
The Therapeutic Potential of ADAMTS8 in Lung Adenocarcinoma without Targetable Therapy

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. NGS and Quantitative Real-Time Reverse Transcription Polymerase Chain Reaction (Q-RT-PCR) Assay
2.2. Cell Culture
2.3. Bioinformatics
2.4. Transcription Factors of ADAMTS8
2.5. Screening for Differentially Expressed miRNAs
2.6. Gene Set Enrichment Analysis
2.7. The Survival Significance of ADAMTS8 in EGFR-Mutant or EGFR-Wild-Type Tumors
2.8. Validation of ADAMTS8 on Databases of Lung Cancer Cell Lines
2.9. Statistical Analysis
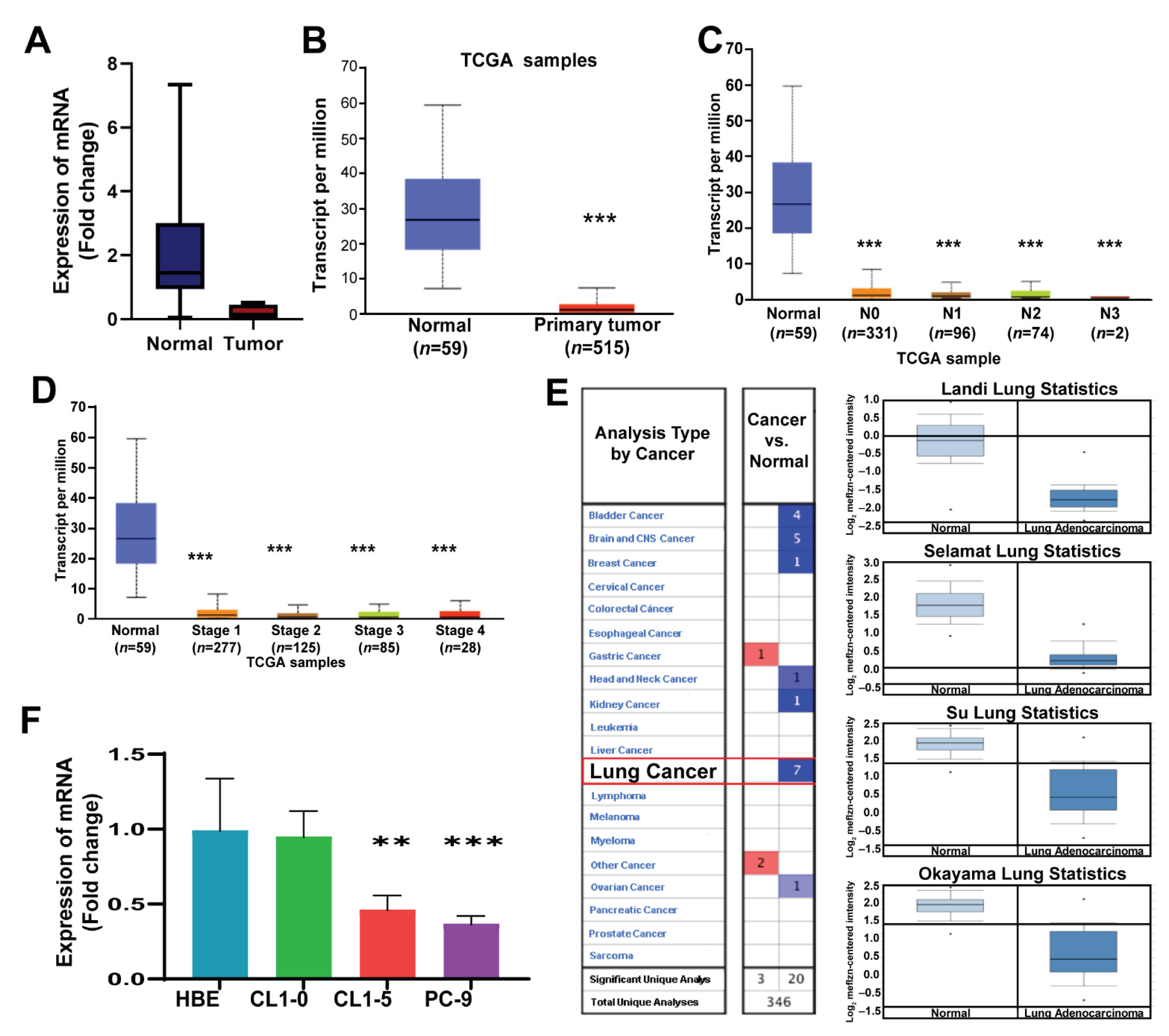
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

References
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer statistics, 2022. CA Cancer J. Clin. 2022, 72, 7–33. [Google Scholar] [CrossRef] [PubMed]
- Tsai, Y.M.; Wu, K.L.; Chang, Y.Y.; Hung, J.Y.; Chang, W.A.; Chang, C.Y.; Jian, S.F.; Tsai, P.H.; Huang, Y.C.; Chong, I.W.; et al. Upregulation of Thr/Tyr kinase Increases the Cancer Progression by Neurotensin and Dihydropyrimidinase-Like 3 in Lung Cancer. Int. J. Mol. Sci. 2020, 21, 1640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uchino, J.; Goldmann, T.; Kimura, H. Editorial: Treatment for Non-Small Cell Lung Cancer in Distinct Patient Populations. Front. Oncol. 2022, 12, 838570. [Google Scholar] [CrossRef] [PubMed]
- Mithoowani, H.; Febbraro, M. Non-Small-Cell Lung Cancer in 2022: A Review for General Practitioners in Oncology. Curr. Oncol. 2022, 29, 1828–1839. [Google Scholar] [CrossRef]
- Tsai, Y.M.; Wu, K.L.; Chang, Y.Y.; Chang, W.A.; Huang, Y.C.; Jian, S.F.; Tsai, P.H.; Lin, Y.S.; Chong, I.W.; Hung, J.Y.; et al. Loss of miR-145-5p Causes Ceruloplasmin Interference with PHD-Iron Axis and HIF-2α Stabilization in Lung Adenocarcinoma-Mediated Angiogenesis. Int. J. Mol. Sci. 2020, 21, 5081. [Google Scholar] [CrossRef]
- Gridelli, C.; Peters, S.; Mok, T.; Forde, P.M.; Reck, M.; Attili, I.; de Marinis, F. First-line immunotherapy in advanced non-small-cell lung cancer patients with ECOG performance status 2: Results of an International Expert Panel Meeting by the Italian Association of Thoracic Oncology. ESMO Open 2022, 7, 100355. [Google Scholar] [CrossRef]
- Chang, Y.-J.; Huang, J.-Y.; Lin, C.-H.; Wang, B.-Y. Survival and Treatment of Lung Cancer in Taiwan between 2010 and 2016. J. Clin. Med. 2021, 10, 4675. [Google Scholar] [CrossRef]
- Wu, L.; Ke, L.; Zhang, Z.; Yu, J.; Meng, X. Development of EGFR TKIs and Options to Manage Resistance of Third-Generation EGFR TKI Osimertinib: Conventional Ways and Immune Checkpoint Inhibitors. Front. Oncol. 2020, 10, 602762. [Google Scholar] [CrossRef]
- Yun, C.-H.; Mengwasser, K.E.; Toms, A.V.; Woo, M.S.; Greulich, H.; Wong, K.-K.; Meyerson, M.; Eck, M.J. The T790M mutation in EGFR kinase causes drug resistance by increasing the affinity for ATP. Proc. Natl. Acad. Sci. USA 2008, 105, 2070–2075. [Google Scholar] [CrossRef] [Green Version]
- Kelwick, R.; Desanlis, I.; Wheeler, G.N.; Edwards, D.R. The ADAMTS (A Disintegrin and Metalloproteinase with Thrombospondin motifs) family. Genome Biol. 2015, 16, 113. [Google Scholar] [CrossRef] [Green Version]
- Apte, S.S. A Disintegrin-like and Metalloprotease (Reprolysin-type) with Thrombospondin Type 1 Motif (ADAMTS) Superfamily: Functions and Mechanisms. J. Biol. Chem. 2009, 284, 31493–31497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mosyak, L.; Georgiadis, K.; Shane, T.; Svenson, K.; Hebert, T.; McDonagh, T.; Mackie, S.; Olland, S.; Lin, L.; Zhong, X.; et al. Crystal structures of the two major aggrecan degrading enzymes, ADAMTS4 and ADAMTS5. Protein Sci. 2008, 17, 16–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakayama, H.; Fukuda, S.; Inoue, H.; Nishida-Fukuda, H.; Shirakata, Y.; Hashimoto, K.; Higashiyama, S. Cell surface annexins regulate ADAM-mediated ectodomain shedding of proamphiregulin. Mol. Biol. Cell 2012, 23, 1964–1975. [Google Scholar] [CrossRef] [PubMed]
- Somerville, R.P.T.; Longpré, J.-M.; Apel, E.D.; Lewis, R.M.; Wang, L.W.; Sanes, J.R.; Leduc, R.; Apte, S.S. ADAMTS7B, the Full-length Product of the ADAMTS7 Gene, Is a Chondroitin Sulfate Proteoglycan Containing a Mucin Domain. J. Biol. Chem. 2004, 279, 35159–35175. [Google Scholar] [CrossRef] [Green Version]
- Tang, B.L. ADAMTS: A novel family of extracellular matrix proteases. Int. J. Biochem. Cell Biol. 2001, 33, 33–44. [Google Scholar] [CrossRef]
- Colige, A.; Sieron, A.L.; Li, S.-W.; Schwarze, U.; Petty, E.; Wertelecki, W.; Wilcox, W.; Krakow, D.; Cohn, D.H.; Reardon, W.; et al. Human Ehlers-Danlos Syndrome Type VII C and Bovine Dermatosparaxis Are Caused by Mutations in the Procollagen I N-Proteinase Gene. Am. J. Hum. Genet. 1999, 65, 308–317. [Google Scholar] [CrossRef] [Green Version]
- Dagoneau, N.; Benoist-Lasselin, C.; Huber, C.; Faivre, L.; Mégarbané, A.; Alswaid, A.; Dollfus, H.; Alembik, Y.; Munnich, A.; Legeai-Mallet, L.; et al. ADAMTS10 Mutations in Autosomal Recessive Weill-Marchesani Syndrome. Am. J. Hum. Genet. 2004, 75, 801–806. [Google Scholar] [CrossRef] [Green Version]
- Levy, G.G.; Nichols, W.C.; Lian, E.C.; Foroud, T.; McClintick, J.N.; McGee, B.M.; Yang, A.Y.; Siemieniak, D.R.; Stark, K.R.; Gruppo, R.; et al. Mutations in a member of the ADAMTS gene family cause thrombotic thrombocytopenic purpura. Nature 2001, 413, 488–494. [Google Scholar] [CrossRef] [Green Version]
- Morales, J.; Al-Sharif, L.; Khalil, D.S.; Shinwari, J.M.A.; Bavi, P.; Al-Mahrouqi, R.A.; Al-Rajhi, A.; Alkuraya, F.S.; Meyer, B.F.; Al Tassan, N. Homozygous Mutations in ADAMTS10 and ADAMTS17 Cause Lenticular Myopia, Ectopia Lentis, Glaucoma, Spherophakia, and Short Stature. Am. J. Hum. Genet. 2009, 85, 558–568. [Google Scholar] [CrossRef] [Green Version]
- Mead, T.J.; Apte, S.S. ADAMTS proteins in human disorders. Matrix Biol. 2018, 71–72, 225–239. [Google Scholar] [CrossRef]
- Wang, Y.; Zhou, J.; Zhang, J.; Cao, H.; Han, F.; Zhang, H.; Xu, E. The expression of ADAMTS14 is regulated by promoter DNA methylation and is associated with poor prognosis in colorectal cancer. Exp. Cell Res. 2022, 410, 112953. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Luo, X.; Huang, B.; Wang, X.; Deng, Y.; Zhong, Z. ADAMTS12 acts as a cancer promoter in colorectal cancer via activating the Wnt/β-catenin signaling pathway in vitro. Ann. Transl. Med. 2020, 8, 301. [Google Scholar] [CrossRef] [PubMed]
- Horgan, D.; Curigliano, G.; Rieß, O.; Hofman, P.; Büttner, R.; Conte, P.; Cufer, T.; Gallagher, W.M.; Georges, N.; Kerr, K.; et al. Identifying the Steps Required to Effectively Implement Next-Generation Sequencing in Oncology at a National Level in Europe. J. Pers. Med. 2022, 12, 72. [Google Scholar] [CrossRef] [PubMed]
- Rhodes, D.R.; Yu, J.; Shanker, K.; Deshpande, N.; Varambally, R.; Ghosh, D.; Barrette, T.; Pander, A.; Chinnaiyan, A.M. ONCOMINE: A cancer microarray database and integrated data-mining platform. Neoplasia 2004, 6, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Chandrashekar, D.S.; Bashel, B.; Balasubramanya, S.A.H.; Creighton, C.J.; Ponce-Rodriguez, I.; Chakravarthi, B.; Varambally, S. UALCAN: A Portal for Facilitating Tumor Subgroup Gene Expression and Survival Analyses. Neoplasia 2017, 19, 649–658. [Google Scholar] [CrossRef]
- Lanczky, A.; Gyorffy, B. Web-Based Survival Analysis Tool Tailored for Medical Research (KMplot): Development and Implementation. J. Med. Internet Res. 2021, 23, e27633. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING v11: Protein–protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2018, 47, D607–D613. [Google Scholar] [CrossRef] [Green Version]
- Yuan, H.; Yan, M.; Zhang, G.; Liu, W.; Deng, C.; Liao, G.; Xu, L.; Luo, T.; Yan, H.; Long, Z.; et al. CancerSEA: A cancer single-cell state atlas. Nucleic Acids Res. 2018, 47, D900–D908. [Google Scholar] [CrossRef] [Green Version]
- Miao, Y.-R.; Zhang, Q.; Lei, Q.; Luo, M.; Xie, G.-Y.; Wang, H.; Guo, A.-Y. ImmuCellAI: A Unique Method for Comprehensive T-Cell Subsets Abundance Prediction and its Application in Cancer Immunotherapy. Adv. Sci. 2020, 7, 1902880. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Liu, W.; Zhang, H.M.; Xie, G.Y.; Miao, Y.R.; Xia, M.; Guo, A.Y. hTFtarget: A Comprehensive Database for Regulations of Human Transcription Factors and Their Targets. Genom. Proteom. Bioinform. 2020, 18, 120–128. [Google Scholar] [CrossRef]
- Nusinow, D.P.; Szpyt, J.; Ghandi, M.; Rose, C.M.; McDonald, E.R., 3rd; Kalocsay, M.; Jané-Valbuena, J.; Gelfand, E.; Schweppe, D.K.; Jedrychowski, M.; et al. Quantitative Proteomics of the Cancer Cell Line Encyclopedia. Cell 2020, 180, 387–402.e16. [Google Scholar] [CrossRef] [PubMed]
- Omura, J.; Satoh, K.; Kikuchi, N.; Satoh, T.; Kurosawa, R.; Nogi, M.; Ohtsuki, T.; Al-Mamun, M.E.; Siddique, M.A.H.; Yaoita, N.; et al. ADAMTS8 Promotes the Development of Pulmonary Arterial Hypertension and Right Ventricular Failure: A Possible Novel Therapeutic Target. Circ. Res. 2019, 125, 884–906. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Tian, R.; Wang, G.; Zhang, J.; Ma, H.; Hu, X.; Xi, J.; Wang, G. ADAMTS8 Inhibits Cell Proliferation and Invasion, and Induces Apoptosis in Breast Cancer. OncoTargets Ther. 2020, 13, 8373–8382. [Google Scholar] [CrossRef] [PubMed]
- Dunn, J.R.; Reed, J.E.; du Plessis, D.G.; Shaw, E.J.; Reeves, P.; Gee, A.L.; Warnke, P.; Walker, C. Expression of ADAMTS-8, a secreted protease with antiangiogenic properties, is downregulated in brain tumours. Br. J. Cancer 2006, 94, 1186–1193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fares, J.; Fares, M.Y.; Khachfe, H.H.; Salhab, H.A.; Fares, Y. Molecular principles of metastasis: A hallmark of cancer revisited. Signal Transduct. Target. Ther. 2020, 5, 28. [Google Scholar] [CrossRef]
- Choi, G.C.; Li, J.; Wang, Y.; Li, L.; Zhong, L.; Ma, B.; Su, X.; Ying, J.; Xiang, T.; Rha, S.Y.; et al. The metalloprotease ADAMTS8 displays antitumor properties through antagonizing EGFR-MEK-ERK signaling and is silenced in carcinomas by CpG methylation. Mol. Cancer Res. MCR 2014, 12, 228–238. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.; Yang, C.; Wu, J.; Nan, Y. ADAMTS8 targets ERK to suppress cell proliferation, invasion, and metastasis of hepatocellular carcinoma. OncoTargets Ther. 2018, 11, 7569–7578. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Hu, K.; Qu, Z.; Xie, Z.; Tian, F. ADAMTS8 inhibited lung cancer progression through suppressing VEGFA. Biochem. Biophys. Res. Commun. 2022, 598, 1–8. [Google Scholar] [CrossRef]
- Wang, L.-L.; Chen, Z.-S.; Zhou, W.-D.; Shu, J.; Wang, X.-H.; Jin, R.; Zhuang, L.-L.; Hoda, M.A.; Zhang, H.; Zhou, G.-P. Down-regulated GATA-1 up-regulates interferon regulatory factor 3 in lung adenocarcinoma. Sci. Rep. 2017, 7, 2551. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Zhang, J.; Li, X.; Zhang, C.; Zhang, H.; Jin, J.; Dai, D. Downregulation of ADAMTS8 by DNA Hypermethylation in Gastric Cancer and Its Clinical Significance. BioMed Res. Int. 2016, 2016, 5083841. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Luo, Y.; Zhou, Y.; Qin, S.; Qiu, Y.; Cui, R.; Yu, M.; Qin, J.; Zhong, M. Promotion of Tumor Growth by ADAMTS4 in Colorectal Cancer: Focused on Macrophages. Cell. Physiol. Biochem. 2018, 46, 1693–1703. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| TF Expression Level a | TF Expression Level b | Correlation with ADAMTS8 b | ||
|---|---|---|---|---|
| Transcription factors | Log2 Fold Change | Log2 Fold Change | Pearson’s r | p-value |
| Low expression in tumor | ||||
| GATA1 | −0.85 | −1.09 * | 0.62 | 3.96 × 10−63 |
| RARA | 0.33 | −0.01 * | 0.19 | 4.13 × 10−6 |
| LMO2 | −1.25 | −0.26 * | 0.59 | 3.27 × 10−54 |
| High expression in tumor | ||||
| E2F4 | 0.24 | 0.12 * | −0.28 | 7.89 × 10−12 |
| CTBP2 | 0.32 | 0.14 * | −0.35 | 2.75 × 10−18 |
| Survival Association a | mRNA Expression Level b | Correlation with ADAMTS8 b | |||
|---|---|---|---|---|---|
| mRNA | HR for OS * | p-Value | Log2 Fold Change | Pearson’s r | p-Value |
| POFUT2 | 0.58 | 5.6 × 10−10 | 1.11 | −0.12 | 3.00 × 10−3 |
| ACAN | 0.57 | 3.7 × 10−11 | 1.44 | −0.17 | 4.50 × 10−5 |
| ADAMTS19 | 0.60 | 3.1 × 10−9 | 0.73 | 0.03 | 4.06 × 10−1 |
| THBS1 | 0.68 | 7.1 × 10−6 | 0.99 | 0.19 | 3.39 × 10−6 |
| ADAMTS13 | 0.60 | 1.4 × 10−9 | 1.12 | 0.11 | 1.00 × 10−2 |
| ADAMTS2 | 0.66 | 1.0 × 10−6 | 1.04 | 0.06 | 1.54 × 10−1 |
| ADAMTS10 | 0.60 | 1.5 × 10−9 | 1.04 | 0.25 | 9.61 × 10−10 |
| THBS2 | 0.81 | 1.4 × 10−2 | 1.36 | −0.36 | 1.29 × 10−18 |
| Immune Cells | Pearson’s r | p-Value |
|---|---|---|
| NKT | 0.630 | 2.50 × 10−64 |
| Tfh | 0.510 | 2.39 × 10−39 |
| NK | 0.450 | 1.02 × 1029 |
| Th2 | 0.440 | 1.09 × 10−28 |
| CD4_T | 0.400 | 1.36 × 10−23 |
| MAIT | 0.400 | 3.76 × 10−23 |
| Tgd | 0.280 | 8.47 × 10−12 |
| Macrophage | 0.190 | 2.53 × 10−6 |
| Tc | 0.130 | 2.00 × 10−3 |
| Th17 | 0.009 | 8.25 × 10−1 |
| CD4_naive | −0.001 | 9.79 × 10−1 |
| DC | −0.020 | 6.20 × 10−1 |
| Neutrophil | −0.020 | 6.79 × 10−1 |
| Tcm | −0.030 | 5.39 × 10−1 |
| CD8_T | −0.050 | 2.59 × 10−1 |
| CD8_naive | −0.060 | 1.27 × 10−1 |
| B_cell | −0.080 | 5.20 × 10−2 |
| Tr1 | −0.110 | 9.00 × 10−3 |
| iTreg | −0.200 | 8.80 × 10−7 |
| Monocyte | −0.260 | 1.70 × 10−10 |
| Th1 | −0.290 | 1.92 × 10−12 |
| Tex | −0.390 | 5.26 × 10−22 |
| Tem | −0.420 | 3.48 × 10−26 |
| nTreg | −0.670 | 3.11 × 10−77 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, H.-C.; Chang, C.-Y.; Wu, K.-L.; Chiang, H.-H.; Chang, Y.-Y.; Liu, L.-X.; Huang, Y.-C.; Hung, J.-Y.; Hsu, Y.-L.; Wu, Y.-Y.; et al. The Therapeutic Potential of ADAMTS8 in Lung Adenocarcinoma without Targetable Therapy. J. Pers. Med. 2022, 12, 902. https://doi.org/10.3390/jpm12060902
Lee H-C, Chang C-Y, Wu K-L, Chiang H-H, Chang Y-Y, Liu L-X, Huang Y-C, Hung J-Y, Hsu Y-L, Wu Y-Y, et al. The Therapeutic Potential of ADAMTS8 in Lung Adenocarcinoma without Targetable Therapy. Journal of Personalized Medicine. 2022; 12(6):902. https://doi.org/10.3390/jpm12060902
Chicago/Turabian StyleLee, Hsiao-Chen, Chao-Yuan Chang, Kuan-Li Wu, Hung-Hsing Chiang, Yung-Yun Chang, Lian-Xiu Liu, Yung-Chi Huang, Jen-Yu Hung, Ya-Ling Hsu, Yu-Yuan Wu, and et al. 2022. "The Therapeutic Potential of ADAMTS8 in Lung Adenocarcinoma without Targetable Therapy" Journal of Personalized Medicine 12, no. 6: 902. https://doi.org/10.3390/jpm12060902