Description of the Nymphal Stages of Hoplopleura affinis (Anoplura: Hoplopleuridae) from the Striped Field Mouse Apodemus agrarius with a Global Checklist of Hoplopleura Species from the Genus Apodemus


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
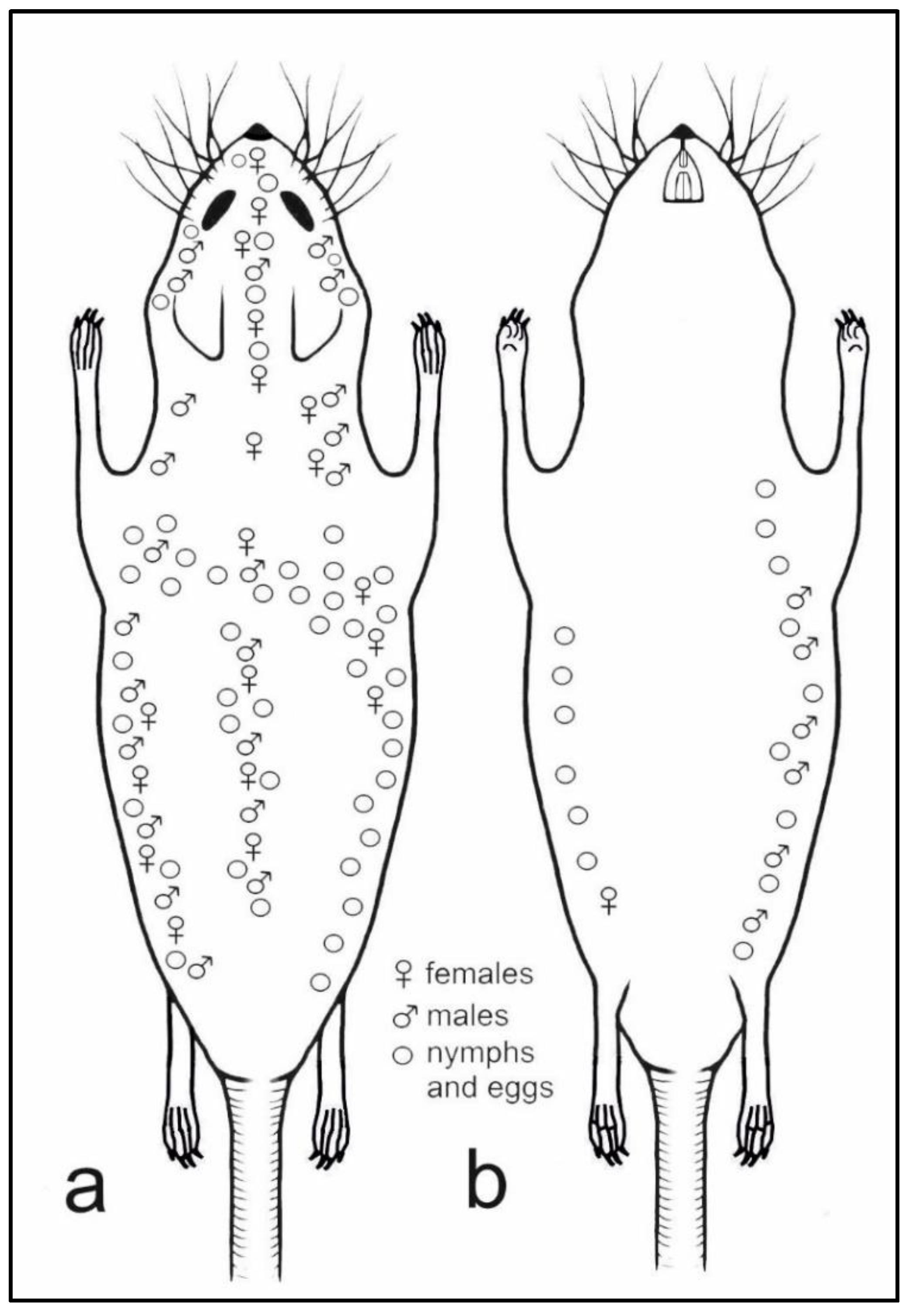
2.1. Lice and Host Material
- Specimens intended for analysis under a light microscope were prepared by slide-mounting in polyvinyl-lactophenol [15].
- Individuals intended for analysis with scanning electron microscopy were subjected to a series of alcohols (80–100%) and then dried in a mix of ethyl alcohol and hexamethyldisilazane (HMDS) in 1:3, 1:1, and 3:1 proportions. Finally, the specimens were transferred to pure HMDS and placed in an incubator for 24 h (37 °C) [16]. The specimens were stuck to double-sided copper tape (by Mierzejewski Materiały Samoprzylepne) and fixed on the table of a scanning electron microscope. Observations and photographs were performed with the use of Field Emission Scanning Electron Microscope JSM—7800F (manufacturer JEOL; stocked in Department of Materials Engineering and Bonding—Faculty of Mechanical Engineering, Gdańsk University of Technology, Gdańsk, Poland).
2.2. The Checklist
3. Results
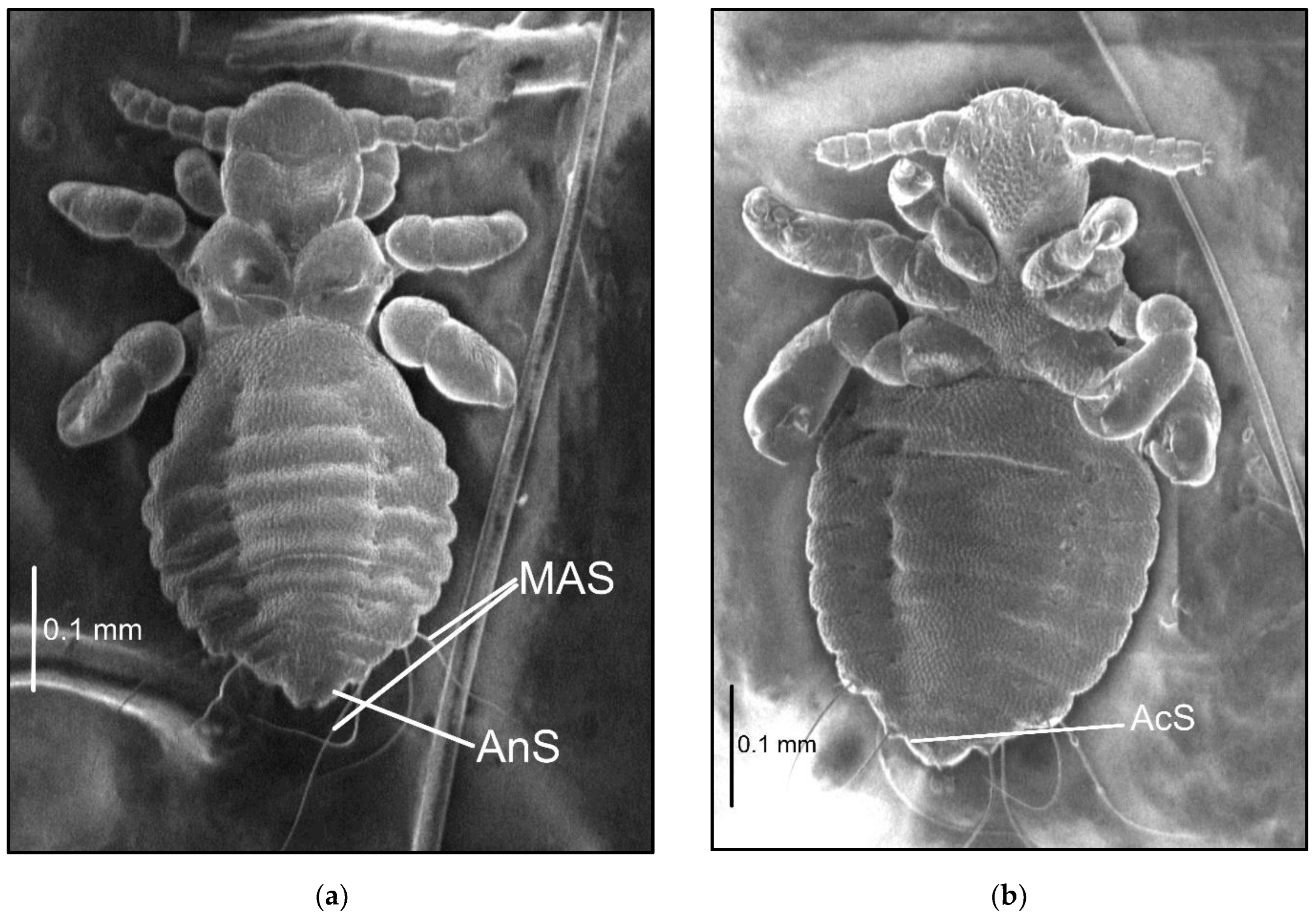
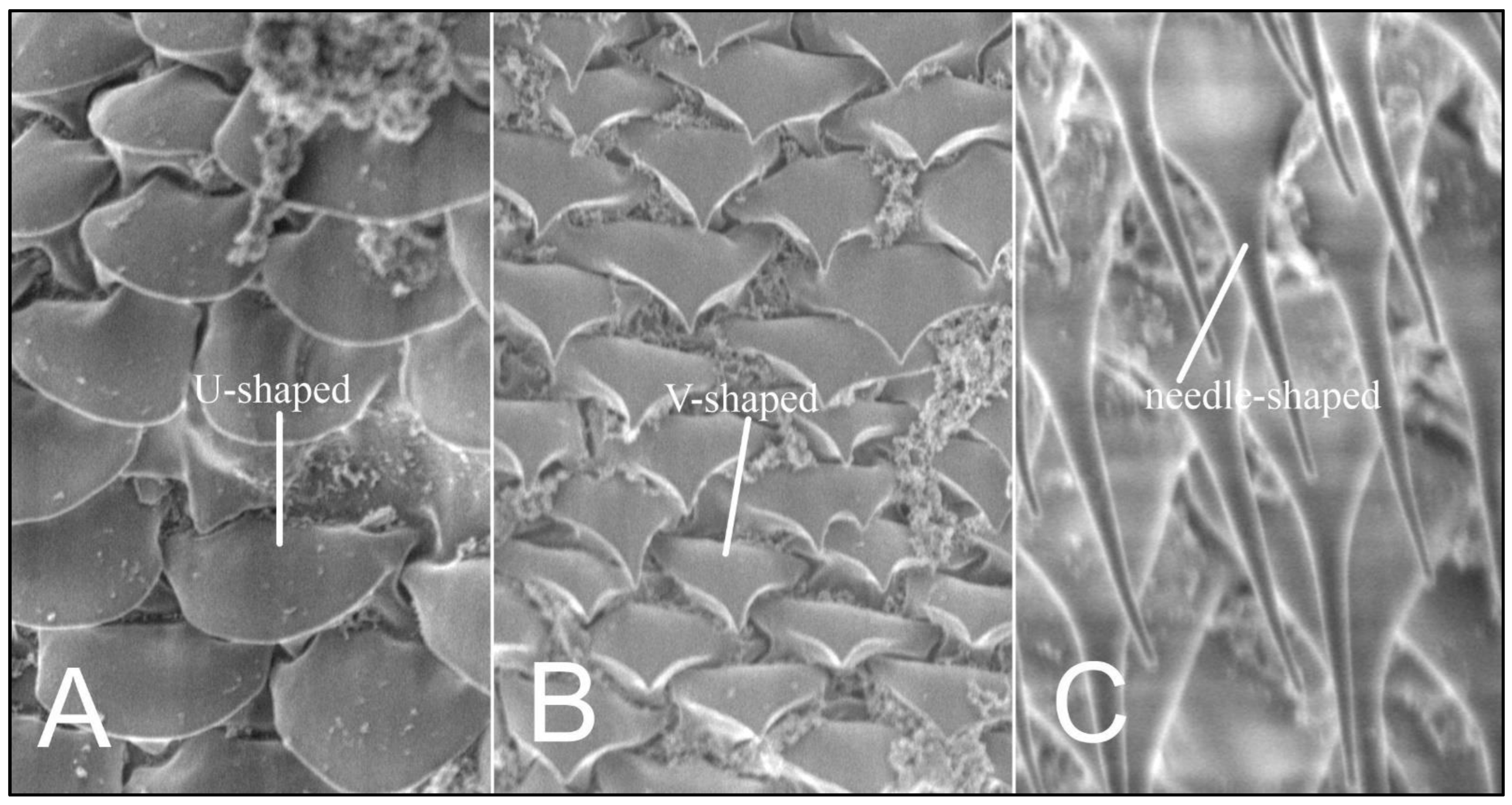
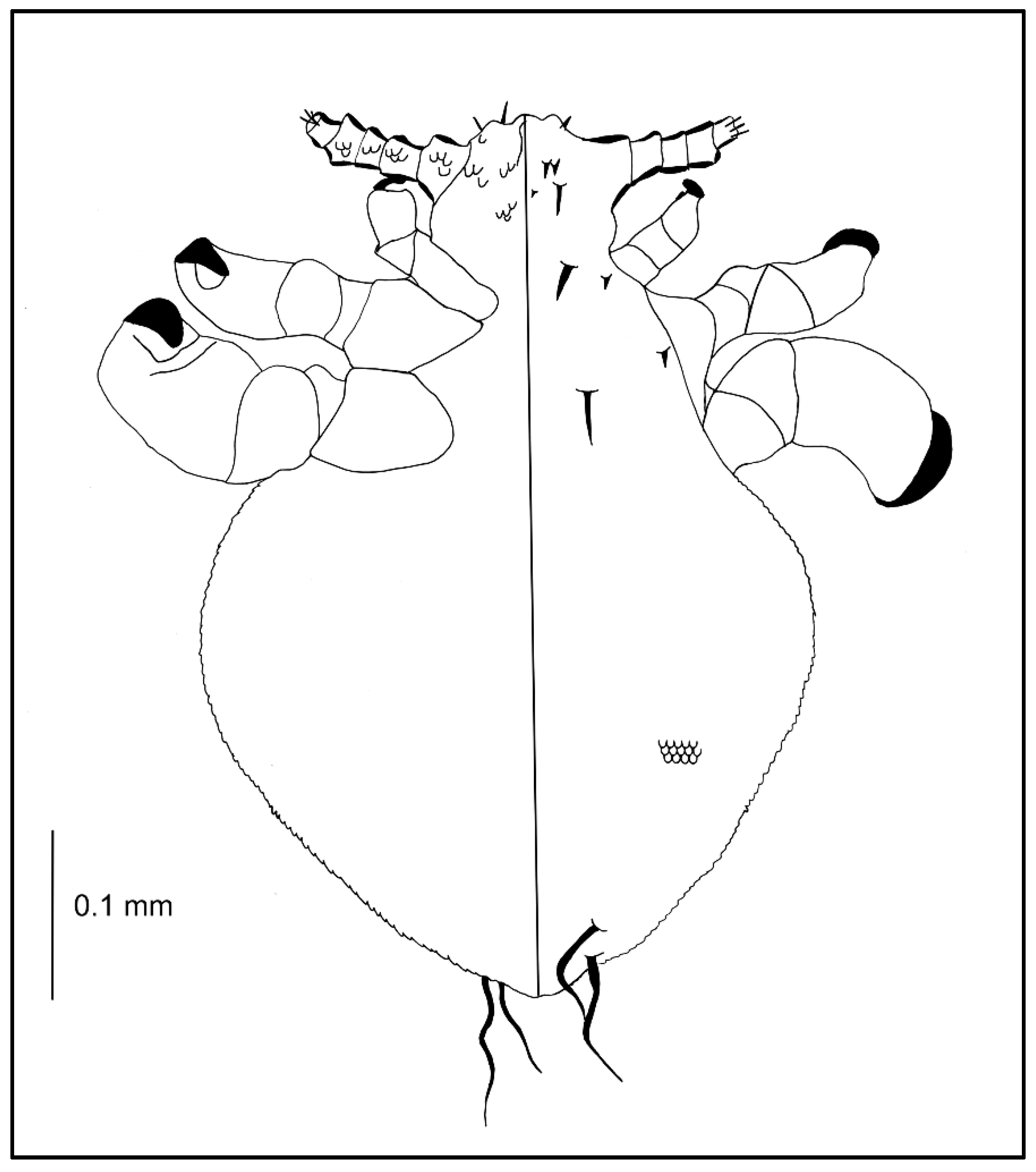
3.1. Description of Nymphal Stages
3.2. Checklist of Hoplopleura spp. on Murid Rodents of the Genus Apodemus
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kim, K.C.; Ludwig, H.W. Phylogenetic relationships of parasitic Psocodea and taxonomic position of the Anoplura. Ann. Entomol. Soc. 1978, 71, 910–922. [Google Scholar] [CrossRef]
- Durden, L.A.; Musser, G.G. The mammalian hosts of the sucking lice (Anoplura) of the world: A host–parasite list. Bull. Soc. Vector Ecol. 1994, 19, 130–168. [Google Scholar]
- Kim, K.C. Blood-sucking lice (Anoplura) of small mammals: True parasites. In Micromammals and Microparasits from Evolutionary to Management; Morand, S., Krasnov, B.R., Poulin, R., Eds.; Springer: Tokyo, Japan, 2006; pp. 141–160. [Google Scholar] [CrossRef]
- Durden, L.A.; Musser, G.G. The sucking lice (Insecta, anoplura) of the world: A taxonomic checklist with records of mammalian hosts and geographical distributions. Bull. Am. Nat. Hist. 1994, 218, 1–90. [Google Scholar]
- Edwards, S.J.; Hood, M.W.; Shaw, J.H.; Rayburn, J.D.; Kirby, M.D.; Hanfman, D.T.; Zidar, J.A. Index-Catalogue of Medical and Veterinary Zoology; Suppl. 21, Pt 5. Parasite-subject catalogue; Parasites: Arthropoda and miscellaneous phyla; United States Department of Agriculture. U.S. Government Printing Office: Washington, DC, USA, 1978; p. 246.
- Krištofík, J.; Lysy, J. Seasonal dynamics of sucking lice (Anoplura) in small mammals (Insectivora, Rodentia) in the natural foci of infections in South West Slovakia. Biológia 1992, 47, 605–617. [Google Scholar]
- Dik, B.; Selçuk, A.Y.; Kefelioğlu, H.; Keskin, A. A preliminary report of the sucking lice (Insecta: Phthiraptera: Anoplura) of some small mammals of Turkey with four new records. Trans. Am. Entomol. Soc. 2019, 145, 395–406. [Google Scholar] [CrossRef]
- Taxonomic Information System (ITIS). Available online: http://www.itis.gov (accessed on 5 November 2021).
- Wilson, D.E.; Reeder, D.M. Mammals Species of the World: A Taxonomic and Geographic Reference, 3rd ed.; The Johns Hopkins University Press: Baltimore, MD, USA, 2005; pp. 1–2142. [Google Scholar]
- Ge, D.; Feijó, A.; Cheng, J.; Lu, L.; Liu, R.; Abramov, A.V.; Xia, L.; Wen, Z.; Zhang, W.; Shi, L.; et al. Evolutionary history of field mice (Murinae: Apodemus), with emphasis on morphological variation among species in China and description of a new species. Zool. J. Linn. Soc. 2019, 187, 518–534. [Google Scholar] [CrossRef]
- Cook, E.F.; Beer, J.R. The immature stages of the genus Hoplopleura (Anoplura: Hoplopleuridae) in North America, with description of two new species. J. Parasitol. 1959, 45, 405–416. [Google Scholar] [CrossRef]
- Kim, K.C. A new species of Hoplopleura from Thailand, with notes and description of nymphal stages of Hoplopleura captiosa Johnson (Anoplura). Parasitology 1966, 56, 603–612. [Google Scholar] [CrossRef] [PubMed]
- Kozina, P.; Izdebska, J.N.; Kowalczyk, R. The first description of the nymphal stages of Hoplopleura longula (Psocodea: Anoplura: Hoplopleuridae) from the harvest mouse Micromys minutus (Rodentia: Muridae). Biodivers. Data J. 2021, 9, e63747. [Google Scholar] [CrossRef]
- Solon, J.; Borzyszkowski, J.; Bidłasik, M.; Richling, A.; Badora, K.; Balon, J.; Brzezińska-Wójcik, T.; Chabudziński, Ł.; Dobrowolski, R.; Grzegorczyk, I.; et al. Physico-geographical mesoregions of Poland: Verification and adjustment of boundaries on the basis of contemporary spatial data. Geogr. Pol. 2018, 91, 143–171. [Google Scholar] [CrossRef]
- Kadulski, S.; Izdebska, J.N. Methods used in studies of parasitic arthropods in mammals. In Arthropods. Epidemiological Importance; Buczek, A., Błaszak, C., Eds.; Koliber: Lublin, Poland, 2006; pp. 113–118. [Google Scholar]
- Murtey, M.D.; Ramasamy, P. Sample preparations for scanning electron microscopy–life sciences. In Modern Electron Microscopy in Physical and Life Sciences; Janecek, M., Kral, R., Eds.; IntechOpen: Rijeka, Croatia, 2016; pp. 161–185. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.Q. Repositories for mite and tick specimens: Acronyms and their nomenclature. Syst. Appl. Acarol. 2018, 23, 2432–2447. [Google Scholar] [CrossRef]
- Kaneko, K. Studies on the Murine Lice in Japan (Part 2). Description of Hoplopleura himenezumi n. sp. (Hoplopleuridae, Anoplura) collected from Apodemus argenteus argenteus. Bull. Tokyo Med. Dent. Univ. 1956, 3, 143. [Google Scholar]
- Zwolski, W. Badania nad ektoparazytofauną drobnych ssaków w ogniskach naturalnych gorączki błotnej. Wiad. Parazytol. 1960, 6, 519–527. [Google Scholar]
- Mishra, A.C.; Kulkarni, S.M.; Bhat, H.R. Hoplopleura himalayana, sp. nov. (Anoplura: Hoplopleuridae), parasitizing Apodemus flavicollis in the Himalayan Region of India. Orient. Insects 1973, 7, 501–506. [Google Scholar] [CrossRef]
- Mitchell, R.M. Accounts of Nepalese mammals and analysis of the host-ectoparasite data by computer techniques. Ph.D. Thesis, Iowa State University, Ames, IA, USA, 1977. [Google Scholar]
- Kadulski, S.; Izdebska, J.N. Anoplura u gryzoni (Rodentia) z terenów Polski Północnej. Wiad. Parazytol. 2004, 50, 329–332. [Google Scholar] [PubMed]
- Stanko, M.; Fričová, J.; Miklisová, D.; Mošanský, L. Host-parasite relationships among parasitic arthropods and common vole (Microtus arvalis, Rodentia) in lowland ecosystems of Slovakia. In Arthropods: Epidemiological Importance; Buczek, A., Błaszak, C., Eds.; Koliber: Lublin, Poland, 2006; pp. 89–97. [Google Scholar]
- Stanko, M.; Fričová, J.; Miklisová, D.; Khokhlova, I.S.; Krasnov, B.R. Environment-related and host-related factors affecting the occurrence of lice on rodents in Central Europe. Parasitology 2015, 142, 938–947. [Google Scholar] [CrossRef] [Green Version]
- Wegner, Z. Klucze do oznaczania owadów Polski. Część XVI. In Wszy-Anoplura; PWN: Warsaw, Poland, 1972; p. 90. [Google Scholar]
- Johnson, P. Some Anoplura of the Oriental region. A study of Hoplopleura pacifica Ewing and allies. J. Med. Entomol. 1972, 9, 219–227. [Google Scholar] [CrossRef] [PubMed]
- Kozina, P.; Izdebska, J.N. Morphological anomalies in the body structure of Hoplopleura spp. (Anoplura: Hoplopleuridae). Ann. Parasitol. 2021, 67, 107–109. [Google Scholar]
- Haitlinger, R. Parasitological investigation of small mammals of Góry Sowie (Middle Sudetes). Pol. Pismo. Entomol. 1976, 46, 207–239. [Google Scholar]
- Grinbergs, A. Review of the animal lice fauna of Latvia. Latvijas Entomol. 1980, 23, 20–28. [Google Scholar]
- Haitlinger, R. Arthropods (Acari, Anoplura, Siphonaptera) of small mammals of Świętokrzyskie Provinence. Zesz. Nauk. Uniw. Przyr. Wroc. 2010, 577, 31–47. [Google Scholar]
- Dubinin, V.B. Parazitofauna mysevidnykh gryzunov i eyo izmeneiya v delt Volgi. Parazitologieeskij Zbornik 1953, 15, 252–301. [Google Scholar]
- Marinis, A.M.D.; Agnelli, P. Guide to the microscope analysis of Italian mammals hairs: Insectivora, Rodentia and Lagomorpha. Ital. J. Zool. 1993, 60, 225–232. [Google Scholar] [CrossRef]
- Kalueff, A.V.; Stewart, A.M.; Song, C.; Berridge, K.C.; Graybiel, A.M.; Fentress, J.C. Neurobiology of rodent self-grooming and its value for translational neuroscience. Nat. Rev. Neurosci. 2016, 17, 45–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, D.; Hao, J.; Yao, X.; Liu, Y.; Peng, T.; Jin, Z.; Meng, F. Observations of the foraging behavior and activity patterns of the Korean wood mouse, Apodemus peninsulae, in China, using infra-red cameras. Zookeys 2020, 992, 139–155. [Google Scholar] [CrossRef]
- Paggi, L.; Mattiucci, S.; Gibson, D.I.; Berland, B.; Nascetti, G.; Cianchi, R.; Cianchi, R.; Bullini, L. Pseudoterranova decipiens species A and B (Nematoda, Ascaridoidea): Nomenclatural designation, morphological diagnostic characters and genetic markers. Syst. Parasitol. 2000, 45, 185–197. [Google Scholar] [CrossRef]
- Shamsi, S.; Norman, R.; Gasser, R.; Beveridge, I. Redescription and genetic characterization of selected Contracaecum spp. (Nematoda: Anisakidae) from various hosts in Australia. Parasitol. Res. 2009, 104, 1507–1525. [Google Scholar] [CrossRef]
- Mattiucci, S.; Cipriani, P.; Levsen, A.; Paoletti, M.; Nascetti, G. Molecular epidemiology of Anisakis and anisakiasis: An ecological and evolutionary road map. Adv. Parasitol. 2018, 99, 93–263. [Google Scholar] [CrossRef]
- Lah, E.F.C.; Yaakop, S.; Ahamad, M.; George, E.; Nor, S.M. Precise identification of different stages of a tick, Ixodes granulatus Supino, 1897 (Acari: Ixodidae). Asian Pac. J. Trop. Biomed. 2016, 6, 597–604. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Physico-Geographical Region | Apodemus agrarius Collection Numbers | Location | Hoplopleura affinis Collection Numbers | Comments | ||
|---|---|---|---|---|---|---|
| City | Voivodeship | GPS Data | ||||
| CENTRAL EUROPEAN LOWLAND | ||||||
| Central Poland Lowlands | 2 specimens (UGDIZPMRMAa122K, UGDIZPMRMAa123K) | Niemce | Lublin | 51°21′ N 22°37′ E | 11 males (UGDIZPMAaAHHaf1m-UGDIZPMAgHaf11m) | 2 cases of co-occurrence of H. affinis-P. serrata |
| 36 females (UGDIZPMAaAHHaf12f-UGDIZPMAaAHHaf47f) | ||||||
| 1 first instar nymph (UGDIZPMAaAHHaf48N1), | ||||||
| 1 third instar nymph (UGDIZPMAaAHHaf49N3) | ||||||
| Southern Baltic Coastlands | 6 specimens (UGDIZPMRMAa156K, UGDIZPMRMAa157K, UGDIZPMRMAa161K-UGDIZPMRMAa163K, UGDIZPMRMAa177K) | Arciszewo | Pomeranian | 54°15′ N 18°33′ E | 1 female (UGDIZPMAaAHHaf50f) | |
| 4 specimens (UGDIZPMRMAa4K, UGDIZPMRMAa172K-UGDIZPMRMAa174K) | Gdynia | Pomeranian | 54°29′ N 18°25′ E | 2 males (UGDIZPMAaAHHaf51m, UGDIZPMAaAHHaf52m) | Co-occurrence of H. affinis-H. acanthopus | |
| 15 females (UGDIZPMAaAHHaf53f- UGDIZPMAaAHHaf67f) | ||||||
| 1 third instar nymph (UGDIZPMAaAHHaf68N3) | measurements—light microscopy; photos—scanning microscopy | |||||
| 1 specimen (UGDIZPMRMAa151K) | Gdańsk 1 | Pomeranian | 54°22′ N 18°36′ E | 3 females (UGDIZPMAaAHHaf69f-UGDIZPMAaAHHaf71f) | ||
| 1 specimen (UGDIZPMRMAa155K) | Gdańsk 2 | Pomeranian | 54°25′ N 18°36′ E | no specimens | ||
| 1 specimen (UGDIZPMRMAa178K) | Hopowo | Pomeranian | 54°15′ N 18°14′ E | 1 female (UGDIZPMAaAHHaf72f) | ||
| 1 specimen (UGDIZPMRMAa179K) | Łebieniec | Pomeranian | 54°43′ N 17°36′ E | 7 males (UGDIZPMAaAHHaf73m- UGDIZPMAaAHHaf79m) | ||
| 21 females (UGDIZPMAaAHHaf80f- UGDIZPMAaAHHaf100f) | ||||||
| 1 first instar nymph (UGDIZPMAaAHHaf101N1) | measurements | |||||
| 7 s instar nymphs (UGDIZPMAaAHHaf102N2- UGDIZPMAaAHHaf108N2) | measurements | |||||
| 2 specimens (UGDIZPMRMAa175K, UGDIZPMRMAa176K) | Swarożyn | Pomeranian | 54°02′ N 18°39′ E | no specimens | ||
| 48 specimens (UGDIZPMRMAa5K- UGDIZPMRMAa50K; UGDIZPMRMAa170K, UGDIZPMRMAa171K) | Tczew | Pomeranian | 54°06′ N 18°46′ E | 1 male (UGDIZPMAaAHHaf109m) | ||
| CZECH MASSIF | ||||||
| Sudety Mts and Sudety Foreland | 3 specimens (UGDIZPMRMAa1K- UGDIZPMRMAa3K) | Nowa Ruda | Lower Silesia | 50°31′ N 16°33′ E | no specimens | |
| POLISH UPLANDS | ||||||
| Lublin-Lviv Upland | 11 specimens (UGDIZPMRMAa66K- UGDIZPMRMAa73K, UGDIZPMRMAa76K, UGDIZPMRMAa127K, UGDIZPMRMAa128K) | Kijany | Lublin | 51°19′ N 22°46′ E | 2 males (UGDIZPMAaAHHaf110m, UGDIZPMAaAHHaf111m) | 2 cases of co-occurrence of H. affinis-P. serrata |
| 22 females (UGDIZPMAaAHHaf112f- UGDIZPMAaAHHaf133f) | ||||||
| 1 first instar nymph (UGDIZPMAaAHHaf134N1) | ||||||
| 26 specimens (UGDIZPMRMAa86K, UGDIZPMRMAa87K, UGDIZPMRMAa89K- UGDIZPMRMAa91K, UGDIZPMRMAa93K- UGDIZPMRMAa114K) | Łęczna | Lublin | 51°18′ N 22°52′ E | 5 males (UGDIZPMAaAHHaf135m- UGDIZPMAaAHHaf139m) | 2 cases of co-occurrence of H. affinis-P. serrata | |
| 23 females (UGDIZPMAaAHHaf140f- UGDIZPMAaAHHaf162f) | ||||||
| 4 first instar nymphs (UGDIZPMAaAHHaf163N1- UGDIZPMAaAHHaf166N1) | ||||||
| 1 s instar nymph (UGDIZPMAaAHHaf167N2) | measurements—light microscopy | |||||
| 2 third instar nymphs (UGDIZPMAaAHHaf168N3, UGDIZPMAaAHHaf169N3) | ||||||
| 10 specimens (UGDIZPMRMAa51K- UGDIZPMRMAa60K) | Nowogród | Lublin | 51°19′ N 22°47′ E | 3 males (UGDIZPMAaAHHaf170m-UGDIZPMAaAHHaf172m) | Co-occurrence of H. affinis-P. serrata | |
| 3 females (UGDIZPMAaAHHaf173f- UGDIZPMAaAHHaf175f) | ||||||
| WESTERN CARPATHIANS and WESTERN AND NORTHERN SUBCARPATHIANS | ||||||
| Outer Western Carpathians | 19 specimens (UGDIZPMRMAa61K- UGDIZPMRMAa65K, UGDIZPMRMAa74K, UGDIZPMRMAa75K, UGDIZPMRMAa77K- UGDIZPMRMAa85K, UGDIZPMRMAa88K, UGDIZPMRMAa92K, UGDIZPMRMAa121K) | Rymanów | Subcarpathian | 49°35′ N 21°50′ E | 10 males (UGDIZPMAaAHHaf176m- UGDIZPMAaAHHaf185m) | 2 cases of co-occurrence of H. affinis-P. serrata |
| 31 females (UGDIZPMAaAHHaf186f- UGDIZPMAaAHHaf216f) | ||||||
| 1 first instar nymph (UGDIZPMAaAHHaf217N1) | ||||||
| 1 s instar nymph (UGDIZPMAaAHHaf218N2) | measurements—light microscopy; photos—scanning microscopy | |||||
| EASTERN BALTIC-BELARUS LOWLAND | ||||||
| 14 specimens (UGDIZPMRMAa115K- UGDIZPMRMAa120K, UGDIZPMRMAa124K, UGDIZPMRMAa125K, UGDIZPMRMAa136K, UGDIZPMRMAa137K, UGDIZPMRMAa140K, UGDIZPMRMAa141K, UGDIZPMRMAa149K, UGDIZPMRMAa150K) | Chełm | Lublin | 51°07′ N 23°28′ E | 3 males (UGDIZPMAaAHHaf219m- UGDIZPMAaAHHaf221m) | 3 cases of co-occurrence of H. affinis-P. serrata | |
| 6 females (UGDIZPMAaAHHaf222f- UGDIZPMAaAHHaf227f) | ||||||
| 17 specimens (UGDIZPMRMAa126K, UGDIZPMRMAa129K- UGDIZPMRMAa135K, UGDIZPMRMAa138K, UGDIZPMRMAa139K, UGDIZPMRMAa142K- UGDIZPMRMAa148K) | Strupin Duży | Lublin | 51°05′ N 23°30′ E | 5 males (UGDIZPMAaAHHaf228m-UGDIZPMAaAHHaf232m) | 4 cases of co-occurrence of H. affinis-P. serrata | |
| 12 females (UGDIZPMAaAHHaf233f- UGDIZPMAaAHHaf244f) | ||||||
| Eastern Baltic Coastland | 3 specimens (UGDIZPMRMAa164K, UGDIZPMRMAa168K, UGDIZPMRMAa169K) | Stare Jabłonki | Warmian-Masurian | 53°42′ N 20°04′ E | no specimens | |
| 4 specimens (UGDIZPMRMAa152K, UGDIZPMRMAa153K, UGDIZPMRMAa165K, UGDIZPMRMAa166K) | Kleszczewo | Greater Poland | 54°11′ N 18°30′ E | no specimens | ||
| 4 specimens (UGDIZPMRMAa154K, UGDIZPMRMAa158K-UGDIZPMRMAa160K) | Słomowo | Greater Poland | 52°21′ N 17°32′ E | 1 male (UGDIZPMAaAHHaf245m) | ||
| no data | UGDIZPMAaAHHaf246f | |||||
| UGDIZPMAaAHHaf247f | ||||||
| Host | Host Distribution | Hoplopleura Species | Parasite Distribution |
|---|---|---|---|
| Apodemus agrarius (Pallas, 1771) | Eastern Europe to Eastern Asia | H. acanthopus [6,7,25] | Holarctic region 1 |
| H. affinis [2,4,6,7,25] | Eurasia | ||
| Apodemus alpicola Heinrich, 1952 | endemic to North West parts of the Alps | no data | no data |
| Apodemus argenteus Temminck, 1844 | endemic to Japan | H. akanezumi [6] | Japan, Taiwan |
| H. himenezumi [2,4,6,20] | Japan | ||
| H. inagakii [6] | Japan | ||
| Apodemus chevrieri Milne-Edwards, 1868 | West Central China | H. affinis [2,4] | Eurasia |
| Apodemus draco Barrett-Hamilton, 1900 = Apodemus ilex Thomas, 1922 | China and North East India | no data | no data |
| Apodemus epimelas (Nehring, 1902) | South Eastern Europe | no data | no data |
| Apodemus flavicolis (Melchior, 1834) | Europe and western Asia | H. acanthopus [6,7,22,23,25] | Holarctic region 1 |
| H. affinis [2,4,7] | Eurasia | ||
| H. edentula [7] | Eurasia | ||
| H. himalayana [4,6,21] | India | ||
| Apodemus gurkha Thomas, 1924 | endemic to Nepal | H. pacifica [5] | cosmopolitan (in tropical, subtropical and southern temperature zones) 2 |
| Apodemus hyrcanicus Vorontsov, Boyeskorov, and Mezhzherin, 1992 | South Caucasus to Central Asia | no data | no data |
| Apodemus latronum Thomas, 1911 | China, India and Burma | no data | no data |
| Apodemus mystacinus Danford and Alston, 1877 | Albania, Bosnia and Herzegovina, Croatia, Georgia, Greece, Iran, Iraq, Israel, Jordan, Lebanon, Saudi Arabia, Serbia and Montenegro | no data | no data |
| Apodemus pallipes Barrett-Hamilton, 1900 = Apodemus wardi (Wroughton, 1908) | Kyrgyzstan, Tajikistan, Afghanistan, India, Iran, Nepal and Pakistan | H. affinis [5] | Eurasia |
| H. captiosa [5] | cosmopolitan (probably) 3 | ||
| H. himalayana [2,4] | India | ||
| H. pacifica [5] | cosmopolitan (in tropical, subtropical and southern temperature zones) 2 | ||
| Apodemus peninsulae Thomas, 1906 | Northeastern Asia, including the Russian Far East, northern China, the Korean Peninsula, Sakhalin and Hokkaidō | no data | no data |
| Apodemus ponticus Sviridenko, 1936 | endemic to the Caucasus | no data | no data |
| Apodemus rusiges Miller, 1913 | India, Nepal and Pakistan | H. himalayana [2,4] | India |
| Apodemus semotus Thomas, 1908 | endemic to Taiwan | H. akanezumi [2,4] | Japan 4, Taiwan |
| Apodemus speciosus Temminck, 1844 | endemic to Japan | H. acanthopus [6] | Holarctic region 1 |
| H. affinis [2,4] | Eurasia | ||
| H. akanezumi [2,4,6] | Japan, Taiwan | ||
| H. himenezumi [6] | Japan | ||
| H. inagakii [6] | Japan | ||
| Apodemus sylvaticus Linnaeus, 1758 | Europe and North Western Africa | H. acanthopus [6,22,23,25] | Holarctic region 1 |
| H. affinis [2,4,6,24,25] | Eurasia | ||
| H. captiosa [25] | cosmopolitan (probably) 3 | ||
| Apodemus uralensis (Pallas, 1811) = A. microps Kratochvíl and Rosicky, 1952 | Central Europe and Asia | H. acanthopus [6,7] | Holarctic region 1 |
| H. affinis [7] | Eurasia | ||
| Apodemus witherbyi Thomas, 1902 | Eastern Europe, Near East and Central Asia | H. affinis [8] | Eurasia |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kozina, P.; Izdebska, J.N.; Łopucki, R. Description of the Nymphal Stages of Hoplopleura affinis (Anoplura: Hoplopleuridae) from the Striped Field Mouse Apodemus agrarius with a Global Checklist of Hoplopleura Species from the Genus Apodemus. Insects 2022, 13, 107. https://doi.org/10.3390/insects13020107
Kozina P, Izdebska JN, Łopucki R. Description of the Nymphal Stages of Hoplopleura affinis (Anoplura: Hoplopleuridae) from the Striped Field Mouse Apodemus agrarius with a Global Checklist of Hoplopleura Species from the Genus Apodemus. Insects. 2022; 13(2):107. https://doi.org/10.3390/insects13020107
Chicago/Turabian StyleKozina, Paulina, Joanna N. Izdebska, and Rafał Łopucki. 2022. "Description of the Nymphal Stages of Hoplopleura affinis (Anoplura: Hoplopleuridae) from the Striped Field Mouse Apodemus agrarius with a Global Checklist of Hoplopleura Species from the Genus Apodemus" Insects 13, no. 2: 107. https://doi.org/10.3390/insects13020107