
Attraction of Egg Parasitoids Trissolcus mitsukurii and Trissolcus japonicus to the chemical cues of Halyomorpha halys and Nezara viridula

 , , , , , ,
, , , , , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Parasitoids
2.2. Hosts
2.3. Foraging Behaviour on Footprints
2.4. Foraging Behaviour on Synthetic Compounds
2.5. Statistical Analysis
3. Results
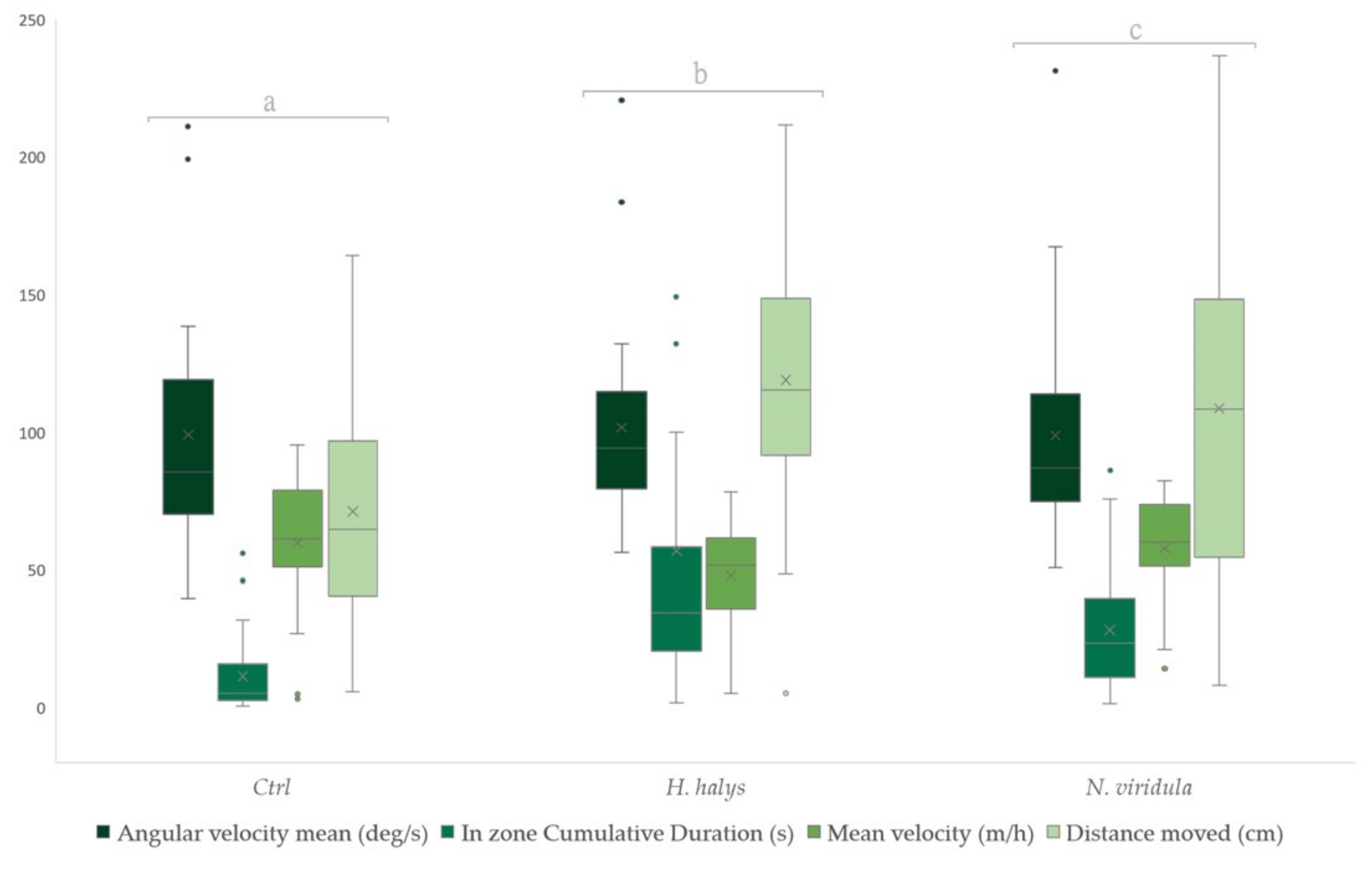
3.1. Foraging Behaviour on Footprints
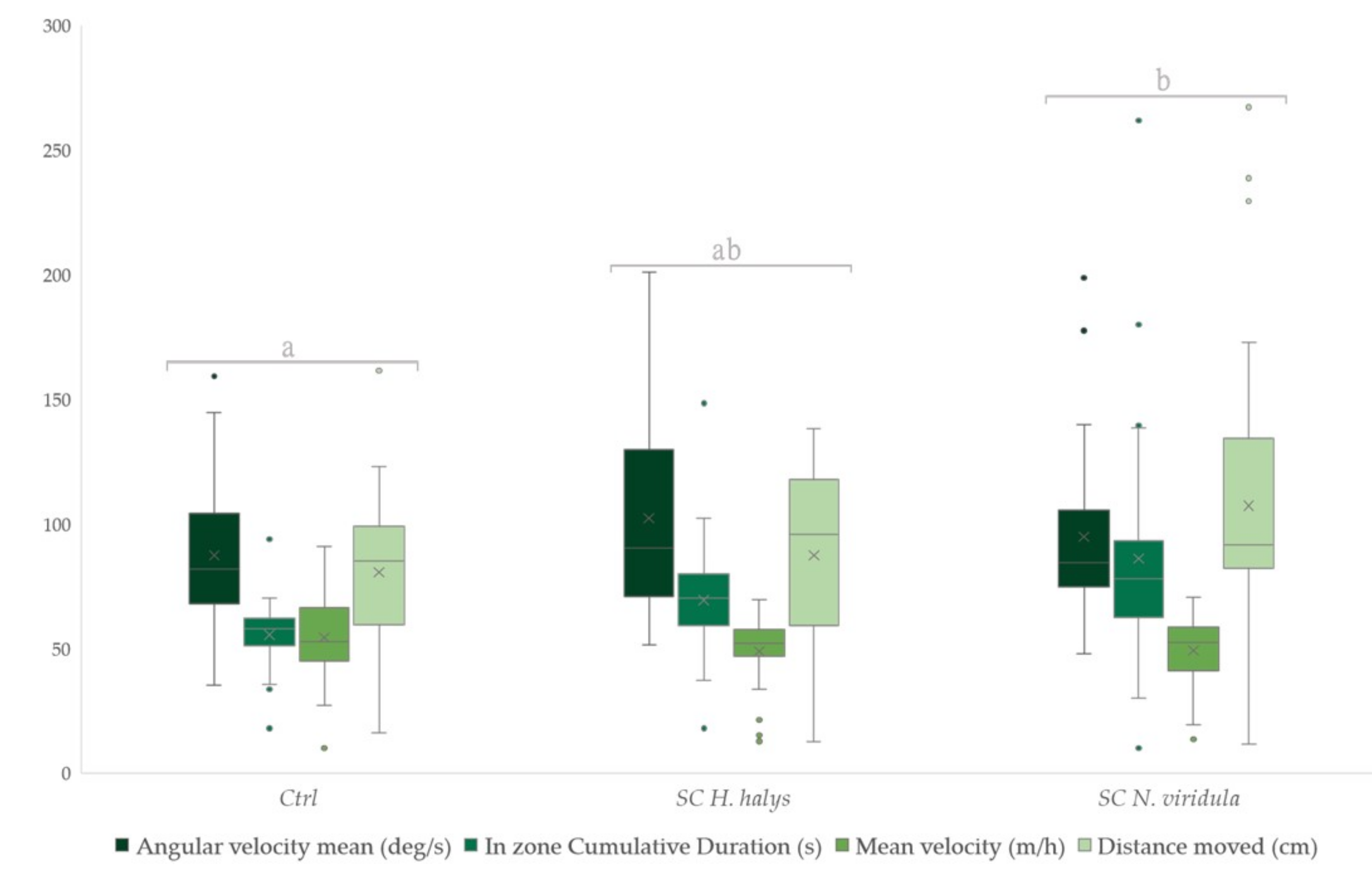
3.2. Foraging Behaviour on Synthetic Compounds
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Hokyo, N.; Kiritani, K. Two species of egg parasites as contemporaneous mortality factors in the egg population of the southern green stink bug, Nezara viridula. Jpn. J. Appl. Entomol. 1963, 7, 214–227. [Google Scholar] [CrossRef]
- Kobayashi, T.; Cosenza, G.W. Integrated control of soybean stink bugs in the Cerrados. Jpn. Agric. Res. Q. 1987, 20, 229–236. [Google Scholar]
- Clarke, A.R. The control of Nezara viridula L. with introduced egg parasitoids in australia. A review of a ‘landmark’ example of classical biological control. Aust. J. Agric. Res. 1990, 41, 1127–1146. [Google Scholar] [CrossRef]
- Ryu, J.; Hirashima, Y. Taxonomic studies on the genus Trissolcus ashmead of Japan and Korea (Hymenoptera, Scelionidae). J. Fac. Agric. Kyushu Univ. 1984, 29, 35–58. [Google Scholar] [CrossRef]
- Cassis, G.; Gross, C.F. HEMIPTERA: Heteroptera (Pentatomomorpha). In Zoological Catalogue of Australia; ABRS/CSIRO Publishing: East Geelong, VIC, Austalia, 2002; Volume 27.3B, ISBN 978-0-643-06875-9. [Google Scholar]
- Johnson, N.F. Revision of Australasian Trissolcus species (Hymenoptera: Scelionidae). Invertebr. Taxon. 1991, 5, 211–239. [Google Scholar] [CrossRef]
- Yang, Z.-Q.; Yao, Y.-X.; Qiu, L.-F.; Li, Z.-X. A new species of Trissolcus (Hymenoptera: Scelionidae) parasitizing eggs of Halyomorpha halys (Heteroptera: Pentatomidae) in China with comments on its biology. Ann. Entomol. Soc. Am. 2009, 102, 39–47. [Google Scholar] [CrossRef]
- Yonow, T.; Kriticos, D.J.; Ota, N.; Avila, G.A.; Hoelmer, K.A.; Chen, H.; Caron, V. Modelling the potential geographic distribution of two Trissolcus Species for the brown marmorated stink bug. Halyomorpha Halys. Insects 2021, 12, 491. [Google Scholar] [CrossRef]
- Maistrello, L.; Vaccari, G.; Caruso, S.; Costi, E.; Bortolini, S.; Macavei, L.; Foca, G.; Ulrici, A.; Bortolotti, P.P.; Nannini, R.; et al. Monitoring of the invasive Halyomorpha halys, a new key pest of fruit orchards in Northern Italy. J. Pest Sci. 2017, 90, 1231–1244. [Google Scholar] [CrossRef]
- Malek, R.; Tattoni, C.; Ciolli, M.; Corradini, S.; Andreis, D.; Ibrahim, A.; Mazzoni, V.; Eriksson, A.; Anfora, G. Coupling traditional monitoring and citizen science to disentangle the invasion of Halyomorpha halys. ISPRS Int. J. Geo-Inf. 2018, 7, 171. [Google Scholar] [CrossRef] [Green Version]
- Leskey, T.C.; Nielsen, A.L. Impact of the invasive brown marmorated stink bug in North America and Europe: History, biology, ecology, and management. Annu. Rev. Entomol. 2018, 63, 599–618. [Google Scholar] [CrossRef] [Green Version]
- Rice, K.B.; Bergh, C.J.; Bergmann, E.J.; Biddinger, D.; Dieckhoff, C.; Dively, G.; Fraser, H.; Gariepy, T.; Hamilton, G.; Haye, T.; et al. Biology, ecology, and management of brown marmorated stink bug (Hemiptera: Pentatomidae). J. Integr. Pest Manag. 2014, 5, A1–A13. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, F.; Gariepy, T.; Mason, P.; Gillespie, D.; Talamas, E.; Haye, T. Seasonal parasitism and host specificity of Trissolcus japonicus in Northern China. J. Pest Sci. 2017, 90, 1127–1141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scaccini, D.; Falagiarda, M.; Tortorici, F.; Martinez-Sañudo, I.; Tirello, P.; Reyes-Domínguez, Y.; Gallmetzer, A.; Tavella, L.; Zandigiacomo, P.; Duso, C.; et al. An insight into the role of Trissolcus mitsukurii as biological control agent of Halyomorpha halys in Northeastern Italy. Insects 2020, 11, 306. [Google Scholar] [CrossRef] [PubMed]
- Zapponi, L.; Bon, M.C.; Fouani, J.M.; Anfora, G.; Schmidt, S.; Falagiarda, M. Assemblage of the egg parasitoids of the invasive stink bug Halyomorpha halys: Insights on plant host associations. Insects 2020, 11, 588. [Google Scholar] [CrossRef]
- Zapponi, L.; Tortorici, F.; Anfora, G.; Bardella, S.; Bariselli, M.; Benvenuto, L.; Bernardinelli, I.; Butturini, A.; Caruso, S.; Colla, R.; et al. Assessing the distribution of exotic egg parasitoids of Halyomorpha halys in europe with a large-scale monitoring program. Insects 2021, 12, 316. [Google Scholar] [CrossRef]
- Biondi, A.; Wang, X.; Daane, K.M. Host preference of three asian larval parasitoids to closely related drosophila species: Implications for biological control of Drosophila suzukii. J. Pest. Sci. 2021, 94, 273–283. [Google Scholar] [CrossRef]
- Godfray, H.C.J. Parasitoids: Behavioral and Evolutionary Ecology; Princeton University Press Books: Princeton, NJ, USA, 1994; ISBN 978-0-691-00047-3. [Google Scholar]
- Vet, L.; Dicke, M. Ecology of infochemical use by natural enemies in a tritrophic context. Annu. Rev. Entomol. 1992, 37, 141–172. [Google Scholar] [CrossRef]
- Nieri, R.; Anfora, G.; Mazzoni, V.; Rossi Stacconi, M.V. Semiochemicals, semiophysicals and their integration for the development of innovative multi-modal systems for agricultural pests’ monitoring and control. Entomol. Gen. 2022, 42, 167–183. [Google Scholar] [CrossRef]
- Meiners, T.; Peri, E. Chemical ecology of insect parasitoids: Essential elements for developing effective biological control programmes. In Chemical Ecology of Insect Parasitoids; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2013; pp. 191–224. ISBN 978-1-118-40958-9. [Google Scholar]
- Wäschke, N.; Meiners, T.; Rostás, M. Foraging strategies of parasitoids in complex chemical environments. In Chemical Ecology of Insect Parasitoids; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2013; pp. 37–63. ISBN 978-1-118-40958-9. [Google Scholar]
- Colazza, S.; Aquila, G.; De Pasquale, C.; Peri, E.; Millar, J.G. The egg parasitoid Trissolcus basalis Uses N-nonadecane, a cuticular hydrocarbon from its stink bug host Nezara viridula, to discriminate between female and male hosts. J. Chem. Ecol. 2007, 33, 1405–1420. [Google Scholar] [CrossRef] [Green Version]
- Federle, W.; Riehle, M.; Curtis, A.S.; Full, R.J. An integrative study of insect adhesion: Mechanics and wet adhesion of pretarsal pads in ants. In Integrative and Comparative Biology; Oxford University Press: Oxford, UK, 2002; Volume 42, pp. 1100–1106. [Google Scholar] [CrossRef]
- Müller, C.; Riederer, M. Plant surface properties in chemical ecology. J. Chem. Ecol. 2005, 31, 2621–2651. [Google Scholar] [CrossRef]
- Lo Giudice, D.; Riedel, M.; Rostás, M.; Peri, E.; Colazza, S. Host sex discrimination by an egg parasitoid on brassica leaves. J. Chem. Ecol. 2011, 37, 622–628. [Google Scholar] [CrossRef] [PubMed]
- Conti, E.; Salerno, G.; Bin, F.; Williams, H.J.; Vinson, S.B. Chemical cues from Murgantia histrionica eliciting host location and recognition in the egg parasitoid Trissolcus brochymenae. J. Chem. Ecol. 2003, 29, 115–130. [Google Scholar] [CrossRef] [PubMed]
- Colazza, S.; Salerno, G.; Wajnberg, E. Volatile and contact chemicals released by Nezara viridula (Heteroptera:Pentatomidae) have a Kairomonal effect on the egg parasitoid Trissolcus basalis (Hymenoptera: Scelionidae). Biol. Control 1999, 16, 310–317. [Google Scholar] [CrossRef] [Green Version]
- Salerno, G.; Conti, E.; Peri, E.; Colazza, S.; Bin, F. Kairomone involvement in the host specificity of the egg parasitoid Trissolcus basalis (Hymenoptera: Scelionidae). Eur. J. Entomol. 2006, 103, 311–318. [Google Scholar] [CrossRef] [Green Version]
- Colazza, S.; Peri, E.; Salerno, G.; Conti, E. Host searching by egg parasitoids: Exploitation of host chemical cues. In Egg Parasitoids in Agroecosystems with Emphasis on Trichogramma; Consoli, F.L., Parra, J.R.P., Zucchi, R.A., Eds.; Progress in Biological Control; Springer: Dordrecht, The Netherlands, 2009; pp. 97–147. ISBN 978-1-4020-9110-0. [Google Scholar]
- Zhong, Y.Z.; Zhang, J.P.; Ren, L.L.; Tang, R.; Zhan, H.X.; Chen, G.H.; Zhang, F. Behavioral responses of the egg parasitoid Trissolcus japonicus to volatiles from adults of its stink bug host, Halyomorpha halys. J. Pest Sci. 2017, 90, 1097–1105. [Google Scholar] [CrossRef]
- Conti, E.; Salerno, G.; Bin, F.; Bradleigh Vinson, S. The role of host semiochemicals in parasitoid specificity: A case study with Trissolcus brochymenae and Trissolcus simoni on Pentatomid Bugs. Biol. Control 2004, 29, 435–444. [Google Scholar] [CrossRef]
- Malek, R.; Kaser, J.M.; Anfora, G.; Ciolli, M.; Khrimian, A.; Weber, D.C.; Hoelmer, K.A. Trissolcus japonicus foraging behavior: Implications for host preference and classical biological control. Biol. Control 2021, 161, 104700. [Google Scholar] [CrossRef]
- Boyle, S.M.; Weber, D.C.; Hough-Goldstein, J.; Hoelmer, K.A. Host kairomones influence searching behavior of Trissolcus japonicus (Hymenoptera: Scelionidae), a parasitoid of Halyomorpha halys (Heteroptera: Pentatomidae). Environ. Entomol. 2020, 49, 15–20. [Google Scholar] [CrossRef] [Green Version]
- Suckling, D.M.; Levy, M.C.; Roselli, G.; Mazzoni, V.; Ioriatti, C.; Deromedi, M.; Cristofaro, M.; Anfora, G. Live traps for adult brown marmorated stink bugs. Insects 2019, 10, 376. [Google Scholar] [CrossRef] [Green Version]
- Aldrich, J.R.; Blum, M.S.; Lloyd, H.A.; Fales, H.M. Pentatomid natural products. J. Chem. Ecol. 1978, 4, 161–172. [Google Scholar] [CrossRef]
- Ellis, A.R.; Burchett, W.; Harrar, S.W.; Bathke, A.C. Nonparametric Inference for multivariate data: The package Npmv. J. Stat. Softw. 2017, 76, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Mayorga-Ch, D.; Castro-Cortés, N.C.; Rodríguez, C.; Sarmiento, C.E. Behavioral responses of the social Wasp Polistes myersi to prey infected with fungi used in biological control. J. Insect Behav. 2021, 34, 136–149. [Google Scholar] [CrossRef]
- Peri, E.; Sole, M.A.; Wajnberg, E.; Colazza, S. Effect of host kairomones and oviposition experience on the arrestment behavior of an egg parasitoid. J. Exp. Biol. 2006, 209, 3629–3635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mustaparta, H. Chemical information processing in the olfactory system of insects. Physiol. Rev. 1990, 70, 199–245. [Google Scholar] [CrossRef]
- Matthews, R.W.; Matthews, J.R. Parental behaviors and social life. In Insect Behavior, 2nd ed.; Springer: New York, NY, USA, 2009; pp. 389–443. ISBN 978-90-481-2389-6. [Google Scholar]
- Rondoni, G.; Chierici, E.; Giovannini, L.; Sabbatini-Peverieri, G.; Roversi, P.F.; Conti, E. Olfactory responses of Trissolcus mitsukurii to plants attacked by target and non-target stink bugs suggest low risk for biological control. Sci. Rep. 2022, 12, 1–9. [Google Scholar] [CrossRef]
- Giovannini, L.; Sabbatini-Peverieri, G.; Marianelli, L.; Rondoni, G.; Conti, E.; Roversi, P.F. Physiological host range of Trissolcus mitsukurii, a candidate biological control agent of halyomorpha halys in Europe. J. Pest Sci. 2021, 95, 605–618. [Google Scholar] [CrossRef]
- Luck, R.F.; Uygun, N. Host recognition and selection by aphytis species: Response to california red, oleander, and cactus scale cover extracts. Entomol. Exp. Appl. 1986, 40, 129–136. [Google Scholar] [CrossRef]
- Boyle, S.M.; Weber, D.C.; Hough-Goldstein, J.; Hoelmer, K.A. Parental host species affects behavior and parasitism by the pentatomid egg parasitoid, Trissolcus japonicus (Hymenoptera: Scelionidae). Biol. Control 2020, 149, 104324. [Google Scholar] [CrossRef]
- Sheehan, W.; Shelton, A.M. The role of experience in plant foraging by the aphid parasitoid Diaeretiella rapae (Hymenoptera: Aphidiidae). J. Insect Behav. 1989, 2, 743–759. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
| Parameters | ||||||
|---|---|---|---|---|---|---|
| Angular Velocity Mean | In Zone Cumulative Duration | Mean Velocity | Distance Moved | |||
| Footprints | T. japonicus | Control | 0.46852 | 0.30000 | 0.56778 | 0.51556 |
| H. halys | 0.53815 | 0.66815 | 0.39000 | 0.73704 | ||
| N. viridula | 0.49333 | 0.53185 | 0.54222 | 0.24741 | ||
| T. mitsukurii | Control | 0.34630 | 0.24352 | 0.64074 | 0.27111 | |
| H. halys | 0.45815 | 0.57389 | 0.47333 | 0.56407 | ||
| N. viridula | 0.69556 | 0.68259 | 0.38593 | 0.66481 | ||
| Synthetic compounds | T. mitsukurii | Control | 0.45185 | 0.31833 | 0.54593 | 0.42704 |
| SC H. halys | 0.55111 | 0.54481 | 0.47296 | 0.51074 | ||
| SC N. viridula | 0.49704 | 0.63685 | 0.48111 | 0.56222 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Scala, M.; Fouani, J.M.; Zapponi, L.; Mazzoni, V.; Wells, K.E.; Biondi, A.; Baser, N.; Verrastro, V.; Anfora, G. Attraction of Egg Parasitoids Trissolcus mitsukurii and Trissolcus japonicus to the chemical cues of Halyomorpha halys and Nezara viridula. Insects 2022, 13, 439. https://doi.org/10.3390/insects13050439
Scala M, Fouani JM, Zapponi L, Mazzoni V, Wells KE, Biondi A, Baser N, Verrastro V, Anfora G. Attraction of Egg Parasitoids Trissolcus mitsukurii and Trissolcus japonicus to the chemical cues of Halyomorpha halys and Nezara viridula. Insects. 2022; 13(5):439. https://doi.org/10.3390/insects13050439
Chicago/Turabian StyleScala, Marica, Jalal Melhem Fouani, Livia Zapponi, Valerio Mazzoni, Karen Elizabeth Wells, Antonio Biondi, Nuray Baser, Vincenzo Verrastro, and Gianfranco Anfora. 2022. "Attraction of Egg Parasitoids Trissolcus mitsukurii and Trissolcus japonicus to the chemical cues of Halyomorpha halys and Nezara viridula" Insects 13, no. 5: 439. https://doi.org/10.3390/insects13050439