
Gut Bacteria Promote Phosphine Susceptibility of Tribolium castaneum by Aggravating Oxidative Stress and Fitness Costs
School of Food and Strategic Reserves, Henan University of Technology, Zhengzhou 450001, China
*
Author to whom correspondence should be addressed.
Insects 2023, 14(10), 815; https://doi.org/10.3390/insects14100815
Submission received: 17 August 2023
/
Revised: 9 October 2023
/
Accepted: 13 October 2023
/
Published: 15 October 2023
(This article belongs to the Section Insect Behavior and Pathology)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
The red flour beetle, Tribolium castaneum, has developed widespread pesticide resistance. Knowledge about resistance mechanisms can provide ideas for pesticide resistance management. Since gut microbes can affect host pesticide resistance, we explored gut bacteria-mediated phosphine susceptibility in T. castaneum and its molecular basis. Among five cultivable gut bacteria excised from a phosphine-resistant T. castaneum, only Enterococcus sp. inoculation significantly promoted host susceptibility to phosphine, while inoculation of any other gut bacteria did not. Furthermore, when T. castaneum was exposed to phosphine, Enterococcus sp. inoculation decreased the female fecundity, promoted host oxidative stress, and suppressed the expression and activity of host antioxidant enzymes. In the absence of phosphine, Enterococcus sp. inoculation also elicited overactive host immune responses, including the dual oxidase–reactive oxygen species system. These results indicate that Enterococcus sp. likely promotes host phosphine susceptibility by aggravating oxidative stress and fitness costs.
Abstract
Knowledge about resistance mechanisms can provide ideas for pesticide resistance management. Although several studies have unveiled the positive or negative impacts of gut microbes on host pesticide resistance, minimal research is available regarding the association between gut microbes and host phosphine resistance. To explore the influence of gut bacteria on host phosphine susceptibility and its molecular basis, mortality, fitness, redox responses, and immune responses of adult Tribolium castaneum were determined when it was challenged by phosphine exposure and/or gut bacteria inoculation. Five cultivable gut bacteria were excised from a population of phosphine-resistant T. castaneum. Among them, only Enterococcus sp. inoculation significantly promoted host susceptibility to phosphine, while inoculation of any other gut bacteria had no significant effect on host phosphine susceptibility. Furthermore, when T. castaneum was exposed to phosphine, Enterococcus sp. inoculation decreased the female fecundity, promoted host oxidative stress, and suppressed the expression and activity of host superoxide dismutase, catalase, and peroxidase. In the absence of phosphine, Enterococcus sp. inoculation also elicited overactive immune responses in T. castaneum, including the immune deficiency and Toll signaling pathways and the dual oxidase–reactive oxygen species system. These results indicate that Enterococcus sp. likely promotes host phosphine susceptibility by aggravating oxidative stress and fitness costs.
1. Introduction
The red flour beetle, Tribolium castaneum (Herbst, 1797) (Coleoptera: Tenebrionidae), is one of the most damaging pests, which can infest 246 grain commodities and cause economic losses in warehouses and mills worldwide [1]. Product losses posed by T. castaneum infestations are up to 34–40% [2]. Owing to the magnitude of such losses, it has been controlled with various pesticides over a long period. However, T. castaneum can easily accommodate to frequently used pesticides, including contact pesticides and the fumigant phosphine. It has developed diverse strategies, including metabolic resistance, increased efflux capacities, and target site insensitivity, to deal with pesticides [3]. Therefore, knowledge of resistance mechanisms is crucial for the development of novel pest management strategies [4]. Recently, the association between gut microbes and host resistance to pesticides and its possible mechanisms have been unveiled in many studies [5], which provides a direction for investigating resistance mechanisms.
Phosphine can suppress the activity of electron transfer chain enzymes and antioxidant enzymes such as catalase and peroxidase, thus eliciting massive accumulation of highly reactive oxygen species (ROS), which contributes to phosphine toxicity [3]. Additionally, considering that infection with a high load of gut bacteria commonly elicits overactive immune responses, subsequently promoting host oxidative stress via a sudden boost of humoral immune effectors such as ROS [6,7], it is assumed that some bacteria can modify host phosphine susceptibility by intervening in the redox system. Furthermore, tradeoffs exist between physiological traits due to competition for limiting resources, i.e., increases in the fitness value of one trait may lead to a corresponding decline in the fitness value of another [8]. Thus, it is speculated that when pests are simultaneously challenged by pesticide exposure and bacterial infection, the tradeoff between immune responses and other fitness traits associated with pesticide resistance will increase host susceptibility to pesticides, as has been found in Diaphorina citri infested with Candidatus Liberibacter asiaticus [9].
Although several studies have unveiled the positive or negative impacts of gut microbes on host pesticide resistance [5,10], minimal research is available regarding the association between gut microbes and host phosphine resistance [11,12]. Herein, we aim to explore gut bacteria-mediated phosphine susceptibility in T. castaneum and its molecular basis, with emphasis on host oxidative stress and fitness costs, through a four-stage research process. When T. castaneum was challenged by phosphine exposure and/or gut bacteria inoculation, (1) the mortality of T. castaneum was measured to unveil the impact of gut bacteria on host phosphine resistance and to screen resistance-mediating gut bacteria, (2) the female fecundity of T. castaneum was measured to unveil the impact of resistance-mediating gut bacteria on host reproductive fitness, (3) the level of hydrogen peroxide and malondialdehyde in T. castaneum was measured to unveil the impact of resistance-mediating gut bacteria on host oxidative stress, and (4) antioxidant and immune responses in T. castaneum were measured to unveil the molecular basis for gut bacteria-induced oxidative stress.
2. Materials and Methods
2.1. Insects
The stock of T. castaneum originated from a population sampled from Shantou, Guangdong, China, in June 2015 and was cultured continuously in laboratory conditions for more than seven years. This stock was strongly resistant to phosphine with a resistance ratio of 208.3, determined via the FAO standard method [13]. The normal diet for the insects was a blend of wholemeal flour and yeast powder (19:1, w:w) sterilized via exposure to ultraviolet radiation (about 254 nm) for 2 h. Insects were cultured at 28 °C and 65% relative humidity in continuous darkness. The pupae were collected and then separated by sex as previously described for subsequent assays [2]. If there were no special instructions, experiments with T. castaneum were performed under the same normal conditions mentioned above.
2.2. Isolation and Identification of Gut Bacteria
To isolate gut bacteria, 7-day-old mixed-sex adult T. castaneum were starved for 1 d, then surface disinfected with 70% alcohol, and subsequently rinsed thrice with aseptic water. Gut bacteria were excised and purified as previously described [14]. The purified gut bacteria were collected and preserved in Luria-Bertani (LB) broth added with 25% glycerine at −80 °C for subsequent assays. Bacterial isolates were revived in LB broth prior to use. Five cultivable gut bacteria were successfully excised from the tested T. castaneum. Bacterial identification based on 16S rDNA genes was conducted as previously described [15]. The 16S rDNA gene primers 27F and 1492R were used to amplify DNA fragments (Supplementary Table S1). Amplified DNA fragments were sequenced and blasted against the 16S rDNA database and submitted to the NCBI database, and their GenBank accession numbers were OM992224.1, OM992227.1, OM992228.1, OM992229.1, and OM992230.1 (Supplementary Figure S1).
2.3. Antibiotic Treatment and Gnotobiotic Inoculation
Since antibiotics could reshape and deplete insect gut bacteria [16], newly emerged beetles were cultured with a normal diet containing 20 μg/g antibiotic cocktail (ciprofloxacin:penicillin:vancomycin = 1:1:1) for 3 d to produce germ-free beetles. After antibiotic treatment, 4-day-old germ-free beetles were cultured with a normal diet mixed with a given bacterial isolate for 3 d to produce gnotobiotic beetles [15]. All treatments were conducted in an aseptic environment. Germ-free and gnotobiotic beetles were then cultured with a normal diet in an aseptic environment for subsequent assays. The beetles cultured with a normal diet under normal conditions were used as conventional beetles.
To confirm whether gut bacteria were successfully depleted and inoculated, their abundance was determined via real-time quantitative PCR (qRT-PCR) using the 2−ΔΔCT method [17] and normalized by the qRT-PCR data of the β-actin gene of T. castaneum. Gut homogenates of thirty 6-, 8-, 10-, 12-, 14-, and 16-day-old beetles were prepared according to the method described above. Total DNA was extracted from the gut homogenate using an E.Z.N.A.® soil DNA kit (Omega Bio-Tek, Norcross, GA, USA). qRT-PCR primers are shown in Supplementary Table S1. The qRT-PCR was run in a 20 μL reactive mixture including 10 μL of SYBR Green Mix (TaKaRa, Otsu, Japan), 0.8 μL of each primer (50 μmol/L), and 5 ng of template DNA. Thermocycling conditions were 95 °C for 30 s, and then 35 cycles of 95 °C for 5 s, 60 °C for 30 s, and 72 °C for 30 s. After the thermal cycles, melting curve analysis was conducted to confirm that the reaction produced a single, specific product. The thermocycling programs for the melting curve analysis were the same as those previously described [6]. Reactions were conducted in triplicate, and the mean of triplicate analyses represented one biological repetition. Each treatment consisted of five biological repetitions.
2.4. Bioassay of Phosphine Susceptibility
Phosphine susceptibility of conventional, germ-free, and gnotobiotic beetles was measured according to the FAO standard method [13] to unveil the impact of bacteria inoculation on host phosphine resistance. Thirty 10-day-old beetles were exposed to phosphine at the concentrations of 0, 1000, 1250, 1500, 1750, and 2000 mL/m3 for 20 h. After exposure to phosphine, the beetles were ventilated and then cultured under normal conditions. Their mortality was assessed after 14 d of culture and corrected for control mortality [18]. Each treatment consisted of five biological repetitions. Since only Enterococcus sp. significantly influenced host phosphine susceptibility, it was used in subsequent assays to explore the molecular basis for gut bacteria-mediated phosphine susceptibility in T. castaneum.
2.5. Determination of Female Fecundity
Female fecundity of conventional, germ-free, and gnotobiotic beetles was measured to assess the impact of Enterococcus sp. inoculation on reproductive fitness of T. castaneum after exposure to phosphine. After exposure of 20 pairs of 10-day-old beetles to 1250 mL/m3 phosphine for 20 h, the beetles were ventilated and then cultured under normal conditions. Pairs were separately cultured with a normal diet in a culture plate with a diameter of 15 mm and checked daily for 45 d, on which most females did not lay eggs anymore. The diet was changed every 2 d. Freshly laid eggs were transferred to a new culture plate and monitored for 10 d for emergence. Female fecundity was defined as the number of viable eggs laid per female. The average fecundity of 20 pairs represented one biological repetition. Each treatment consisted of five biological repetitions.
2.6. Qualification of MDA and H2O2
Since malondialdehyde (MDA) and hydrogen peroxide (H2O2) are commonly used as biological indicators of oxidative stress [19,20], the level of MDA and H2O2 in conventional, germ-free, and gnotobiotic beetles was measured to assess the impact of Enterococcus sp. inoculation on oxidative stress in T. castaneum after exposure to phosphine. After exposure of 10-day-old beetles to 1250 mL/m3 phosphine for 0, 6, 12, and 24 h, the level of MDA and H2O2 was determined using BC0020 and BC3590 assay kits (Solarbio, Beijing, China), respectively. MDA reacts with thiobarbituric acid to produce tridione with a maximum absorbance at 532 nm, and H2O2 reacts with titanic sulfate to produce a titanium peroxide complex with characteristic absorbance at 415 nm [21]. The level of MDA and H2O2 was calculated based on the absorbance according to the assay kit protocol. The samples were conducted in triplicate, and the mean of triplicate analyses represented one biological repetition. Each treatment consisted of five biological repetitions.
2.7. Determination of SOD, CAT, and POD Activities
The activity of three antioxidant enzymes, namely, superoxide dismutase (SOD), catalase (CAT), and peroxidase (POD), in conventional, germ-free, and gnotobiotic beetles was measured to assess the impact of Enterococcus sp. inoculation on the antioxidant system of T. castaneum after exposure to phosphine. After exposure of 10-day-old beetles to 1250 mL/m3 phosphine for 0, 6, 12, and 24 h, the activity of SOD, CAT, and POD was determined using BC0170, BC0200, and BC0090 assay kits (Solarbio) according to the protocols of the manufacturer, respectively.
SOD activity was determined by detecting superoxide radicals generated by xanthine oxidase and hypoxanthine and monitoring absorbance at 560 nm [22]. One unit of SOD activity refers to the quantity of enzyme necessary to display 50% dismutation of the superoxide radical. CAT activity was determined by detecting the decomposition rate of H2O2 and monitoring absorbance at 240 nm [23]. One unit of CAT activity refers to the quantity of CAT necessary to decompose 1 µmol of H2O2 per min per g of sample. POD activity was determined by detecting guaiacol oxidation and monitoring absorbance at 470 nm [24]. One unit of POD activity refers to a 0.01 change in absorbance at 470 nm per min per g of sample per mL of the reactive mixture. The samples were conducted in triplicate, and the mean of triplicate analyses represented one biological repetition. Each treatment consisted of five biological repetitions.
2.8. Determination of SOD, CAT, and POD Gene Expression
The expression levels of SOD, CAT, and POD genes in conventional, germ-free, and gnotobiotic beetles were measured to explore the molecular basis for the impact of Enterococcus sp. inoculation on the antioxidant system in T. castaneum after exposure to phosphine. After exposure of 10-day-old beetles to 1250 mL/m3 phosphine for 0, 6, 12, and 24 h, the total RNA of 50 beetles was extracted using TRIzol® reagent (Accurate Biotechnology, Changsha, China). Contaminating genomic DNA was depleted using DNase I (Accurate Biotechnology). Subsequently, cDNA libraries were constructed from 1 μg of RNA using M-MLV Reverse Transcriptase (Accurate Biotechnology). The relative gene expression data were assessed using the 2−ΔΔCT method and normalized with the qRT-PCR data of the Rps18 gene of T. castaneum [25]. qRT-PCR primers are shown in Supplementary Table S1. The 20 μL qRT-PCR reactive mixture included 10 μL of SYBR Green Mix (TaKaRa, Otsu, Japan), 0.8 μL of each primer (50 μmol/L), and 2 μL of cDNA (diluted 1:10). qRT-PCR conditions were the same as those described above. Reactions were conducted in triplicate, and the mean of triplicate analyses represented one biological repetition. Each treatment consisted of five biological repetitions.
2.9. Measurement of Immune Responses
The expression levels of PLCβ, Atf2, and DUOX from the dual oxidase-reactive oxygen species (DUOX-ROS) system, IMD from the immune deficiency (IMD) signaling pathway, and Toll from the Toll signaling pathway in conventional, germ-free, and gnotobiotic beetles were measured to assess the impact of Enterococcus sp. inoculation on host immune responses in the absence of phosphine. The total RNA of fifty 6-, 8-, 10-, 12-, 14-, and 16-day-old beetles was sequentially extracted, processed, and analyzed according to the method described above. qRT-PCR primers are shown in Supplementary Table S1. Reactions were conducted in triplicate, and the mean of triplicate analyses represented one biological repetition. Each treatment consisted of five biological repetitions.
2.10. Statistical Analysis
Datasets were tested for normality using the Kolmogorov–Smirnov test. All datasets were normally distributed and statistically compared with one-way analysis of variance (ANOVA) and Tukey’s honest significant difference (HSD) test. All statistics were conducted using SPSS Statistics 22.0 (IBM, NY, USA), and differences among means were considered significant at p < 0.05.
3. Results
3.1. Impact of Bacterial Treatment on Bacterial Abundance
Five cultivable bacteria, namely, Brevibacterium sp., Enterococcus sp., Microbacterium sp., Micrococcus sp., and Staphylococcus sp., were excised from the gut of phosphine-resistant T. castaneum (Supplementary Figure S1). Since germ-free and gnotobiotic beetles were used to assess the impact of Enterococcus sp. on host traits, the methodology for producing these beetles required technical approval. The gut load of Brevibacterium sp., Enterococcus sp., Microbacterium sp., Micrococcus sp., and Staphylococcus sp. (Figure 1) varied substantially among conventional, germ-free, and gnotobiotic beetles (F2,12 = 68.40, p < 0.001; F2,12 = 55.57, p < 0.001; F2,12 = 10.60, p = 0.002; F2,12 = 84.91, p < 0.001; F2,12 = 21.46, p < 0.001, respectively). Five gut bacteria were successfully depleted via antibiotic treatment to produce germ-free beetles, and the gut load of a given bacteria was restored to its normal level after inoculation of germ-free beetles with the bacterial isolate, thus producing gnotobiotic beetles.
3.2. Impact of Bacteria Inoculation on Host Phosphine Susceptibility
The impact of five gut bacterial isolates on host phosphine susceptibility was assessed to screen resistance-mediating bacteria. Enterococcus sp. inoculation significantly promoted host susceptibility to phosphine, and the mortality of the beetles inoculated with Enterococcus sp. was substantially higher than those of conventional and germ-free beetles after exposure to 1000, 1250, 1500, 1750, and 2000 mL/m3 phosphine for 20 h (F6,28 = 4.09, p = 0.005; F6,28 = 42.72, p < 0.001; F6,28 = 13.21, p < 0.001; F6,28 = 15.54, p < 0.001; F6,28 = 4.26, p = 0.004, respectively) (Figure 2). Moreover, inoculation of any other gut bacterial isolate had no significant effect on host phosphine susceptibility, and no significant difference was found in the mortality between germ-free and gnotobiotic beetles. Therefore, Enterococcus sp. was used in subsequent assays to explore molecular mechanisms of gut bacteria-induced phosphine susceptibility in T. castaneum.
3.3. Impact of Enterococcus sp. Inoculation on Host Fecundity
Enterococcus sp. inoculation significantly reduced the female fecundity of T. castaneum. After exposure of beetles to 1250 mL/m3 phosphine for 20 h, the number of eggs laid per female of gnotobiotic beetles was substantially lower than that of conventional and germ-free beetles (F2,57 = 106.20, p < 0.001) (Figure 3), suggesting that Enterococcus sp. inoculation results in fitness costs in T. castaneum after exposure to phosphine.
3.4. Impact of Enterococcus sp. Inoculation on Host Oxidative Stress
After exposure to 1250 mL/m3 phosphine for 0, 6, 12, and 24 h, the level of MDA and H2O2 in T. castaneum was measured to assess the impact of phosphine exposure and Enterococcus sp. inoculation on host oxidative stress. Phosphine exposure induced serious oxidative stress in T. castaneum, and the MDA level (Figure 4A) in conventional, germ-free, and gnotobiotic beetles increased as the phosphine exposure period was extended (F3,16 = 33.78, p < 0.001; F3,16 = 4.64, p = 0.016; F3,16 = 24.30, p < 0.001, respectively). Enterococcus sp. inoculation further promoted oxidative stress, and the MDA level in gnotobiotic beetles was substantially higher than that in germ-free beetles after exposure to phosphine for 6, 12, and 24 h (F2,12 = 15.67, p < 0.001; F2,12 = 45.98, p < 0.001; F2,12 = 13.44, p = 0.001, respectively).
Phosphine-induced oxidative stress in T. castaneum was further verified by massive accumulation of H2O2. The H2O2 level in conventional and germ-free beetles (Figure 4B) increased as the phosphine exposure period was extended (F3,16 = 10.64, p < 0.001; F3,16 = 10.41, p < 0.001, respectively). However, Enterococcus sp. inoculation inhibited H2O2 production in T. castaneum after exposure to phosphine, and the H2O2 level in gnotobiotic beetles was substantially lower than that in germ-free beetles after exposure to phosphine for 24 h (F2,12 = 8.60, p = 0.005).
3.5. Impact of Enterococcus sp. Inoculation on the Host Antioxidant System
The impact of Enterococcus sp. inoculation on the activity of SOD, CAT, and POD in T. castaneum was assessed after exposure to 1250 mL/m3 phosphine for 0, 6, 12, and 24 h. Phosphine exposure promoted SOD activity in conventional, germ-free, and gnotobiotic beetles, which increased as the phosphine exposure period was extended (F3,16 = 25.56, p < 0.001; F3,16 = 61.87, p < 0.001; F3,16 = 11.28, p < 0.001, respectively) (Figure 5A). However, Enterococcus sp. inoculation suppressed SOD activity in T. castaneum after exposure to phosphine. SOD activity in gnotobiotic beetles was substantially lower than that in germ-free beetles after exposure to phosphine for 6 and 24 h (F2,12 = 6.80, p = 0.011; F2,12 = 31.60, p < 0.001, respectively).
However, phosphine exposure suppressed the activity of CAT (Figure 5B) and POD (Figure 5C), which declined in conventional, germ-free, and gnotobiotic beetles as the phosphine exposure period was extended (F3,16 = 748.39, p < 0.001; F3,16 = 768.52, p < 0.001; F3,16 = 424.97, p < 0.001 for CAT, respectively; F3,16 = 33.38, p < 0.001; F3,16 = 431.92, p < 0.001; F3,16 = 1474.92, p < 0.001 for POD, respectively). Enterococcus sp. inoculation further suppressed the activity of CAT and POD. CAT activity in gnotobiotic beetles was substantially lower than that in germ-free beetles after exposure to phosphine for 6 and 24 h (F2,12 = 46.52, p < 0.001; F2,12 = 19.91, p < 0.001, respectively), and POD activity in gnotobiotic beetles was substantially lower than that in germ-free beetles after exposure to phosphine for 6, 12, and 24 h (F2,12 = 24.26, p < 0.001; F2,12 = 27.93, p < 0.001; F2,12 = 9.83, p = 0.003, respectively).
The expression levels of SOD1a, SOD1b, SOD1c (three transcript variants of the SOD1 gene in T. castaneum), SOD2, CAT3, and PRDX6 in T. castaneum were measured after exposure to 1250 mL/m3 phosphine for 0, 6, 12, and 24 h (Figure 6). Phosphine exposure led to upregulation of SOD gene expression. The expression levels of SOD1a, SOD1b, SOD1c, and SOD2 first increased and then decreased in conventional, germ-free, and gnotobiotic beetles as the phosphine exposure period was extended (F3,16 = 47.88, p < 0.001; F3,16 = 18.37, p < 0.001; F3,16 = 7.12, p = 0.003 for SOD1a, respectively; F3,16 = 94.45, p < 0.001; F3,16 = 179.90, p < 0.001; F3,16 = 29.06, p < 0.001 for SOD1b, respectively; F3,16 = 54.07, p < 0.001; F3,16 = 21.35, p < 0.001; F3,16 = 29.63, p < 0.001 for SOD1c, respectively; F3,16 = 123.62, p < 0.001; F3,16 = 54.13, p < 0.001; F3,16 = 3.92, p = 0.028 for SOD2, respectively), reaching the maximal values after exposure to phosphine for 24, 12, 6, and 6 h, respectively.
However, Enterococcus sp. inoculation led to downregulation of SOD gene expression in T. castaneum after exposure to phosphine. The expression level of SOD1a in gnotobiotic beetles was substantially lower than that in germ-free beetles after exposure to phosphine for 6, 12, and 24 h (F2,12 = 39.10, p < 0.001; F2,12 = 72.94, p < 0.001; F2,12 = 19.18, p < 0.001, respectively). Similarly, the expression level of SOD1b in gnotobiotic beetles was substantially lower than that in germ-free beetles after exposure to phosphine for 12 and 24 h (F2,12 = 53.72, p < 0.001; F2,12 = 28.17, p < 0.001, respectively), the expression level of SOD1c in gnotobiotic beetles was substantially lower than that in germ-free beetles after exposure to phosphine for 6 h (F2,12 = 31.13, p < 0.001), and the expression level of SOD2 in gnotobiotic beetles was substantially lower than that in germ-free beetles after exposure to phosphine for 6 and 12 h (F2,12 = 243.76, p < 0.001; F2,12 = 7.41, p = 0.008, respectively).
Similar variation trends in CAT and POD gene expression occurred in T. castaneum when challenged by phosphine exposure and Enterococcus sp. inoculation. The expression level of CAT3 first increased and then decreased in conventional and gnotobiotic beetles as the phosphine exposure period was extended (F3,16 = 9.12, p = 0.001; F3,16 = 4.60, p = 0.017, respectively). The expression level of PRDX6 first increased and then decreased in conventional and germ-free beetles as the phosphine exposure period was extended (F3,16 = 219.17, p < 0.001; F3,16 = 174.68, p < 0.001, respectively). The expression levels of CAT3 and PRDX6 reached the maximal values after exposure to phosphine for 6 h. Enterococcus sp. inoculation led to downregulation of the expression of CAT3 and PRDX6 in T. castaneum after exposure to phosphine. The expression level of CAT3 in gnotobiotic beetles was substantially lower than that in germ-free beetles after exposure to phosphine for 12 h (F2,12 = 11.03, p < 0.001), and the expression level of PRDX6 in gnotobiotic beetles was substantially lower than that in germ-free beetles after exposure to phosphine for 0, 6, and 12 h (F2,12 = 6.91, p = 0.010; F2,12 = 176.51, p < 0.001; F2,12 = 5.29, p = 0.023, respectively).
3.6. Impact of Enterococcus sp. Inoculation on Host Immune Responses
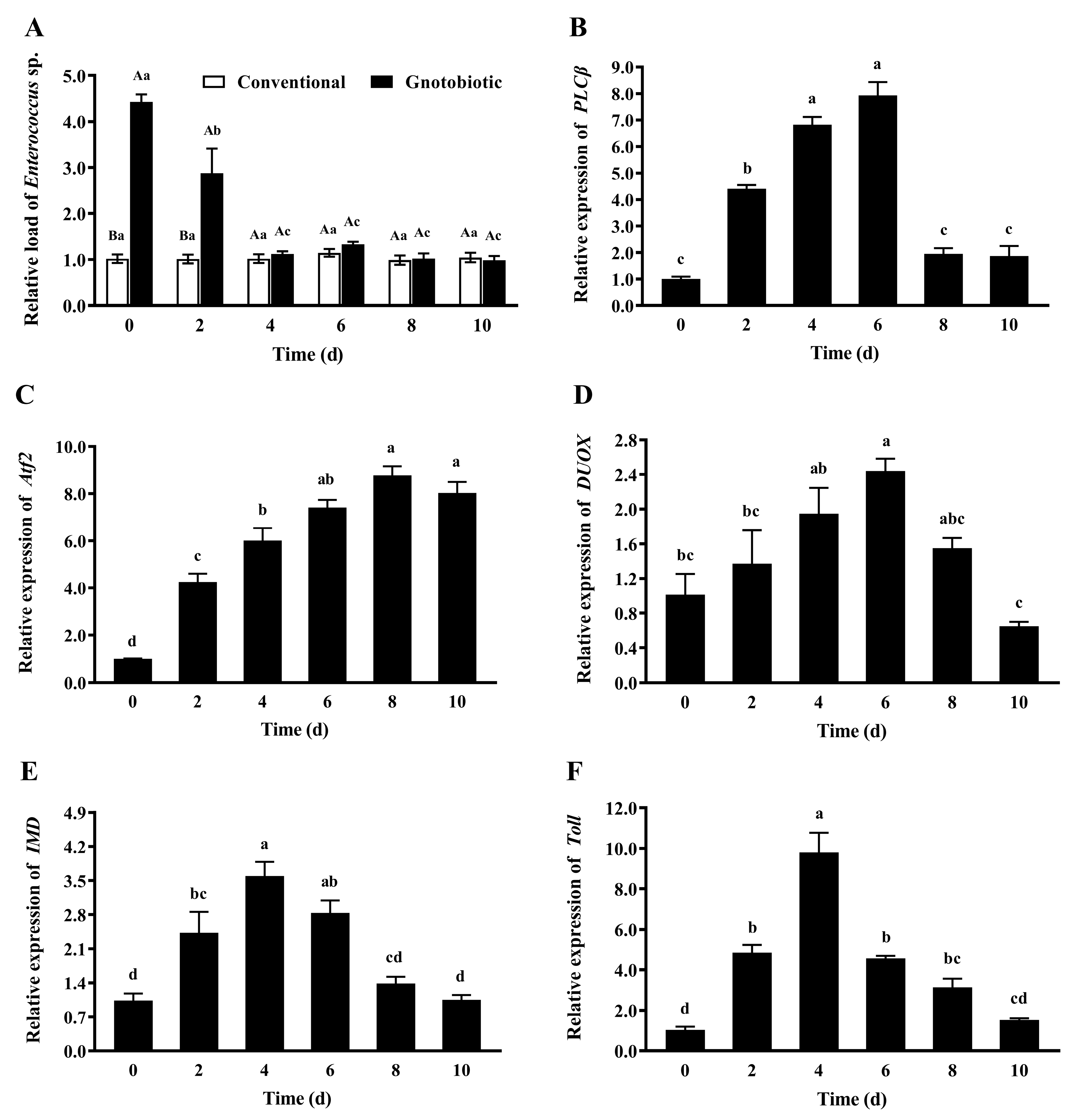
To assess the impact of Enterococcus sp. inoculation on host immune responses, the gut load of Enterococcus sp. and the expression levels of PLCβ, Atf2, DUOX, IMD, and Toll in gnotobiotic T. castaneum were evaluated on days 0, 2, 4, 6, 8, and 10 after inoculation of Enterococcus sp. to germ-free beetles. After Enterococcus sp. was inoculated into germ-free beetles, the gut load of Enterococcus sp. in gnotobiotic beetles (Figure 7A) was significantly higher than that in conventional beetles during the first two days after bacteria inoculation (F1,8 = 313.73, p < 0.001; F1,8 = 11.77, p = 0.009) and recovered to the normal level from the fourth day after bacteria inoculation (F1,8 = 0.89, p = 0.374 for the fourth day; F1,8 = 3.40, p = 0.103 for the sixth day; F1,8 = 0.06, p = 0.808 for the eighth day; F1,8 = 0.19, p = 0.674 for the tenth day).
The expression levels of PLCβ (Figure 7B), Atf2 (Figure 7C), DUOX (Figure 7D), IMD (Figure 7E), and Toll (Figure 7F) first increased and then decreased (F5,24 = 91.72, p < 0.001; F5,24 = 56.96, p < 0.001; F5,24 = 7.46, p < 0.001; F5,24 = 17.71, p < 0.001; F5,24 = 45.87, p < 0.001, respectively), reaching the maximal values on days 6, 8, 6, 4, and 4 after bacteria inoculation, respectively. These results suggest that T. castaneum recruits different immune signaling pathways to cope with a high gut load of Enterococcus sp. and maintain microbial homeostasis.
4. Discussion
Five cultivable gut bacteria, namely, Brevibacterium sp., Enterococcus sp., Microbacterium sp., Micrococcus sp., and Staphylococcus sp., were excised from phosphine-resistant T. castaneum. Gut microbes may increase host pesticide resistance by directly degrading pesticides [10], modulating the host immune system [26], and producing nutrients and other beneficial chemicals [27,28]. However, Enterococcus faecalis inoculation impaired phosphine resistance of adult T. castaneum [12]. In this study, inoculation with another Enterococcus sp. also promoted host phosphine susceptibility, while inoculation of any other gut bacteria had no significant effect on host phosphine susceptibility. The negative influence of gut microbes on host pesticide resistance to contact pesticides, which has also been found in other studies [29,30], will provide ideas for pesticide resistance management by integrating pesticides with biocontrol agents containing resistance-mediating bacteria [31,32].
Phosphine exposure elicited an increased level of MDA and H2O2, indicating high oxidative stress in T. castaneum. On the other hand, inoculation with a high load of Enterococcus sp. upregulated the expression of PLCβ, Atf2, and DUOX from the DUOX–ROS system in T. castaneum, further aggravating oxidative stress in T. castaneum [7]. Furthermore, when T. castaneum was exposed to phosphine, Enterococcus sp. inoculation suppressed the activity of host antioxidant enzymes, including SOD, CAT, and POD, which play a vital role in scavenging ROS [33]. Additionally, since SOD is responsible for the transformation of active superoxide anions into stable H2O2, a decreased level of H2O2 formation in T. castaneum due to the suppression of SOD activity by Enterococcus sp. inoculation will further aggravate oxidative stress. Considering that phosphine-induced oxidative stress and its damage are considered to be among the primary mechanisms of phosphine toxicity [34], it is not difficult to infer that increased mortality rates of T. castaneum simultaneously challenged by phosphine exposure and Enterococcus sp. inoculation could be attributed to overgeneration of ROS, which will cause damage to essential cellular building blocks, including DNA, proteins, and lipids [35].
Besides the DUOX–ROS system, other immune responses, including IMD and Toll signaling pathways, were also activated by Enterococcus sp. inoculation. As an essential life-preserving process, the immune response is privileged over other processes regarding energy supply [36]. The tradeoff between immune responses and other fitness traits associated with pesticide resistance because of alternative allocation of limiting energetic resources will increase host susceptibility to pesticides. For example, infection of Culex pipiens with the bacterium Wolbachia increased the fitness cost of resistance to pesticides, rendering hosts more susceptible to pesticides [37], and parasitism by Vavraia culicis caused fitness costs in Culex pipiens quinquefasciatus bearing organophosphate resistance alleles, potentially altering the strength and direction of selection against resistance mutations in untreated environments [38]. Since both antioxidant and immune responses are energetically costly [8,39], it is speculated that tradeoff between them will lead to Enterococcus sp.-induced phosphine susceptibility in T. castaneum.
During the initial stage of phosphine fumigation, T. castaneum attempted to mitigate oxidative stress from phosphine by upregulating the expression of SOD1a, SOD1b, SOD1c, SOD2, CAT3, and PRDX6 and promoting SOD activity, as has been found in other phosphine-resistant insects [40]. However, Enterococcus sp. inoculation constantly suppressed the expression and activity of SOD, CAT, and POD in T. castaneum after exposure to phosphine, demonstrating that beetles inoculated with Enterococcus sp. could not allocate enough resources to contradict oxidative stress due to tradeoff between immune and antioxidant responses. Similarly, infection of D. citri with Ca. Liberibacter asiaticus reduced the activity of host detoxifying enzymes such as glutathione-S-transferase (GST) and cytochrome P450 [31], and infection with a high titer of Phytoplasma inhibited the activity of host detoxifying enzymes such as β-esterase and GST in Amplicephalus curtulus [41]. Meanwhile, Enterococcus sp. inoculation decreased reproductive fitness of T. castaneum after exposure to phosphine, suggesting that beetles could not allocate enough resources to conduct normal physiological activities. Increased mortality, reduced antioxidant enzyme activity, and low fecundity indicate increased fitness costs in T. castaneum simultaneously challenged by phosphine exposure and Enterococcus sp. inoculation. From an evolutionary perspective, gut microbe-induced fitness costs will delay the development of phosphine resistance among insect populations.
Previous research showed that bacteria inoculation can activate the Toll signaling pathway and subsequently suppress the insulin signaling pathway [36]. The low expression of AGC kinase (Akt), an element in the insulin signaling pathway, can inhibit the activity of the downstream transcription factor erythroid 2-related factor 2 (Nrf2). Thus, Nrf2 cannot translocate into the nucleus to activate antioxidant enzyme expression [42]. Therefore, it is speculated that Enterococcus sp. regulates the expression of antioxidant enzymes by sequentially activating the Toll–Akt–Nrf2 pathway. The expression level of Toll was upregulated when T. castaneum was inoculated with a high load of Enterococcus sp., partially illustrating the possibility of this regulatory mechanism. Future studies of the molecular mechanisms behind gut microbe-mediated host pesticide resistance with an emphasis on the immune system will inform development of new pesticide targets. For example, the miR-34-5p target gene in the immune pathway of Spodoptera frugiperda has been developed as a target of nucleic acid pesticides [43].
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/insects14100815/s1, Table S1: Primers used for qRT-PCR. Figure S1: Identification of gut bacterial isolates excised from phosphine-resistant Tribolium castaneum adults based on 16S rDNA sequence similarity analysis.
Author Contributions
Conceptualization, Z.W.; methodology, Z.L. and H.H.; investigation, Z.L., Z.C. and H.H.; formal analysis, S.Z.; writing—original draft, Z.W. and S.Z.; writing—review and editing, Z.W., Z.L. and Z.C.; funding acquisition, Z.W. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by the National Natural Science Foundation of China (Grant no. 32272531) and the Henan Science and Technology Research Project (Grant no. 222103810069).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Conflicts of Interest
All authors declare no conflict of interest.
References
- Hagstrum, D.W.; Subramanyam, B. Stored-Product Insect Resource; AACC International Inc.: St. Paul, MN, USA, 2009; pp. 183–184. [Google Scholar]
- Astuti, L.P.; Mutala’liah, M. Host preference of Tribolium castaneum (Herbst) on six kinds of flour. Indones. J. Entomol. 2020, 17, 149–155. [Google Scholar] [CrossRef]
- Rosner, J.; Wellmeyer, B.; Merzendorfer, H. Tribolium castaneum: A model for investigating the mode of action of insecticides and mechanisms of resistance. Curr. Pharm. Des. 2020, 26, 3554–3568. [Google Scholar] [CrossRef] [PubMed]
- Fusetto, R.; Denecke, S.; Perry, T.; O’Hair, R.A.J.; Batterham, P. Partitioning the roles of CYP6G1 and gut microbes in the metabolism of the insecticide imidacloprid in Drosophila melanogaster. Sci. Rep. 2017, 7, 11339. [Google Scholar] [CrossRef]
- Pietri, J.E.; Liang, D. The links between insect symbionts and insecticide resistance: Causal relationships and physiological tradeoffs. Ann. Entomol. Soc. Am. 2018, 111, 92–97. [Google Scholar] [CrossRef]
- Yao, Z.; Wang, A.; Li, Y.; Cai, Z.; Lemaitre, B.; Zhang, H. The dual oxidase gene BdDuox regulates the intestinal bacterial community homeostasis of Bactrocera dorsalis. ISME J. 2016, 10, 1037–1050. [Google Scholar] [CrossRef]
- Wang, Z.; Yong, H.; Zhang, S.; Liu, Z.; Zhao, Y. Colonization resistance of symbionts in their insect hosts. Insects 2023, 14, 594. [Google Scholar] [CrossRef] [PubMed]
- Schwenke, R.A.; Lazzaro, B.P.; Wolfner, M.F. Reproduction–immunity trade-offs in insects. Annu. Rev. Entomol. 2016, 61, 239–256. [Google Scholar] [CrossRef]
- Tiwari, S.; Pelz-Stelinski, K.; Stelinski, L.L. Effect of Candidatus Liberibacter asiaticus infection on susceptibility of Asian citrus psyllid, Diaphorina citri, to selected insecticides. Pest Manag. Sci. 2011, 67, 94–99. [Google Scholar] [CrossRef]
- Siddiqui, J.A.; Khan, M.M.; Bamisile, B.S.; Hafeez, M.; Qasim, M.; Rasheed, M.T.; Rasheed, M.A.; Ahmad, S.; Shahid, M.I.; Xu, Y. Role of insect gut microbiota in pesticide degradation: A review. Front. Microbiol. 2022, 13, 870462. [Google Scholar] [CrossRef] [PubMed]
- Gowda, G.B.; Patil, N.B.; Sahu, M.; Prabhukarthikeyan, S.R.; Raghu, S.; Pandi, G.P.; Adak, T.; Swain, C.K.; Pokhare, S.; Mohapatra, S.D.; et al. Differential gut bacteria in phosphine resistant and susceptible population of Tribolium castaneum (Herbst) and their biochemical and molecular characterization. Pak. J. Zool. 2022, 54, 1331–1338. [Google Scholar] [CrossRef]
- Wang, Z.; Hu, H.; Chang, Z.; Zhang, S.; Lu, Y. Molecular mechanism of Enterococcus faecalis-induced phosphine sensitivity in Tribolium castaneum (Coleoptera: Tenebrionidae). J. Econ. Entomol. 2023, 116, 1820–1829. [Google Scholar] [CrossRef] [PubMed]
- Food and Agriculture Organization. Recommended methods for the detection and measurement of resistance of agricultural pests to pesticides: Tentative method for beetles of some major pest species of stored cereals with methyl bromide and phosphine. FAO method no. 16. FAO Plant Prot. Bull. 1975, 23, 12–25. [Google Scholar]
- Wang, Z.; Zhao, Y.; Yong, H.; Liu, Z.; Wang, W.; Lu, Y. The contribution of gut bacteria to pesticide resistance of Tribolium castaneum (Herbst). J. Stored Prod. Res. 2023, 103, e102160. [Google Scholar] [CrossRef]
- Wang, Z.; Wang, W.; Lu, Y. Biodegradation of insecticides by gut bacteria isolated from stored grain beetles and its implication in host insecticide resistance. J. Stored Prod. Res. 2022, 96, e101943. [Google Scholar] [CrossRef]
- Douglas, A.E. Mycetocyte symbiosis in insects. Biol. Rev. 1989, 64, 409–434. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Abbott, W.S. A method of computing the effectiveness of an insecticide. J. Econ. Entomol. 1925, 18, 265–267. [Google Scholar] [CrossRef]
- Seif, M.M.; Madboli, A.N.; Marrez, D.A.; Aboulthana, W.M.K. Hepato-renal protective effects of Egyptian purslane extract against experimental cadmium toxicity in rats with special emphasis on the functional and histopathological changes. Toxicol. Rep. 2019, 6, 625–631. [Google Scholar] [CrossRef] [PubMed]
- Pellavio, G.; Rossino, G.; Gastaldi, G.; Rossi, D.; Linciano, P.; Collina, S.; Laforenza, U. Sigma-1 receptor agonists acting on aquaporin-mediated H2O2 permeability: New tools for counteracting oxidative stress. Int. J. Mol. Sci. 2021, 22, 9790. [Google Scholar] [CrossRef]
- Yin, Q.; Liu, C.; Bao, H.; Li, S.; Huang, Z.; Gu, D.; Xiong, L.; Miao, L. Estimation of gingival crevicular fluid oxidative stress markers in school-aged children and teenagers with insufficient sleep. BMC Oral Health 2022, 22, 616. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Oberley, L.W.; Li, Y. A simple method for clinical assay of superoxide dismutase. Clin. Chem. 1988, 34, 497–500. [Google Scholar] [CrossRef] [PubMed]
- Lian, H.; Lin, K.; Yang, C.; Cai, P. Generation and propagation of yeast prion [URE3] are elevated under electromagnetic field. Cell Stress Chaperones 2018, 23, 581–594. [Google Scholar] [CrossRef] [PubMed]
- Xiang, L.; Li, Y.; Wang, Z.; Liu, B.; Zhao, H.; Li, H.; Cai, Q.; Mo, C.; Li, Q. Bioaccumulation and phytotoxicity and human health risk from microcystin-LR under various treatments: A pot study. Toxins 2020, 12, 523. [Google Scholar] [CrossRef]
- Li, X.; Xu, Y.; Liu, J.; Yu, X.; Zhang, W.; You, C. Biological activities and gene expression of detoxifying enzymes in Tribolium castaneum induced by Moutan cortex essential oil. J. Toxicol. Environ. Health A 2022, 85, 591–602. [Google Scholar] [CrossRef]
- Xia, X.; Sun, B.; Gurr, G.M.; Vasseur, L.; Xue, M.; You, M. Gut microbiota mediate insecticide resistance in the diamondback moth, Plutella xylostella (L.). Front. Microbiol. 2018, 9, 25. [Google Scholar] [CrossRef] [PubMed]
- Kohl, K.D.; Dearing, M.D. The woodrat gut microbiota as an experimental system for understanding microbial metabolism of dietary toxins. Front. Microbiol. 2016, 7, 1165. [Google Scholar] [CrossRef] [PubMed]
- Mason, C.J.; Jones, A.G.; Felton, G.W. Co-option of microbial associates by insects and their impact on plant–folivore interactions. Plant. Cell Environ. 2019, 42, 1078–1086. [Google Scholar] [CrossRef] [PubMed]
- Broderick, N.A.; Raffa, K.F.; Handelsman, J. Midgut bacteria required for Bacillus thuringiensis insecticidal activity. Proc. Natl. Acad. Sci. USA 2006, 103, 15196–15199. [Google Scholar] [CrossRef]
- Kontsedalov, S.; Zchori-Fein, E.; Chiel, E.; Gottlieb, Y.; Inbar, M.; Ghanim, M. The presence of Rickettsia is associated with increased susceptibility of Bemisia tabaci (Homoptera: Aleyrodidae) to insecticides. Pest Manag. Sci. 2008, 64, 789–792. [Google Scholar] [CrossRef]
- Tiwari, S.; Pelz-Stelinski, K.; Mann, R.S.; Stelinski, L.L. Glutathione transferase and cytochrome P450 (general oxidase) activity levels in Candidatus Liberibacter asiaticus-infected and uninfected Asian citrus psyllid (Hemiptera: Psyllidae). Ann. Entomol. Soc. Am. 2011, 104, 297–305. [Google Scholar] [CrossRef]
- Daisley, B.A.; Trinder, M.; McDowell, T.W.; Collins, S.L.; Sumarah, M.W.; Reid, G. Microbiota-mediated modulation of organophosphate insecticide toxicity by species-dependent interactions with Lactobacilli in a Drosophila melanogaster insect model. Appl. Environ. Microbiol. 2018, 84, e02820-17. [Google Scholar] [CrossRef] [PubMed]
- Miao, Z.; Tu, Y.; Guo, P.; He, W.; Jing, T.; Wang, J.; Wei, D. Antioxidant enzymes and heat shock protein genes from Liposcelis bostrychophila are involved in stress defense upon heat shock. Insects 2020, 11, 839. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Li, L.; Zhang, F.; Wang, Y. Transcriptional inhibition of the catalase gene in phosphine-induced oxidative stress in Drosophila melanogaster. Pestic. Biochem. Physiol. 2015, 124, 1–7. [Google Scholar] [CrossRef]
- Bae, Y.S.; Choi, M.K.; Lee, W.J. Dual oxidase in mucosal immunity and host–microbe homeostasis. Trends Immunol. 2010, 31, 278–287. [Google Scholar] [CrossRef] [PubMed]
- Dolezal, T.; Krejcova, G.; Bajgar, A.; Nedbalova, P.; Strasser, P. Molecular regulations of metabolism during immune response in insects. Insect Biochem. Mol. Biol. 2019, 109, 31–42. [Google Scholar] [CrossRef]
- Duron, O.; Labbe, P.; Berticat, C.; Rousset, F.; Guillot, S.; Raymond, M.; Weill, M. High Wolbachia density correlates with cost of infection for insecticide resistant Culex pipiens mosquitoes. Evolution 2006, 60, 303–314. [Google Scholar] [CrossRef]
- Agnew, P.; Berticat, C.; Bedhomme, S.; Sidobre, C.; Michalakis, Y. Parasitism increases and decreases the costs of insecticide resistance in mosquitoes. Evolution 2004, 58, 579–586. [Google Scholar] [CrossRef] [PubMed]
- Slos, S.; Stoks, R. Predation risk induces stress proteins and reduces antioxidant defense. Funct. Ecol. 2008, 22, 637–642. [Google Scholar] [CrossRef]
- Yadav, S.K.; Srivastava, C.; Sabtharishi, S. Phosphine resistance and antioxidant enzyme activity in Trogoderma granarium Everts. J. Stored Prod. Res. 2020, 87, 101636. [Google Scholar] [CrossRef]
- Arismendi, N.L.; Reyes, M.; Miller, S.A.; Wijeratne, A.J.; Carrillo, R. Infection of ‘Candidatus Phytoplasma ulmi’ reduces the protein content and alters the activity of detoxifying enzymes in Amplicephalus curtulus. Entomol. Exp. Appl. 2015, 157, 334–345. [Google Scholar] [CrossRef]
- Wu, Y.; Huang, J.; Deng, M.; Jin, Y.; Yang, H.; Liu, Y.; Cao, Q.; Mennigen, J.A.; Tu, W. Acute exposure to environmentally relevant concentrations of Chinese PFOS alternative F-53B induces oxidative stress in early developing zebrafish. Chemosphere 2019, 235, 945–951. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Guo, X.; Li, Y.; Zhang, J.; Fu, Y. miR-34-5p, encoded by Spodoptera frugiperda, participates in anti-baculovirus by regulating innate immunity in the insect host. Int. J. Biol. Macromol. 2022, 222, 2190–2199. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Mean ± SE relative gut load of Brevibacterium sp., Enterococcus sp., Microbacterium sp., Micrococcus sp., and Staphylococcus sp. in adult Tribolium castaneum with different treatments of gut bacteria (n = 5). Germ-free means that gut bacteria are successfully depleted from beetles via antibiotic treatment. Gnotobiotic means that a given gut bacterium is successfully inoculated into germ-free beetles. The same letters indicate no significant difference among the means in the same cluster (one-way ANOVA and Tukey HSD test, p > 0.05).
Figure 1.
Mean ± SE relative gut load of Brevibacterium sp., Enterococcus sp., Microbacterium sp., Micrococcus sp., and Staphylococcus sp. in adult Tribolium castaneum with different treatments of gut bacteria (n = 5). Germ-free means that gut bacteria are successfully depleted from beetles via antibiotic treatment. Gnotobiotic means that a given gut bacterium is successfully inoculated into germ-free beetles. The same letters indicate no significant difference among the means in the same cluster (one-way ANOVA and Tukey HSD test, p > 0.05).

Figure 2.
Mean ± SE mortality of adult Tribolium castaneum after exposure to different concentrations of phosphine for 20 h (n = 5). Germ-free means that gut bacteria are successfully depleted from beetles via antibiotic treatment. Gnotobiotic means that a given gut bacterium is successfully inoculated into germ-free beetles. The same letters indicate no significant difference among the means in the same cluster (one-way ANOVA and Tukey HSD test, p > 0.05).
Figure 2.
Mean ± SE mortality of adult Tribolium castaneum after exposure to different concentrations of phosphine for 20 h (n = 5). Germ-free means that gut bacteria are successfully depleted from beetles via antibiotic treatment. Gnotobiotic means that a given gut bacterium is successfully inoculated into germ-free beetles. The same letters indicate no significant difference among the means in the same cluster (one-way ANOVA and Tukey HSD test, p > 0.05).
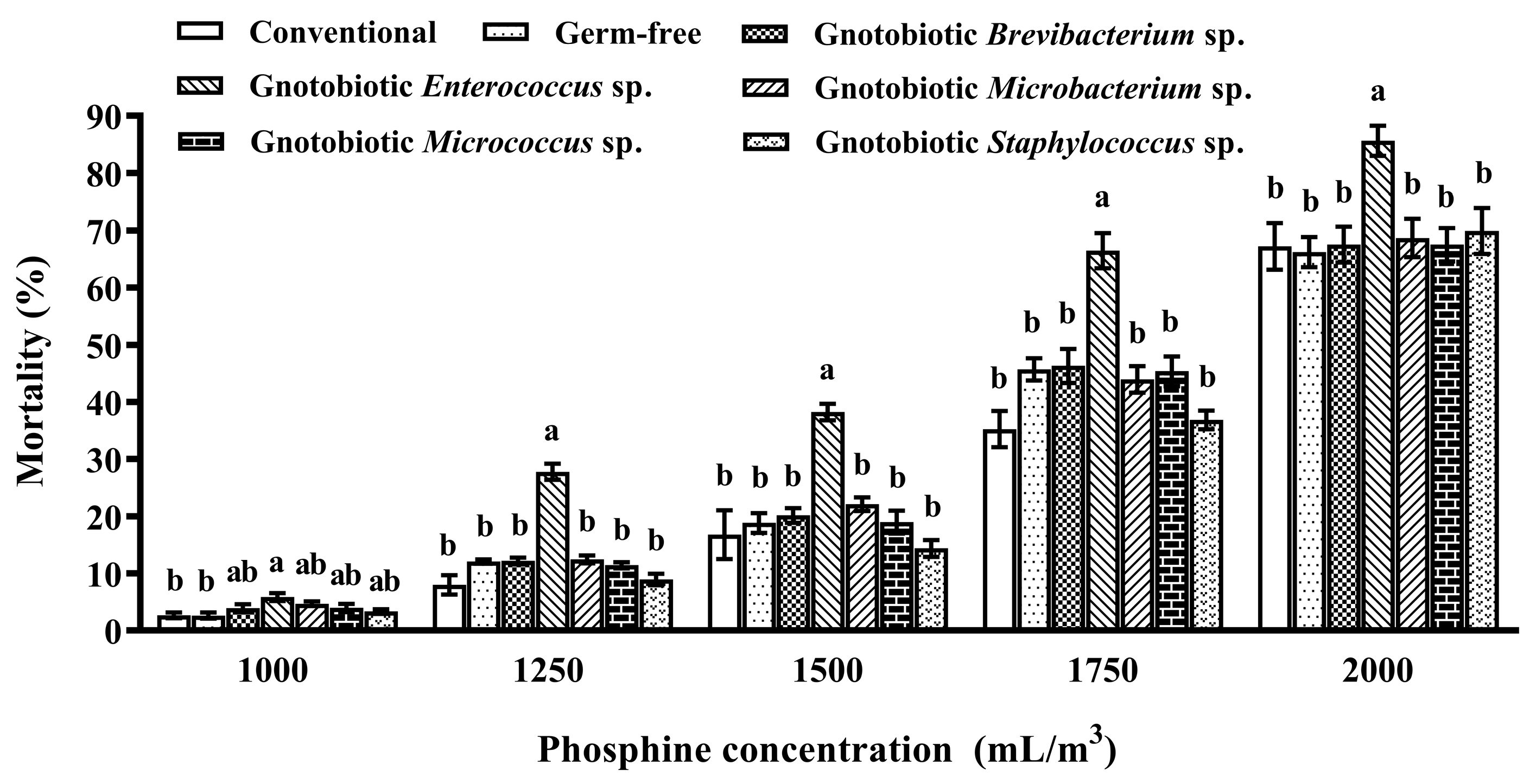
Figure 3.
Mean ± SE number of eggs laid per female after exposure of adult Tribolium castaneum to 1250 mL/m3 phosphine for 20 h (n = 5). Germ-free means that Enterococcus sp. is successfully depleted from beetles via antibiotic treatment. Gnotobiotic means that Enterococcus sp. is successfully inoculated into germ-free beetles. The same letters indicate no significant difference among different treatments (one-way ANOVA and Tukey HSD test, p > 0.05).
Figure 3.
Mean ± SE number of eggs laid per female after exposure of adult Tribolium castaneum to 1250 mL/m3 phosphine for 20 h (n = 5). Germ-free means that Enterococcus sp. is successfully depleted from beetles via antibiotic treatment. Gnotobiotic means that Enterococcus sp. is successfully inoculated into germ-free beetles. The same letters indicate no significant difference among different treatments (one-way ANOVA and Tukey HSD test, p > 0.05).
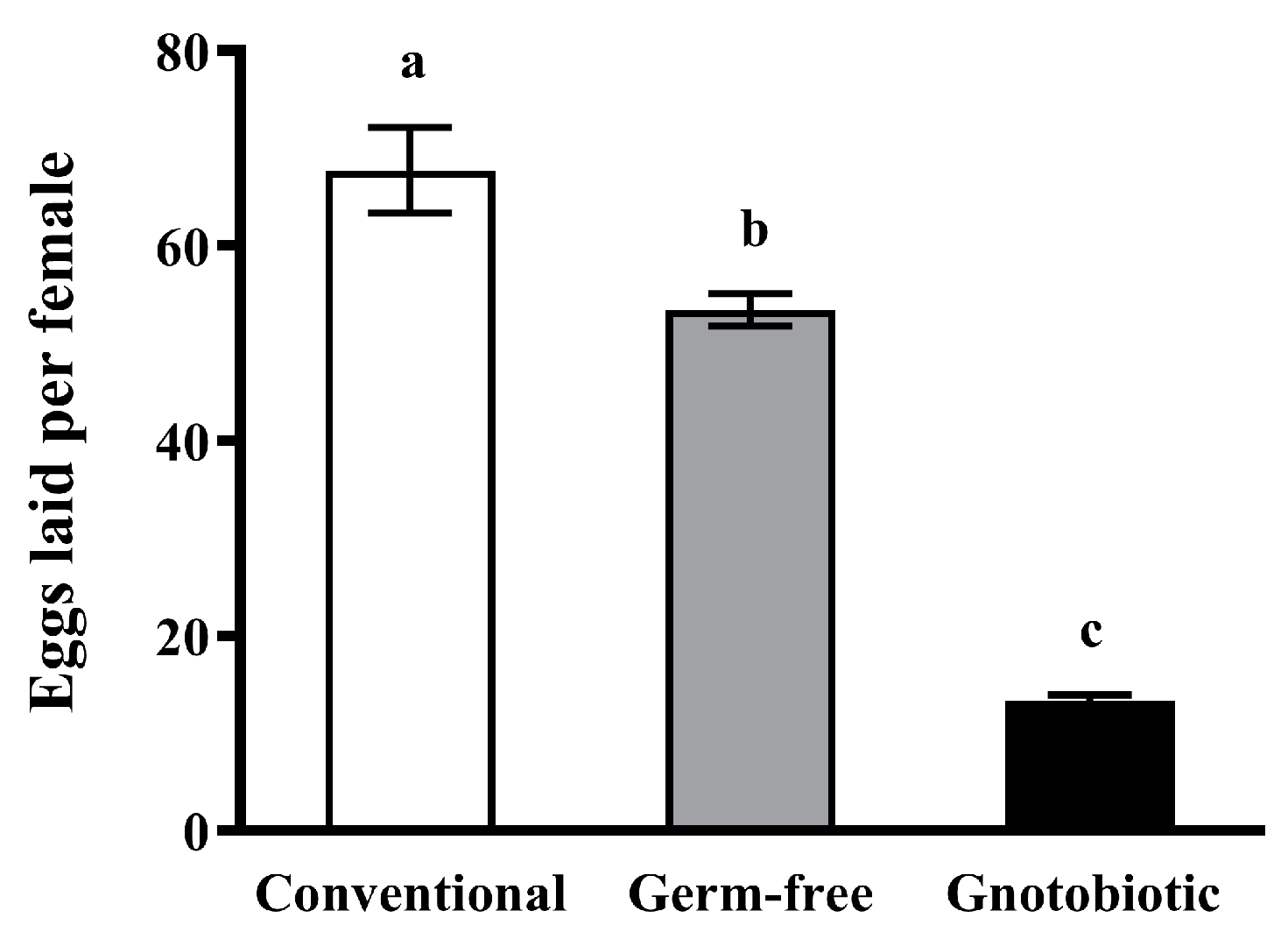
Figure 4.
Mean ± SE level of malondialdehyde (MDA, A) and hydrogen peroxide (H2O2, B) in adult Tribolium castaneum after exposure to 1250 mL/m3 phosphine for different periods (n = 5). Germ-free means that Enterococcus sp. is successfully depleted from beetles via antibiotic treatment. Gnotobiotic means that Enterococcus sp. is successfully inoculated into germ-free beetles. The same uppercase letters indicate no significant difference among the means with the same treatment of gut bacteria across different exposure periods, and the same lowercase letters indicate no significant difference among the means of conventional, germ-free, and gnotobiotic beetles within the same exposure period (one-way ANOVA and Tukey HSD test, p > 0.05).
Figure 4.
Mean ± SE level of malondialdehyde (MDA, A) and hydrogen peroxide (H2O2, B) in adult Tribolium castaneum after exposure to 1250 mL/m3 phosphine for different periods (n = 5). Germ-free means that Enterococcus sp. is successfully depleted from beetles via antibiotic treatment. Gnotobiotic means that Enterococcus sp. is successfully inoculated into germ-free beetles. The same uppercase letters indicate no significant difference among the means with the same treatment of gut bacteria across different exposure periods, and the same lowercase letters indicate no significant difference among the means of conventional, germ-free, and gnotobiotic beetles within the same exposure period (one-way ANOVA and Tukey HSD test, p > 0.05).
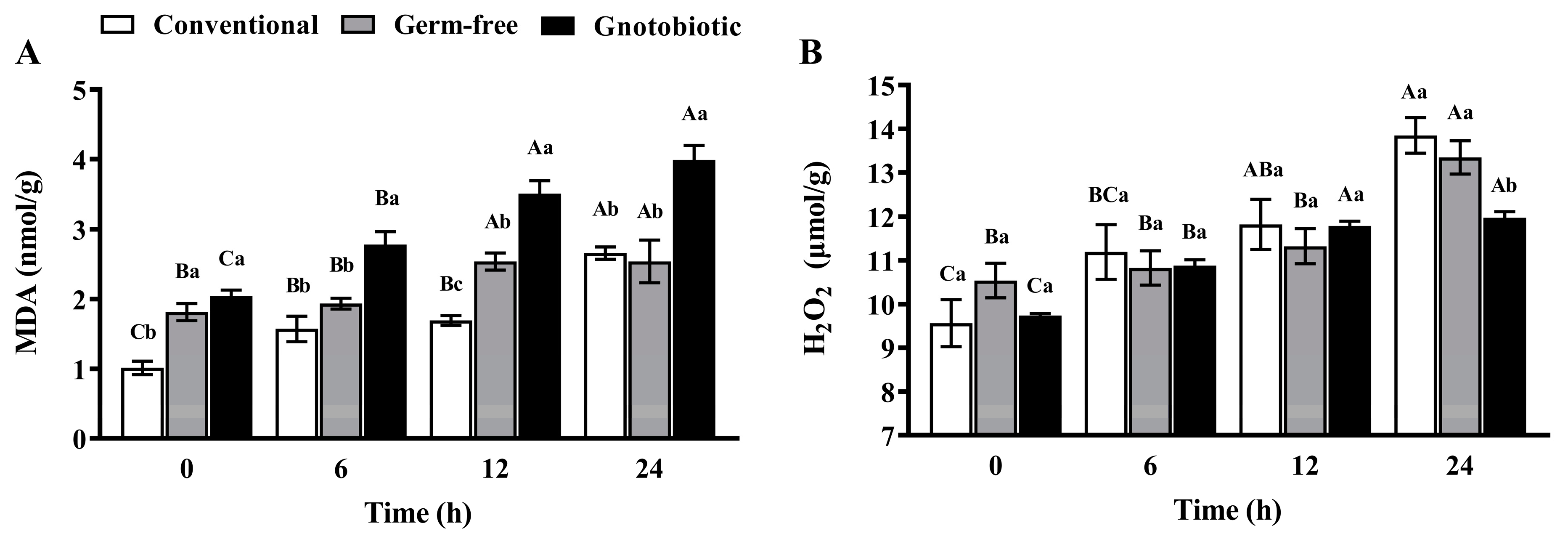
Figure 5.
Mean ± SE activity of superoxide dismutase (SOD, A), catalase (CAT, B), and peroxidase (POD, C) in adult Tribolium castaneum after exposure to 1250 mL/m3 phosphine for different periods (n = 5). Germ-free means that Enterococcus sp. is successfully depleted from beetles via antibiotic treatment. Gnotobiotic means that Enterococcus sp. is successfully inoculated into germ-free beetles. The same uppercase letters indicate no significant difference among the means with the same treatment of gut bacteria across different exposure periods, and the same lowercase letters indicate no significant difference among the means of conventional, germ-free, and gnotobiotic beetles within the same exposure period (one-way ANOVA and Tukey HSD test, p > 0.05).
Figure 5.
Mean ± SE activity of superoxide dismutase (SOD, A), catalase (CAT, B), and peroxidase (POD, C) in adult Tribolium castaneum after exposure to 1250 mL/m3 phosphine for different periods (n = 5). Germ-free means that Enterococcus sp. is successfully depleted from beetles via antibiotic treatment. Gnotobiotic means that Enterococcus sp. is successfully inoculated into germ-free beetles. The same uppercase letters indicate no significant difference among the means with the same treatment of gut bacteria across different exposure periods, and the same lowercase letters indicate no significant difference among the means of conventional, germ-free, and gnotobiotic beetles within the same exposure period (one-way ANOVA and Tukey HSD test, p > 0.05).
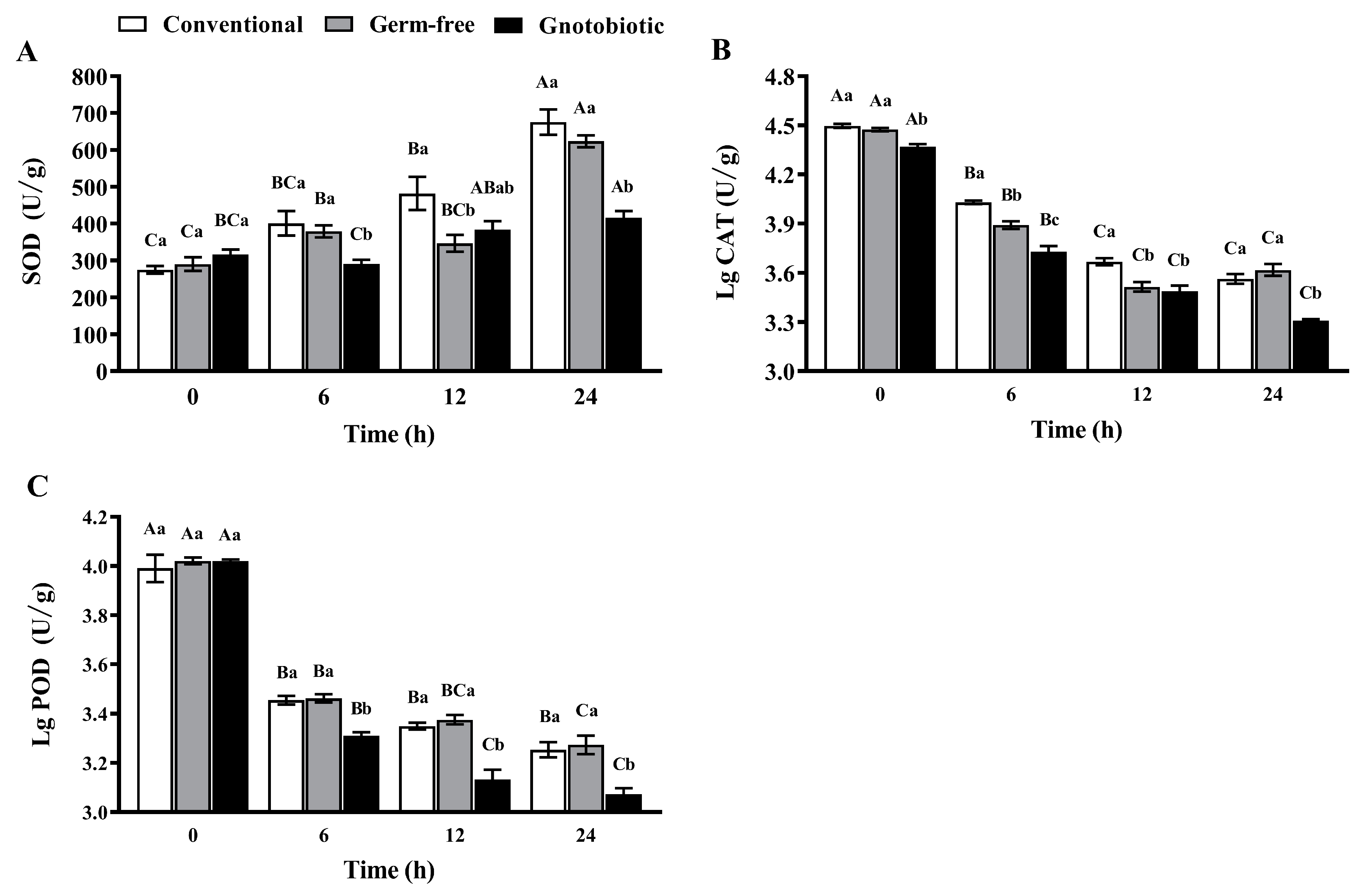
Figure 6.
Mean ± SE relative expression levels of SOD1a (A), SOD1b (B), SOD1c (C), SOD2 (D), CAT3 (E), and PRDX6 (F) in adult Tribolium castaneum after exposure to 1250 mL/m3 phosphine for different periods (n = 5). Germ-free means that Enterococcus sp. is successfully depleted from beetles via antibiotic treatment. Gnotobiotic means that Enterococcus sp. is successfully inoculated into germ-free beetles. The same uppercase letters indicate no significant difference among the means with the same treatment of gut bacteria across different exposure periods, and the same lowercase letters indicate no significant difference among the means of conventional, germ-free, and gnotobiotic beetles within the same exposure period (one-way ANOVA and Tukey HSD test, p > 0.05).
Figure 6.
Mean ± SE relative expression levels of SOD1a (A), SOD1b (B), SOD1c (C), SOD2 (D), CAT3 (E), and PRDX6 (F) in adult Tribolium castaneum after exposure to 1250 mL/m3 phosphine for different periods (n = 5). Germ-free means that Enterococcus sp. is successfully depleted from beetles via antibiotic treatment. Gnotobiotic means that Enterococcus sp. is successfully inoculated into germ-free beetles. The same uppercase letters indicate no significant difference among the means with the same treatment of gut bacteria across different exposure periods, and the same lowercase letters indicate no significant difference among the means of conventional, germ-free, and gnotobiotic beetles within the same exposure period (one-way ANOVA and Tukey HSD test, p > 0.05).
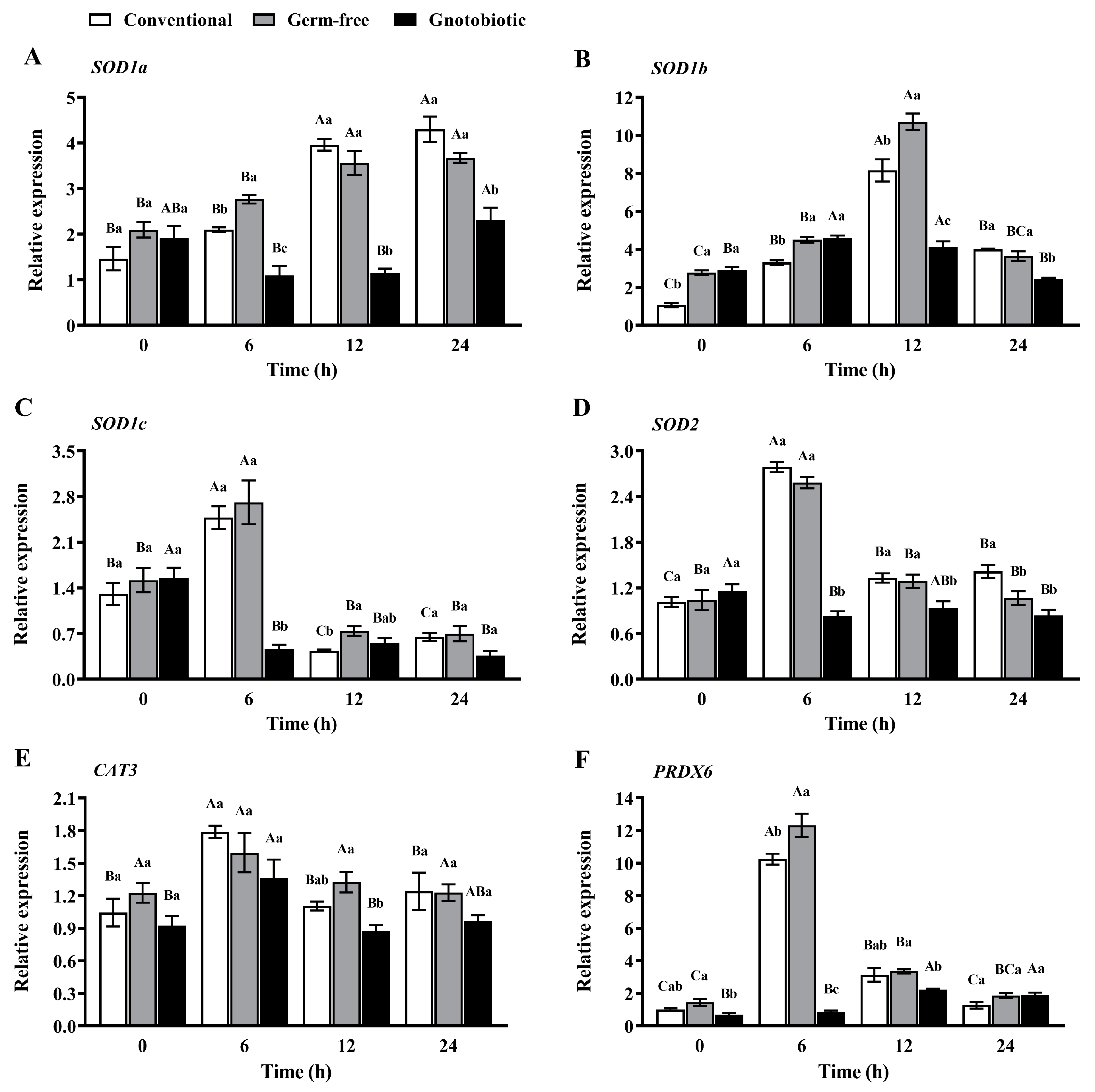
Figure 7.
Mean ± SE relative gut load of Enterococcus sp. (A) and relative expression levels of PLCβ (B), Atf2 (C), DUOX (D), IMD (E), and Toll (F) in adult Tribolium castaneum after inoculation of Enterococcus sp. into germ-free beetles for different periods (n = 5). Gnotobiotic means that Enterococcus sp. is successfully inoculated into germ-free beetles. The same uppercase letters indicate no significant difference among the means of conventional and gnotobiotic beetles within the same period (one-way ANOVA, p > 0.05), and the same lowercase letters indicate no significant difference among the means with the same gut bacteria treatment across different periods (one-way ANOVA and Tukey HSD test, p > 0.05).
Figure 7.
Mean ± SE relative gut load of Enterococcus sp. (A) and relative expression levels of PLCβ (B), Atf2 (C), DUOX (D), IMD (E), and Toll (F) in adult Tribolium castaneum after inoculation of Enterococcus sp. into germ-free beetles for different periods (n = 5). Gnotobiotic means that Enterococcus sp. is successfully inoculated into germ-free beetles. The same uppercase letters indicate no significant difference among the means of conventional and gnotobiotic beetles within the same period (one-way ANOVA, p > 0.05), and the same lowercase letters indicate no significant difference among the means with the same gut bacteria treatment across different periods (one-way ANOVA and Tukey HSD test, p > 0.05).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Wang, Z.; Zhang, S.; Liu, Z.; Chang, Z.; Hu, H. Gut Bacteria Promote Phosphine Susceptibility of Tribolium castaneum by Aggravating Oxidative Stress and Fitness Costs. Insects 2023, 14, 815. https://doi.org/10.3390/insects14100815
AMA Style
Wang Z, Zhang S, Liu Z, Chang Z, Hu H. Gut Bacteria Promote Phosphine Susceptibility of Tribolium castaneum by Aggravating Oxidative Stress and Fitness Costs. Insects. 2023; 14(10):815. https://doi.org/10.3390/insects14100815
Chicago/Turabian StyleWang, Zhengyan, Shan Zhang, Zhiyuan Liu, Zhenzhen Chang, and Haisheng Hu. 2023. "Gut Bacteria Promote Phosphine Susceptibility of Tribolium castaneum by Aggravating Oxidative Stress and Fitness Costs" Insects 14, no. 10: 815. https://doi.org/10.3390/insects14100815
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.