
Global Habitat Suitability of Spodoptera frugiperda (JE Smith) (Lepidoptera, Noctuidae): Key Parasitoids Considered for Its Biological Control

 , , , , , , , , , , , , , and
, , , , , , , , , , , , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Species Studied and Presence Records
2.2. Environmental Variables
2.3. Modeling Procedures
2.4. Prediction Maps
2.5. Model Calibration
3. Results
3.1. Evaluation of Models
3.2. Variable Importance
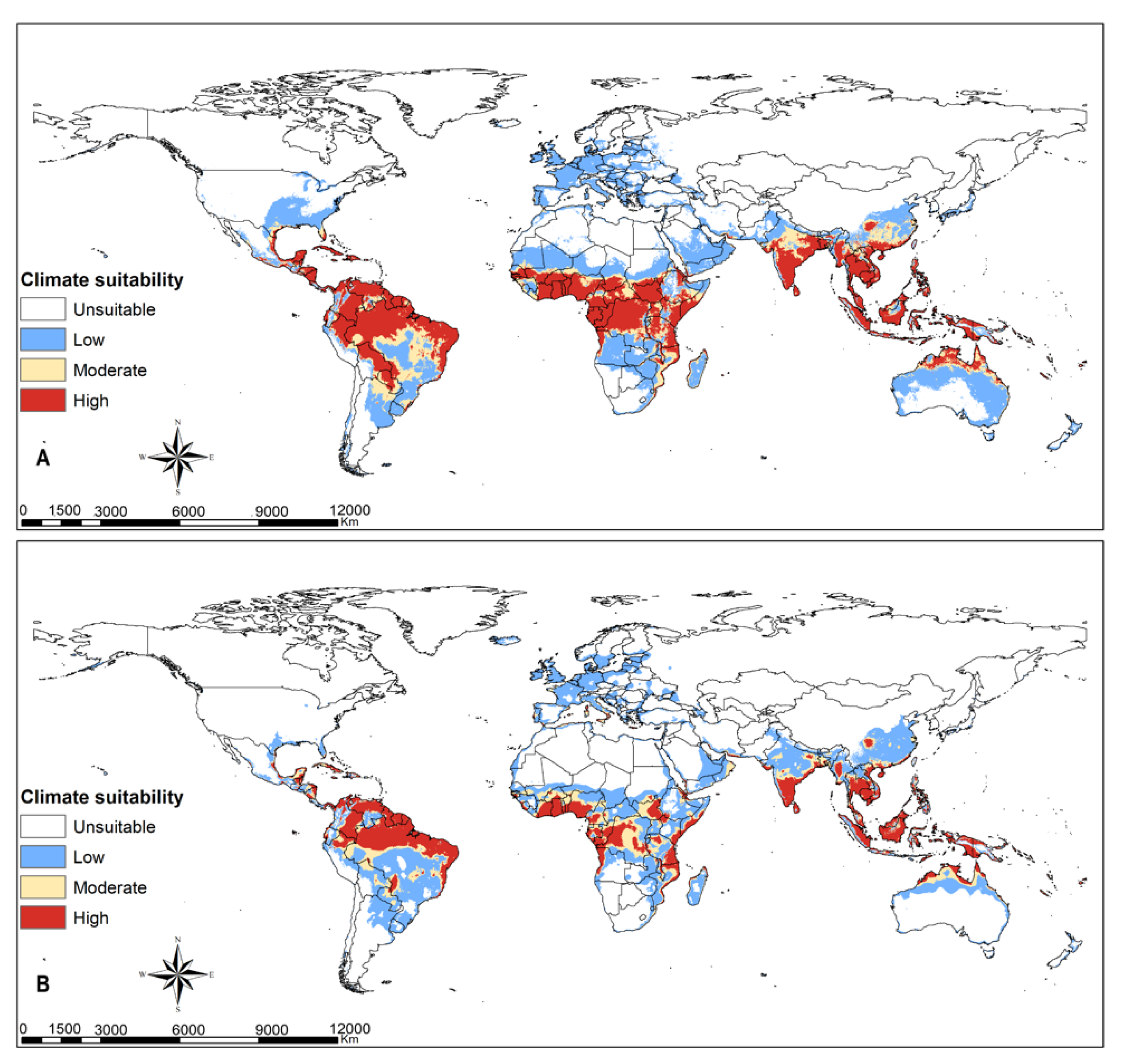
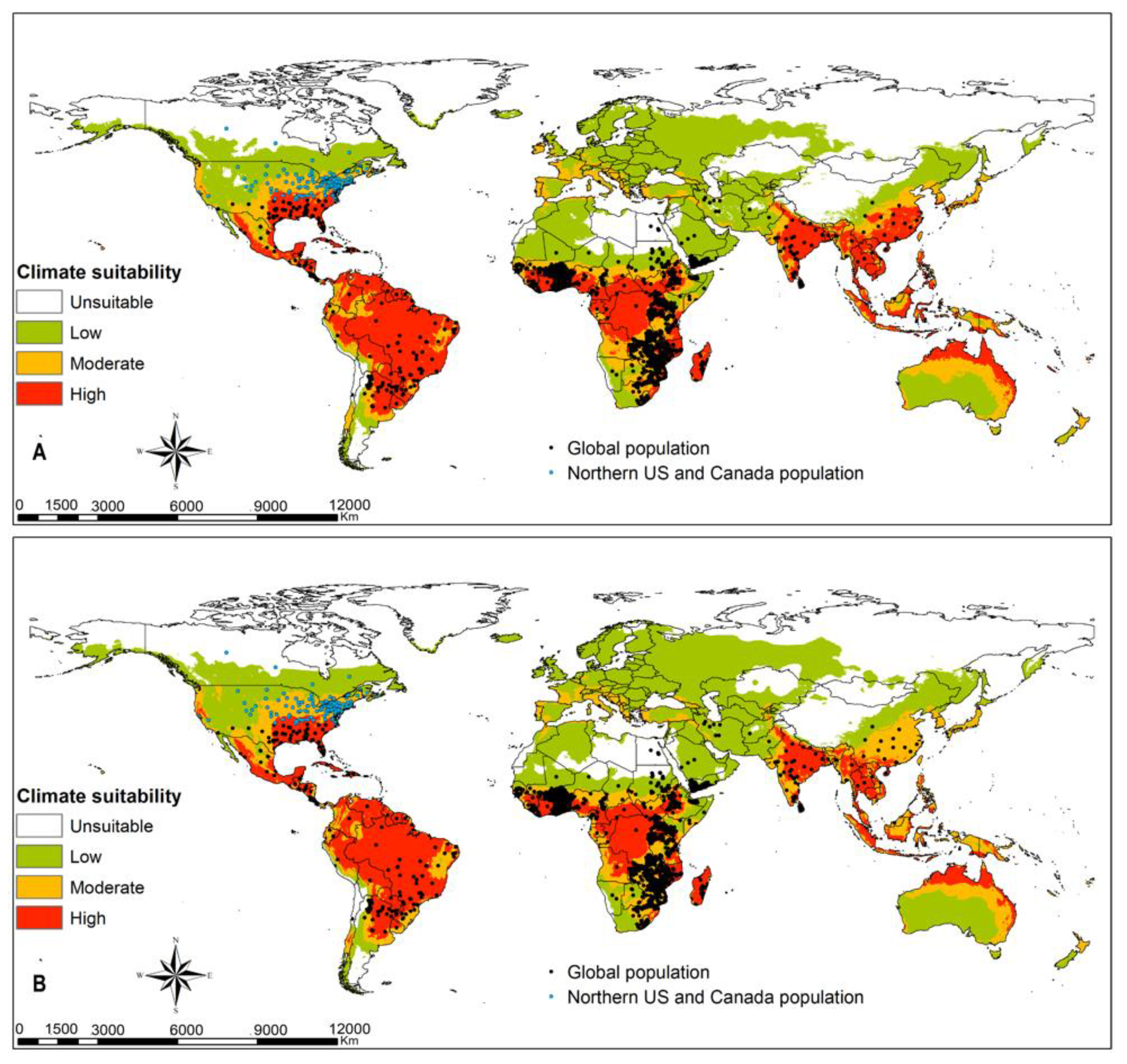
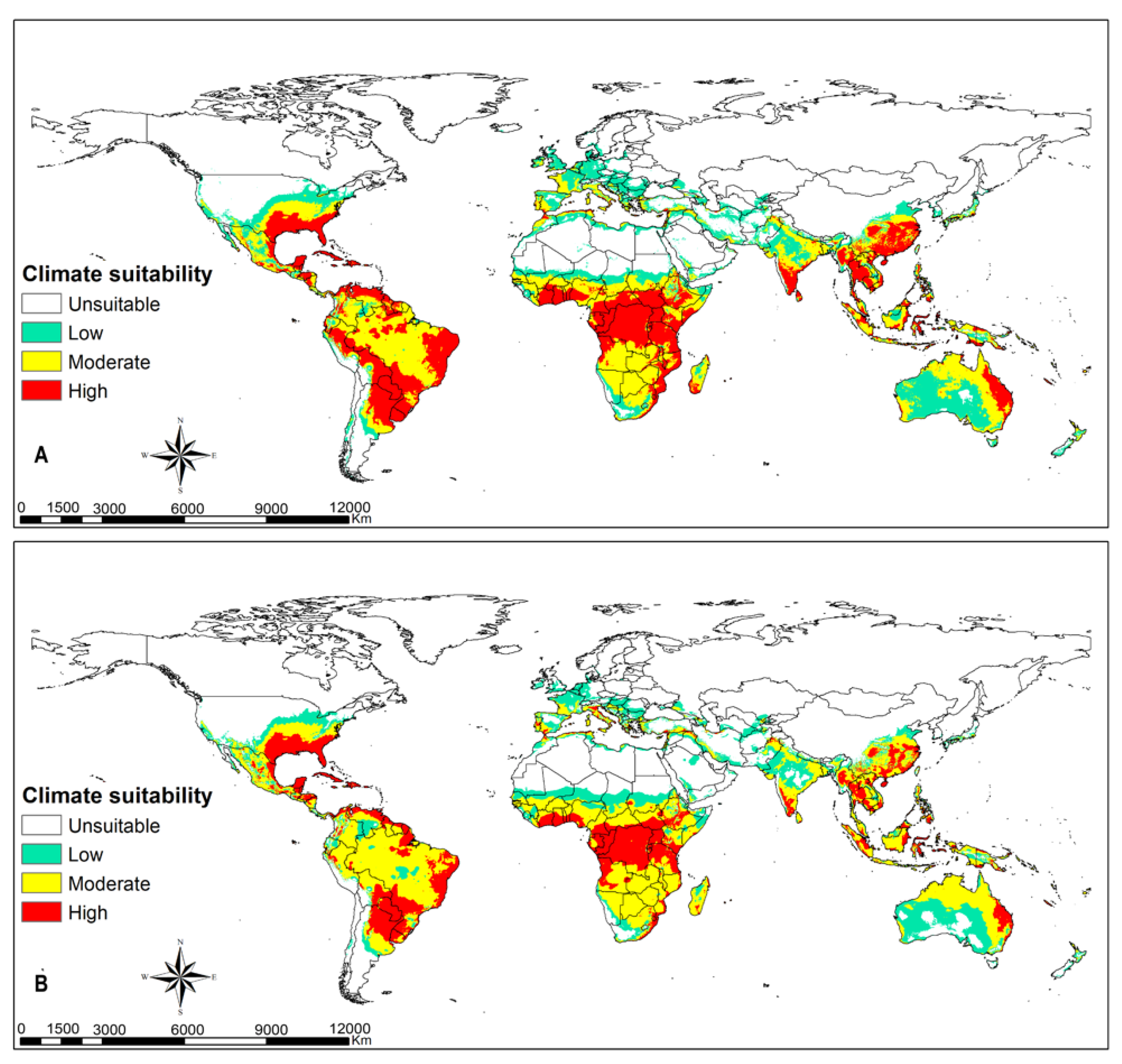
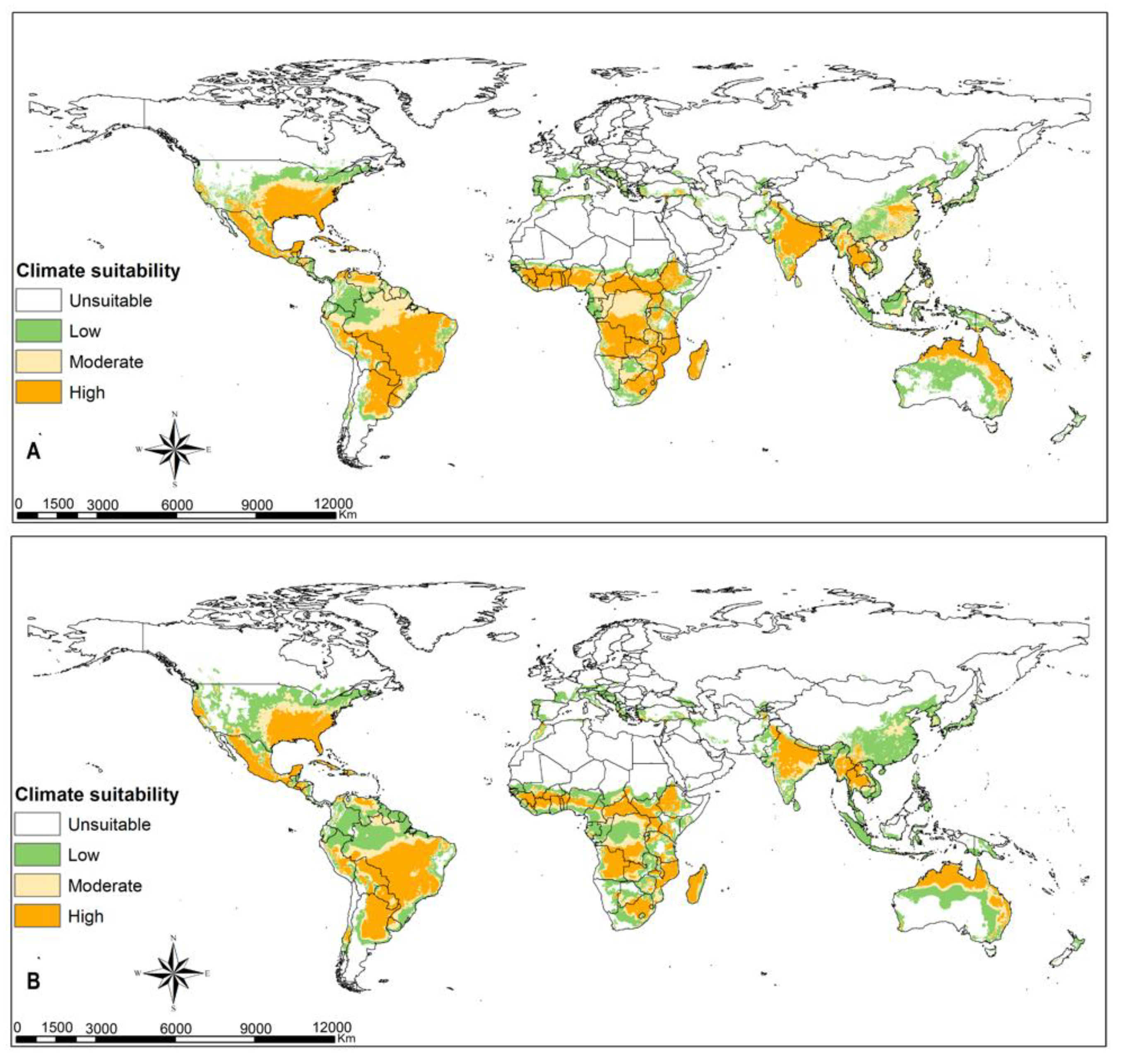
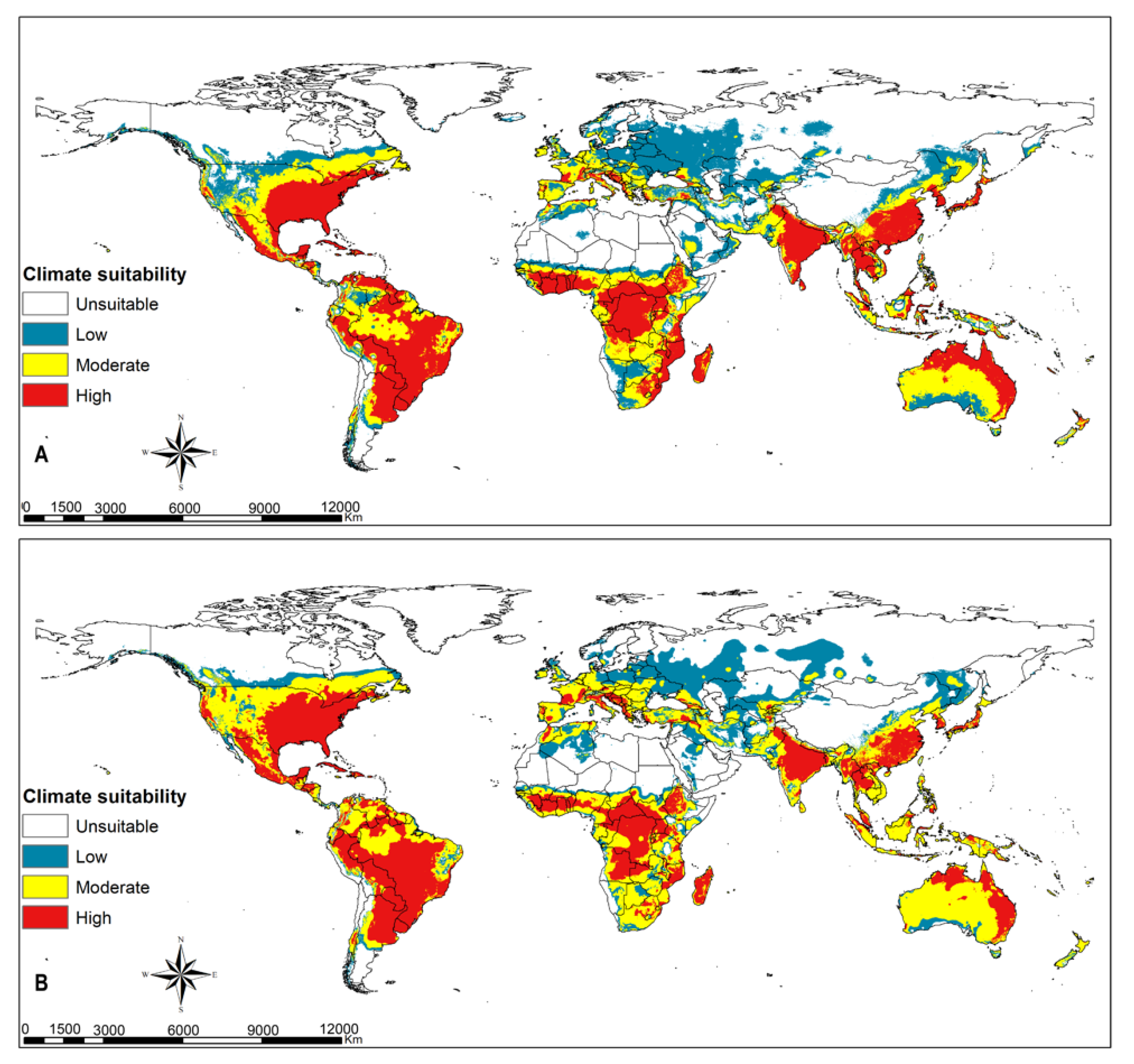
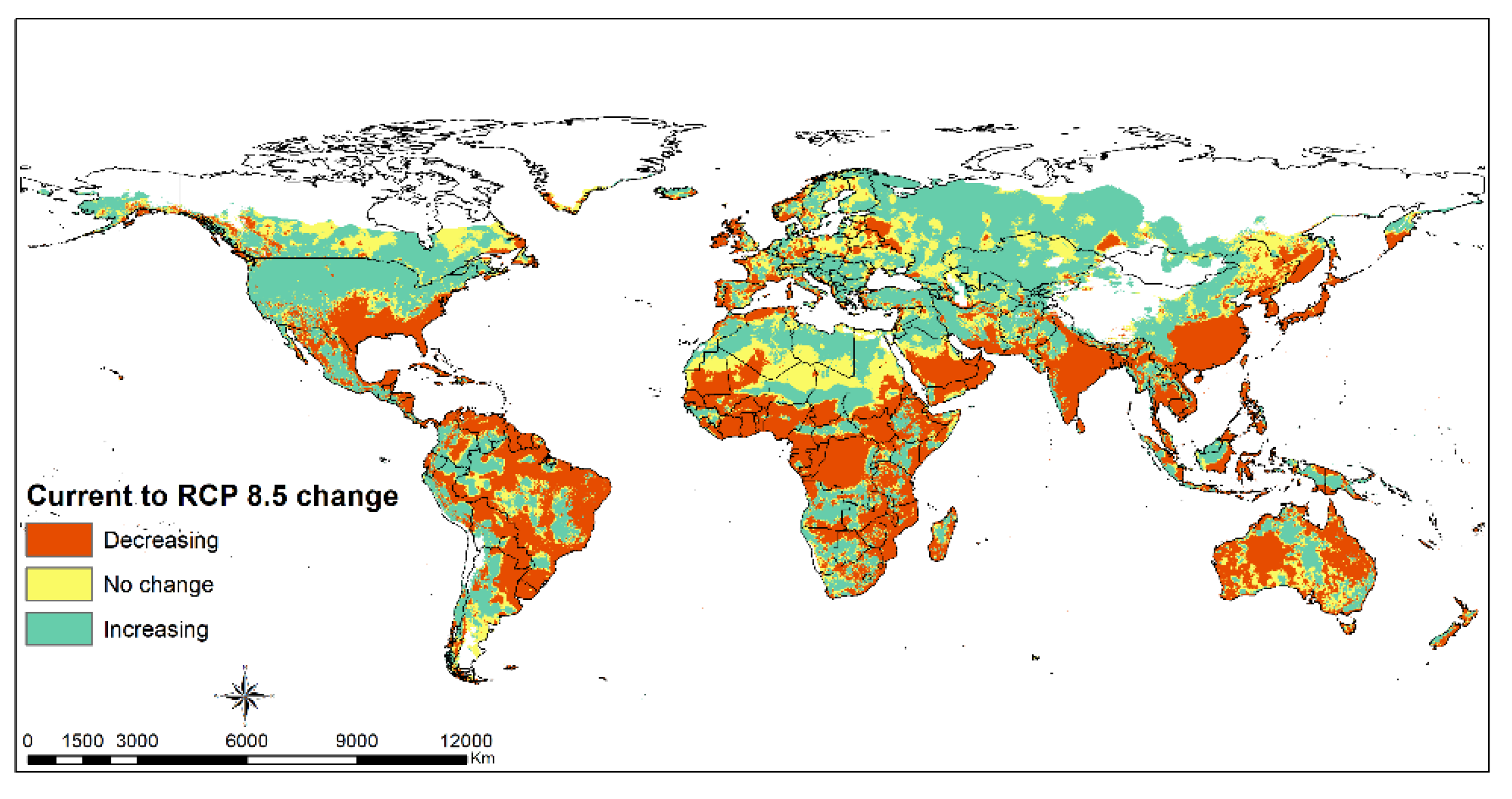
3.3. FAW Habitat Suitability
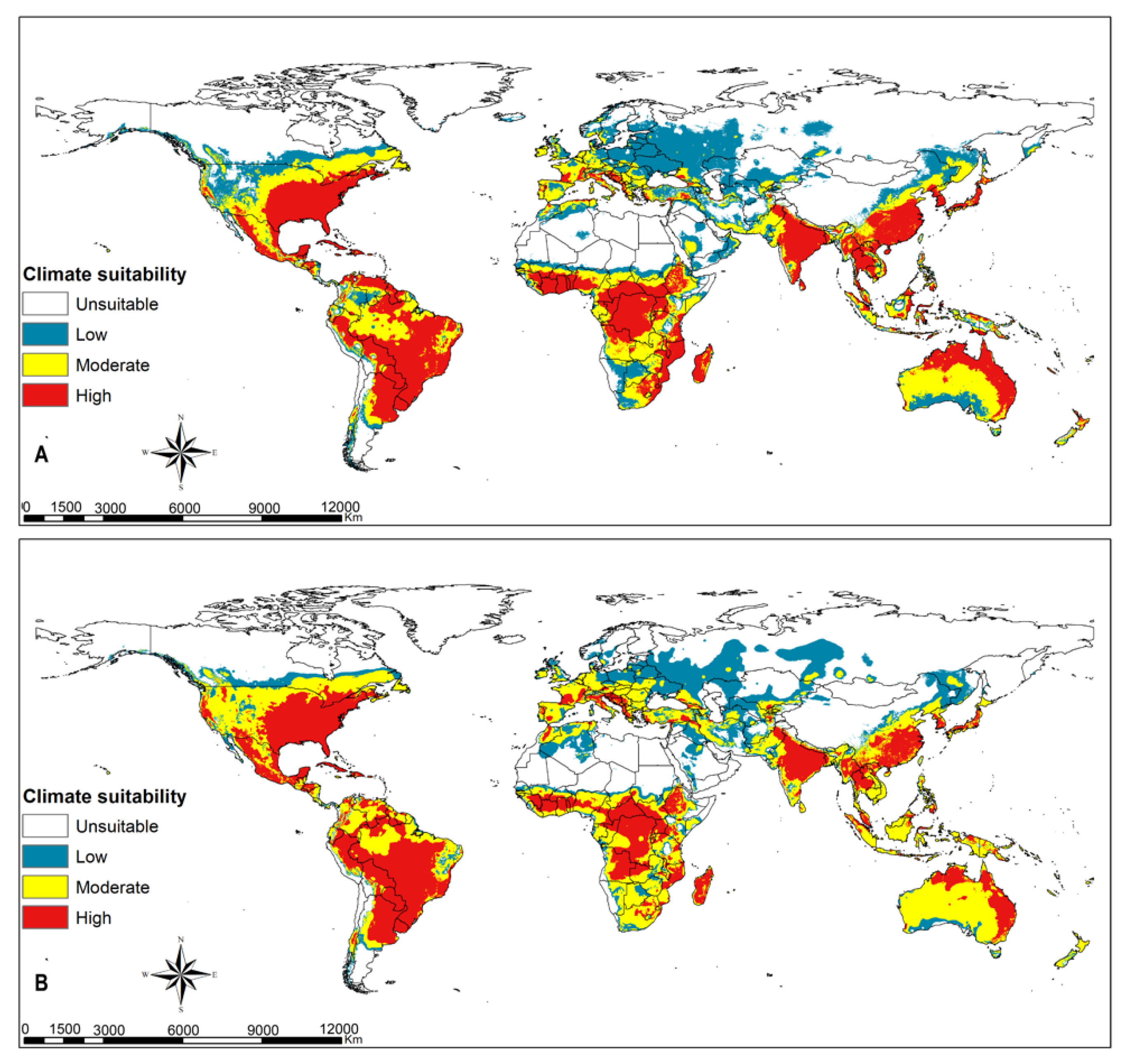
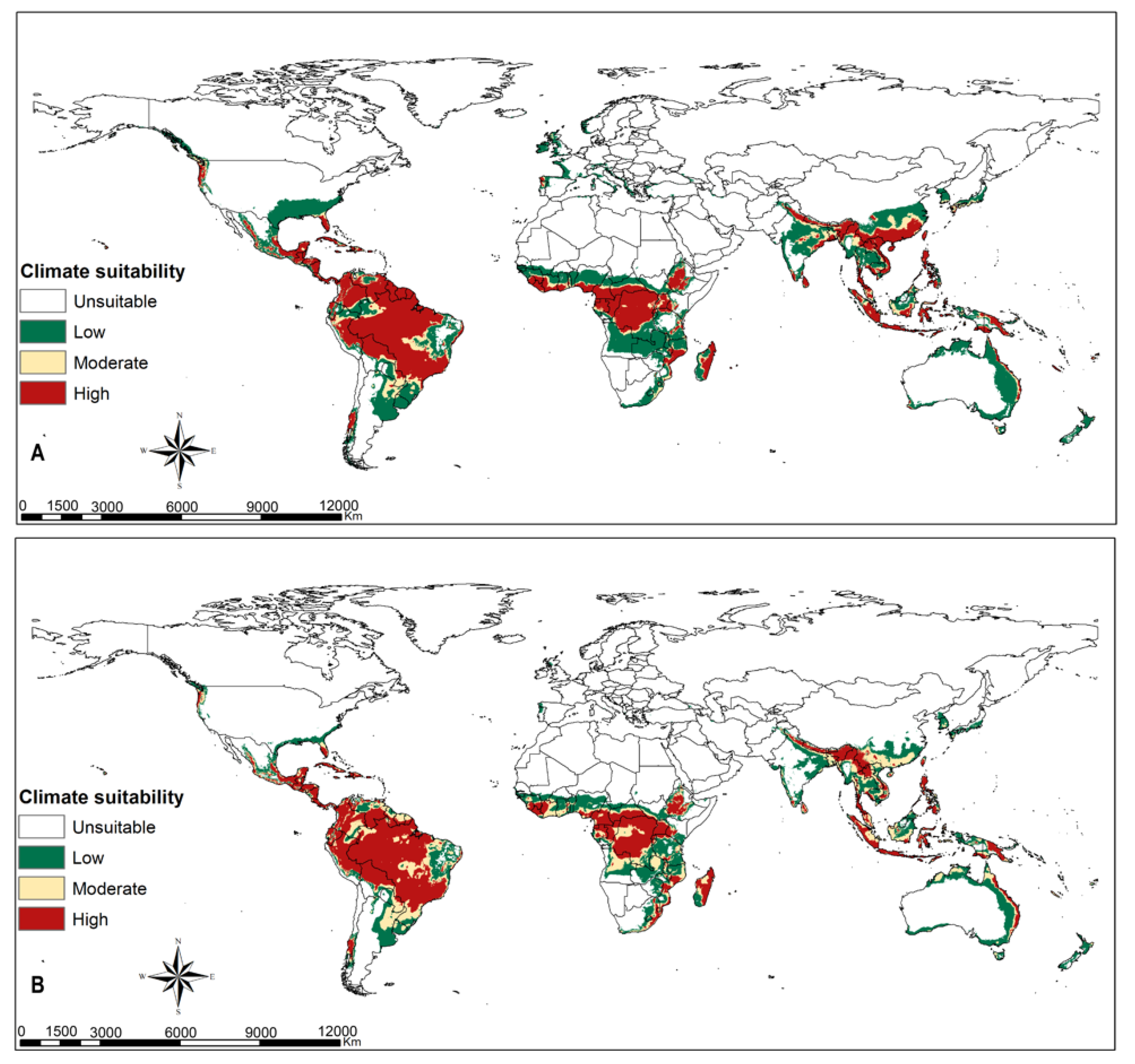
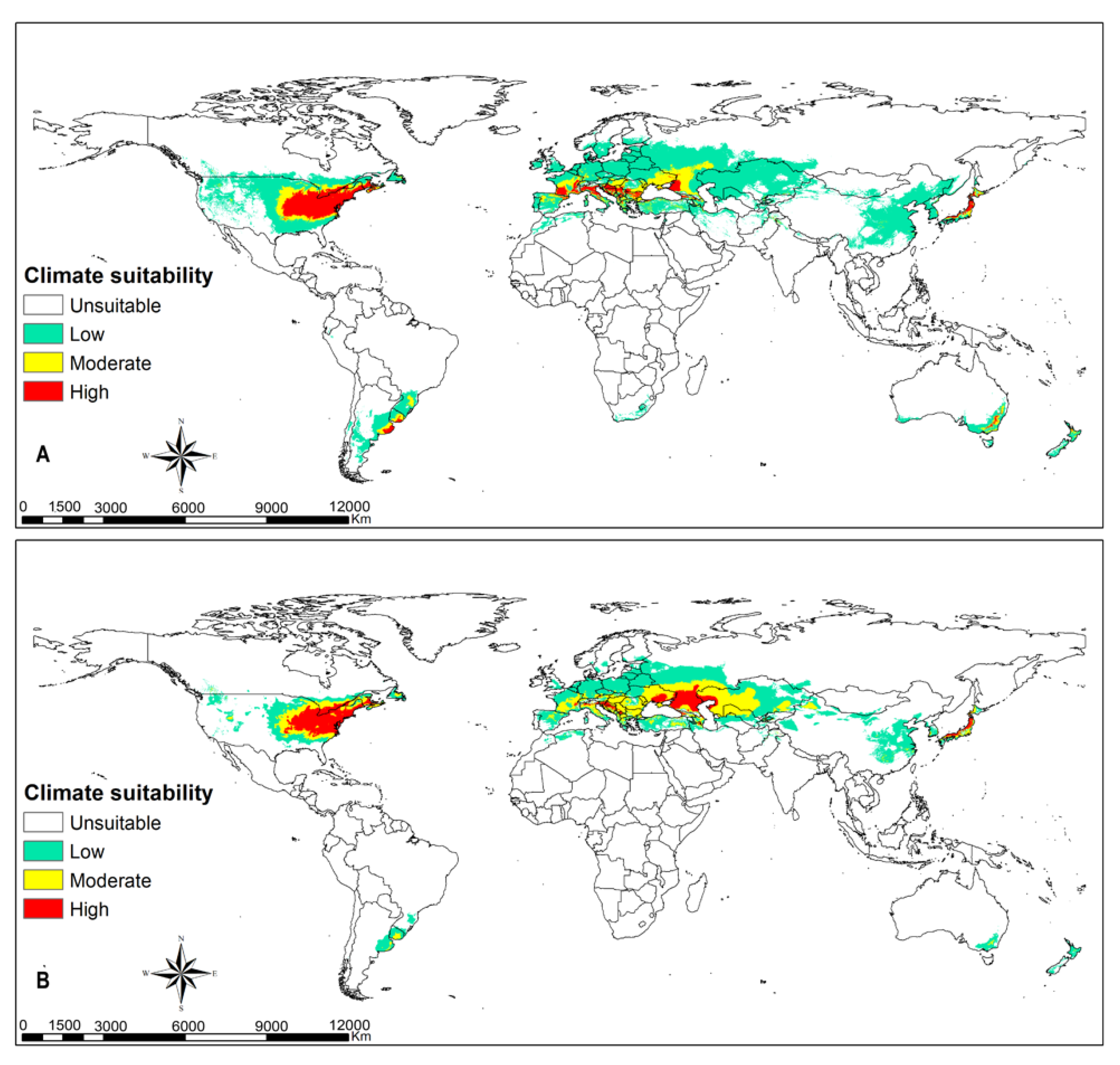
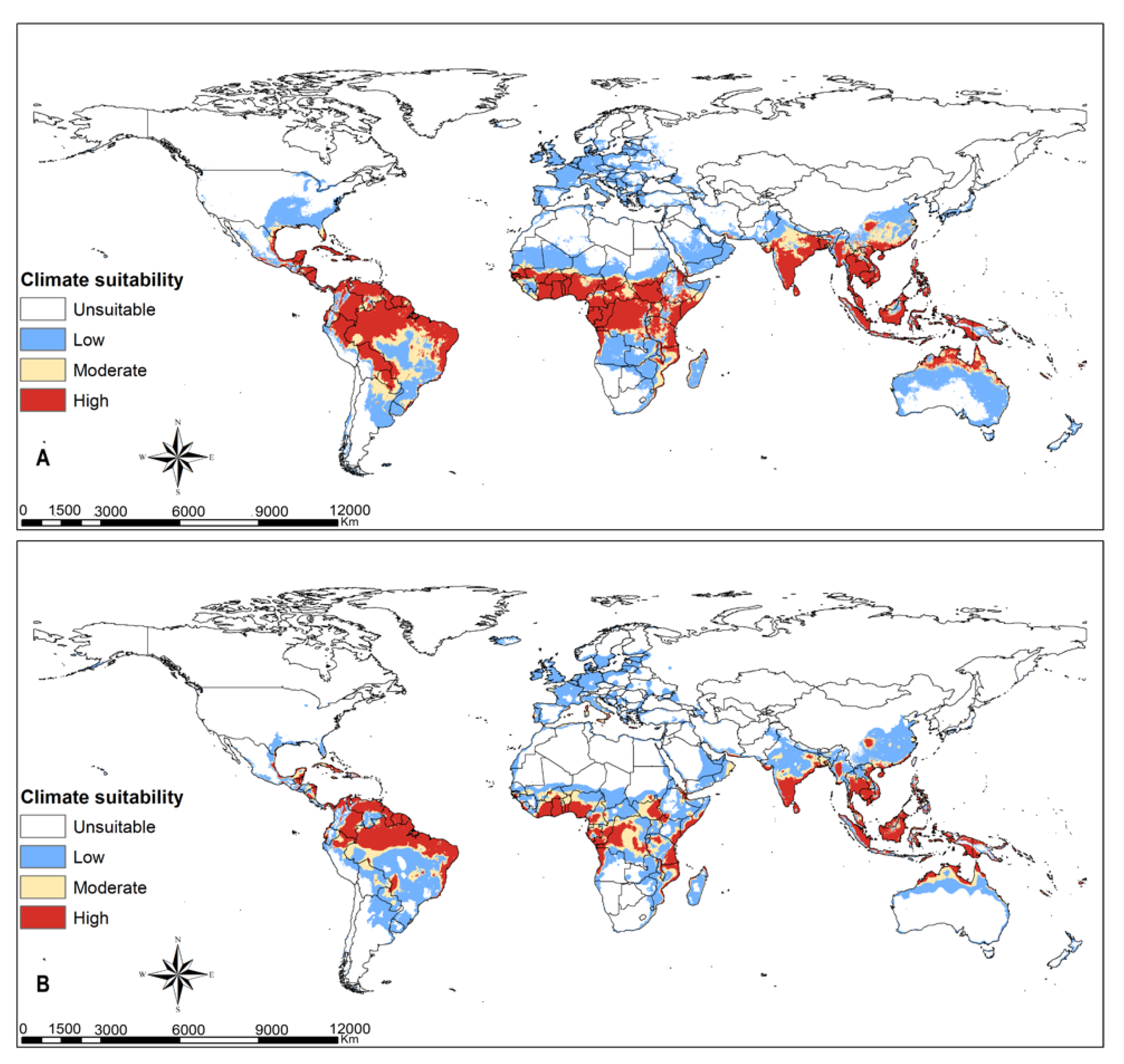
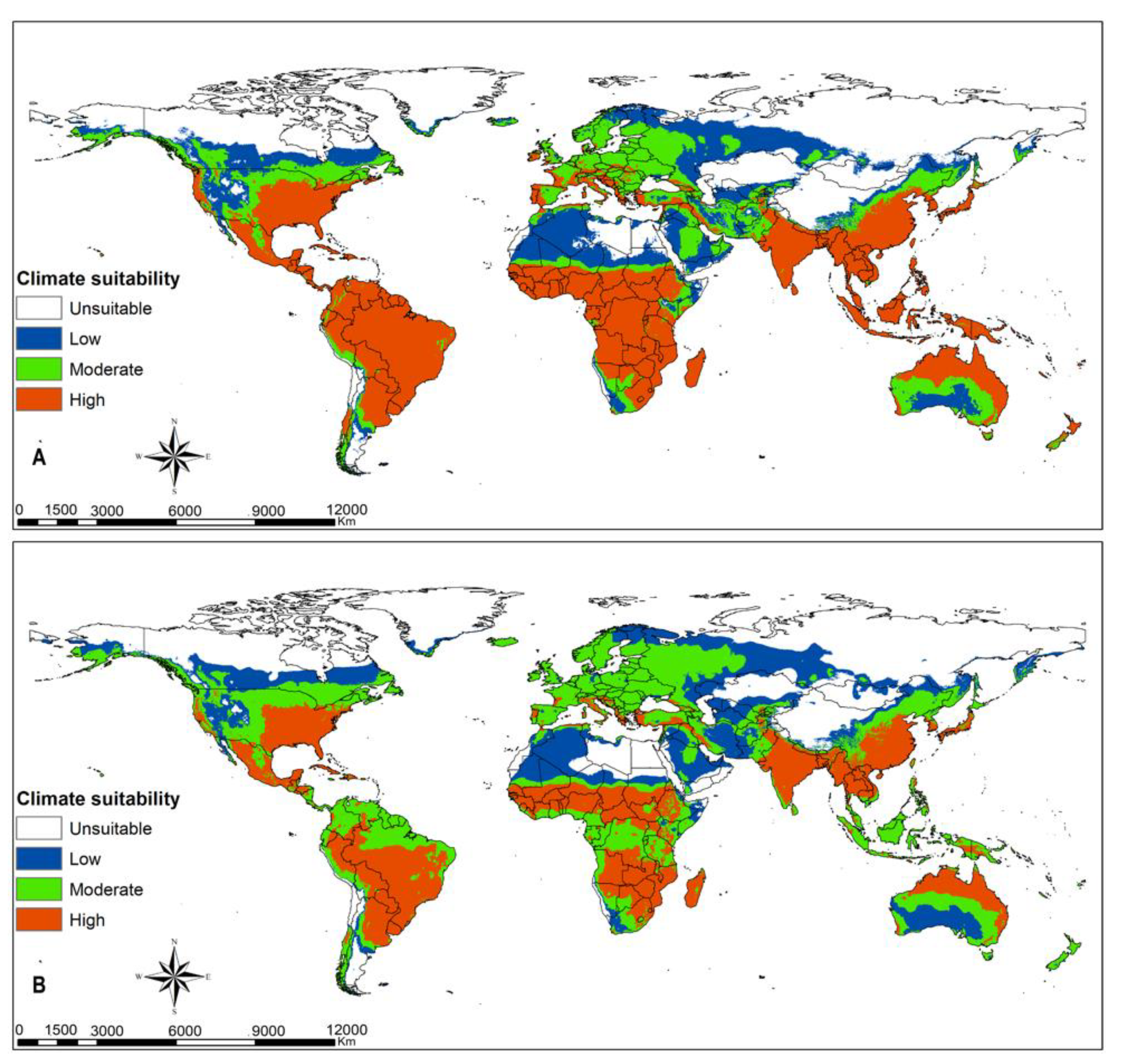
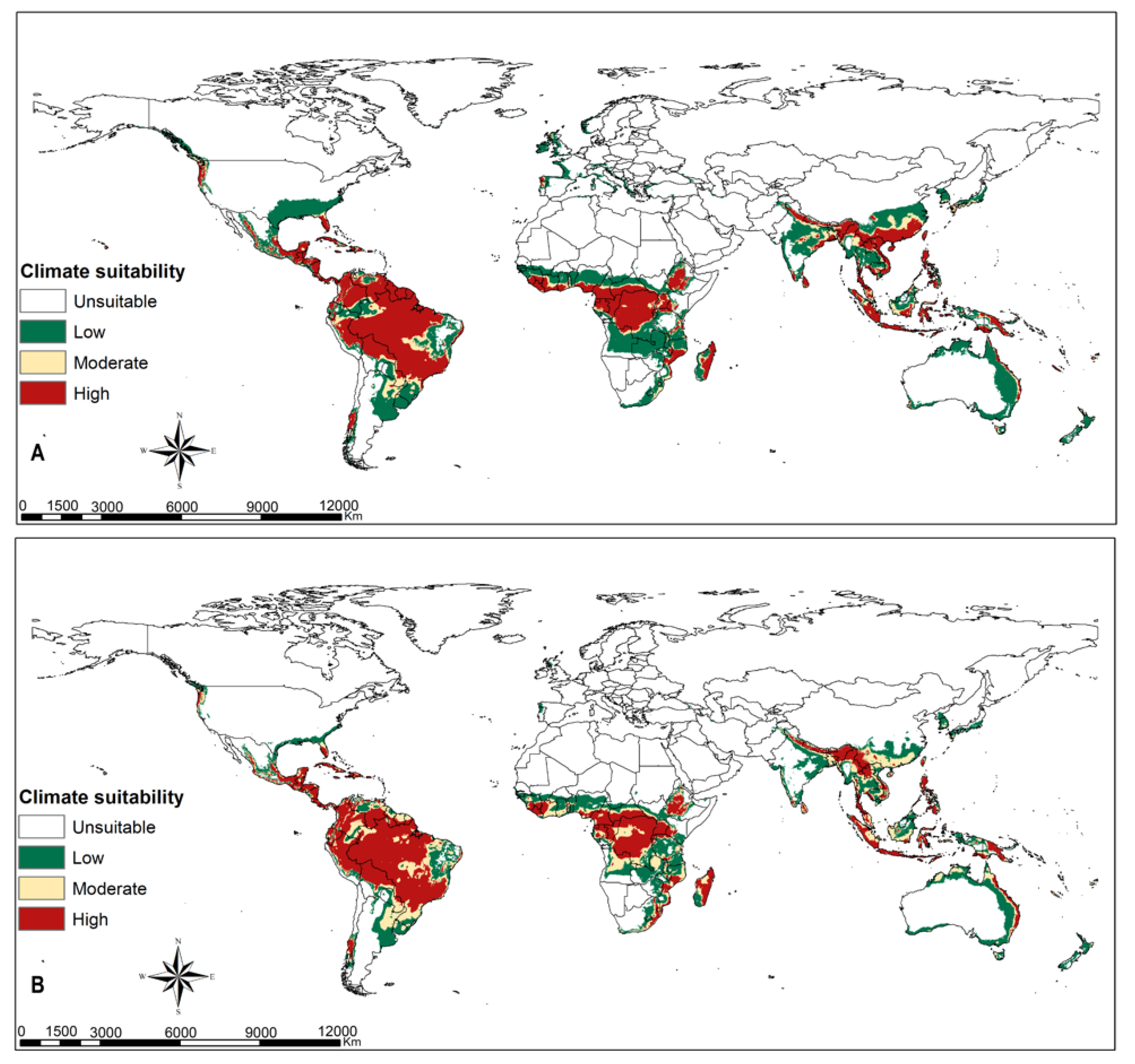
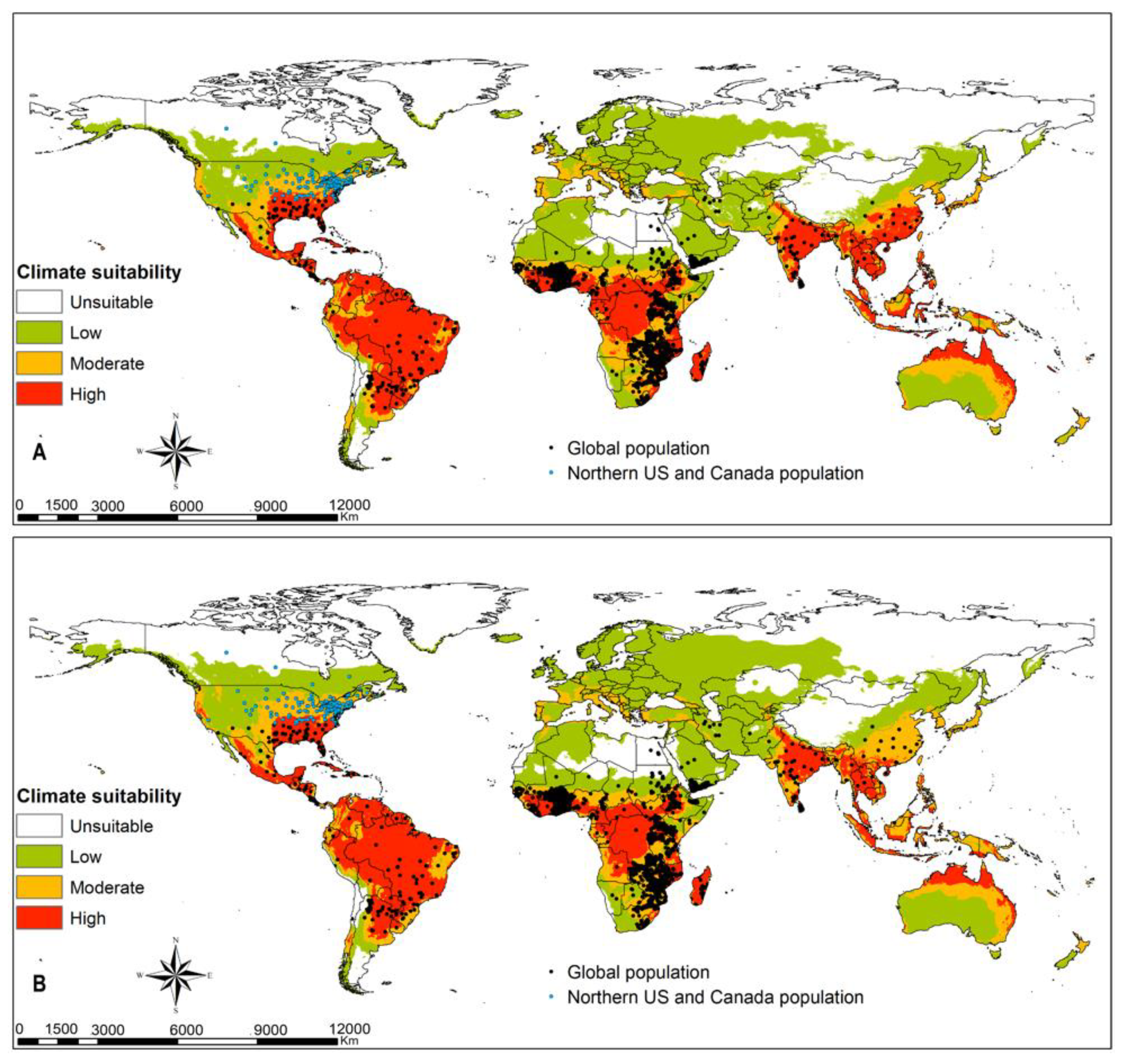
3.4. Parasitoids Habitat Suitability
4. Discussion
4.1. Model Evaluations
4.2. FAW Habitat Suitability
4.3. Parasitoid Habitat Suitability
4.4. Parasitoids Potential for Biocontrol
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Paini, D.R.; Sheppard, A.W.; Cook, D.C.; De Barro, P.J.; Worner, S.P.; Thomas, M.B. Global threat to agriculture from invasive species. Proc. Natl. Acad. Sci. USA 2016, 113, 7575–7579. [Google Scholar] [CrossRef]
- Montezano, D.G.; Specht, A.; Sosa-Gómez, D.R.; Roque-Specht, V.F.; Sousa-Silva, J.C.; Paula-Moraes, S.D.; Peterson, J.A.; Hunt, T.E. Host Plants of Spodoptera frugiperda (Lepidoptera: Noctuidae) in the Americas. Afr. Entomol. 2018, 26, 286–300. [Google Scholar] [CrossRef]
- Goergen, G.; Kumar, P.L.; Sankung, S.B.; Togola, A.; Tamò, M. First Report of Outbreaks of the Fall Armyworm Spodoptera frugiperda (J E Smith) (Lepidoptera, Noctuidae), a New Alien Invasive Pest in West and Central Africa. PLoS ONE 2016, 11, e0165632. [Google Scholar] [CrossRef]
- Sharanabasappa; Kalleshwaraswamy, C.M.; Asokan, R.; Swamy, H.M.M.; Maruthi, M.S.; Pavithra, H.B.; Kavita, H.; Shivaray, N.; Prabhu, S.T.; Goergen, G. First report of the Fall armyworm, Spodoptera frugiperda (J E Smith) (Lepidoptera: Noctuidae), an alien invasive pest on maize in India. Pest Manag. Hortic. Ecosyst. 2018, 24, 23–29. [Google Scholar]
- FAO. Fall Armyworm: Map of the Worldwide Spread of Fall Armyworm Since 2016 (as of March 2020), Compiled Using Information from a Range of Sources, Including FAO, International Plant Protection Convention, CABI, the European and Mediterranean Plant Protection Organization, and National Governments. Published online at Fao.org. 2020. Available online: http://www.fao.org/fall-armyworm/monitoring-tools/faw-map/en/ (accessed on 7 December 2020).
- CABI. Spodoptera frugiperda (Fall Armyworm), Invasive Species Compendium 2020: Datasheet 29810. Available online: https://www.cabi.org/isc/datasheet/29810#96ab1f68-7d15-4147-a6e3-4840893c75e6 (accessed on 15 January 2020).
- Malo, M.; Hore, J. The emerging menace of fall armyworm (Spodoptera frugiperda JE Smith) in maize: A call for attention and action. J. Entomol. Zool. Stud. 2020, 8, 455–465. [Google Scholar]
- Early, R.; González-Moreno, P.; Murphy, S.T.; Day, R. Forecasting the global extent of invasion of the cereal pest Spodoptera frugiperda, the fall armyworm. NeoBiota 2018, 40, 25–50. [Google Scholar] [CrossRef]
- Du Plessis, H.; Van den Berg, J.; Kriticos, D.J.; Ota, N. Spodoptera frugiperda (Fall Armyworm), CLIMEX Modelling. CSIRO-InSTePP Pest Geography 2018. ResearchGate. 2018 Sept. Available online: https://www.researchgate.net/publication/327423712_Spodoptera_frugiperde_-_CLIMEX_modelling_du_Plessis_van_den_Berg_Ota_Kriticos_2018_CSIRO-InSTePP_Pest_Geography_June_2018_Canberra (accessed on 7 April 2020).
- Jeger, M.; Bragard, C.; Caffier, D.; Candresse, T.; Chatzivassiliou, E.; Dehnen-Schmutz, K.; Gilioli, G.; Gregoire, J.; Miret, J.A.J.; Navarro, M.N.; et al. Scientific Opinion on the pest categorisation of Spodoptera frugiperda. EFSA J. 2017, 15, e04927. [Google Scholar] [CrossRef]
- Ferreira, F.J.B.S.; Alves, L.; Gottardo, L.; Georgino, M. Dimensionamento do custo econômico representado por Spodoptera frugiperda na cultura do milho no Brasil. In Proceedings of the 48 Congresso Sociedade Brasileira de Economia, Campo Grande, Brazil, 25–28 July 2010; Administração e Sociologia Rural: Campo Grande, Brazil, 2010; p. 21. [Google Scholar]
- Rwomushana, I.; Bateman, M.; Beale, T.; Beseh, P.; Cameron, K.; Chiluba, M.; Clottey, V.; Davis, T.; Day, R.; Early, R.; et al. Fall Armyworm: Impacts and Implications for Africa; Evidence Note Update; CABI: London, UK, 2018; p. 51. [Google Scholar]
- Day, R.; Abrahams, P.; Bateman, M.; Beale, T.; Clottey, V.; Cock, M.; Colmenarez, Y.; Corniani, N.; Early, R.; Godwin, J.; et al. Fall armyworm: Impacts and implications for Africa. Outlooks Pest Manag. 2017, 28, 196–201. [Google Scholar] [CrossRef]
- Molina-Ochoa, J.; Carpenter, J.E.; Heinrichs, E.A.; Foster, J.E. Parasitoids and parasites of Spodoptera frugiperda (Lepidoptera: Noctuidae) in the Americas and Caribbean Basin: An inventory. Fla. Entomol. 2003, 86, 254–289. [Google Scholar] [CrossRef]
- Cruz, I.; Bruce, A.; Sevgan, S.; Akutse, K.S.; Mohamed, F.S.; Niassy, S.; Rangaswamy, M.; Sidhu, J.; Goergen, G.; Rwomushana, I.; et al. Biological Control and Biorational Pesticides for Fall Armyworm Management. In Fall Armyworm in Africa: A Guide for Integrated Pest Management; Prasanna, B.M., Huesing, J.E., Eddy, R., Peschke, V.M., Eds.; CIMMYT: Mexico City, Mexico, 2018; pp. 63–88. [Google Scholar]
- Figueredo, M.C.; Cruz, I.; Martins-Dias, A.M.P.; da Silva, R.B. Associação entre inimigos naturais e Spodoptera frugiperda (J.E. Smith, 1797) (Lepidoptera: Noctuidae) na cultura do milho. Rev. Bras. Milho Sorgo 2006, 5, 340–350. [Google Scholar] [CrossRef]
- Shimbori, E.M.; Onody, H.C.; Fernandes, D.R.R.; Silvestre, R.; Tavares, M.T.; Penteado-Dias, A.M. Hymenoptera “Parasitica” in the state of Mato Grosso do Sul, Brazil. Iheringia Sér Zool. 2017, 107, 1–12. [Google Scholar] [CrossRef]
- Hajek, A.E. Natural Enemies: An Introduction to Biological Control; Cambridge University Press: Cambridge, MA, USA, 2004. [Google Scholar]
- Zanuncio, J.C.; Matos Neto, F.D.C.; Tavares, W.D.S.; Cruz, I.; Leite, G.L.D.; Serrão, J.E. Functional and numerical responses and reproduction of Campoletis flavicincta parasitizing Spodoptera frugiperda caterpillars. Acta Sci. Agron. 1933, 35, 419–426. [Google Scholar] [CrossRef]
- Pomari-Fernandes, A.; Bueno, A.D.F.; Bueno, R.C.F.; Junior, M.; de Oliveiras, A.; Fonseca, A.C. Releasing number of Telenomus remus (Nixon) (Hymenoptera: Platygastridae) against Spodoptera frugiperda Smith (Lepidoptera: Noctuidae) in corn, cotton and soybean. Ciênc. Rural 2013, 43, 377–382. [Google Scholar] [CrossRef]
- Pomari-Fernandes, A.; Bueno, A.F.; De Bortoli, S.A.; Favetti, B.M. Dispersal capacity of the egg parasitoid Telenomus remus Nixon (Hymenoptera: Platygastridae) in maize and soybean crops. Biol. Control 2018, 126, 158–168. [Google Scholar] [CrossRef]
- Escobar-Ramírez, S.; Grass, I.; Armbrecht, I.; Tscharntke, T. Biological control of the coffee berry borer: Main natural enemies, control success, and landscape influence. Biol. Control 2019, 136, 103992. [Google Scholar] [CrossRef]
- Harrison, R.D.; Thierfelder, C.; Baudron, F.; Chinwada, P.; Midega, C.; Schaffner, U.; van den Berg, J. Agro-ecological options for fall armyworm (Spodoptera frugiperda JE Smith) management: Providing low-cost, smallholder friendly solutions to an invasive pest. J. Environ. Manag. 2019, 243, 318–330. [Google Scholar] [CrossRef]
- Elith, J.; Graham, C.H.; Anderson, R.P.; Dudik, M.; Ferrier, S.; Guisan, A.; Hijmans, R.J.; Huettmann, F.; Leathwick, J.R.; Lehmann, A.; et al. Novel methods improve prediction of species’ distributions from occurrence data. Ecography 2006, 29, 129–151. [Google Scholar] [CrossRef]
- Elith, J.; Phillips, S.J.; Hastie, T.; Dudík, M.; Chee, Y.E.; Yates, C.J. A statistical explanation of maxent for ecologists. Divers. Distrib. 2011, 17, 43–57. [Google Scholar] [CrossRef]
- Phillips, S.; Anderson, R.; Schapire, R. Maximum entropy modelling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef]
- GBIF. Occurrence Download. 2018. Available online: https://www.gbif.org/occurrence/download/0001963-181108115102211 (accessed on 15 January 2020).
- CABI. Crop Protection Compendium; CAB International: Wallingford, UK, 2020; Available online: https://www.cabi.org/cpc/ (accessed on 15 January 2020).
- Kenis, M.; Du Plessis, H.; Van den Berg, J.; Ba, M.N.; Goergen, G.; Kwadjo, K.E.; Baoua, I.; Tefera, T.; Buddie, A.; Cafà, G.; et al. Telenomus remus, a Candidate Parasitoid for the Biological Control of Spodoptera frugiperda in Africa, is already Present on the Continent. Insects 2019, 10, 92. [Google Scholar] [CrossRef] [PubMed]
- Agboyi, L.K.; Goergen, G.; Beseh, P.; Mensah, S.A.; Clottey, V.A.; Glikpo, R.; Buddie, A.; Cafà, G.; Offord, L.; Day, R.; et al. Parasitoid complex of fall armyworm, Spodoptera frugiperda, in Ghana and Benin. Insects 2020, 11, 68. [Google Scholar] [CrossRef]
- Liao, Y.L.; Yang, B.; Xu, M.F.; Lin, W.; Wang, D.S.; Chen, K.W.; Chen, H.Y. First report of Telenomus remus parasitizing Spodoptera frugiperda and its field parasitism in southern China. J. Hymenopt Res. 2019, 73, 95–102. [Google Scholar] [CrossRef]
- Fick, S.E.; Hijmans, R.J. Worldclim 2: New 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Hijmans, R.J.; Cameron, S.E.; Parra, J.L.; Jones, P.G.; Jarvis, A. Very high resolution interpolated climate surfaces for global land areas. Int. J. Climatol. 2005, 25, 1965–1978. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing. 2020. Available online: https://www.R-project.org/ (accessed on 5 March 2021).
- Alsamadisi, A.G.; Tran, L.T.; Papeş, M. Employing inferences across scales: Integrating spatial data with different resolutions to enhance Maxent models. Ecol. Model. 2020, 415, 108857. [Google Scholar] [CrossRef]
- Ruane, A.C.; McDermid, S.P. Selection of a representative subset of global climate models that captures the profile of regional changes for integrated climate impacts assessment. Earth Perspect. 2017, 4, 1. [Google Scholar] [CrossRef]
- Qin, A.; Liu, B.; Guo, Q.; Bussmann, R.W.; Ma, F.; Jian, Z.; Xu, G.; Pei, S. Maxent modeling for predicting impacts of climate change on the potential distribution of Thuja sutchuenensis Franch., an extremely endangered conifer from southwestern China. Glob. Ecol. Conserv. 2017, 10, 139–146. [Google Scholar] [CrossRef]
- Biber-Freudenberger, L.; Ziemacki, J.; Tonnang, H.E.Z.; Borgemeister, C. Future Risks of Pest Species under Changing Climatic Conditions. PLoS ONE 2016, 11, e0153237. [Google Scholar] [CrossRef] [PubMed]
- Pearson, R.G.; Dawson, T.P. Predicting the impacts of climate change on the distribution of species: Are bioclimate envelope models useful? Glob. Ecol. Biogeogr. 2003, 12, 361–371. [Google Scholar] [CrossRef]
- Stireman, J.O.; Dyer, L.A.; Janzen, D.H.; Singer, M.S.; Lill, J.T.; Marquis, R.J.; Ricklefs, R.E.; Gentry, G.L.; Hallwachs, W.; Coley, P.D.; et al. Climatic unpredictability and parasitism of caterpillars: Implications of global warming. Proc. Natl. Acad. Sci. USA 2005, 102, 17384–17387. [Google Scholar] [CrossRef]
- Thomson, L.J.; Macfadyen, S.; Hoffmann, A.A. Predicting the effects of climate change on natural enemies of agricultural pests. Biol. Control 2010, 52, 296–306. [Google Scholar] [CrossRef]
- Hannah, L.; Midgley, G.F.; Millar, D. Climate change integrated conservation strategies. Glob. Ecol. Biogeogr. 2002, 11, 485–495. [Google Scholar] [CrossRef]
- IPPC. Report of First Detection of Spodoptera frugiperda—Fall Armyworm (FAW) in Egypt. 2019. Available online: https://www.ippc.int/en/countries/egypt/pestreports/2019/06/report-of-first-detection-of-spodoptera-frugiperda-fall-armyworm-faw-in-egypt/ (accessed on 17 April 2020).
- Durocher-Granger, L.; Babendreier, D.; Huesing, J.E.; Jepson, P.C.; Eddy, R.; Prassana, B.M. Fall Armyworm (FAW) on Maize in Mauritania. 2018. Available online: https://www.cabdirect.org/cabdirect/abstract/20197800136 (accessed on 15 January 2020).
- IPPC. Detections of Spodoptera frugiperda (Fall Armyworm) on Mainland Australia. 2020. Available online: https://www.ippc.int/en/countries/australia/pestreports/2020/03/detections-of-spodoptera-frugiperda-fall-armyworm-on-mainland-australia/ (accessed on 17 April 2020).
- WFP. WFP Timor-Leste Country Brief February 2020. Available online: https://reliefweb.int/sites/reliefweb.int/files/resources/WFP-0000113450.pdf (accessed on 17 April 2020).
- Du Plessis, H.; Schlemmer, M.-L.; Van den Berg, J. The Effect of Temperature on the Development of Spodoptera frugiperda (Lepidoptera: Noctuidae). Insects 2020, 11, 228. [Google Scholar] [CrossRef]
- Bueno, R.F.; Carneiro, T.R.; Bueno, A.F.; Pratissoli, D.; Fernandes, O.A.; Vieira, S.S. Parasitism capacity of Telenomus remus Nixon (Hymenoptera: Scelionidae) on Spodoptera frugiperda (Smith) (Lepidoptera: Noctuidae) eggs. Braz. Arch. Biol. Technol. 2010, 53, 133–139. [Google Scholar] [CrossRef]
- Carneiro, T.R.; Fernandes, O.A. Interspecific interaction between Telenomus remus (Hymenoptera: Platygastridae) and Trichogramma pretiosum (Hymenoptera: Trichogrammatidae) on Spodoptera frugiperda (Lepidoptera: Noctuidae) eggs. An. Acad. Bras. Ciênc. 2012, 84, 1127–1135. [Google Scholar] [CrossRef]
- Díaz, M.F.; Ramírez, A.; Poveda, K. Efficiency of different egg parasitoids and increased floral diversity for the biological control of noctuid pests. Biol. Control 2012, 60, 182–191. [Google Scholar] [CrossRef]
- Sisay, B.; Simiyu, J.; Mendesil, E.; Likhayo, P.; Ayalew, G.; Mohamed, S.; Subramanian, S.; Tefera, T. Fall armyworm, Spodoptera frugiperda infestations in East Africa: Assessment of damage and parasitism. Insects 2019, 10, 195. [Google Scholar] [CrossRef]
- Cock, M.J.W. Review of Biological Control of Pests in the Commonwealth Caribbean and Bermuda up to 1982. CAB CIBC Tech. Commun. 1985, 9, 218. [Google Scholar]
- Bueno, R.F.; Bueno, A.D.F.; Parra, J.R.P.; Vieira, S.S.; Oliveira, L.J.D. Biological characteristics and parasitism capacity of Trichogramma pretiosum Riley (Hymenoptera, Trichogrammatidae) on eggs of Spodoptera frugiperda (JE Smith) (Lepidoptera, Noctuidae). Rev. Bras. Entomol. 2010, 54, 322–327. [Google Scholar] [CrossRef]
- Chapman, J.W.; Williams, T.; Escribano, A.; Caballero, P.; Cave, R.D.; Goulson, D. Fitness consequences of cannibalism in the fall armyworm, Spodoptera frugiperda. Behav. Ecol. 1999, 10, 298–303. [Google Scholar] [CrossRef]
- Gauld, I.D. The Ichneumonidae of Costa Rica, 3; Memoirs of the American Entomological Institute; American Entomological Institute: Gainesville, FL, USA, 2000; Volume 63, pp. 1–453. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| GCM | Institution | Horizontal Resolution | 2× [CO2] Equilibrium Climate Sensitivity (°C) |
|---|---|---|---|
| GISS-E2-R * | National Aeronautics and Space Association Goddard Institute for Space Studies (NASA GISS) | 2° × 2.5° | 2.1 |
| HadGEM2-ES * | UK Meteorological Office—Hadley Centre | 1.25° × 1.875° | 4.6 |
| Spodoptera frugiperda | Current | RCP8.5 | ||
|---|---|---|---|---|
| Training | Test | Training | Test | |
| Native and invaded regions model | 0.929 | 0.925 | 0.902 | 0.913 |
| Temporary migrated regions model | 0.952 | 0.942 | 0.951 | 0.931 |
| Parasitoid Species | Current | RCP8.5 | ||
|---|---|---|---|---|
| Training | Test | Training | Test | |
| Chelonus insularis | 0.934 | 0.953 | 0.920 | 0.941 |
| Cotesia marginiventris | 0.900 | 0.946 | 0.869 | 0.958 |
| Eiphosoma laphygmae | 0.969 | 0.971 | 0.965 | 0.966 |
| Telenomus remus | 0.931 | 0.956 | 0.916 | 0.917 |
| Trichogramma pretiosum | 0.856 | 0.864 | 0.802 | 0.807 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tepa-Yotto, G.T.; Tonnang, H.E.Z.; Goergen, G.; Subramanian, S.; Kimathi, E.; Abdel-Rahman, E.M.; Flø, D.; Thunes, K.H.; Fiaboe, K.K.M.; Niassy, S.; et al. Global Habitat Suitability of Spodoptera frugiperda (JE Smith) (Lepidoptera, Noctuidae): Key Parasitoids Considered for Its Biological Control. Insects 2021, 12, 273. https://doi.org/10.3390/insects12040273
Tepa-Yotto GT, Tonnang HEZ, Goergen G, Subramanian S, Kimathi E, Abdel-Rahman EM, Flø D, Thunes KH, Fiaboe KKM, Niassy S, et al. Global Habitat Suitability of Spodoptera frugiperda (JE Smith) (Lepidoptera, Noctuidae): Key Parasitoids Considered for Its Biological Control. Insects. 2021; 12(4):273. https://doi.org/10.3390/insects12040273
Chicago/Turabian StyleTepa-Yotto, Ghislain T., Henri E. Z. Tonnang, Georg Goergen, Sevgan Subramanian, Emily Kimathi, Elfatih M. Abdel-Rahman, Daniel Flø, Karl H. Thunes, Komi K. M. Fiaboe, Saliou Niassy, and et al. 2021. "Global Habitat Suitability of Spodoptera frugiperda (JE Smith) (Lepidoptera, Noctuidae): Key Parasitoids Considered for Its Biological Control" Insects 12, no. 4: 273. https://doi.org/10.3390/insects12040273