. 2019, 18, 209–215. [Google Scholar] [CrossRef] [Green Version]
- Di Ieva, A.; Butz, H.; Niamah, M.; Rotondo, F.; De Rosa, S.; Sav, A.; Yousef, G.M.; Kovacs, K.; Cusimano, M.D. MicroRNAs as Biomarkers in Pituitary Tumors. Neurosurgery 2014, 75, 181–189. [Google Scholar] [CrossRef]
- Botelho, C.H.A.; De Magalhães, A.V.; De Mello, P.A.; Schmitt, F.C.; Casulari, L.A. Expression of p53, KI-67 and c-erb B2 in growth hormone- and/or prolactin-secreting pituitary adenomas. Arq. Neuro-Psiquiatr. 2006, 64, 60–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cloonan, N.; Brown, M.K.; Steptoe, A.L.; Wani, S.; Chan, W.L.; Forrest, A.R.; Kolle, G.; Gabrielli, B.; Grimmond, S.M. The miR-17-5p microRNA is a key regulator of the G1/S phase cell cycle transition. Genome Biol. 2008, 9, R127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aguda, B.D.; Kim, Y.; Piper-hunter, M.G.; Friedman, A.; Marsh, C.B. MicroRNA regulation of a cancer network: MiR-17-92, E2F, and Myc. Proc. Natl. Acad. Sci. USA 2008, 105, 19678–19683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luque, R.M.; Ibáñez-Costa, A.; Sánchez-Tejada, L.; Rivero-Cortés, E.; Robledo, M.; Madrazo-Atutxa, A.; Mora, M.; Álvarez, C.V.; Lucas-Morante, T.; Álvarez-Escolá, C.; et al. El Registro Molecular de Adenomas Hipofisarios (REMAH): Una apuesta de futuro de la Endocrinología española por la medicina individualizada y la investigación traslacional. Endocrinol. Nutr. 2016, 63, 274–284. [Google Scholar] [CrossRef]
- Sanchez-Tejada, L.; Sanchez-Ortiga, R.; Lamas, C.; Camara, R.; Riesgo, P.; Fajardo, C.; Aranda, F.I.; Pico, A. Contribution of molecular analysis to the typification of the non-functioning pituitary adenomas. PLoS ONE 2017, 12, e0180039. [Google Scholar] [CrossRef] [Green Version]
- García-Martínez, A.; Sottile, J.; Fajardo, C.; Riesgo, P.; Cámara, R.; Simal, J.A.; Lamas, C.; Sandoval, H.; Aranda, I.; Picó, A. Is it time to consider the expression of specific-pituitary hormone genes when typifying pituitary tumours? PLoS ONE 2018, 13, e0198877. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Micko, A.S.G.; Wöhrer, A.; Wolfsberger, S.; Knosp, E. Invasion of the cavernous sinus space in pituitary adenomas: Endoscopic verification and its correlation with an MRI-based classification. J. Neurosurg. 2015, 122, 803–811. [Google Scholar] [CrossRef]
- Trouillas, J.; Roy, P.; Sturm, N.; Dantony, E.; Cortet-Rudelli, C.; Viennet, G.; Bonneville, J.-F.; Assaker, R.; Auger, C.; Brue, T.; et al. A new prognostic clinicopathological classification of pituitary adenomas: A multicentric case-control study of 410 patients with 8 years post-operative follow-up. Acta Neuropathol. 2013, 126, 123–135. [Google Scholar] [CrossRef]
- Trouillas, J.; Jaffrain-Rea, M.L.; Vasiljevic, A.; Raverot, G.; Roncaroli, F.; Villa, C.C. How to classify the pituitary neuroendocrine tumors (PitNET)s in 2020. Cancers 2020, 12, 514. [Google Scholar] [CrossRef] [Green Version]
- Yang, Q.; Li, X. Molecular network basis of invasive pituitary adenoma: A review. Front. Endocrinol. 2019, 10, 7. [Google Scholar] [CrossRef] [Green Version]
- Scheithauer, B.W.; Kovacs, K.T.; Laws, E.R.; Randall, R.V. Pathology of invasive pituitary tumors with special reference to functional classification. J. Neurosurg. 1986, 65, 733–744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suliman, M.; Royds, J.; Cullen, D.; Timperley, W.; Powell, T.; Battersby, R.; Jones, T.H. Mdm2 and the p53 pathway in human pituitary adenomas. Clin. Endocrinol. 2001, 54, 317–325. [Google Scholar] [CrossRef] [PubMed]
- Madsen, H.; Borges, T.M.; Knox, A.J.; Michaelis, K.A.; Xu, M.; Lillehei, K.O.; Wierman, M.E.; Kleinschmidt-DeMasters, B.K. Giant pituitary adenomas: Pathologic-radiographic correlations and lack of role for p53 and MIB-1 labeling. Am. J. Surg. Pathol. 2011, 35, 1204–1213. [Google Scholar] [CrossRef]
- Hentschel, S.J.; McCutcheon, I.E.; Moore, W.; Durity, F.A. P53 and MIB-1 immunohistochemistry as predictors of the clinical behavior of nonfunctioning pituitary adenomas. Can. J. Neurol. Sci. 2003, 30, 215–219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, D.; Zhang, Y.; Cui, H. Long noncoding RNA CCAT2 is activated by E2F1 and exerts oncogenic properties by interacting with PTTG1 in pituitary adenomas. Am. J. Cancer Res. 2018, 8, 245–255. [Google Scholar]
- Araki, T.; Liu, N.A.; Tone, Y.; Cuevas-Ramos, D.; Heltsley, R.; Tone, M.; Melmed, S. E2F1-mediated human POMC expression in ectopic Cushing’s syndrome. Endocr. Relat. Cancer 2016, 23, 857–870. [Google Scholar] [CrossRef] [Green Version]
- Araki, T.; Liu, X.; Kameda, H.; Tone, Y.; Fukuoka, H.; Tone, M.; Melmed, S. EGFR Induces E2F1-Mediated Corticotroph Tumorigenesis. J. Endocr. Soc. 2017, 1, 127–143. [Google Scholar] [CrossRef] [Green Version]
- Metin-Armagan, D.; Comunoglu, N.; Bulut, G.; Kadioglu, P.; Kameda, H.; Gazioglu, N.; Tanriover, N.; Ozturk, M. A Novel Expression Profile of Cell Cycle and DNA Repair Proteins in Nonfunctioning Pituitary Adenomas. Endocr. Pathol. 2020, 31, 2–13. [Google Scholar] [CrossRef]
- Albihn, A.; Johnsen, J.I.; Henriksson, M.A. MYC in oncogenesis and as a target for cancer therapies. Adv. Cancer Res. 2010, 107, 163–224. [Google Scholar] [CrossRef]
- Fernandez, P.C.; Frank, S.R.; Wang, L.; Schroeder, M.; Liu, S.; Greene, J.; Cocito, A.; Amati, B. Genomic targets of the human c-Myc protein. Genes Dev. 2003, 17, 1115–1129. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Wu, Y.; Yu, S.; Bai, J.; Li, C.; Wu, D.; Zhang, Y. Increased β-catenin and c-myc expression predict aggressive growth of non-functioning pituitary adenomas: An assessment using a tissue microarray-based approach. Mol. Med. Rep. 2017, 15, 1793–1799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, D.G.; Johnston, C.F.; Atkinson, A.B.; Heaney, A.P.; Mirakhur, M.; Buchanan, K.D. Expression of bcl-2 oncoprotein in pituitary tumours: Comparison with c-myc. J. Clin. Pathol. 1996, 49, 795–797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, T.X.; Rothenberg, M.E. MicroRNA. J. Allergy Clin. Immunol. 2018, 141, 1202–1207. [Google Scholar] [CrossRef] [Green Version]
- Gulyaeva, L.F.; Kushlinskiy, N.E. Regulatory mechanisms of microRNA expression. J. Transl. Med. 2016, 14, 143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wierinckx, A.; Roche, M.; Legras-Lachuer, C.; Trouillas, J.; Raverot, G.; Lachuer, J. MicroRNAs in pituitary tumors. Mol. Cell. Endocrinol. 2017, 456, 51–61. [Google Scholar] [CrossRef]
- Fusco, A.; Fedele, M. Roles of HMGA proteins in cancer. Nat. Rev. Cancer 2007, 7, 899–910. [Google Scholar] [CrossRef]
- Palmieri, D.; D’angelo, D.; Valentino, T.; De Martino, I.; Ferraro, A.; Wierinckx, A.; Trouillas, J.; Fusco, A. Downregulation of HMGA-targeting microRNAs has a critical role in human pituitary tumorigenesis. Oncogene 2012, 31, 3857–3865. [Google Scholar] [CrossRef] [Green Version]
- D’Angelo, D.; Palmieri, D.; Mussnich, P.; Roche, M.; Wierinckx, A.; Raverot, G.; Fedele, M.; Croce, C.M.; Trouillas, J.; Fusco, A. Altered microRNA expression profile in human pituitary GH adenomas: Down-regulation of miRNA targeting HMGA1, HMGA2, and E2F1. J. Clin. Endocrinol. Metab. 2012, 97. [Google Scholar] [CrossRef] [PubMed]
- D’Angelo, D.; Esposito, F.; Fusco, A. Epigenetic mechanisms leading to overexpression of HMGA proteins in human pituitary adenomas. Front. Med. 2015, 2. [Google Scholar] [CrossRef] [Green Version]
- Leone, V.; Langella, C.; D’Angelo, D.; Mussnich, P.; Wierinckx, A.; Terracciano, L.; Raverotd, G.; Lachuere, J.; Rotondif, S.; Jaffrain-Rea, M.L.; et al. MiR-23b and miR-130b expression is downregulated in pituitary adenomas. Mol. Cell Endocrinol. 2014, 390, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Roche, M.; Wierinckx, A.; Croze, S.; Rey, C.; Legras-Lachuer, C.; Morel, A.P.; Fusco, A.; Raverot, G.; Trouillas, J.; Lachuer, J. Deregulation of miR-183 and KIAA0101 in aggressive and malignant pituitary tumors. Front. Med. 2015, 2, 54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, Z.; Zhou, C.; Liu, M.; Yao, Y.; Sun, J.; Xiao, J.; Ma, W.; Zhu, H.; Wang, R. MicroRNA involvement in a metastatic non-functioning pituitary carcinoma. Pituitary 2015, 18, 710–721. [Google Scholar] [CrossRef] [PubMed]
- Tani, Y.; Inoshita, N.; Sugiyama, T.; Kato, M.; Yamada, S.; Shichiri, M.; Hirata, Y. Upregulation of CDKN2A and suppression of cyclin D1 gene expressions in ACTH-secreting pituitary adenomas. Eur. J. Endocrinol. 2010, 163, 523–529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Chen, L.; Yang, S.; Fang, D. E2F1: A potential negative regulator of hTERT transcription in normal cells upon activation of oncogenic c-Myc. Med. Sci. Monit. 2012, 18, RA12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Zhang, A.; Shen, C.; Zhang, B.; Rao, Z.; Wang, R.; Yang, S.; Ning, S.; Mao, G.; Fang, D. E2F1 acts as a negative feedback regulator of c-Myc-induced hTERT transcription during tumorigenesis. Oncol. Rep. 2014, 32, 1273–1280. [Google Scholar] [CrossRef]
- Matsumura, I.; Tanaka, H.; Kanakura, Y. E2F1 and c-Myc in cell growth and death. Cell Cycle 2003, 2, 333–338. [Google Scholar] [CrossRef] [Green Version]
- Leone, G.; DeGregori, J.; Sears, R.; Jakoi, L.; Nevins, J.R. Myc and Ras collaborate in inducing accumulation of active cyclin E/Cdk2 and E2F. Nature 1997, 387, 422–426. [Google Scholar] [CrossRef]
- Coller, H.A.; Forman, J.J.; Legesse-Miller, A. “Myc’ed messages”: Myc induces transcription of E2F1 while inhibiting its translation via a microRNA polycistron. PLoS Genet. 2007, 3, 1319–1324. [Google Scholar] [CrossRef] [Green Version]
- O’Donnell, K.A.; Wentzel, E.A.; Zeller, K.I.; Dang, C.V.; Mendell, J.T. c-Myc-regulated microRNAs modulate E2F1 expression. Nature 2005, 435, 839–843. [Google Scholar] [CrossRef]
- Sylvestre, Y.; De Guire, V.; Querido, E.; Mukhopadhyay, U.K.; Bourdeau, V.; Major, F.; Ferbeyre, G.; Chartrand, P. An E2F/miR-20a autoregulatory feedback loop. J. Biol. Chem. 2007, 282, 2135–2143. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | TaqMan Gene Expression Assays | |
|---|---|---|
| E2F1 | Hs00153451_m1 | |
| MYC | Hs00153408_m1 | |
| Ki-67 | Hs01032443_m1 | |
| Pri-miR~17-92 | Hs03295901_pri | |
| Housekeeping genes | ||
| ACTB | Hs99999903_m1 | |
| GAPDH | Hs99999905_m1 | |
| YWHAZ | Hs00237047_m1 | |
| microRNA | TaqMan miRNA Expression Assays | |
| Hsa-miR-17-5p | 002308 | |
| Hsa-miR-20a | 000580 | |
| Small nucleoar RNA (for normalisation) | ||
| RNU43 | 001095 | |
| U54 | 001210 | |
| Variables | Frequency or Mean ± SD | ||
|---|---|---|---|
| Age (years) | 54 ± 15 | ||
| Women | 33/60 (55%) | ||
| Maximum tumour diameter (mm) | 22.51 ± 10.92 | ||
| PitNET subtypes | |||
| ST tumours | 15/60 (25%) | ||
| fCT tumours | 8/60 (13.33%) | ||
| sCT tumours | 8/60 (13.33%) | ||
| GT tumours | 29/60 (48.33%) | ||
| Invasiveness | Knosp grade | 59/60 * | |
| Non-invasive | Grades I–II | 28/59 (47.5%) | |
| Invasive | Grades III–IV | 31/59 (52.5%) | |
| Proliferation | Ki-67 | 59/60 * | |
| Non-proliferative | <2.59 | 42/59 (71.2%) | |
| Proliferative | ≥2.59 | 17/59 (28.8%) | |
| Grades | 59/60 * | ||
| Grade 1 | Non-invasive (regardless Ki-67 index) | 28/59 (47.5%) | |
| Grade 2 | Invasive and non-proliferative (Ki-67 < 2.59) | 21/59 (35.6%) | |
| Grade 3 | Invasive and proliferative (Ki-67 ≥ 2.59) | 10/59 (16.9%) | |
| Subtype | E2F1 | MYC |
|---|---|---|
| Mean Fold Change (IQR) | ||
| All | 2.09 (1.16–3.78) | 0.32 (0.11–0.64) |
| GT | 3.03 (2.01–4.03) * | 0.17 (0.07–0.41) † |
| fCT | 1.32 (1.17–1.90) * | 0.59 (0.44–1.02) † |
| sCT | 3.45 (1.01–5.00) | 0.70 (0.25–1.88) † |
| ST | 0.96 (0.69–1.41) * | 0.28 (0.13–0.54) † |
| p value | <0.001 | 0.004 |
| Subtype | Pri-miR-17~92 | miR-17-5p | miR-20a |
|---|---|---|---|
| Mean Fold Change (IQR) | |||
| All | 0.12 (0.08–0.27) | 1.15 (0.53–2.61) | 1.21 (0.51–2.54) |
| GT | 0.11 (0.07–0.25) | 1.62 (0.92–2.77) | 1.69 (0.80–2.82) |
| fCT | 0.13 (0.09–0.24) | 0.69 (0.53–1.27) | 1.02 (0.53–1.28) |
| sCT | 0.22 (0.13–0.31) | 1.59 (0.54–6.84) | 1.56 (0.55–8.44) |
| ST | 0.12 (0.09–0.42) | 0.92 (0.42–1.56) | 0.83 (0.46–1.93) |
| p value | 0.421 | 0.286 | 0.477 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
García-Martínez, A.; López-Muñoz, B.; Fajardo, C.; Cámara, R.; Lamas, C.; Silva-Ortega, S.; Aranda, I.; Picó, A. Increased E2F1 mRNA and miR-17-5p Expression Is Correlated to Invasiveness and Proliferation of Pituitary Neuroendocrine Tumours. Diagnostics 2020, 10, 227. https://doi.org/10.3390/diagnostics10040227
García-Martínez A, López-Muñoz B, Fajardo C, Cámara R, Lamas C, Silva-Ortega S, Aranda I, Picó A. Increased E2F1 mRNA and miR-17-5p Expression Is Correlated to Invasiveness and Proliferation of Pituitary Neuroendocrine Tumours. Diagnostics. 2020; 10(4):227. https://doi.org/10.3390/diagnostics10040227
Chicago/Turabian StyleGarcía-Martínez, Araceli, Beatriz López-Muñoz, Carmen Fajardo, Rosa Cámara, Cristina Lamas, Sandra Silva-Ortega, Ignacio Aranda, and Antonio Picó. 2020. "Increased E2F1 mRNA and miR-17-5p Expression Is Correlated to Invasiveness and Proliferation of Pituitary Neuroendocrine Tumours" Diagnostics 10, no. 4: 227. https://doi.org/10.3390/diagnostics10040227