
The Impact of Climate on the Energetics of Overwintering Paper Wasp Gynes (Polistes dominula and Polistes gallicus)
 , , and
, , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Species, Locations, and Climates
2.2. Temperature Measurement
2.3. Energetic Expenditure Calculations
2.4. Data Analysis and Statistics
3. Results
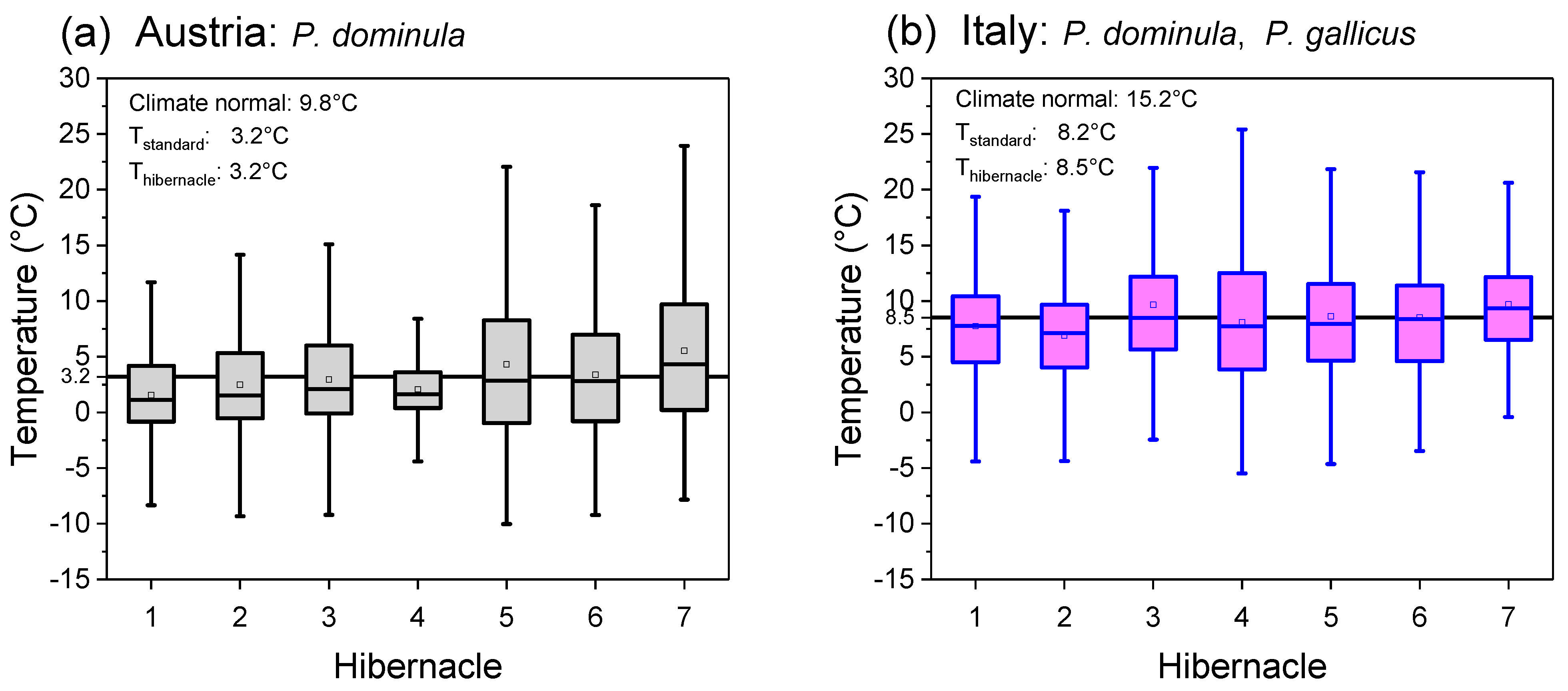
3.1. Hibernacle Micro- and Macroclimate
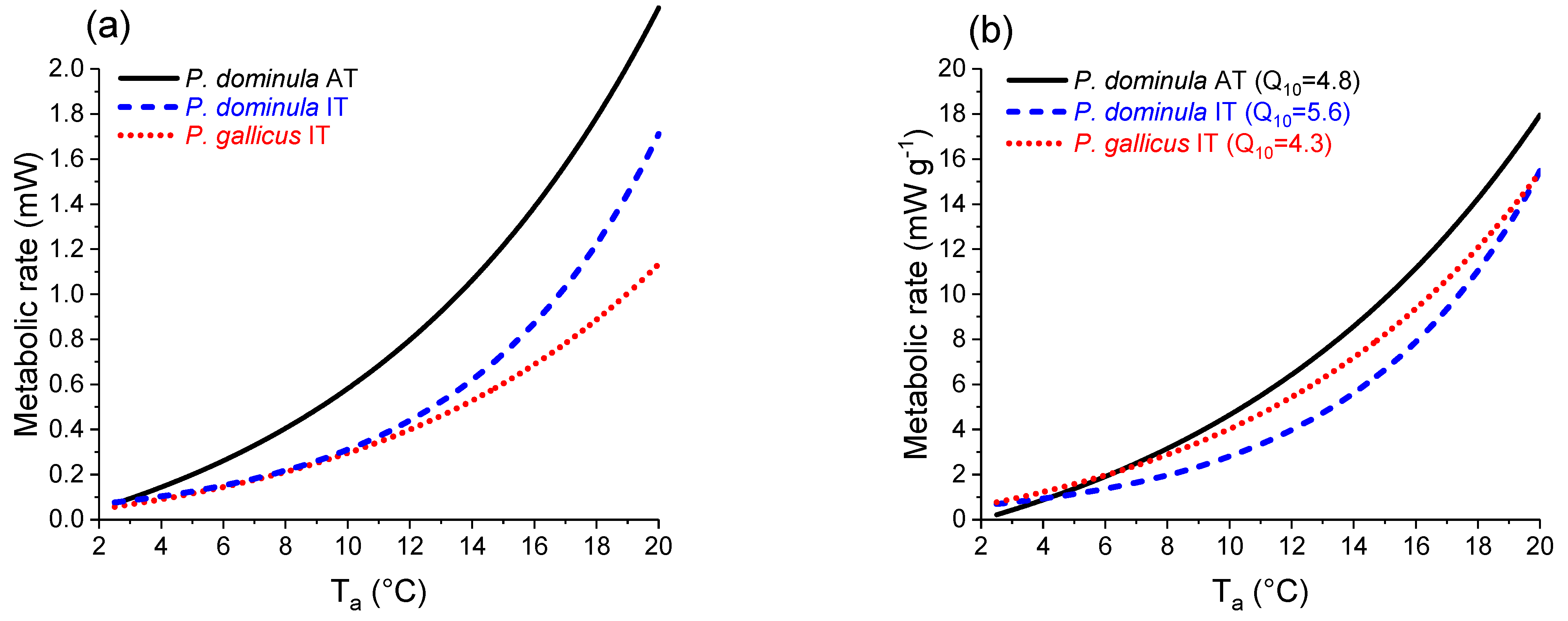
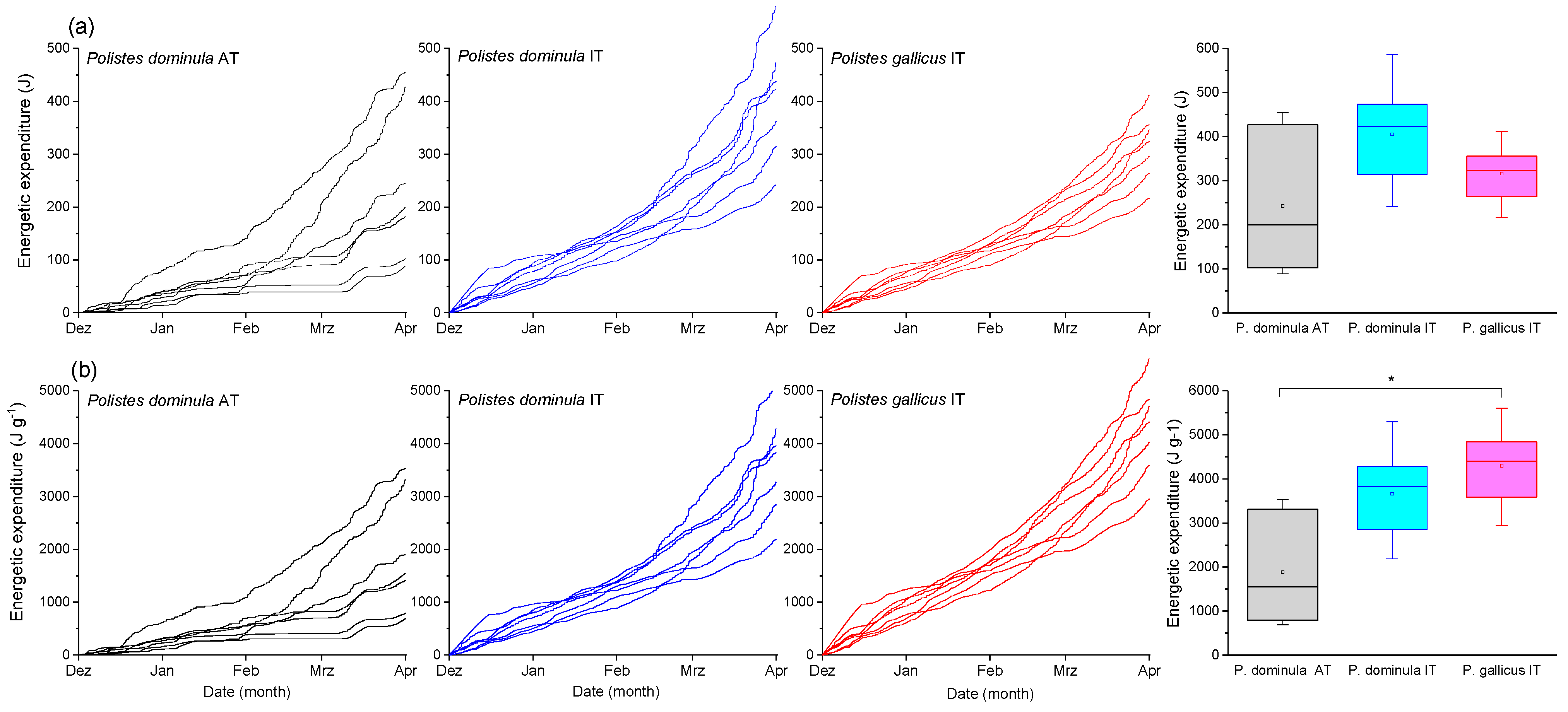
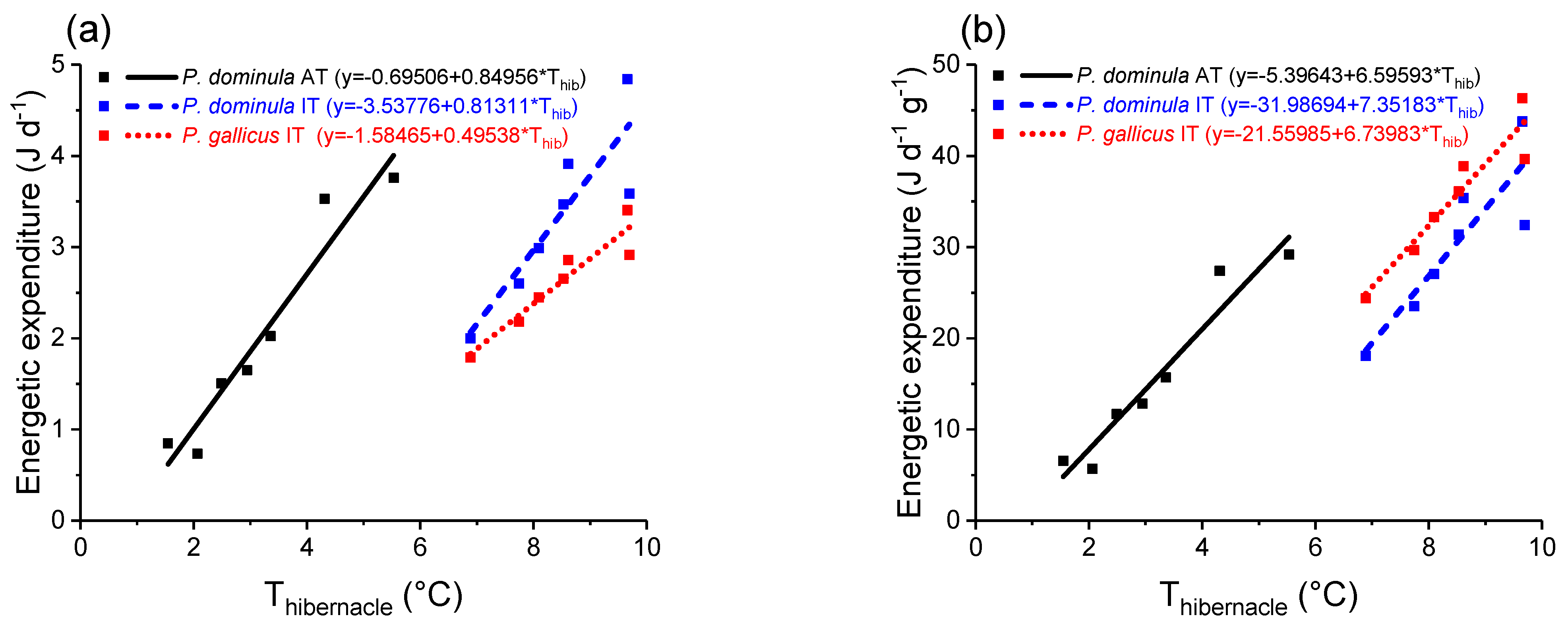
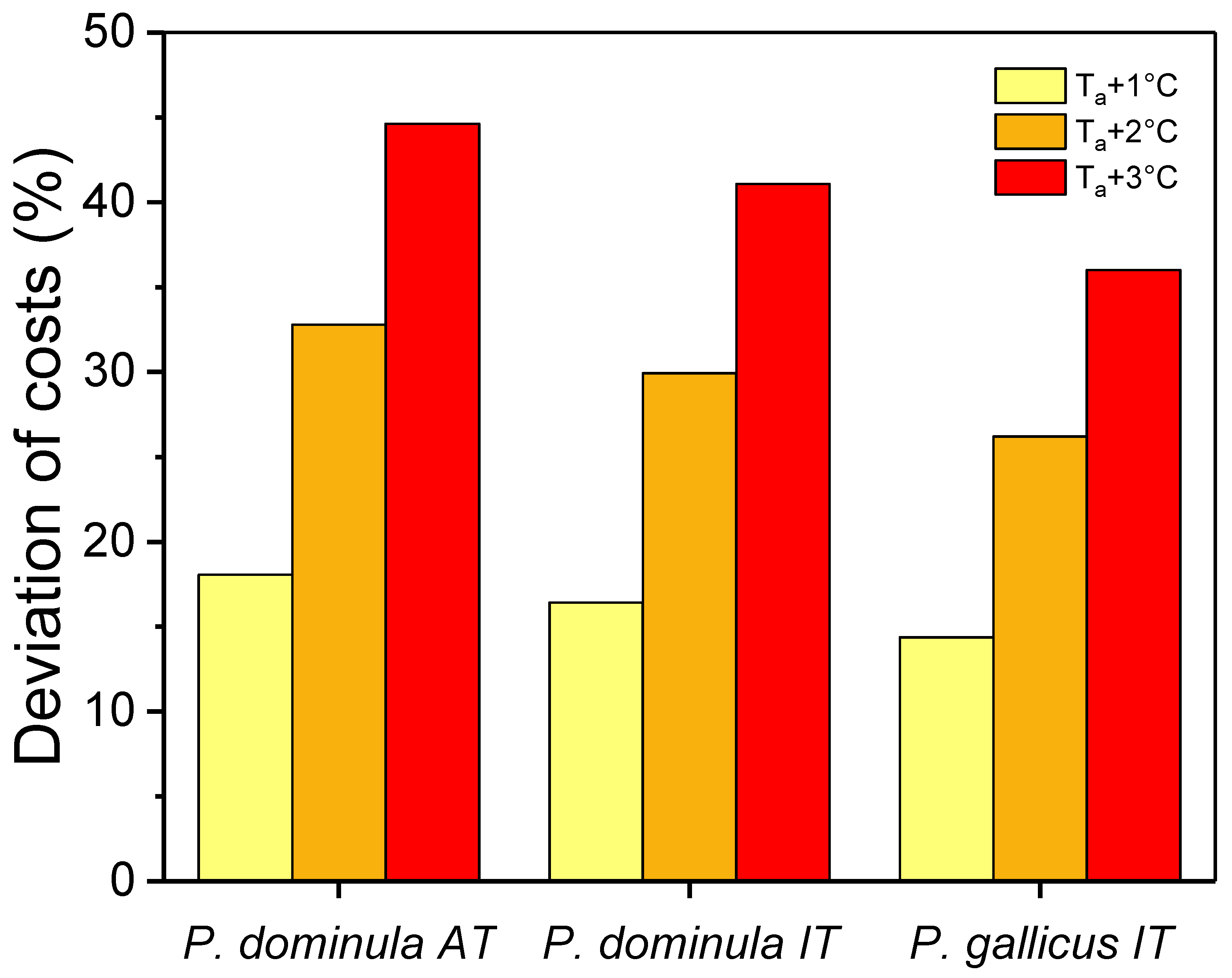
3.2. Energetic Costs of Overwintering
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Koštál, V. Eco-physiological phases of insect diapause. J. Insect Physiol. 2006, 52, 113–127. [Google Scholar] [CrossRef]
- Hahn, D.A.; Denlinger, D.L. Energetics of Insect Diapause. Annu. Rev. Entomol. 2011, 56, 103–121. [Google Scholar] [CrossRef]
- Kovac, H.; Käfer, H.; Petrocelli, I.; Stabentheiner, A. The respiratory metabolism of overwintering paper wasp gynes (Polistes dominula and Polistes gallicus). Physiol. Entomol. 2022, 47, 62–71. [Google Scholar] [CrossRef]
- Popović, Ž.D.; Maier, V.; Avramov, M.; Uzelac, I.; Gošić-Dondo, S.; Blagojević, D.; Koštál, V. Acclimations to Cold and Warm Conditions Differently Affect the Energy Metabolism of Diapausing Larvae of the European Corn Borer Ostrinia nubilalis (Hbn.). Front. Physiol. 2021, 12, 768593. [Google Scholar] [CrossRef] [PubMed]
- Käfer, H.; Kovac, H.; Oswald, B.; Stabentheiner, A. Respiration and metabolism of the resting European paper wasp (Polistes dominulus). J. Comp. Physiol. B 2015, 185, 647–658. [Google Scholar] [CrossRef] [PubMed]
- Kovac, H.; Käfer, H.; Petrocelli, I.; Stabentheiner, A. Comparison of thermal traits of Polistes dominula and Polistes gallicus, two European paper wasps with strongly differing distribution ranges. J. Comp. Physiol. B 2017, 187, 277–290. [Google Scholar] [CrossRef] [PubMed]
- Sinclair, B.J. Linking energetics and overwintering in temperate insects. J. Therm. Biol. 2015, 54, 5–11. [Google Scholar] [CrossRef] [PubMed]
- Williams, C.M.; Hellmann, J.; Sinclair, B.J. Lepidopteran species differ in susceptibility to winter warming. Clim. Res. 2012, 53, 119–130. [Google Scholar] [CrossRef]
- Sgolastra, F.; Kemp, W.P.; Buckner, J.S.; Pitts-Singer, T.L.; Maini, S.; Bosch, J. The long summer: Pre-wintering temperatures affect metabolic expenditure and winter survival in a solitary bee. J. Insect Physiol. 2011, 57, 1651–1659. [Google Scholar] [CrossRef]
- Williams, C.M.; Marshall, K.E.; MacMillan, H.A.; Dzurisin, J.D.K.; Hellmann, J.J.; Sinclair, B.J. Thermal variability increases the impact of autumnal warming and drives metabolic depression in an overwintering butterfly. PLoS ONE 2012, 7, e34470. [Google Scholar] [CrossRef]
- Koštál, V.; Renault, D.; Rozsypal, J. Seasonal changes of free amino acids and thermal hysteresis in overwintering heteropteran insect, Pyrrhocoris apterus. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2011, 160, 245–251. [Google Scholar] [CrossRef] [PubMed]
- Williams, J.B.; Shorthouse, J.D.; Lee, R.E. Deleterious effects of mild simulated overwintering temperatures on survival and potential fecundity of rose-galling Diplolepis wasps (Hymenoptera: Cynipidae). J. Exp. Zool. Part A Comp. Exp. Biol. 2003, 298, 23–31. [Google Scholar] [CrossRef] [PubMed]
- Williams, C.M.; Watanabe, M.; Guarracino, M.R.; Ferraro, M.B.; Edison, A.S.; Morgan, T.J.; Boroujerdi, A.F.B.; Hahn, D.A. Cold adaptation shapes the robustness of metabolic networks in Drosophila melanogaster. Evolution 2014, 68, 3505–3523. [Google Scholar] [CrossRef] [PubMed]
- Bosch, J.; Kemp, W.P. Effect of Wintering Duration and Temperature on Survival and Emergence Time in Males of the Orchard Pollinator Osmia lignaria (Hymenoptera: Megachilidae). Environ. Entomol. 2003, 32, 711–716. [Google Scholar] [CrossRef]
- Bosch, J.; Kemp, W.P. Effect of pre-wintering and wintering temperature regimes on weight loss, survival, and emergence time in the mason bee Osmia cornuta (Hymenoptera: Megachilidae). Apidologie 2004, 35, 469–479. [Google Scholar] [CrossRef]
- Irwin, J.T.; Lee, J.R.E. Cold winter microenvironments conserve energy and improve overwintering survival and potential fecundity of the goldenrod gall fly, Eurosta solidaginis. Oikos 2003, 100, 71–78. [Google Scholar] [CrossRef]
- Klockmann, M.; Fischer, K. Strong reduction in diapause survival under warm and humid overwintering conditions in a temperate-zone butterfly. Popul. Ecol. 2019, 61, 150–159. [Google Scholar] [CrossRef]
- Mercader, R.J.; Scriber, J.M. Asymmetrical thermal constraints on the parapatric species boundaries of two widespread generalist butterflies. Ecol. Entomol. 2008, 33, 537–545. [Google Scholar] [CrossRef]
- Sorvari, J.; Haatanen, M.-K.; Vesterlund, S.-R. Combined effects of overwintering temperature and habitat degradation on the survival of boreal wood ant. J. Insect Conserv. 2011, 15, 727–731. [Google Scholar] [CrossRef]
- Rozsypal, J.; Moos, M.; Rudolf, I.; Košťál, V. Do energy reserves and cold hardiness limit winter survival of Culex pipiens? Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2021, 255, 110912. [Google Scholar] [CrossRef]
- Williams, C.M.; Chick, W.D.; Sinclair, B.J. A cross-seasonal perspective on local adaptation: Metabolic plasticity mediates responses to winter in a thermal-generalist moth. Funct. Ecol. 2015, 29, 549–561. [Google Scholar] [CrossRef]
- Silbernagl. Taschenatlas der Physiologie; Silbernagl: Castelrotto, Italy, 1991. [Google Scholar]
- Masson-Delmotte, V.; Zhai, P.; Pirani, A.; Connors, S.L.; Péan, C.; Berger, S.; Caud, N.; Chen, Y.; Goldfarb, L.; Gomis, M.I.; et al. Climate Change 2021: The Physical Science Basis: Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK, 2021; 2391p. [Google Scholar] [CrossRef]
- Potter, K.A.; Arthur Woods, H.; Pincebourde, S. Microclimatic challenges in global change biology. Glob. Change Biol. 2013, 19, 2932–2939. [Google Scholar] [CrossRef] [PubMed]
- Kearney, M.R.; Isaac, A.P.; Porter, W.P. microclim: Global estimates of hourly microclimate based on long-term monthly climate averages. Sci. Data 2014, 1, 140006. [Google Scholar] [CrossRef] [PubMed]
- Pincebourde, S.; Casas, J. Narrow safety margin in the phyllosphere during thermal extremes. Proc. Natl. Acad. Sci. USA 2019, 116, 5588–5596. [Google Scholar] [CrossRef] [PubMed]
- Pincebourde, S.; Woods, H.A. There is plenty of room at the bottom: Microclimates drive insect vulnerability to climate change. Curr. Opin. Insect Sci. 2020, 41, 63–70. [Google Scholar] [CrossRef]
- Garcia, R.A.; Allen, J.L.; Clusella-Trullas, S. Rethinking the scale and formulation of indices assessing organism vulnerability to warmer habitats. Ecography 2019, 42, 1024–1036. [Google Scholar] [CrossRef]
- Maclean, I.M.D.; Duffy, J.P.; Haesen, S.; Govaert, S.; de Frenne, P.; Vanneste, T.; Lenoir, J.; Lembrechts, J.J.; Rhodes, M.W.; van Meerbeek, K. On the measurement of microclimate. Methods Ecol. Evol. 2021, 12, 1397–1410. [Google Scholar] [CrossRef]
- Woods, H.A.; Dillon, M.E.; Pincebourde, S. The roles of microclimatic diversity and of behavior in mediating the responses of ectotherms to climate change. J. Therm. Biol. 2015, 54, 86–97. [Google Scholar] [CrossRef]
- Pincebourde, S.; Suppo, C. The Vulnerability of Tropical Ectotherms to Warming Is Modulated by the Microclimatic Heterogeneity. Integr. Comp. Biol. 2016, 56, 85–97. [Google Scholar] [CrossRef]
- Kovac, H.; Käfer, H.; Petrocelli, I.; Amstrup, A.B.; Stabentheiner, A. Energetics of Paper Wasps (Polistes sp.) from Differing Climates during the Breeding Season. Insects 2022, 13, 800. [Google Scholar] [CrossRef]
- Dapporto, L.; Palagi, E. Wasps in the shadow: Looking at the pre-hibernating clusters of Polistes dominulus. Ann. Zool. Fenn. 2006, 43, 583–594. [Google Scholar]
- ZAMG-Zentralanstalt für Meteorologie und Geodynamik. Klimamittelwerte für den Zeitraum 1981–2010 von rund 160 österreichischen Messstationen. Available online: https://www.zamg.ac.at/cms/de/klima/informationsportal-klimawandel/daten-download/klimamittel (accessed on 20 December 2022).
- LaMMA Consorzio. Climatologia di Firenze 1981–2010. Available online: https://www.lamma.toscana.it/clima-e-energia/climatologia/clima-firenze (accessed on 20 December 2022).
- Gibo, D.L. Cold-hardiness in Fall and Winter Adults of the social Wasp Polistes fuscatus (Hymenoptera: Vespidae) in southern Ontario. Can. Entomol. 1976, 108, 801–806. [Google Scholar] [CrossRef]
- Gibo, D.L. Hibernation Sites and Temperature Tolerance of Two Species of Vespula and One Species of Polistes (Hymenoptera: Vespidae). J. N. Y. Entomol. Soc. 1972, 80, 105–108. [Google Scholar]
- Pratte, M. Relations antérieures et association de fondation chezPolistes gallicus L. Insectes Sociaux 1982, 29, 352–357. [Google Scholar] [CrossRef]
- Starks, P.T. Alternative reproductive tactics in the paper wasp Olistes dominulus with specific focus on the sit-and-wait tactic. Ann. Zool. Fenn. 2001, 38, 189–199. [Google Scholar]
- Gamboa, G.J.; Noble, M.A.; Thom, M.C.; Togal, J.L.; Srinivasan, R.; Murphy, B.D. The comparative biology of two sympatric paper wasps in Michigan, the native Polistes fuscatus and the invasive Polistes dominulus (Hymenoptera, Vespidae). Insectes Sociaux 2004, 51, 153–157. [Google Scholar] [CrossRef]
- Barnes, B.M.; Barger, J.L.; Seares, J.; Tacquard, P.C.; Zuercher, G.L. Overwintering in Yellowjacket Queens (Vespula vulgaris) and Green Stinkbugs (Elasmostethus interstinctus) in Subarctic Alaska. Physiol. Zool. 1996, 69, 1469–1480. [Google Scholar] [CrossRef]
- Popović, Ž.D.; Subotić, A.; Nikolić, T.V.; Radojičić, R.; Blagojević, D.P.; Grubor-Lajšić, G.; Koštál, V. Expression of stress-related genes in diapause of European corn borer (Ostrinia nubilalis Hbn.). Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2015, 186, 1–7. [Google Scholar] [CrossRef]
- Sgolastra, F.; Bosch, J.; Molowny-Horas, R.; Maini, S.; Kemp, W.P. Effect of temperature regime on diapause intensity in an adult-wintering Hymenopteran with obligate diapause. J. Insect Physiol. 2010, 56, 185–194. [Google Scholar] [CrossRef]
- Williams, C.M.; Henry, H.A.L.; Sinclair, B.J. Cold truths: How winter drives responses of terrestrial organisms to climate change. Biol. Rev. 2015, 90, 214–235. [Google Scholar] [CrossRef]
- Sinclair, B.J.; Klok, C.J.; Chown, S.L. Metabolism of the sub-Antarctic caterpillar Pringleophaga marioni during cooling, freezing and thawing. J. Exp. Biol. 2004, 207, 1287–1294. [Google Scholar] [CrossRef] [PubMed]
- Sømme, L.; Block, W. Adaptations to Alpine and Polar Environments in Insects and Other Terrestrial Arthropods. In Insects at Low Temperature; Lee, R.E., Denlinger, D.L., Eds.; Springer: Boston, MA, USA, 1991; pp. 318–359. ISBN 978-1-4757-0192-0. [Google Scholar]
- Hoffmann, A.A. Acclimation: Increasing survival at a cost. Trends Ecol. Evol. 1995, 10, 1–2. [Google Scholar] [CrossRef]
- Addo-Bediako, A.; Chown, S.L.; Gaston, K.J. Metabolic cold adaptation in insects: A large-scale perspective. Funct. Ecol. 2002, 16, 332–338. [Google Scholar] [CrossRef]
- Chown, S.L.; Nicolson, S.W. Insect Physiological Ecology: Mechanisms and Patterns; Oxford University Press: Oxford, NY, USA, 2004; ISBN 978-0198515494. [Google Scholar]
- Devlin, J.J.; Unfried, L.; Lecheta, M.C.; McCabe, E.A.; Gantz, J.D.; Kawarasaki, Y.; Elnitsky, M.A.; Hotaling, S.; Michel, A.P.; Convey, P.; et al. Simulated winter warming negatively impacts survival of Antarctica’s only endemic insect. Funct. Ecol. 2022, 36, 1949–1960. [Google Scholar] [CrossRef]
- Dillon, M.E.; Wang, G.; Huey, R.B. Global metabolic impacts of recent climate warming. Nature 2010, 467, 704–706. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kovac, H.; Käfer, H.; Petrocelli, I.; Amstrup, A.B.; Stabentheiner, A. The Impact of Climate on the Energetics of Overwintering Paper Wasp Gynes (Polistes dominula and Polistes gallicus). Insects 2023, 14, 849. https://doi.org/10.3390/insects14110849
Kovac H, Käfer H, Petrocelli I, Amstrup AB, Stabentheiner A. The Impact of Climate on the Energetics of Overwintering Paper Wasp Gynes (Polistes dominula and Polistes gallicus). Insects. 2023; 14(11):849. https://doi.org/10.3390/insects14110849
Chicago/Turabian StyleKovac, Helmut, Helmut Käfer, Iacopo Petrocelli, Astrid B. Amstrup, and Anton Stabentheiner. 2023. "The Impact of Climate on the Energetics of Overwintering Paper Wasp Gynes (Polistes dominula and Polistes gallicus)" Insects 14, no. 11: 849. https://doi.org/10.3390/insects14110849