
Identification and Characterization of Antioxidant Enzyme Genes in Parasitoid Aphelinus asychis (Hymenoptera: Aphelinidae) and Expression Profiling Analysis under Temperature Stress



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insect Species
2.2. Temperature Treatments
2.3. Identification of the Antioxidant Enzyme Genes in A. asychis
2.4. Phylogenetic and Structural Feature Analysis of Antioxidant Enzymes Genes
2.5. RNA Extraction and Expression Profiles of Antioxidant Enzyme Genes
2.6. Enzyme Activity Assay
2.7. Statistical Analyses
3. Results
3.1. Effect of Short-Term High Temperate on the Survival Rate of A. asychis
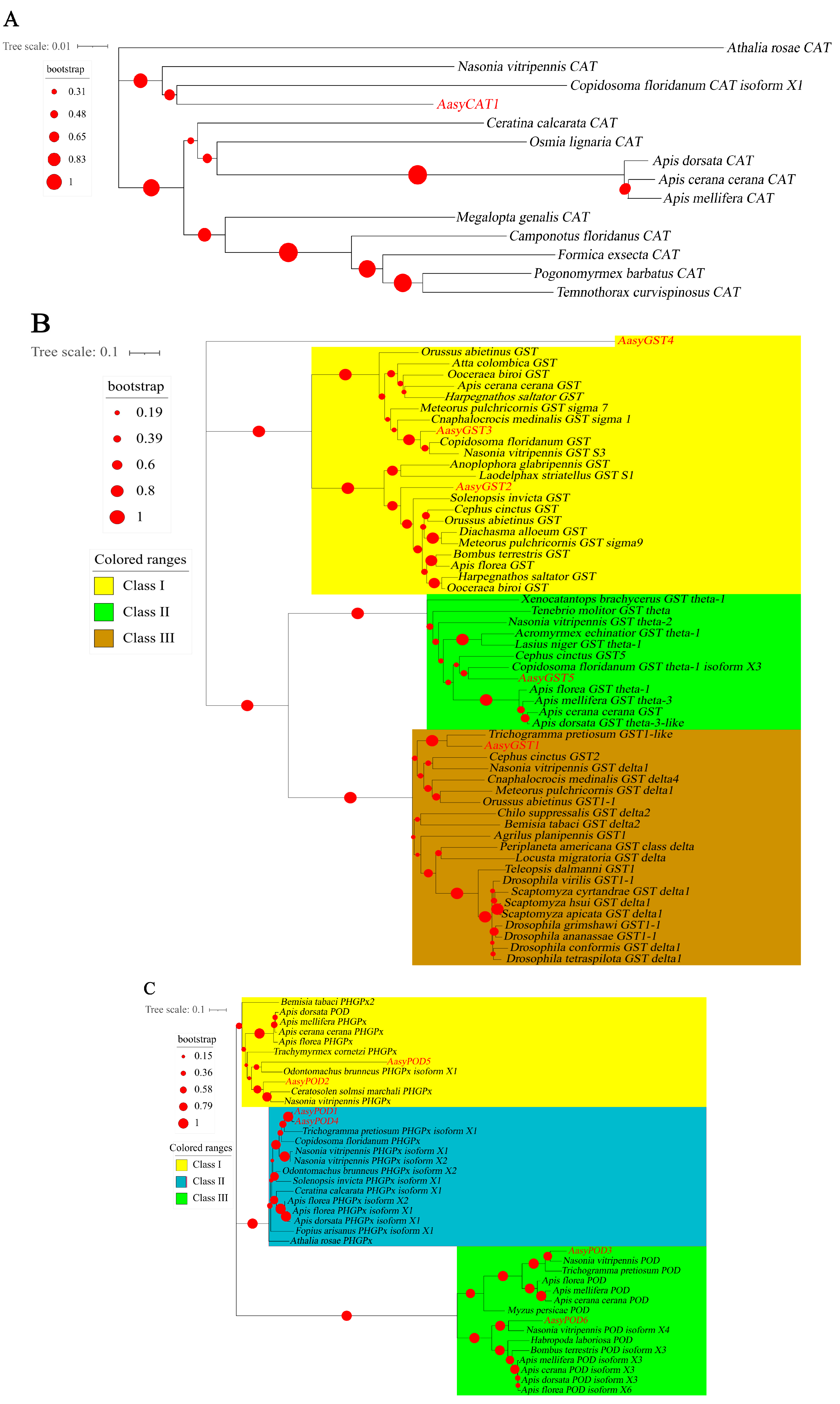
3.2. The Identification and Phylogenetic Relationship Analysis of Antioxidant Enzyme Genes in A. asychis
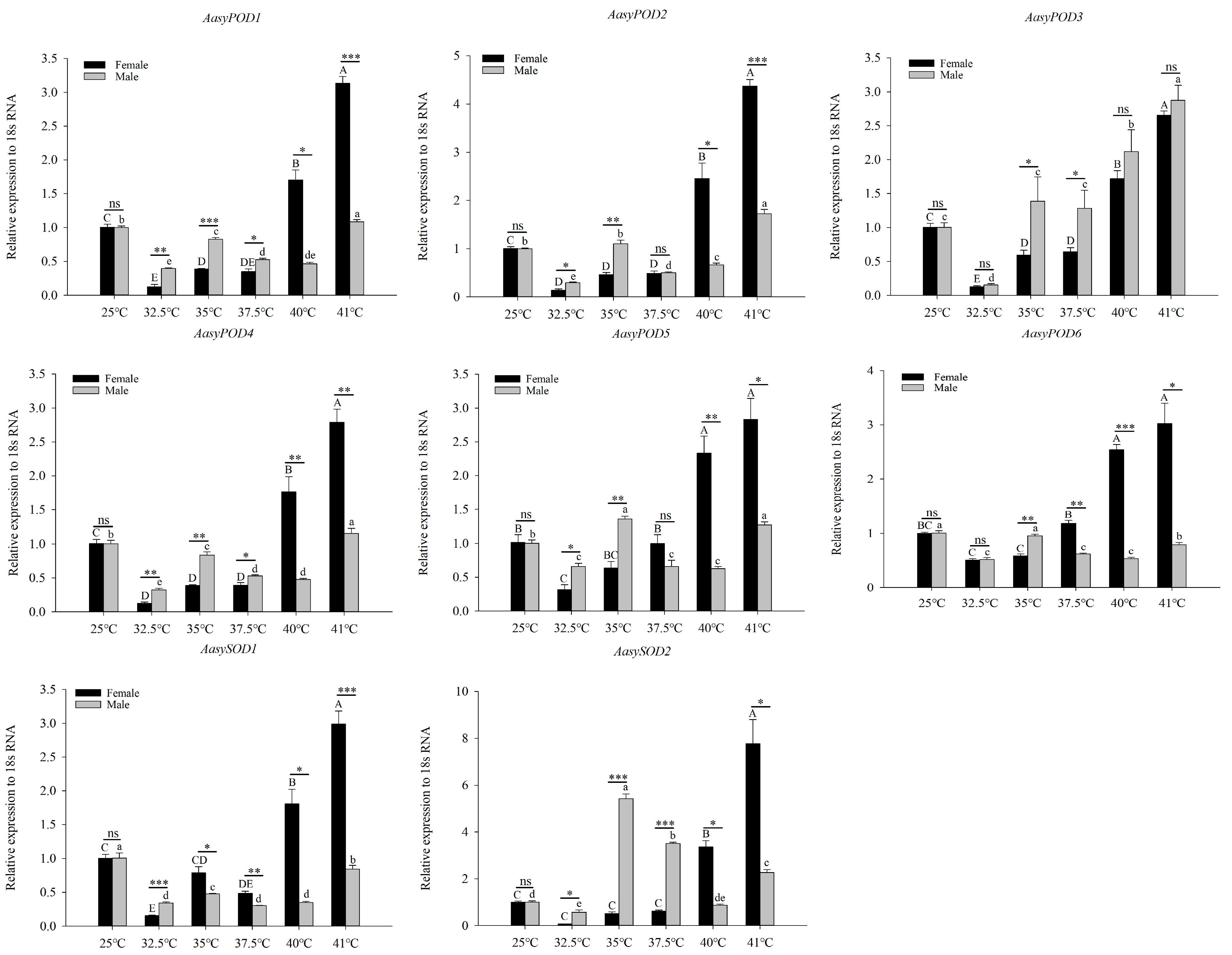
3.3. Differential Expression of Antioxidant Enzyme Genes in A. asychis under High-Temperature Stress
3.4. Antioxidant Enzyme Activities of A. asychis in Response to High-Temperature Stress
4. Discussions
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Livingstone, D.R. Contaminant-stimulated reactive oxygen species production and oxidative damage in aquatic organisms. Mar. Pollut. Bull. 2001, 42, 656–666. [Google Scholar] [CrossRef]
- Hernández-García, D.; Wood, C.D.; Castro-Obregón, S.; Covarrubias, L. Reactive oxygen species: A radical role in development? Free Radic. Biol. Med. 2010, 49, 130–143. [Google Scholar] [CrossRef]
- Aucoin, R.R.; Philogène, B.J.R.; Arnason, J.T. Antioxidant enzymes as biochemical defenses against phototoxin-induced oxidative stress in three species of herbivorous Lepidoptera. Arch. Insect Biochem. Physiol. 1991, 16, 139–152. [Google Scholar] [CrossRef]
- Felton, G.W.; Summers, C.B. Antioxidant system in insects. Arch. Insect Biochem. Physiol. 1995, 29, 187–197. [Google Scholar] [CrossRef]
- Yang, L.-H.; Huang, H.; Wang, J.-J. Antioxidant responses of citrus red mite, Panonychus citri (McGregor) (Acari: Tetranychidae), exposed to thermal stress. J. Insect Physiol. 2010, 56, 1871–1876. [Google Scholar] [CrossRef]
- Ahmad, S. Biochemical defence of pro-oxidant plant allelochemicals by herbivorous insects. Biochem. Syst. Ecol. 1992, 20, 269–296. [Google Scholar] [CrossRef]
- Ahmad, S.; Duval, D.L.; Weinhold, L.C.; Pardini, R.S. Cabbage looper antioxidant enzymes: Tissue specificity. Insect Biochem. 1991, 21, 563–572. [Google Scholar] [CrossRef]
- Felton, G.W.; Duffey, S.S. Protective action of midgut catalase in lepidopteran larvae against oxidative plant defenses. J. Chem. Ecol. 1991, 17, 1715–1732. [Google Scholar] [CrossRef]
- Pickett, J.; Wadhams, L.J.; Woodcock, C.M.; Hardie, J. The chemical ecology of aphids. Annu. Rev. Entomol. 1992, 37, 67–90. [Google Scholar] [CrossRef]
- Dubovskiy, I.M.; Martemyanov, V.V.; Vorontsova, Y.L.; Rantala, M.J.; Gryzanova, E.V.; Glupov, V.V. Effect of bacterial infection on antioxidant activity and lipid peroxidation in the midgut of Galleria mellonella larvae (Lepidoptera: Pyralidae). Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2008, 148, 1–5. [Google Scholar] [CrossRef]
- Campbell, A.; Frazer, B.D.; Gilbert, N.; Gutierrez, A.P.; Mackauer, M. Temperature requirements of some aphids and their parasites. J. Appl. Ecol. 1974, 11, 431–438. [Google Scholar] [CrossRef]
- Logan, J.A.; Wollkind, D.J.; Hoyt, S.C.; Tanigoshi, L.K. An analytic model for description of temperature dependent rate phenomena in arthropods. Environ. Entomol. 1976, 5, 1133–1140. [Google Scholar] [CrossRef]
- Joanisse, D.R.; Stotey, K. Oxidative stress and antioxidants in overwintering larvae of cold-hardy goldenrod gall insects. J. Exp. Biol. 1996, 199, 1483–1491. [Google Scholar] [CrossRef]
- Shirley, X.A.; Woolley, J.B.; Hopper, K.R. Revision of the asychis species group of Aphelinus (Hymenoptera: Aphelinidae). J. Hymenopt. Res. 2017, 54, 1–32. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.-C.; Fang, Q.-Q. The phylogenetic relationships of introduced Aphelinus (Hymenoptera: Aphelinidae), biological control agents of the Russian wheat aphid (Homoptera: Aphididae). Insect Sci. 2009, 16, 277–285. [Google Scholar] [CrossRef]
- Dong, Z.-K.; Hou, R.-Z.; Ouyang, Z.; Zhang, R.-Z. Tritrophic interaction influenced by warming and tillage: A field study on winter wheat, aphids and parasitoids. Agric. Ecosyst. Environ. 2013, 181, 144–148. [Google Scholar] [CrossRef]
- Sentis, A.; Morisson, J.; Boukal, D.S. Thermal acclimation modulates the impacts of temperature and enrichment on trophic interaction strengths and population dynamics. Glob. Chang. Biol. 2015, 21, 3290–3298. [Google Scholar] [CrossRef]
- Wang, S.-Y.; Wang, B.-L.; Yan, G.-L.; Liu, Y.-H.; Zhang, D.-Y.; Liu, T.-X. Temperature-dependent demographic characteristics and control potential of Aphelinus asychis reared from Sitobion avenae as a biological control agent for Myzus persicae on Chili Peppers. Insects 2020, 11, 475. [Google Scholar] [CrossRef]
- Wang, S.-Y.; Zhang, D.-Y.; Liu, T.-X. Influence of temperature on the demographics and control efficiency of Aphelinus asychis, a parasitoid of the cabbage pest, Myzus persicae. Phytoparasitica 2020, 48, 767–783. [Google Scholar] [CrossRef]
- Kang, Z.-W.; Liu, F.-H.; Liu, X.; Yu, W.-B.; Tan, X.-L.; Zhang, S.-Z.; Tian, H.-G.; Liu, T.-X. The potential coordination of the heat-shock proteins and antioxidant enzyme genes of Aphidius gifuensis in response to thermal stress. Front. Physiol. 2017, 8, 976. [Google Scholar] [CrossRef] [Green Version]
- Zhang, G.-H.; Liu, H.; Wang, J.-J.; Wang, Z.-Y. Effects of thermal stress on lipid peroxidation and antioxidant enzyme activities of the predatory mite, Neoseiulus cucumeris (Acari: Phytoseiidae). Exp. Appl. Acarol. 2014, 64, 73–85. [Google Scholar] [CrossRef]
- Ju, R.-T.; Wang, F.; Li, B. Effects of temperature on the development and population growth of the sycamore lace bug, Corythucha ciliata. J. Insect Sci. 2011, 11, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Marchler-Bauer, A.; Derbyshire, M.K.; Gonzales, N.R.; Lu, S.; Chitsaz, F.; Geer, L.Y.; Geer, R.C.; He, J.; Gwadz, M.; Hurwitz, D.I.; et al. CDD: NCBI’s conserved domain database. Nucleic. Acids. Res. 2015, 43, D222–D226. [Google Scholar] [CrossRef] [Green Version]
- Bjellqvist, B.; Basse, B.; Olsen, E.; Celis, J.E. Reference points for comparisons of two-dimensional maps of proteins from different human cell types defined in a pH scale where isoelectric points correlate with polypeptide compositions. Electrophoresis 1994, 15, 529–539. [Google Scholar] [CrossRef]
- Yu, C.-S.; Lin, C.-J.; Hwang, J.K. Predicting subcellular localization of proteins for Gram-negative bacteria by support vector machines based on n-peptide compositions. Protein Sci. 2004, 13, 1402–1406. [Google Scholar] [CrossRef] [Green Version]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [Green Version]
- Robert, X.; Gouet, P. Deciphering key features in protein structures with the new ENDscript server. Nucleic Acids Res. 2011, 42, W320–W324. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.-Z.; Fu, W.-Y.; Li, N.; Zhang, F.; Liu, T.-X. Antioxidant responses of Propylaea japonica (Coleoptera: Coccinellidae) exposed to high temperature stress. J. Insect Physiol. 2015, 73, 47–52. [Google Scholar] [CrossRef]
- Paaijmans, K.P.; Heinig, R.L.; Seliga, R.A.; Blandford, J.I.; Blanford, S.; Murdock, C.C.; Thomas, M.B. Temperature variation makes ectotherms more sensitive to climate change. Glob. Chang. Biol. 2013, 19, 2373–2380. [Google Scholar] [CrossRef] [Green Version]
- Colinet, H.; Sinclair, B.J.; Vernon, P.; Renault, D. Insects in fluctuating thermal environments. Annu. Rev. Entomol. 2015, 60, 123–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- An, M.I.; Choi, C.Y. Activity of antioxidant enzymes and physiological responses in ark shell, Scapharca broughtonii, exposed to thermal and osmotic stress: Effects on hemolymph and biochemical parameters. Comp. Biochem. Physiol. Part B. Biochem. Mol. Biol. 2010, 155, 34–42. [Google Scholar] [CrossRef] [PubMed]
- Rajarapu, S.P.; Mamidala, P.; Herms, D.A.; Bonello, P.; Mittapalli, O. Antioxidant genes of the emerald ash borer (Agrilus planipennis): Gene characterization and expression profiles. J. Insect Physiol. 2010, 57, 819–824. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Wang, W.-N.; Wang, A.-L. Effects of dietary vitamin E supplementation on antioxidant enzyme activities in Litopenaeus vannamei exposed to acute salinity changes. Aquaculture 2007, 265, 351–358. [Google Scholar] [CrossRef]
- Choi, C.Y.; An, K.W.; An, M.I. Molecular characterization and mRNA expression of glutathione peroxidase and glutathione S-transferase during osmotic stress in olive flounder (Paralichthys olivaceus). Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2008, 149, 330–337. [Google Scholar] [CrossRef]
- Zhou, Y.-F.; Tang, J.-L. Lethal effect of high temperature on leafminer fly, Liriomyza huidobrensis in a greenhouse. China Bull. Entomol. 2003, 40, 372–373. [Google Scholar] [CrossRef]
- Lu, Z.-Z.; Zhang, J.-G.; Luo, L.; Gao, G.-Z.; Lu, G.-L. Effect of high temperature on survival of Aphis gossypii. China Bull. Entomol. 2010, 47, 685–689. [Google Scholar] [CrossRef]
- Palter, K.B.; Watanabe, M.; Stinson, L.; Mahowald, A.P.; Craig, E.A. Expression and localization of Drosophila melanogaster hsp70 cognate proteins. Mol. Cell. Biol. 1986, 6, 1187–1203. [Google Scholar] [CrossRef]
- Folk, D.G.; Zwollo, P.; Rand, D.M.; Gilchrist, G.W. Selection on knockdown performance in Drosophila melanogaster impacts thermotolerance and heat-shock response differently in females and males. J. Exp. Biol. 2006, 209, 3964–3973. [Google Scholar] [CrossRef] [Green Version]
- Krebs, R.A.; Thompson, K.A. Direct and correlated effects of selection on flight after exposure to thermal stress in Drosophila melanogaster. Genetica 2006, 128, 217–225. [Google Scholar] [CrossRef]
- Wilbert, H. The causes of sexual difference in the size of Aphelinus asychis Walker (Hym, Aphelinidae). Parasitol. Res. 2006, 32, 220–236. [Google Scholar] [CrossRef]
- Sies, H. Oxidative stress: Oxidants and antioxidants. Exp. Physiol. 1997, 82, 291–297. [Google Scholar] [CrossRef] [PubMed]
- Bafana, A.; Dutt, S.; Kumar, A.; Kumar, S.; Ahuja, P.S. The basic and applied aspects of superoxide dismutase. J. Mol. Catal. B Enzym. 2011, 68, 129–138. [Google Scholar] [CrossRef]
- Celino, F.T.; Yamaguchi, S.; Miura, C.; Ohta, T.; Tozawa, Y.; Iwai, T.; Miura, T. Tolerance of spermatogonia to oxidative stress is due to high levels of Zn and Cu/Zn superoxide dismutase. PLoS ONE 2011, 6, e16938. [Google Scholar] [CrossRef] [Green Version]
- Jia, F.-X.; Dou, W.; Hu, F.; Wang, J.-J. Effects of thermal stress on lipid peroxidation and antioxidant enzyme activities of oriental fruit fly, Bactrocera dorsalis (Diptera: Tephritidae). Fla. Entomol. 2011, 94, 956–963. [Google Scholar] [CrossRef]
- Park, M.S.; Jo, P.G.; Choi, Y.K.; An, K.W.; Choi, C.Y. Characterization and mRNA expression of Mn-SOD and physiological responses to stresses in the Pacific oyster Crassostrea gigas. Mar. Biol. Res. 2009, 5, 451–461. [Google Scholar] [CrossRef]
- Li, D.; Blasevich, F.; Theopold, U.; Schmidt, O. Possible function of two insect phospholipid-hydroperoxide glutathione peroxidases. J. Insect Physiol. 2003, 49, 1–9. [Google Scholar] [CrossRef]
- Li, Z.-M.; Chen, Q.; Jin, Q.-A.; Tang, C.; Wen, H.-B.; Peng, Z.-Q. Effects of high temperature on the activities of protective enzymes in the Tetrastichus brontispae. China J. Trop. Crop. 2010, 31, 994–998. [Google Scholar] [CrossRef]
- Lu, Y.; Bai, Q.; Zheng, X.; Lu, Z.-X. Expression and enzyme activity of catalase in Chilo suppressalis (Lepidoptera: Crambidae) is responsive to environmental stresses. J. Econ. Entomol. 2017, 110, 1803–1812. [Google Scholar] [CrossRef]
- Marutani-Hert, M.; Hunter, W.B.; Hall, D.G. Gene response to stress in the asian citrus psyllid (hemiptera: Psyllidae). Fla. Entomol. 2010, 93, 519–525. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Accession Number | FPKM | Blast P Hit | E-Value | Identity (%) | |
|---|---|---|---|---|---|---|
| Catalase | AasyCAT1 | OK169278 | 118.72 | XP_031785372.1 catalase [Nasonia vitripennis] | 0.0 | 84.28% |
| Superoxide dismutase | AasySOD1 | OK169290 | 54.94 | KMQ95190.1 superoxide dismutase [Lasius niger] | 3 × 10−65 | 67.10% |
| AasySOD2 | OK169291 | 36.33 | XP_031787604.1 superoxide dismutase [Cu-Zn] isoform X3 [Nasonia vitripennis] | 3 × 10−94 | 89.54% | |
| Peroxidase | AasyPOD1 | OK169284 | 14.12 | XP_008210669.1 probable phospholipid hydroperoxide glutathione peroxidase isoform X1 [Nasonia vitripennis] | 1 × 10−109 | 90.48% |
| AasyPOD2 | OK169285 | 299.22 | XP_011503666.1 PREDICTED: phospholipid hydroperoxide glutathione peroxidase [Ceratosolen solmsi marchali] | 3 × 10−94 | 80.36% | |
| AasyPOD3 | OK169286 | 4.57 | XP_008203489.1 peroxidase [Nasonia vitripennis] | 0.0 | 77.03% | |
| AasyPOD4 | OK169287 | 35.98 | XP_008210672.1 probable phospholipid hydroperoxide glutathione peroxidase isoform X2 [Nasonia vitripennis] | 9 × 10−110 | 90.48% | |
| AasyPOD5 | OK169288 | 27.94 | XP_001606751.1 phospholipid hydroperoxide glutathione peroxidase-like [Nasonia vitripennis] | 6 × 10−57 | 52.97% | |
| AasyPOD6 | OK169289 | 7.55 | XP_008203493.1 peroxidase isoform X4 [Nasonia vitripennis] | 0.0 | 70.03% | |
| Glutathione | AasyGST1 | OK169279 | 12.59 | XP_014225564.1 glutathione-S-transferase 1-like [Trichogramma pretiosum] | 3 × 10−121 | 78.24% |
| AasyGST2 | OK169280 | 34.36 | XP_031783860.1 glutathione-S-transferase isoform X1 [Nasonia vitripennis] | 8 × 10−122 | 88.78% | |
| AasyGST3 | OK169281 | 135.33 | NP_001165920.1 glutathione-S-transferase S3 [Nasonia vitripennis] | 5 × 10−133 | 88.61% | |
| AasyGST4 | OK169282 | 19.78 | OXU29987.1 hypothetical protein TSAR_001104 [Trichomalopsis sarcophagae] | 3 × 10−44 | 50.61% | |
| AasyGST5 | OK169283 | 45.22 | XP_014215774.1 glutathione-S-transferase theta-1 isoform X3 [Copidosoma floridanum] | 7 × 10−124 | 73.68% |
| Family | Gene Name | Coding Sequence | Mw (kDa) | pI | Subcelluar Location | Strand |
|---|---|---|---|---|---|---|
| Catalase | AasyCAT1 | 1818 | 67.8 | 9.00 | Mitochondrial | plus |
| Superoxide dismutase | AasySOD1 | 531 | 18.5 | 6.29 | Extracellular | minus |
| AasySOD2 | 675 | 23.4 | 5.88 | Cytoplasmic | plus | |
| Peroxidase | AasyPOD1 | 507 | 19.1 | 6.90 | Cytoplasmic | minus |
| AasyPOD2 | 579 | 21.7 | 6.44 | Cytoplasmic | minus | |
| AasyPOD3 | 2115 | 79.4 | 6.13 | Nuclear | minus | |
| AasyPOD4 | 576 | 21.6 | 8.61 | Cytoplasmic | minus | |
| AasyPOD5 | 552 | 20.0 | 5.37 | Cytoplasmic | plus | |
| AasyPOD6 | 2325 | 86.3 | 7.78 | Extracellular | minus | |
| Glutathione-S-transferase | AasyGST1 | 654 | 25.0 | 6.63 | Cytoplasmic | plus |
| AasyGST2 | 624 | 23.8 | 6.45 | Cytoplasmic | plus | |
| AasyGST3 | 609 | 23.0 | 6.00 | Cytoplasmic | minus | |
| AasyGST4 | 1014 | 38.6 | 11.74 | Nuclear | minus | |
| AasyGST5 | 684 | 26.6 | 7.01 | Cytoplasmic | plus |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, X.; Fu, Z.-X.; Kang, Z.-W.; Li, H.; Liu, T.-X.; Wang, D. Identification and Characterization of Antioxidant Enzyme Genes in Parasitoid Aphelinus asychis (Hymenoptera: Aphelinidae) and Expression Profiling Analysis under Temperature Stress. Insects 2022, 13, 447. https://doi.org/10.3390/insects13050447
Liu X, Fu Z-X, Kang Z-W, Li H, Liu T-X, Wang D. Identification and Characterization of Antioxidant Enzyme Genes in Parasitoid Aphelinus asychis (Hymenoptera: Aphelinidae) and Expression Profiling Analysis under Temperature Stress. Insects. 2022; 13(5):447. https://doi.org/10.3390/insects13050447
Chicago/Turabian StyleLiu, Xiang, Zhi-Xiao Fu, Zhi-Wei Kang, Hao Li, Tong-Xian Liu, and Dun Wang. 2022. "Identification and Characterization of Antioxidant Enzyme Genes in Parasitoid Aphelinus asychis (Hymenoptera: Aphelinidae) and Expression Profiling Analysis under Temperature Stress" Insects 13, no. 5: 447. https://doi.org/10.3390/insects13050447