
Accumulation of Fungal Pathogens Infecting the Invasive Spotted Lanternfly, Lycorma delicatula

1
Department of Entomology, Cornell University, Ithaca, NY 14853, USA
2
Research & Development, BioWorks Inc., Victor, NY 14564, USA
*
Author to whom correspondence should be addressed.
Insects 2023, 14(12), 912; https://doi.org/10.3390/insects14120912
Submission received: 30 October 2023
/
Revised: 21 November 2023
/
Accepted: 24 November 2023
/
Published: 27 November 2023
(This article belongs to the Section Insect Behavior and Pathology)
Abstract
:Simple Summary
Populations of the invasive spotted lanternfly in the eastern United States threaten vineyards and pose a public nuisance. Methods for control are being investigated, with an interest in mortality caused by insect-specific pathogens and naturally occurring predators. Two species of insect-pathogenic fungi caused extensive mortality in spotted lanternfly populations in fall 2018, and two additional fungal pathogens have been reported. Our extensive surveys, which emphasized a year with a summer drought, document that while Beauveria bassiana was most abundant, fifteen additional fungal pathogens killed spotted lanternflies. Although levels of infection for many pathogen species were low, infection was greatest in adults sampled in September and October, when spotted lanternfly reproduction occurs. Thus, we report an increased diversity of naturally occurring generalist fungal pathogens attacking this relatively new invasive planthopper.
Abstract
In the eastern United States, populations of the invasive spotted lanternfly, Lycorma delicatula, are abundant and spreading. Four species of naturally occurring entomopathogenic fungi have previously been reported as infecting these planthoppers, with two of these causing epizootics. Nymphal- and adult-stage lanternflies in Pennsylvania and New York were surveyed for entomopathogenic fungal infections from October 2021 to November 2023, and assays were conducted to confirm the pathogenicity of species that were potentially pathogenic. Beauveria bassiana was the most abundant pathogen, but we report an additional 15 previously unreported species of entomopathogenic fungi infecting spotted lanternflies, all in the order Hypocreales (Ascomycota). The next most common pathogens were Fusarium fujikuroi and Sarocladium strictum. While infection prevalence by species was often low, probably impacted to some extent by the summer drought in 2022, together these pathogens caused a total of 6.7% mortality. A significant trend was evident over time within a season, with low levels of infection among nymphs and higher infection levels in mid- and late-stage adults, the stages when mating and oviposition occur.
1. Introduction
The success of invasive species in their introduced ranges is often attributed to escape from natural enemies that provide control in native regions [1]. However, over time, natural enemies can adapt to using an invasive species through host shifts, evolutionary changes in resident enemy populations, or introductions of co-evolved natural enemies [2,3,4]. In the case of pathogens, the accumulation of pathogens attacking invasives has been hypothesized to potentially (1) lead to population decline of invasives or (2) create increases in pathogen populations that could spill over to impact co-occurring native host species [4]. With no saturation in numbers of invasive insect species predicted over time [5], the potential accumulation of entomopathogens attacking invasives, relative to both their biodiversity and their impact, requires investigation.
Entomopathogenic fungi are often found infecting invasive insect populations, especially when those invasive hosts are abundant [6]. In simplified ecosystems, epizootics in populations of one host species can occur, predominantly driven by one or two fungal species, and these are frequently well-known species in just a few genera, including species of Beauveria, Metarhizium, Akanthomyces (previously Lecanicillium), and Cordyceps [6]. However, there are more than 1000 species of entomopathogenic fungi in many different genera [7,8]. There are numerous reports of multiple entomopathogenic fungi infecting invasive insect populations [9,10,11,12,13,14,15,16,17,18], with many of these entomopathogens belonging to the order Hypocreales and considered native to the region where the pest is invading. These hypocrealean entomopathogens infecting invasives range from well-known pathogens, such as Beauveria bassiana, to opportunists requiring specific conditions (e.g., impaired host immunity) to infect insects [19].
The spotted lanternfly, Lycorma delicatula, is originally from Asia but was first discovered in Berks County, Pennsylvania, in 2014 and has since spread into 15 additional eastern US states [20]. This univoltine species feeds on woody plants, with nymphs feeding on leaf midribs, petioles, and twigs and adults feeding through bark. Once an area has been invaded, this planthopper increases in abundance, often in areas where its preferred Asian host tree, tree of heaven (Ailanthus altissima), occurs. Another preferred plant for L. delicatula is grape (Vitis spp.), and feeding has caused decreased yield in vineyards, requiring new control tactics [21]. Hemiptera, including planthoppers, are particularly susceptible to entomopathogenic fungi [7,22], and a species known worldwide, B. bassiana, has been reported to infect L. delicatula where it is native, in China [23].
In 2018, epizootics caused by entomopathogenic fungi were documented in abundant populations of L. delicatula in southeastern Pennsylvania [24]. One fungus causing these epizootics was the well-known insect pathogen B. bassiana, while the second was the lesser-known Batkoa major. Batkoa major is a poorly known obligate insect pathogen [25], while B. bassiana is a significant pathogen of insects, known to infect >700 insect species, but which can also live as a saprophyte and plant endophyte [26]. Late in December 2018 and in November 2020, two new species of entomopathogenic fungi were discovered infecting L. delicatula: Metarhizium pemphigi and Ophiocordyceps delicatula [27]. These discoveries were not previously known from North America and not previously described, respectively. Thus, by 2020, four entomopathogenic fungi were known to naturally infect L. delicatula in the US. In 2021, three more species were isolated from L. delicatula adults (see below), and the need for the present study became apparent. The goal of this study was to evaluate the diversity and abundance of entomopathogenic fungi naturally infecting L. delicatula of different stages in invasive infestations in the northeastern US. Fungal species were isolated and identified using molecular methods, and tests were conducted to confirm pathogenicity when needed.
2. Materials and Methods
2.1. Sample Collection
Throughout our studies, because L. delicatula is not considered well established in the area around Cornell University, all work with living L. delicatula was conducted under permit (USDA APHIS permit # P526P-21-02895) within the Cornell arthropod quarantine building, the Sarkaria Arthropod Research Laboratory (SARL). Any living L. delicatula collected at field sites were transported in triple containment to SARL for our studies.
This study was initiated with fungal entomopathogens isolated from 3 L. delicatula collected from two Pennsylvania sites in October and early November 2021. All three were adults; one was collected living and died while being reared on A. altissima (see below), and the other two were cadavers of recently-killed adults. In 2022, 4 field sites in eastern Pennsylvania were chosen where L. delicatula and A. altissima were well established in forested areas. Between 8 June and 2 November, 15 trips were made to these sites for collections (Table 1). Two additional trips were made to Angora Fruit Farm and Sinking Spring on 29 September and 12 October. On each trip, approximately 30 L. delicatula, representing the stages present at that site at that time, were collected. The L. delicatula collected alive were reared on potted A. altissima plants in netting cages in a growth chamber at 22.5 °C (days) and 15 °C (nights), 14:10 (L:D), and 65% relative humidity, as described in Clifton and Hajek [28]. Reared L. delicatula were monitored daily for 14 d after collection, and any L. delicatula that died were removed daily from cages. To promote fungal outgrowth, cadavers of first–third-instar nymphs were moved into individual wells in 24-well plates (Falcon 353047). Each of these plates was maintained in a tightly closed plastic container (17 × 17 × 5 cm), with wet paper towels to maintain high humidity. Cadavers of fourth-instar nymphs and adults were placed individually in 29 mL clear, tightly-lidded plastic cups containing 6 mL of 1.5% water agar.
During each collection trip in 2022, the ground was also searched for cadavers of L. delicatula that had died very recently, with no evidence of scavenging. Searching for cadavers occurred for either 15 min or until 30 cadavers had been collected. Cadavers were placed individually in water agar cups and were treated similarly to cadavers of reared L. delicatula that died in the quarantine laboratory. All cadavers from both rearing and cadaver collections in the field were kept under humid conditions at 23 °C for 1–4 weeks. In 2022, a total of 2177 living and 373 dead L. delicatula were collected. Among the live L. delicatula, 1097 (50.4%) died within 14 d while being reared.
In 2023, L. delicatula populations at Glen Run and Angora Fruit Farm were sampled on 20 July, and at each site, 20–38 live L. delicatula were collected and reared in the quarantine laboratory. An infested site in Owego, New York (Table 1), was also visited on 7 and 12 July and on 9 August, and 43–55 living L. delicatula were reared from each collection date. In 2023, the same procedures were employed, monitoring insects for 14 d and maintaining any resulting cadavers under high humidity. However, in 2023, immediately after discovery, cadavers were surface-sterilized with the following methods: rinsing in 0.75% sodium hypochlorite for 30 s, followed by rinsing with deionized water for 30 s before incubating at high humidity to promote fungal outgrowth.
2.2. Fungal Isolation and Identification
Fungi were isolated from outgrowth on each cadaver and grown on potato dextrose agar (PDA) with 0.3 g/L streptomycin at 23 °C for several weeks before being moved to 4 °C. Cultures were then grouped into morphotypes based on physical characteristics. Representatives of each morphotype were selected for sequencing and were then subcultured and grown at 23–25 °C until sampling for DNA extraction.
DNA was extracted from young cultures using the DNeasy Plant Mini Kit (Qiagen, Germantown, MD, USA). Fungal mycelium was added to AP1 buffer with 0.5 gm of 0.7 mm zirconia beads (BioSpec, Bartlesville, Oklahoma, USA) and beat for 1 min at 4.8 krpm. After bead beating, samples were incubated at 65 °C for 10 min with inversion every 3 min; all other steps were carried out following the manufacturer’s instructions. PCR was conducted on up to 5 loci for each isolate; primers used are listed in Table 2. The nuclear ribosomal DNA internal transcribed spacer region (ITS1-5.8S-ITS2 = ITS) was amplified for exemplar isolates of each species (Table 3) and most of the additional samples requiring sequencing (see Table S1) following the protocol of White et al. [29]. The nuclear ribosomal DNA large subunit (LSU) was amplified for some Cordycipitaceae samples following the protocol in Vilgalys and Hester [30]. The protein-encoding gene RNA polymerase II largest subunit (RPB1) was amplified for some samples of Cordycipitaceae following the protocol of Castlebury et al. [31] but with an initial denaturation of 2 min at 94 °C and annealing temperatures of 52 °C for Flavocillium bifurcatum and 45 °C for other Cordycipitaceae. The protein-encoding gene RNA polymerase II second-largest subunit (RPB2) was amplified for Samsoniella sp., Fusarium fujikuroi, and Fusarium graminearum (primer pair RPB2-5F2 and RPB2-7cR) following protocols of Sung et al. [32] but with an initial denaturation of 2 min at 94 °C, an annealing temperature of 57 °C [33], and a final extension of 5 min at 72 °C. For Akanthomyces muscarius, F. bifurcatum, F. fujikuroi, F. graminearum, and Samsoniella sp., RPB2 was amplified following Liu et al. [34] (primer pair RPB2-5F and RPB2-7cR). The protein-encoding gene translation elongation factor 1-α (TEF1-α) was amplified for most samples, following protocols of Rehner and Buckley [35] for Cordycipitaceae (primer pair 983F and 2218R) and Karlsson et al. [36] for Fusarium and Clonostachys spp. (primer pair EF-1 and EF-2). The protein-encoding gene ß-tubulin (TUB2) was amplified for Colletotrichum fioriniae, Cordyceps javanica, and Samsoniella sp. following the protocol in Weir et al. [37].
For specific identification, genera and species with records as insect pathogens in the literature were emphasized, and genera that are often considered common saprophytes, e.g., Cladosporium, Mucor, and Penicillium [15], were not evaluated further. Beauveria bassiana samples were excluded from molecular analyses because these cultures could be identified morphologically, and B. bassiana is the only Beauveria species that has been isolated from L. delicatula during extensive sampling in eastern Pennsylvania [45]. For each species, an exemplar isolate was chosen, sequences were deposited in GenBank, and the culture was deposited in the ARSEF culture collection (Table 4). When there were multiple samples of one species, usually these were also sequenced (Table S1), except in cases where the morphotype of the cultures were very distinctive (i.e., Sarocladium strictum). All samples were sequenced by the Biotechnology Resource Center, Cornell University, and resulting sequences were aligned and trimmed with Geneious Prime (2023.0.4). For identification, Fusarium samples were compared with sequences in the FUSARIOID-ID database [72], and all other samples were compared with sequences in the NCBI GenBank database. In cases where a clear species identity was not apparent after comparing sequences of multiple loci to the relevant database (i.e., A. muscarius and F. bifurcatum), morphology of the isolate was also evaluated for identification.
2.3. Testing Pathogenicity
In 2021 and 2022, cadavers of L. delicatula from rearing field collections were not surface-sterilized after death, creating the possibility that fungal isolates from cadavers were saprophytic and not pathogenic. Therefore, tests were conducted to confirm pathogenicity to L. delicatula, using the sequenced exemplar isolate for each fungal species. In 2023, a pathogenicity test was also conducted with C. fioriniae, although the cadaver had been surface-sterilized. Based on field and laboratory studies with B. bassiana [24,28], tests of pathogenicity were not necessary, and based on laboratory bioassays with C. javanica [28], complete pathogenicity testing was unnecessary (i.e., controls were not included during pathogenicity tests and numbers of L. delicatula tested were decreased).
To test pathogenicity, exemplar isolates were grown on PDA or SDA (Sabouraud dextrose agar) (Table S2) until adequate sporulation occurred. For some Fusarium isolates, growth under black lights without parafilm was necessary to promote conidial production (K. Myers and G. Bergstrom pers. comm.). Conidia were removed from plates by adding 6 mL of sterile 0.05% Tween per plate, disrupting the cultures with a glass cell spreader, and pouring the resulting suspension through cheesecloth to remove hyphae and agar. Most pathogenicity tests were conducted with 1 × 107 conidia/mL, although the initial tests with isolates from 2021 collections used 3.5 × 107 conidia/mL for Fusarium falsibabinda, 7.4 × 106 for Cordyceps cateniannulata, and 6.7 × 107 for A. muscarius.
Lycorma delicatula used in pathogenicity tests were collected in the field and caged on plants in the quarantine growth chamber for 1–12 d prior to pathogenicity tests. Because L. delicatula is univoltine and year-round, non-diapausing colonies were not available, and we used the life stages present in the field at the times that sporulating cultures were ready (Table S2). For inoculations, 20 L. delicatula nymphs or adults (as an exception, 6 were used for C. javanica) were cold-anesthetized at 4 °C for 2 min, placed in groups of 1–3 in centrifuge tubes containing a conidial suspension, and shaken by hand for 10 s. Insects were then briefly placed on a paper towel so that excess water would drain off and were then reared on A. altissima in cages for 14 d. For the first 3 d after inoculation, cages were enclosed in clear plastic bags with wet paper towels to maintain high humidity. For 14 d after inoculation, cages were monitored daily, and cadavers were removed and maintained individually under high humidity. For controls, 20 L. delicatula of the same stage being challenged with fungi were treated the same way but were initially shaken in groups in 0.05% Tween. For pathogenicity tests with the 3 species first collected in 2021, controls were not included, but additional L. delicatula caged nearby were observed, and mortality due to these fungi (A. muscarius, C. cateniannulata, and F. falsibabinda) was not observed. For pathogenicity tests with 2021 samples, subsequent fungal outgrowth from dead insects was examined to confirm fungal identity, while for tests with samples from 2022–23, fungi were reisolated from cadavers and cultures were examined to confirm identity.
2.4. Photographing Fungal Forms
Photos of fungal outgrowth from L. delicatula cadavers, conidia, and obverse and reverse sides of cultures were taken for exemplar isolates known to be pathogens. After the death of an infected insect, cadavers were maintained under high humidity. For most species, approximately 1 wk after death, fungal outgrowth was adequate, and the cadaver was stored at 4 °C until a photo was taken under a dissecting microscope at 7.5× magnification for adults and 10–15× for nymphs. Conidia were harvested and photos were taken at 400× under phase contrast, except in the case of F. graminearum, for which phase contrast was not used. Photos of cultures were all taken on PDA in 100 mm petri dishes. Photos of B. bassiana are not included because the appearance of this species is already well-documented [24,45].
2.5. Analyses
Comparisons of infection prevalence were conducted using data from 2022, when consistent, extensive sampling occurred across the season. To evaluate levels of infection by stage across the 2022 season, first–third instar nymphs were merged, 4th-instar nymphs were considered separately, and adults were grouped into three stages. Stage A1, extending from eclosion to 7 September, is spent feeding, aggregating, and beginning to disperse [73]. Stage A2, extending from 8 September to 5 October, is when courtship and mating occur and oviposition begins, and Stage A3, extending from 6 October to 2 November, includes oviposition as the major behavior, with limited courtship and mating. For analyses, aside from B. bassiana, insect pathogens are grouped as ‘opportunists’ [19], based on their lower abundance in this study or the fact that most were not generally considered major fungal entomopathogens [6].
The percentage of L. delicatula infected was calculated for each instar/stage by the number infected insects by all insect pathogens identified in this study divided by the total number of that instar/stage collected, merging data across sample dates. Frequencies by instar/stage were compared using chi-squared tests with a Bonferroni correction. The numbers of males versus females that died from B. bassiana or opportunistic pathogens (all other pathogens) were compared using chi-squared tests. Analyses were conducted using SAS (Version 9.4, SAS Institute, Inc., Cary, NC, USA).
3. Results
3.1. Pathogen Biodiversity
A total of sixteen species in six families of the Hypocreales were isolated from field-collected L. delicatula (Table 3). For most species, fungal outgrowth from cadavers did not provide quick identification; in the field, there was no fungal outgrowth from living insects and rarely from cadavers, and only after collection and incubation under humid conditions did outgrowth occur. Then, for most opportunistic pathogens, white, cobweb-like outgrowth occurred (Figure 1, Figure 2, Figure 3, Figure 4 and Figure 5), and both microscopy and sequencing were required for identification. The exception was B. bassiana, for which cadavers were more readily recognizable with abundant bright white mycelium and dense balls of conidia (=‘spore balls’) [74]. Stromata grew from one cadaver of an L. delicatula adult killed by C. cateniannulata (Figure 2(b1)), but this growth occurred long after collection.
3.2. Pathogen Prevalence
Merging reared insects and collected cadavers in 2022, B. bassiana was collected most frequently (59.2% of all entomopathogenic fungi), while F. fujikuroi (16.8%) and Sarocladium strictum (12.3%) were the next most frequently found pathogens (Table 5). These three species were also the most widely distributed, being found at 5–6 of the 6 collection sites. Although we sampled throughout the season, the remaining species were rarely collected and were never found at all sites (Table 5). The most common genus was Fusarium, with five species collected, yet only F. fujikuroi-infected L. delicatula were collected more than five times.
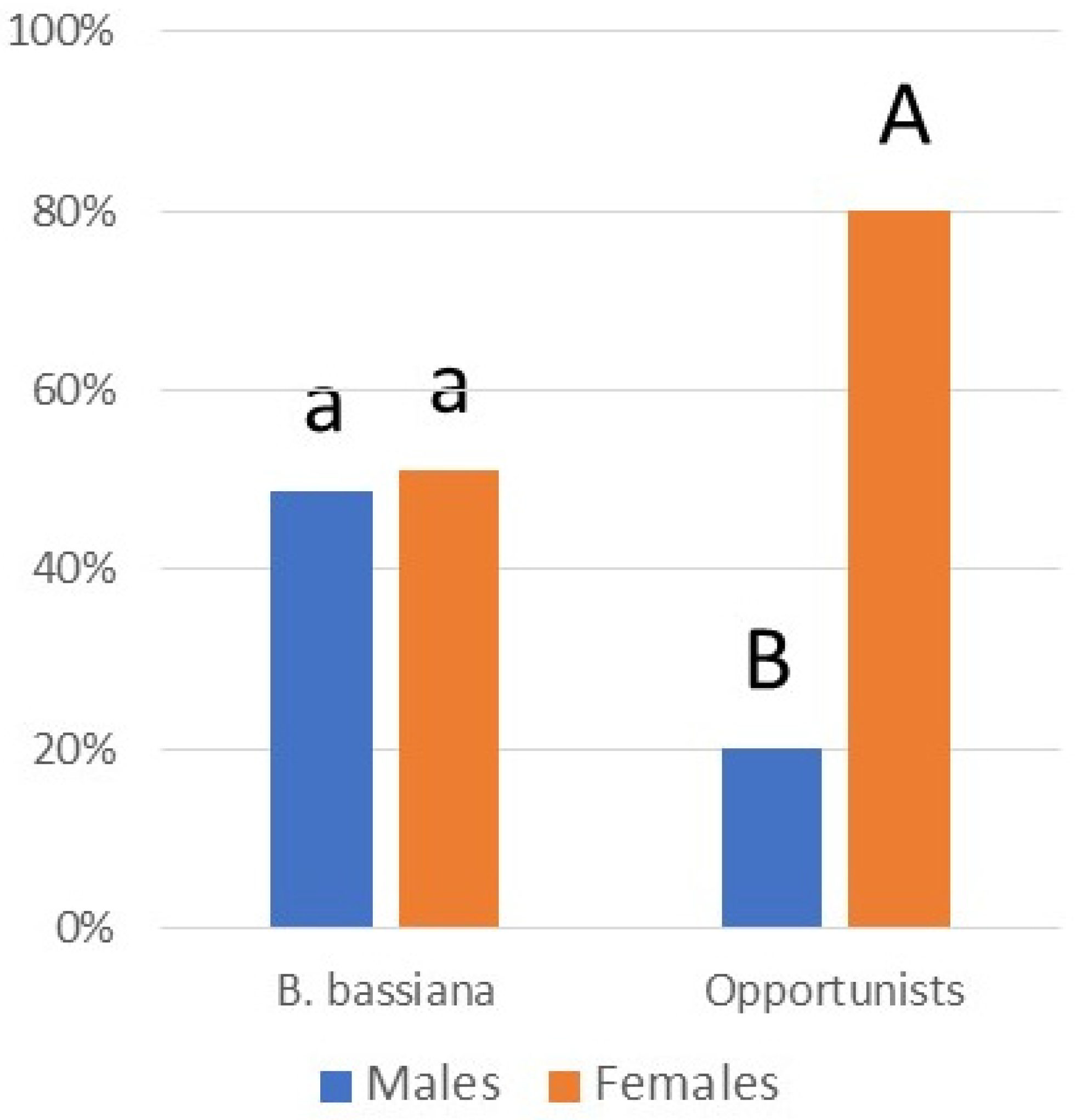
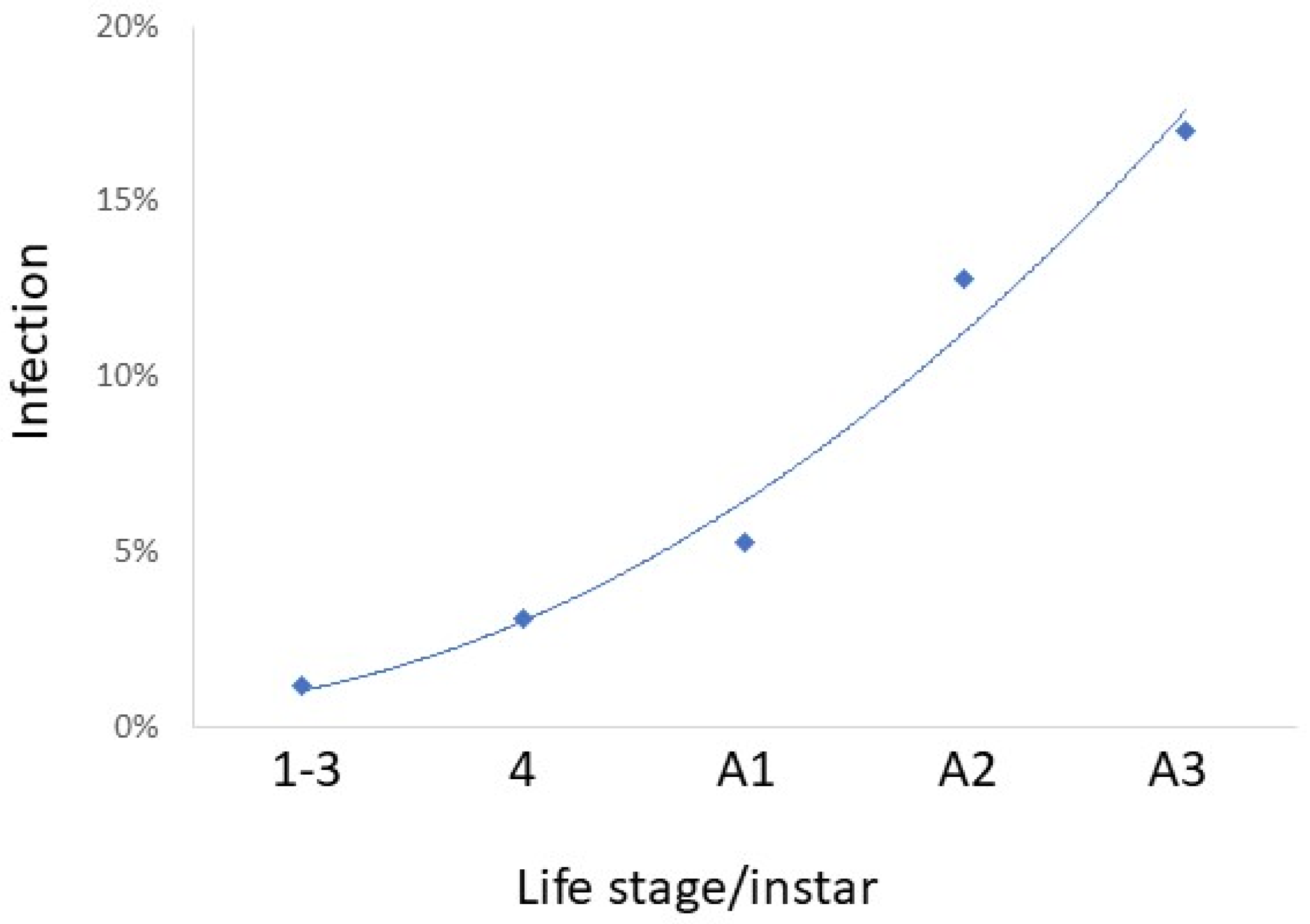
While infection prevalence by species was often low, together, these pathogens caused a total of 6.7% mortality. Considering all 14 pathogens collected in the field in 2022 (this includes all species in Table 3 except C. javanica and C. fioriniae), percentages of L. delicatula infection increased through the season, with little infection in nymphs and infection increasing especially through adult stages A2 and A3 (R2 = 0.9777) (Figure 6). Comparisons among instars/stages revealed that infection was equivalent for stages A2 and A3, when infection levels were greater than other instars/stages (chi-squared tests; p < 0.05). Among adults infected by opportunistic pathogens (i.e., all species except B. bassiana), significantly more adults females were infected than adult males (chi-squared = 21.600; p < 0.0001) (Figure 7). Among adults infected by B. bassiana, numbers of infected females were not significantly different from those of males (chi-squared = 0.047; p = 0.8292) (Figure 7).
In 2022, among cadavers of recently dead L. delicatula, infection by B. bassiana was much more abundant (81.7%) than B. bassiana infections among reared insects (18.7%) (chi-squared = 40.8427; p < 0.00001). Few cadavers of nymphs were found (Table 5). Also, few cadavers of the opportunistic pathogens were found at sites (18.8% of cadavers), and most of these pathogens were discovered through rearings (68.6% of entomopathogenic fungi from rearings).
4. Discussion
This study increased the known number of naturally occurring entomopathogenic fungi infecting L. delicatula from 4 species [24,27] to a total of 19. Numerous isolated fungal species that we report as pathogens of L. delicatula are not well known as entomopathogens or have not been reported as entomopathogens previously, e.g., F. falsibabinda. Our sampling demonstrated the highest levels of infection among mid- and late-stage adults (i.e., from September to November) (Figure 6). Lycorma delicatula adults generally live from late July to early November [75], which is a long life compared to that of adults of many insect species. This host is invasive, with no other members of the same family (Fulgoridae) in the areas sampled [76], so we assume that these pathogens are generalists and not specialists. Boomsma et al. [77] hypothesized that in the case of generalist entomopathogenic fungi, average spore doses usually present in the environment would be low and, as such, these possible insect pathogens would usually only be able to kill compromised hosts. Studies conducted with B. bassiana have shown that as L. delicatula adults age, they become weaker ([28]). As confirmation, we found the highest levels of infection in the A2 and A3 adult stages that had already been adults for at least one month. Because L. delicatula are invasives relatively new to the areas sampled, many of the pathogens being reported in this study could be considered opportunists that have adopted this invasive as a new food source.
In general, fungi have diverse ecological functions, including acting as saprophytes, endophytes, and pathogens of plants, animals, and fungi. Aside from B. bassiana [45], the pathogens identified in this study have not been reported previously as L. delicatula pathogens and have usually been reported as having additional ecological functions, leading to multifunctional lifestyles. For example, C. fioriniae, which killed L. delicatula in this study, is also well known as a plant endophyte and plant pathogen, and F. avenaceum and F. graminearum are well known as plant pathogens (see Table 3). This provides further evidence that many of the entomopathogenic fungi identified in this study are accidental opportunists that, in the space and time when they were collected, were acting as insect pathogens. In fact, secondary metabolites that are considered toxic to insects have been identified from fungi not principally known as insect pathogens; for example, many Fusarium species, including Fusarium concentricum, produce beauvericin [56], and S. strictum produces bassianolide, among other compounds [62].
Perhaps the occurrence and impact of these accidental opportunists are not recognized more frequently because their fungal outgrowth on cadavers is often not evident or easily identifiable. For example, several days after host death, bodies of L. delicatula killed by B. bassiana become covered with white mycelium, while many species in this study often became covered with paler, cobweb-like mycelium (Figure 1, Figure 2, Figure 3, Figure 4 and Figure 5). One significant exception was C. cateniannulata (Figure 2(b1)), from which large stromata grew. However, we only collected one specimen killed by C. cateniannulata, and cadavers killed by this fungus during pathogenicity tests did not produce stromata.
It is possible that our intensive sampling in 2022, when we sampled nymphs extensively as well as adults, could have led to underestimated infection levels compared with other years due to abiotic factors. During July and August 2022, there were drought conditions in the areas being sampled (Reading area and East Stroudsburg, Pennsylvania [78]), which would have decreased fungal transmission. It was surprising not to find M. pemphigi or O. delicatula [27] in 2022, especially as M. pemphigi had been collected in 2018 at one of the sites that were intensively sampled in 2022. Batkoa major had been very abundant in early October 2018 during a period with abundant rainfall and was recovered from field-collected L. delicatula during 2019–2021 (E.H.C. and D.C. Harris, unpublished data). However, during 2022, under intensive sampling but drier conditions, B. major infections were not observed.
Although the low levels of infection by most of these species will not individually have a strong impact on population suppression, when added together, the impact is greater. Most infections were recorded in stages A2 and A3 (R2 = 0.9777) (Figure 6), when mating and oviposition occur [73]. Among infected A2- and A3-stage adults, equal numbers of females and males were infected with B. bassiana, but among the opportunistic species, far more females were infected than males (Figure 7). Further studies are necessary to quantify the impact of infection by these fungi on L. delicatula population densities under field conditions.
Why would so much more infection occur among adults than nymphs? First, adults begin to eclose in July, and many live until early November [75]. Each nymphal instar persists for approximately 2 weeks [73], while individual adults can live >4 months [75]. So, adults are present in the environment much longer than nymphs. The soil is generally considered a reservoir for spores of hypocrealean entomopathogenic fungi [79]. At the sites sampled, nymphs were rarely recovered near the forest floor. In contrast, winged adults are highly mobile and often feed on tree roots or just above the base of the tree, close to the soil (unpubl. data). We hypothesize that by feeding in these locations, adults are more likely to become infected compared with nymphs.
Fusarium falsibabinda has not been reported previously as an insect pathogen. In this case, our laboratory isolated and assessed Koch’s postulates for this species by random chance. During this study, many more fungal species, including more Fusarium species, were isolated from dead L. delicatula in 2022, but we prioritized testing species known to be insect pathogens. Worldwide, several dozen Fusarium species have been reported as insect pathogens, but all of these are generally known from few isolates and host species [56,80]. In some reported instances, Fusarium species were only isolated from a challenged insect with the ‘Galleria bait method’ [81], and Koch’s postulates were confirmed using lab colonies of Galleria mellonella instead of wild insects. Epizootics in insect populations caused by Fusarium species have been documented, but not due to any of the Fusarium species isolated in this study [82]. However, the white, cobweb-like growth on cadavers exhibited by Fusarium species (Figure 3(b1,c1) and Figure 4(a1,b1,c1)) is shared with the other opportunistic species in this study, making species identifications in the field difficult and likely contributing to an observation bias against species of Fusarium and hypocrealean genera other than the more easily recognizable B. bassiana.
Numerous opportunistic pathogens we have reported are less well known. This is the first report of C. cateniannulata, Clonostachys eriocamporesii, F. bifurcatum, F. falsibabinda, and Samsoniella sp. in North America. However, North American isolates of some of these taxa may have been previously reported under other names. This is the first study to conduct Koch’s postulates with a species of Samsoniella against insects, although species of Samsoniella have been isolated from dead insects [83] and have also been found to be pathogens of nematodes [84]. Future taxonomic work is needed to determine which species of this recently named genus are present in North America.
Cladosporium and Mucor species have recently been confirmed as important pathogens in hemipteran populations, with potential for biological control [85,86,87]. Unfortunately, during this study, we were unable to evaluate the pathogenicity of additional potentially pathogenic species such as these that that were isolated from dead L. delicatula. Although in 2022, fungi were isolated from dead L. delicatula, the cadavers had not been surface-sterilized, so tests of pathogenicity were needed to confirm pathogenicity. Because we emphasized species already known to be entomopathogens, this study perhaps underestimated opportunists present in the field that were infecting L. delicatula, and future trials using Koch’s postulates could help to identify additional entomopathogenic species.
Although in this study we report 15 species that were previously not known as pathogens of L. delicatula, we hypothesize that 2021–2023 were probably not the first years for the infection of these invasives by these opportunistic generalists. Lycorma delicatula was first found in only one county in Pennsylvania in 2014 [21] and has been spreading since [20]. During our earlier studies [24,27], we were limited in methodology, and these opportunists could have gone undetected or could have been removed from the environment by scavengers, saprophytes, or weathering, preventing collection. As L. delicatula is constantly spreading, further studies could discover more species of entomopathogens infecting this invasive in new areas.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/2075-4450/14/12/912/s1: Table S1: GenBank accession numbers for additional sequenced isolates of enotomopathogenic fungi infecting L. delicatula; Table S2: Methods used for challenging L. delicatula to confirm pathogenicity of fungal species.
Author Contributions
Conceptualization, E.H.C., A.E.H.; methodology, E.H.C., T.A.E. and A.E.H.; formal analysis, T.A.E., E.H.C. and A.E.H.; investigation, E.H.C. and T.A.E.; writing—original draft preparation, A.E.H.; writing—review and editing, E.H.C., T.A.E. and A.E.H.; supervision, A.E.H. All authors have read and agreed to the published version of the manuscript.
Funding
This work was funded by USDA NIFA 2019-51181-30014 (A.E.H.).
Data Availability Statement
The data supporting the findings of study are available upon reasonable request to the corresponding author.
Acknowledgments
We thank Anais Ocegueda, Destiny Smith, and David Harris for technical assistance in the laboratory and field. We thank Brendan Lederer of Berks County Parks and Recreation, Matt Schultz and the Pocono Heritage Land Trust, Smithfield Township (Minisink), and John Rost (Sinking Spring) for assistance with finding sites and allowing us to use study sites. We thank Jennifer Gonzalez, Kathie Hodge, Kevin Myers, and Gary Bergstrom for advice and assistance.
Conflicts of Interest
Author Eric H. Clifton was employed by the company BioWorks Inc. The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
- Keane, R.M.; Crawley, M.J. Exotic plant invasions and the enemy release hypothesis. Trends Ecol. Evol. 2002, 17, 164–170. [Google Scholar] [CrossRef]
- Mitchell, C.E.; Blumenthal, D.; Jarosik, V.; Puckett, E.E.; Pyšek, P. Controls on pathogen species richness in plants’ introduced and native ranges: Roles of residence time, range size and host traits. Ecol. Lett. 2010, 13, 1525–1535. [Google Scholar] [CrossRef]
- Phillips, B.L.; Kelehear, C.; Pizzatto, L.; Brown, G.P.; Baarton, D.; Shine, R. Parasites and pathogens lag behind their host during periods of host range advance. Ecology 2010, 91, 872–881. [Google Scholar] [CrossRef]
- Flory, S.L.; Clay, K. Pathogen accumulation and long-term dynamics of plant invasions. J. Ecol. 2013, 101, 607–613. [Google Scholar] [CrossRef]
- Seebens, H.; Blackburn, T.M.; Dyer, E.E.; Benovesi, P.; Hulme, P.E.; Jeschke, J.M.; Pagad, S.; Pyšek, P.; Winter, M.; Arianoutsou, M.; et al. No saturation in the accumulation of alien species worldwide. Nat. Comm. 2017, 8, 14435. [Google Scholar] [CrossRef]
- Dara, S.K.; Montalva, C.; Barta, M. Microbial control of invasive forest pests with entomopathogenic fungi: A review of the current situation. Insects 2019, 10, 341. [Google Scholar] [CrossRef]
- Araújo, J.P.M.; Hughes, D.P. Diversity of entomopathogenic fungi: Which groups conquered the insect body? Adv. Gen. 2016, 94, 1–39. [Google Scholar]
- Sacco, N.E.; Hajek, A.E. Diversity and breadth of host specificity among arthropod pathogens in the Entomophthoromycotina. Microorganisms 2023, 11, 1658. [Google Scholar] [CrossRef]
- Domingues, M.M.; dos Santos, P.L.; Costa Gea, B.C.; de Carvalho, V.R.; de Oliveira, F.N.; Solian, E.P.; Serrao, J.E.; Zanuncio, J.C.; Zanetti, R.; Wilcken, C.F. Entomopathogenic fungi, isolated from soils and Bemisia tabaci (Hemiptera: Aleyrodidae) adults, to manage the Eucalyptus red gum lerp psyllid Glycaspis brimblecombei (Hemiptera: Aphalaridae). J. Econ. Entomol. 2022, 115, 1886–1893. [Google Scholar] [CrossRef]
- Biryol, S.; Araz, N.; Eski, A.; Akturk, R.; Aksu, Y.; Gokturk, B.C.; Bilgin, L.; Demir, I. Biodiversity and pathogenicity of entomopathogenic fungi associated with the lesser spruce sawfly, Pristophora abietina. Entomol. Exp. Appl. 2021, 169, 414–423. [Google Scholar] [CrossRef]
- Kovač, M.; Gorczak, M.; Wrzosek, M.; Tkaczuk, C.; Pernek, M. Identification of entomopathogenic fungi as naturally occurring enemies of the invasive oak lace bug, Corythuca arcuata (Say) (Hemiptera: Tingidae). Insects 2020, 11, 679. [Google Scholar] [CrossRef] [PubMed]
- Corallo, B.; Simeto, S.; Martinez, G.; Gomez, D.; Abreo, E.; Altier, N.; Lupo, S. Entomopathogenic fungi naturally infecting the eucalypt bronze bug, Thaumastocoris peregrinus (Heteroptera: Thaumastocoridae), in Uruguay. J. Appl. Entomol. 2019, 143, 542–555. [Google Scholar] [CrossRef]
- Sharma, L.; Gonçalves, F.; Oliveira, I.; Torres, L.; Marques, G. Insect-associated fungi from naturally mycosed vine mealybug Planococcus ficus (Signoret) (Hemiptera: Pseudococcidae). Biocontr. Sci. Technol. 2018, 28, 122–141. [Google Scholar] [CrossRef]
- Dao, H.T.; Beattie, G.A.C.; Rossman, A.Y.; Burgess, L.W.; Holford, P. Four putative entomopathogenic fungi of armoured scale insects on Citrus in Australia. Mycol. Progr. 2016, 15, 47. [Google Scholar] [CrossRef]
- Gouli, V.; Gouli, S.; Marcelino, J.A.P.; Skinner, M.; Parker, B.L. Entomopathogenic fungi associated with exotic invasive insect pests in northeastern forests of the USA. Insects 2013, 4, 631–645. [Google Scholar] [CrossRef] [PubMed]
- Reid, W.R.; Parker, B.L.; Gouli, S.Y.; Skinner, M.; Gouli, V.V.; Teillon, H.B. Fungi associated with the hemlock woolly adelgid, Adelges tsugae, and assessment of entomopathogenic isolates for management. J. Insect Sci. 2010, 10, 62. [Google Scholar] [CrossRef] [PubMed]
- Tanyeli, E.; Sevin, A.; Demirbag, Z.; Eroglu, M.; Demir, I. Isolation and virulence of entomopathogenic fungi against the great spruce bark beetle, Dendroctonus micans (Kugelann) (Coleoptera: Scolytidae). Biocontr. Sci. Technol. 2010, 20, 695–701. [Google Scholar] [CrossRef]
- Venugopal Rao, N.; Reddy, A.S.; Tirumala Rao, K. Natural enemies of cotton whitefly, Bemisia tabaci Gennadius in relation to host population and weather factors. J. Biol. Control 1989, 3, 10–12. [Google Scholar]
- Kaya, H.K.; Vega, F.E. Scope and basic principles of insect pathology. In Insect Pathology, 2nd ed.; Vega, F.E., Kaya, H.K., Eds.; Academic Press: Amsterdam, The Netherlands, 2012; pp. 1–12. [Google Scholar]
- NYS IPM (New York State Integrated Pest Management Program). Interactive Spotted Lanternfly Map. Available online: http://Go.nysipm.org/slf-map-i (accessed on 26 October 2023).
- Urban, J.M.; Leach, H. Biology and management of the spotted lanternfly, Lycorma delicatula (Hemiptera: Fulgoridae), in the United States. Annu. Rev. Entomol. 2023, 68, 151–167. [Google Scholar] [CrossRef]
- Soper, R.S. Pathogens of leafhoppers and planthoppers. In The Leafhoppers and Planthoppers; Nault, L.R., Rodriguez, J.G., Eds.; John Wiley & Sons: New York, NY, USA, 1985; pp. 469–488. [Google Scholar]
- Li, M.-Y.; Lin, H.-F.; Li, S.-G.; Xu, A.; Feng, M.-F. Efficiency of entomopathogenic fungi in the control of eggs of the brown planthopper Nilaparvata lugens Stål (Homoptera: Delphacidae). Afr. J. Microbiol. Res. 2012, 6, 7162–7167. [Google Scholar]
- Clifton, E.H.; Castrillo, L.A.; Gryganskyi, A.; Hajek, A.E. A pair of native fungal pathogens drives decline of a new invasive herbivore. Proc. Natl. Acad. Sci. USA 2019, 116, 9178–9180. [Google Scholar] [CrossRef] [PubMed]
- Hajek, A.E.; Clifton, E.H.; Stefanik, S.E.; Harris, D.C. Batkoa major infecting the invasive planthopper Lycorma delicatula. J. Invertebr. Pathol. 2022, 194, 107821. [Google Scholar] [CrossRef] [PubMed]
- Ortiz-Urquiza, A. The split personality of Beauveria bassiana: Understanding the molecular basis of fungal parasitism and mutualism. mSystems 2021, 6, e00766-21. [Google Scholar] [CrossRef] [PubMed]
- Clifton, E.H.; Castrillo, L.A.; Hajek, A.E. Discovery of two hypocrealean fungi infecting spotted lanternflies, Lycorma delicatula: Metarhizium pemphigi and a novel species, Ophiocordyceps delicatula. J. Invertebr. Pathol. 2021, 186, 107689. [Google Scholar] [CrossRef] [PubMed]
- Clifton, E.H.; Hajek, A.E. Efficacy of Beauveria bassiana and Cordyceps javanica mycoinsecticides against spotted lanternflies, Lycorma delicatula, in laboratory bioassays. Biocontr. Sci. Technol. 2021, 32, 824–836. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M., Gelfand, D., Sninsky, J., White, T., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef]
- Castlebury, L.A.; Rossman, A.Y.; Sung, G.H.; Hyten, A.S.; Spatafora, J.W. Multigene phylogeny reveals new lineage for Stachybotrys chartarum, the indoor air fungus. Mycol. Res. 2004, 108, 864–872. [Google Scholar] [CrossRef]
- Sung, G.-H.; Sung, J.-M.; Hywel-Jones, N.L.; Spatafora, J.W. A multi-gene phylogeny of Clavicipitaceae (Ascomycota, Fungi): Identification of localized incongruence using a combinational bootstrap approach. Mol. Phylogenet. Evol. 2007, 44, 1204–1223. [Google Scholar] [CrossRef]
- Wang, M.M.; Crous, P.W.; Sandoval-Denis, M.; Han, S.L.; Liu, F.; Liang, J.M.; Duan, W.J.; Cai, L. Fusarium and allied genera from China: Species diversity and distribution. Persoonia 2022, 48, 1–53. [Google Scholar] [CrossRef]
- Liu, Y.J.; Whelen, S.; Hall, B.D. Phylogenetic relationships among Ascomycetes: Evidence from an RNA polymer[a]se II subunit. Mol. Biol. Evol. 1999, 16, 1799–1808. [Google Scholar] [CrossRef]
- Rehner, S.A.; Buckley, E. A Beauveria phylogeny inferred from nuclear ITS and EF1-α sequences: Evidence for cryptic diversification and links to Cordyceps teleomorphs. Mycologia 2005, 97, 84–98. [Google Scholar] [CrossRef] [PubMed]
- Karlsson, I.; Edel-Hermann, V.; Gautheron, N.; Durling, M.B.; Kolseth, A.K.; Steinberg, C.; Persson, P.; Friberg, H. Genus-specific primers for study of Fusarium communities in field samples. Appl. Environ. Microbiol. 2016, 82, 491–501. [Google Scholar] [CrossRef] [PubMed]
- Weir, B.S.; Johnston, P.R.; Damm, U. The Colletotrichum gloeosporioides species complex. Stud. Mycol. 2012, 73, 115–180. [Google Scholar] [CrossRef] [PubMed]
- Korabečná, M.; Liška, V.; Fajfrlík, K. Primers ITS1, ITS2 and ITS4 detect the intraspecies variability in the internal transcribed spacers and 5.8S rRNA gene region in clinical isolates of fungi. Folia Microbiol. 2003, 48, 233–238. [Google Scholar] [CrossRef] [PubMed]
- Lutzoni, F. Lutzoni Lab Evolution, Ecology, and Genomics of Fungal Symbioses: Primer Sequences. 2014. Available online: https://lutzonilab.org/primer-sequences/ (accessed on 3 September 2023).
- Matheny, P.B.; Liu, Y.J.; Ammirati, J.F.; Hall, B.D. Using RPB1 sequences to improve phylogenetic inference among mushrooms (Inocybe, Agaricales). Amer. J. Bot. 2002, 89, 688–698. [Google Scholar] [CrossRef]
- O’Donnell, K.; Kistler, H.C.; Cigelnik, E.; Ploetz, R.C. Multiple evolutionary origins of the fungus causing Panama disease of banana: Concordant evidence from nuclear and mitochondrial gene genealogies. Proc. Natl. Acad. Sci. USA 1998, 95, 2044–2049. [Google Scholar] [CrossRef]
- O’Donnell, K.; Cigelnik, E. Two divergent intragenomic rDNA ITS2 types within a monophyletic lineage of the fungus Fusarium are nonorthologous. Mol. Phylogenet. Evol. 1997, 7, 103–116. [Google Scholar] [CrossRef]
- Saidi, A.; Mebdoua, S.; Mecelem, D.; Al-Hoshani, N.; Sadrati, N.; Boufahja, F.; Bendif, H. Dual biocontrol potential of the entomopathogenic fungus Akanthomyces muscarius against Thaumetopoea pityocampa and plant pathogenic fungi. Saudi J. Biol. Sci. 2023, 30, 103719. [Google Scholar] [CrossRef]
- Broumandnia, F.; Rajabpour, A. Morphological and molecular identification of four isolates of the entomopathogenic fungal genus Akanthomyces and their effects against Bemisia tabaci on cucumber. Bull. Entomol. Res. 2021, 111, 628–636. [Google Scholar] [CrossRef]
- Clifton, E.H.; Castrillo, L.A.; Jaronski, S.T.; Hajek, A.E. Cryptic diversity and virulence of Beauveria bassiana recovered from Lycorma delicatula (spotted lanternfly) in eastern Pennsylvania. Front. Insect Sci. 2023, 3, 1127682. [Google Scholar] [CrossRef]
- Hajek, A.E.; Meyling, N.V. Ecology of invertebrate pathogens: Fungi. In Ecology of Invertebrate Diseases; Hajek, A.E., Shapiro-Ilan, D., Eds.; John Wiley & Sons: Hoboken, NJ, USA, 2018; pp. 327–377. [Google Scholar]
- Montes-Bazurto, L.G.; Bustillo-Pardey, A.E.; Medina-Cárdenas, H.C. Cordyceps cateniannulata, a novel entomopathogenic fugus to control Stenoma impresella Busck (Lepidoptera: Elachistidae) in Colombia. J. Appl. Entomol. 2020, 144, 788–796. [Google Scholar] [CrossRef]
- Wang, Y.-B.; Wang, Y.; Fan, Q.; Duan, D.-E.; Zhang, G.-D.; Dai, R.-Q.; Dai, Y.-D.; Zeng, W.-B.; Chen, Z.-H.; Li, D.-D.; et al. Multigene phylogeny of the family Cordycipitaceae (Hypocreales): New taxa and the systematic position of the Chinese cordycipitoid fungus Paecilomyces hepiali. Fungal Div. 2020, 103, 1–46. [Google Scholar] [CrossRef]
- Rodrigues, J.; Rocha, L.F.N.; Martinez, J.M.; Montalva, C.; Humber, R.A. Clonostachys spp., natural mosquito antagonists, and their prospects for biological control of Aedes aegypti. Parasitol. Res. 2022, 121, 2979–2984. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Wu, Q.; Fan, J.; Huang, J.; Wu, Z.; Lin, J.; Bin, S.; Shu, B. Effects of the entomopathogenic fungus Clonostachys rosea on mortality rates and gene expression profiles in Diaphorina citri adults. J. Invertebr. Pathol. 2021, 179, 107539. [Google Scholar] [CrossRef]
- Sun, Z.-B.; Li, S.-D.; Ren, Q.; Xu, J.-L.; Lu, X.; Sun, L.-H. Biology and applications of Clonostachys rosea. J. Appl. Microbiol. 2020, 129, 486–495. [Google Scholar] [CrossRef]
- Batta, Y.A. The first report on entomopathogenic effect of Fusarium avenaceum (Fries) Saccardo (Hypocreales, Ascomycota) against rice weevil (Sitophilus oryzae L.: Curculionidae, Coleoptera). J. Entomol. Acarol. Res. 2012, 44, e11. [Google Scholar] [CrossRef]
- Rajagopal, K.; Suryanarayanan, T.S. Isolation of endophytic fungi from leaves of neem (Azadirachta indica A. Juss.). Curr. Sci. 2000, 78, 1375–1378. [Google Scholar]
- Uhlig, S.; Jestoi, M.; Parikka, P. Fusarium avenaceum—The North European situation. Internatl. J. Food Microbiol. 2007, 119, 17–24. [Google Scholar] [CrossRef]
- Qiu, H.-L.; Fox, E.G.P.; Qin, C.-S.; Yang, H.; Tian, L.-Y.; Wang, D.-S.; Xu, J.-Z. First record of Fusarium concentricum (Hypocreales: Hypocreaceae) isolated from the moth Polychrosis cunninhamiacola (Lepidoptera: Tortricidae) as an entomopathogenic fungus. J. Insect Sci. 2023, 23, 2. [Google Scholar] [CrossRef] [PubMed]
- Sharma, L.; Marques, G. Fusarium, an entomopathogen—A myth or reality? Pathogens 2018, 7, 93. [Google Scholar] [CrossRef]
- Dewing, C.; Van der Nest, M.A.; Santana, Q.C.; Proctor, R.H.; Wingfield, B.D.; Steenkamp, E.T.; De Vos, L. Characterization of host-specific genes from pine- and grass-associated species of the Fusarium fujikuroi species complex. Pathogens 2022, 11, 858. [Google Scholar] [CrossRef] [PubMed]
- Ameen, M.K.M. Screening of Fusarium isolates pathogenicity in vitro by using the larvae of Galleria mellonella L. J. Basrah Res. Sci. 2012, 38, 19–28. [Google Scholar]
- Trail, F. For blighted waves of grain: Fusarium graminearum in the postgenomics era. Plant Physiol. 2009, 149, 103–110. [Google Scholar] [CrossRef] [PubMed]
- Lofgren, L.A.; LeBlanc, N.R.; Certano, A.K.; Nachtigall, J.; LaBine, K.M.; Riddle, J.; Broz, K.; Dong, Y.; Bethan, B.; Kafer, C.W.; et al. Fusarium graminearum: Pathogen or endophyte of North American grasses. New Phytol. 2018, 217, 1203–1212. [Google Scholar] [CrossRef] [PubMed]
- El-Sayed, A.S.A.; Moustafa, A.H.; Hussein, H.A.; El-Sheikh, A.A.; El-Shafrey, S.N.; Fathy, N.A.M.; Enan, G.A. Potential insecticidal activity of Sarocladium strictum, an endophyte of Cynanchum acutum, against Spodoptera littoralis, a polyphagous insect pest. Biocat. Agric. Biotechnol. 2020, 24, 101524. [Google Scholar] [CrossRef]
- Blaszczyk, L.; Waskiewicz, A.; Gromadzka, K.; Mikolajczak, K.; Chelkowski, J. Sarocladium and Lecanicillium associated with maize seeds and their potential to form selected secondary metabolites. Biomolecules 2021, 11, 98. [Google Scholar] [CrossRef]
- Tagne, A.; Neergaard, E.; Hansen, H. Studies of host-pathogen interaction between maize and Acremonium strictum from Cameroon. Eur. J. Plant Pathol. 2002, 108, 93–102. [Google Scholar] [CrossRef]
- Racedo, J.; Salazar, S.M.; Castagnaro, A.P.; Díaz Ricci, J.C. A strawberry disease caused by Acremonium strictum. Eur. J. Plant Pathol. 2013, 137, 649–654. [Google Scholar] [CrossRef]
- Kim, J.-C.; Choi, G.J.; Kim, H.-J.; Kim, H.T.; Ahn, J.W.; Cho, K.Y. Verlamelin, an antifungal compound produced by a mycoparasite, Acremonium strictum. Plant Pathol. J. 2002, 18, 102–105. [Google Scholar] [CrossRef]
- Gonzalez, J.B.; Lambert, C.A.; Foley, A.M.; Hajek, A.E. First report of Colletotrichum fioriniae infections in brown marmorated stink bugs, Halyomorpha halys. J. Invertebr. Pathol. 2023, 199, 107939. [Google Scholar] [CrossRef]
- Marcelino, J.A.P.; Gouli, S.; Giordano, R.; Gouli, V.V.; Parker, B.L.; Skinner, M. Fungi associated with a natural epizootic in Fiorinia externa Ferris (Hemiptera: Diaspididae) populations. J. Appl. Entomol. 2009, 133, 82–89. [Google Scholar] [CrossRef]
- Marcelino, J.A.P.; Gouli, S.; Parker, B.L.; Skinner, M.; Schwarzberg, L.; Giordano, R. Host plant associations of an entomopathogenic variety of the fungus, Colletotrichum acutatum, recovered from the elongate hemlock scale, Fiorinia externa. J. Insect Sci. 2009, 9, 25. [Google Scholar] [CrossRef]
- Batta, Y. Entomopathogenic effect of Trichothecium roseum (Pers.) Link (Hypocreales: Ascomycota) against Pauropsylla buxtoni (Psylloidea: Hemiptera) infesting Ficus carica leaves and its potential use as biocontrol agent of the insect. J. Appl. Microbiol. 2020, 129, 400–410. [Google Scholar] [CrossRef] [PubMed]
- Götz, M.; Karbowy-Thongbai, B. First detection of Trichothecium roseum causing leaf spots on tomato in Germany. Plant Dis. 2023, 107, 1233. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.; Duan, X.; Cai, P.; Zhang, W.; Liu, Y.; Cui, J.; Li, Z.; Qiu, Z. Biocontrol action of Trichothecium roseum against the wheat powdery mildew fungus Blumeria graminis f. sp. tritici. Front. Sustain. Food Syst. 2022, 6, 998830. [Google Scholar] [CrossRef]
- Crous, P.W.; Lombard, L.; Sandoval-Denis, M.; Seifert, K.A.; Schroers, H.-J.; Chaverri, P.; Gené, J.; Guarro, J.; Hirooka, Y.; Bensch, K.; et al. Fusarium: More than a node or a foot-shaped basal cell. Stud. Mycol. 2021, 98, 100116. [Google Scholar] [CrossRef] [PubMed]
- Cooperband, M.F.; Murman, K. Responses of adult spotted lanternflies to artificial aggregations composed of all males or females. Front. Insect Sci. 2022, 2, 981832. [Google Scholar] [CrossRef]
- Humber, R.A. Identification of entomopathogenic fungi. In Manual of Techniques in Invertebrate Pathology; Lacey, L.A., Ed.; Academic Press: Amsterdam, The Netherlands, 2012; pp. 151–187. [Google Scholar]
- Urban, J.M. Perspective: Shedding light on spotted lanternfly impacts in the USA. Pest Mgmt. Sci. 2020, 76, 10–17. [Google Scholar] [CrossRef]
- Bartlett, C.R.; O’Brien, L.B.; Wilson, W.W. A review of the planthoppers (Hemiptera: Fulgoroidea) of the United States. Mem. Amer. Entomol. Soc. 2014, 50, 39–69. [Google Scholar]
- Boomsma, J.J.; Jensen, A.B.; Meyling, N.V.; Eilenberg, J. Evolutionary interaction networks of insect pathogenic fungi. Annu. Rev. Entomol. 2014, 59, 467–485. [Google Scholar] [CrossRef]
- NRCC (Northeast Regional Climate Center). NOWData (NOAA Online Weather Data). Available online: https://www.nrcc.cornell.edu/wxstation/nowdata.html (accessed on 15 September 2023).
- Chandler, D. Basic and applied research on entomopathogenic fungi. In Microbial Control of Insect and Mite Pests; Lacey, L.A., Ed.; Academic Press: Cambridge, MA, USA, 2017; pp. 69–89. [Google Scholar]
- Santos, A.C.S.; Diniz, A.G.; Tiago, P.V.; Oliveira, N.T. Entompathogenic Fusarium species: A review of their potential for biological control of insects, implications and prospects. Fungal Biol. Rev. 2020, 34, 41–57. [Google Scholar] [CrossRef]
- Zimmermann, G. The ‘Galleria bait method’ for detection of entomopathogenic fungi in soil. J. Appl. Entomol. 1986, 102, 213–215. [Google Scholar] [CrossRef]
- Teetor-Barsch, G.H.; Roberts, D.W. Entomogenous Fusarium species. Mycopathologia 1983, 84, 3–16. [Google Scholar] [CrossRef] [PubMed]
- Mongkolsamrit, S.; Noisripoom, W.; Thanakitpipattana, D.; Wutikhun, T.; Spatafora, J.W.; Luangsa-ard, J. Disentangling cryptic species with isaria-like morphs in Cordycipitaceae. Mycologia 2018, 110, 230–257. [Google Scholar] [CrossRef]
- Liang, Y.-J.; Ariyawansa, H.A.; Becker, J.O.; Yang, J.-I. The evaluation of egg-parasitic fungi Paraboeremia taiwanensis and Samsoniella sp. for the biological control of Meloidogyne enterolobii on Chinese cabbage. Microorganisms 2020, 8, 828. [Google Scholar] [CrossRef]
- Islam, T.; Gupta, D.R.; Surovy, M.Z.; Mahmuc, N.U.; Mazlan, N.; Islam, T. Identification and application of a fungal biocontrol agent Cladosporium cladosporioides against Bemisia tabaci. Biotech. Biotechnol. Equip. 2019, 33, 1698–1705. [Google Scholar] [CrossRef]
- Liu, W.; Yu, S.-H.; Zhang, H.-P.; Fu, Z.-Y.; An, J.-Q.; Zhang, J.-Y.; Yang, P. Two Cladosporium fungi with opposite functions to the Chinese white wax scale insect have different genome characters. J. Fungi 2022, 8, 286. [Google Scholar] [CrossRef]
- Zhu, G.; Ding, W.; Xue, M.; Zhao, Y.; Li, M.; Li, Z. Identification and pathogenicity of a new entomopathogenic fungus, Mucor hiemalis (Mucorales: Mucorales), on the root maggot, Bradysia odoriphaga (Diptera: Sciaridae). J. Insect Sci. 2022, 22, 2. [Google Scholar] [CrossRef]
Figure 1.
(a) Akanthomyces muscarius: (a1) Infected L. delicatula adult. (a2) Conidia. (a3) Colony on potato dextrose agar (PDA) at 23 d, obverse. (a4) Colony on PDA at 24 d, reverse. (b) Clonostachys eriocamporesii: (b1) Infected L. delicatula adult. (b2) Conidia. (b3) Colony on PDA at 36 d, obverse. (b4) Colony on PDA at 36 d, reverse. (c) Clonostachys rosea: (c1) Infected L. delicatula third-instar nymph. (c2) Conidia. (c3) Colony on PDA at 24 d, obverse. (c4) Colony on PDA at 24 d, reverse. Cadaver scale bars: 3 mm. Conidial scale bars: 10 µm.
Figure 1.
(a) Akanthomyces muscarius: (a1) Infected L. delicatula adult. (a2) Conidia. (a3) Colony on potato dextrose agar (PDA) at 23 d, obverse. (a4) Colony on PDA at 24 d, reverse. (b) Clonostachys eriocamporesii: (b1) Infected L. delicatula adult. (b2) Conidia. (b3) Colony on PDA at 36 d, obverse. (b4) Colony on PDA at 36 d, reverse. (c) Clonostachys rosea: (c1) Infected L. delicatula third-instar nymph. (c2) Conidia. (c3) Colony on PDA at 24 d, obverse. (c4) Colony on PDA at 24 d, reverse. Cadaver scale bars: 3 mm. Conidial scale bars: 10 µm.

Figure 2.
(a) Colletotrichum fioriniae: (a1) Infected L. delicatula adult. (a2) Conidia. (a3) Colony on PDA at 12 d, obverse. (a4) Colony on PDA at 12 d, reverse. (b) Cordyceps cateniannulata: (b1) Infected L. delicatula adult. Arrows indicate two of the stromata. (b2) Conidia. (b3) Colony on PDA at 28 d, obverse. (b4) Colony on PDA at 28 d, reverse. (c) Cordyceps javanica: (c1) Infected L. delicatula adult. (c2) Conidia. (c3) Colony on PDA at 37 d, obverse. (c4) Colony on PDA at 37 d, reverse. Cadaver scale bars: 3 mm. Conidial scale bars: 10 µm.
Figure 2.
(a) Colletotrichum fioriniae: (a1) Infected L. delicatula adult. (a2) Conidia. (a3) Colony on PDA at 12 d, obverse. (a4) Colony on PDA at 12 d, reverse. (b) Cordyceps cateniannulata: (b1) Infected L. delicatula adult. Arrows indicate two of the stromata. (b2) Conidia. (b3) Colony on PDA at 28 d, obverse. (b4) Colony on PDA at 28 d, reverse. (c) Cordyceps javanica: (c1) Infected L. delicatula adult. (c2) Conidia. (c3) Colony on PDA at 37 d, obverse. (c4) Colony on PDA at 37 d, reverse. Cadaver scale bars: 3 mm. Conidial scale bars: 10 µm.
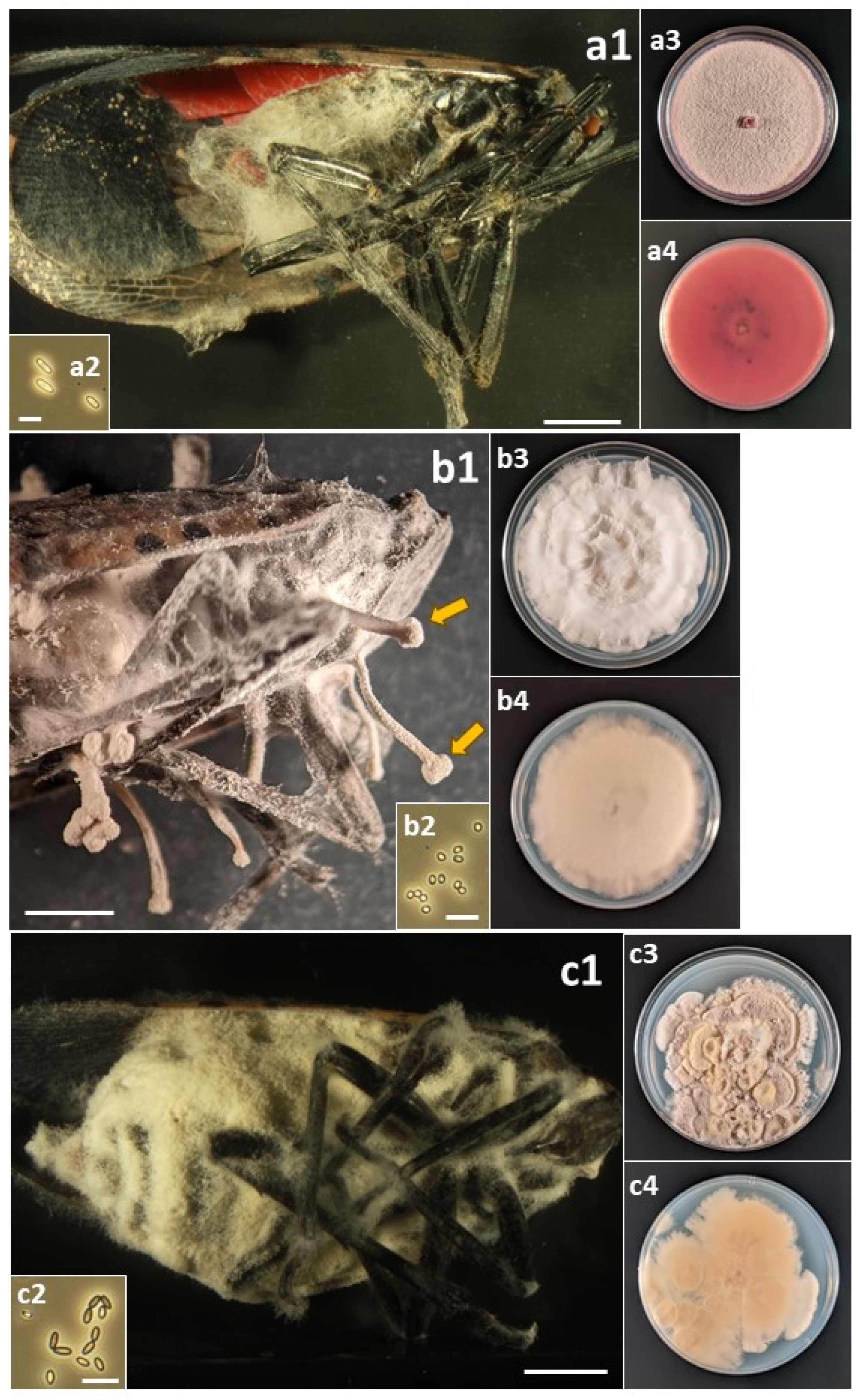
Figure 3.
(a) Flavocillium bifurcatum. (a1) Infected L. delicatula adult. (a2) Conidia. (a3) Colony on PDA at 12 d, obverse. (a4) Colony on PDA at 12 d, reverse. (b) Fusarium avenaceum: (b1) Infected L. delicatula adult. (b2) Macroconidia. (b3) Colony on PDA at 10 d, obverse. (b4) Colony on PDA at 10 d, reverse. (c) Fusarium concentricum: (c1) Infected L. delicatula fourth-instar nymph. (c2) Microconidia. (c3) Colony on PDA at 12 d, obverse. (c4) Colony on PDA at 12 d, reverse. Cadaver scale bars: 3 mm. Conidial scale bars: 10 µm.
Figure 3.
(a) Flavocillium bifurcatum. (a1) Infected L. delicatula adult. (a2) Conidia. (a3) Colony on PDA at 12 d, obverse. (a4) Colony on PDA at 12 d, reverse. (b) Fusarium avenaceum: (b1) Infected L. delicatula adult. (b2) Macroconidia. (b3) Colony on PDA at 10 d, obverse. (b4) Colony on PDA at 10 d, reverse. (c) Fusarium concentricum: (c1) Infected L. delicatula fourth-instar nymph. (c2) Microconidia. (c3) Colony on PDA at 12 d, obverse. (c4) Colony on PDA at 12 d, reverse. Cadaver scale bars: 3 mm. Conidial scale bars: 10 µm.
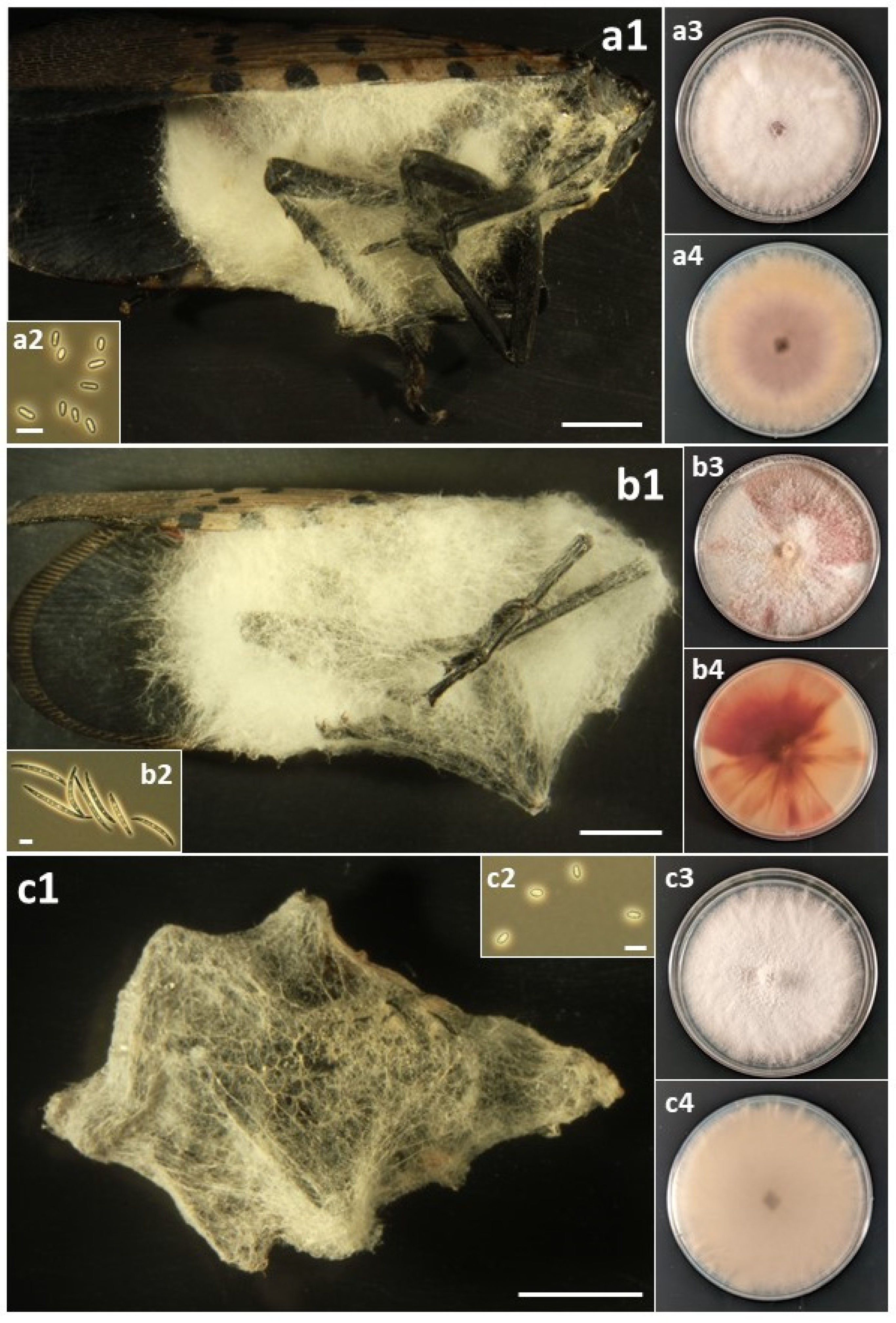
Figure 4.
(a) Fusarium falsibabinda: (a1) Infected L. delicatula adult. (a2) Microconidia. (a3) Colony on PDA at 12 d, obverse. (a4) Colony on PDA at 12 d, reverse. (b) Fusarium fujikuroi: (b1) Infected L. delicatula adult. (b2) Microconidia. (b3) Colony on PDA at 11 d, obverse. (b4) Colony on PDA at 11 d, reverse. (c) Fusarium graminearum: (c1) Infected L. delicatula adult. (c2) Macroconidia. (c3) Colony on PDA at 10 d, obverse. (c4) Colony on PDA at 10 d, reverse. Cadaver scale bars: 3 mm. Conidial scale bars: 10 µm.
Figure 4.
(a) Fusarium falsibabinda: (a1) Infected L. delicatula adult. (a2) Microconidia. (a3) Colony on PDA at 12 d, obverse. (a4) Colony on PDA at 12 d, reverse. (b) Fusarium fujikuroi: (b1) Infected L. delicatula adult. (b2) Microconidia. (b3) Colony on PDA at 11 d, obverse. (b4) Colony on PDA at 11 d, reverse. (c) Fusarium graminearum: (c1) Infected L. delicatula adult. (c2) Macroconidia. (c3) Colony on PDA at 10 d, obverse. (c4) Colony on PDA at 10 d, reverse. Cadaver scale bars: 3 mm. Conidial scale bars: 10 µm.
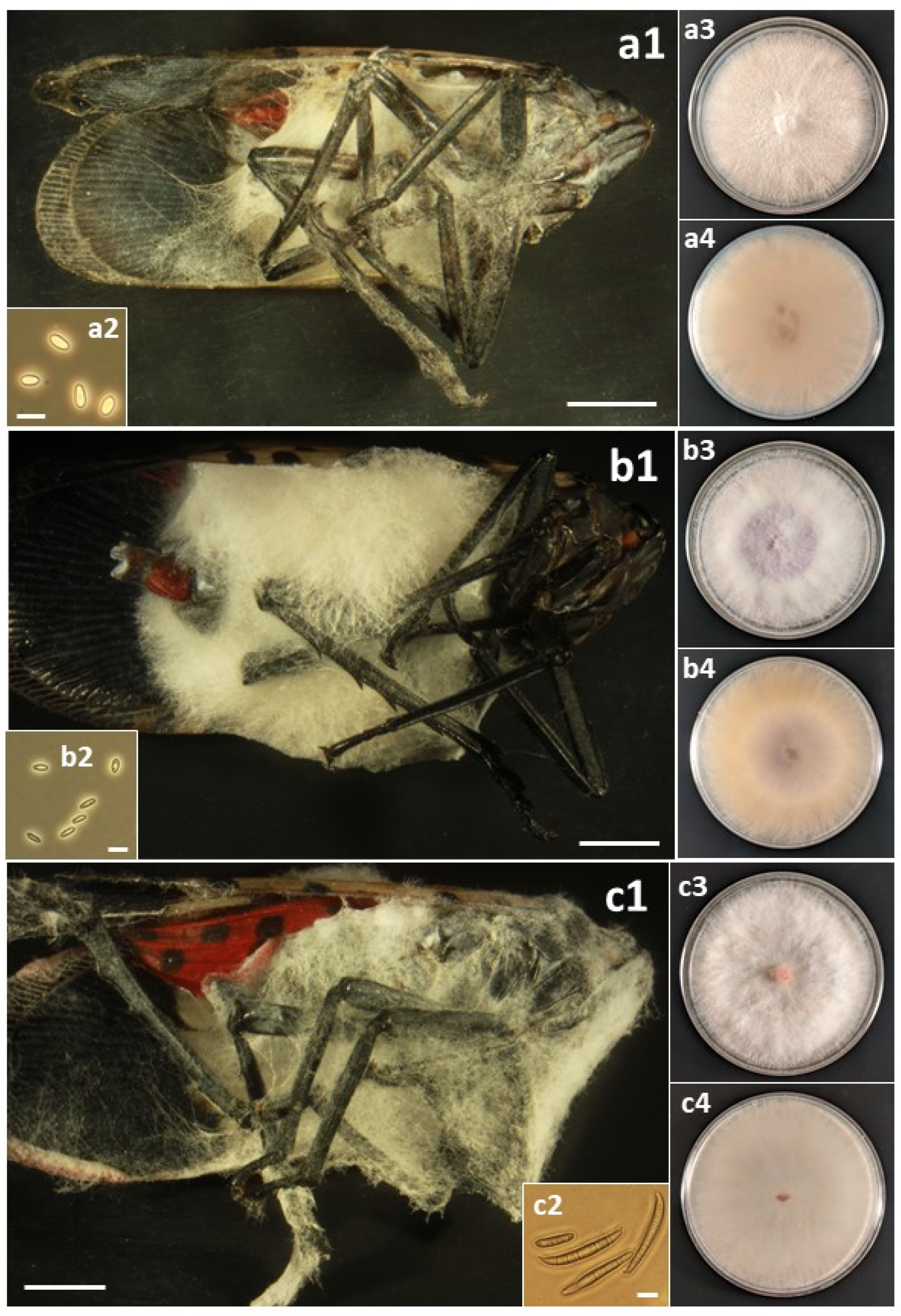
Figure 5.
(a) Samsoniella sp.: (a1) Infected L. delicatula adult. (a2) Conidia. (a3) Colony on PDA at 48 d, obverse. (a4) Colony on PDA at 48 d, reverse. (b) Sarocladium strictum: (b1) Infected L. delicatula fourth-instar nymph. (b2) Conidia. (b3) Colony on PDA at 48 d, obverse. (b4) Colony on PDA at 48 d, reverse. (c) Trichothecium roseum: (c1) Infected L. delicatula adult. (c2) Conidia. (c3) Colony on PDA at 10 d, obverse. (c4) Colony on PDA at 10 d, reverse. Cadaver scale bars: 3 mm. Conidial scale bars: 10 µm.
Figure 5.
(a) Samsoniella sp.: (a1) Infected L. delicatula adult. (a2) Conidia. (a3) Colony on PDA at 48 d, obverse. (a4) Colony on PDA at 48 d, reverse. (b) Sarocladium strictum: (b1) Infected L. delicatula fourth-instar nymph. (b2) Conidia. (b3) Colony on PDA at 48 d, obverse. (b4) Colony on PDA at 48 d, reverse. (c) Trichothecium roseum: (c1) Infected L. delicatula adult. (c2) Conidia. (c3) Colony on PDA at 10 d, obverse. (c4) Colony on PDA at 10 d, reverse. Cadaver scale bars: 3 mm. Conidial scale bars: 10 µm.

Figure 6.
Percent Lycorma delicatula collected in the field that were infected with the 14 fungal pathogens collected in 2022 by life stage/instar. Field-collected living nymphs and adults were reared on potted Ailanthus altissima under quarantine for 14 d after collection or were collected in the field as cadavers of recently dead L. delicatula. All fungal species (Table 3) except Cordyceps javanica and Colletotrichum fioriniae were collected in 2022.
Figure 6.
Percent Lycorma delicatula collected in the field that were infected with the 14 fungal pathogens collected in 2022 by life stage/instar. Field-collected living nymphs and adults were reared on potted Ailanthus altissima under quarantine for 14 d after collection or were collected in the field as cadavers of recently dead L. delicatula. All fungal species (Table 3) except Cordyceps javanica and Colletotrichum fioriniae were collected in 2022.

Figure 7.
Male versus female adults infected by B. bassiana versus opportunistic pathogens in 2022. ‘Opportunistic pathogens’ killing adults included A. muscarius, F. concentricum, F. falsibabinda, F. fujikuroi, F. graminearum, and S. strictum. Letters above bars demonstrate statistical differences between numbers of males versus females infected, separately by pathogen type.
Figure 7.
Male versus female adults infected by B. bassiana versus opportunistic pathogens in 2022. ‘Opportunistic pathogens’ killing adults included A. muscarius, F. concentricum, F. falsibabinda, F. fujikuroi, F. graminearum, and S. strictum. Letters above bars demonstrate statistical differences between numbers of males versus females infected, separately by pathogen type.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Sites for sampling Lycorma delicatula in Pennsylvania and New York, 2021–2023.
| Site | County | State | GPS | Year(s) Sampled |
|---|---|---|---|---|
| Angora Fruit Farm, Lower Alsace Twp. | Berks | Pennsylvania | 40.35846 N, 75.88323 W | 2021, 2022, 2023 |
| Penn Ave., Sinking Spring, Lower Heidelberg Twp. | Berks | Pennsylvania | 40.32700 N, 76.04683 W | 2022, 2023 |
| Glen Run Nature Preserve, Stroud Twp. | Monroe | Pennsylvania | 40.96950 N, 75.19004 W | 2021, 2022, 2023 |
| Minisink Park, Delaware Water Gap | Monroe | Pennsylvania | 40.98622 N, 75.13695 W | 2022 |
| Cherry Valley Rd., Stroud Twp. | Monroe | Pennsylvania | 40.97055 N, 75.17941 W | 2022 |
| Boys & Girls Club, Main St., Owego | Tioga | New York | 42.11008 N, 76.25717 W | 2023 |
Table 2.
Primers used for PCR and sequencing.
| Locus | Primer | Primer Sequence | Reference |
|---|---|---|---|
| ITS | ITS1 | TCCGTAGGTGAACCTGCGG | White et al., 1990 [29] |
| ITS1 mod | TCCGTAGGTGAACCTTGCGG | Korabečná et al., 2003 [38] | |
| ITS4 | TCCTCCGCTTATTGATATGC | White et al., 1990 [29] | |
| LSU | LR5 | ATCCTGAGGGAAACTTC | Vilgalys and Hester, 1990 [30] |
| LR0R 1 | ACCCGCTGAACTTAAGC | Lutzoni, 2014 [39] | |
| RPB1 | CRPB1 | CCWGGYTTYATCAAGAARGT | Castlebury et al., 2004 [31] |
| RPB1-Cr | CNGCDATNTCRTTRTCCATRTA | Matheny et al., 2002 [40] | |
| RPB2 | RPB2-5F | GAYGAYMGWGATCAYTTYGG | Liu et al., 1999 [34] |
| RPB2-5F2 | GGGGWGAYCAGAAGAAGGC | Sung et al., 2007 [32] | |
| RPB2-7cR | CCCATRGCTTGYTTRCCCAT | Liu et al., 1999 [34] | |
| TEF1-α | EF-1 | ATGGGTAAGGARGACAAGAC | O’Donnell et al., 1998 [41] |
| EF-2 | GGARGTACCAGTSATCATGTT | O’Donnell et al., 1998 [41] | |
| 983F | GCYCCYGGHCAYCGTGAYTTYAT | Rehner and Buckley, 2005 [35] | |
| 2218R | ATGACACCRACRGCRACRGTYTG | Rehner and Buckley, 2005 [35] | |
| TUB2 | T1 | AACATGCGTGAGATTGTAAGT | O’Donnell and Cigelnik, 1997 [42] |
| T2 | TAGTGACCCTTGGCCCAGTTG | O’Donnell and Cigelnik, 1997 [42] |
1 Referenced as (Vilgalys, unpublished).
Table 3.
Hypocrealean pathogens isolated from Lycorma delicatula (White, 1845).
| Kingdom Fungi | Functional Roles 1 | References for Roles |
|---|---|---|
| Division Ascomycota | ||
| Subdivision Pezizomycotina | ||
| Class Sordariomycetes | ||
| Order Hypocreales | ||
| Family Cordycipitaceae | ||
| Akanthomyces muscarius (Petch) Spatafora, Kepler and B. Shrestha, 2017 | EPF, MP | Saidi et al., 2023 [43]; Broumandnia and Rajabpour, 2021 [44] |
| Beauveria bassiana (Bals.-Criv.) Vuill., 1912 | EPF, PEP | Clifton et al., 2019, 2023 [24,45]; Hajek and Meyling, 2018 [46] |
| Cordyceps cateniannulata (Z.Q. Liang) Kepler, B. Shrestha, and Spatafora, 2017 | EPF | Montes-Bazurto et al., 2020 [47] |
| Cordyceps javanica (Bally) Kepler, B. Shrestha, and Spatafora, 2017 | EPF | Clifton and Hajek, 2021 [28] |
| Flavocillium bifurcatum H. Yu, Y.B. Wang, Y. Wang, Q. Fan, and Zhu L. Yang, 2020 | EPF | Wang et al., 2020 [48] |
| Samsoniella sp. | EPF | This study |
| Family Bionectriaceae | ||
| Clonostachys eriocamporesii R.H. Perera and K.D. Hyde, 2020 | EPF | Rodrigues et al., 2022 [49] |
| Clonostachys rosea (Link) Schroers, Samuels, Seifert, and W. Gams, 1999 | EPF, MP | Yang et al., 2021 [50]; Sun et al., 2020 [51] |
| Family Nectriaceae | ||
| Fusarium avenaceum (Fr.) Sacc., 1886 | EPF, PP, PEP | Batta, 2012 [52]; Rajagopal and Suryanarayanan, 2000 [53]; Uhlig et al., 2007 [54] |
| Fusarium concentricum Nirenberg and O’Donnell, 1998 | EPF, PP | Qiu et al., 2023 [55] |
| Fusarium falsibabinda M.M. Wang and L. Cai, 2022 | EPF | This study |
| Fusarium fujikuroi Nirenberg, 1976 | EPF, PP | Sharma et al., 2018 [56]; Dewing et al., 2022 [57] |
| Fusarium graminearum Schwabe, 1839 | EPF, PP, PEP | Ameen, 2012 [58]; Trail, 2009 [59]; Lofgren et al., 2018 [60] |
| Family Sarocladiaceae | ||
| Sarocladium strictum (W. Gams) Summerb., 2011 | EPF, PP, PEP, MP | El-Sayed et al., 2020 [61]; Blaszczyk et al., 2021 [62]; Tagne et al., 2002 [63]; Racedo et al., 2013 [64]; Kim 2002 [65]; This study |
| Family Glomerellaceae | ||
| Colletotrichum fioriniae (Marcelino and Gouli) Pennycook, 2017 | EPF, PP, PEP | Gonzalez et al., 2023 [66]; Marcelino et al., 2009 [67,68] |
| Family Myrotheciomycetaceae | ||
| Trichothecium roseum (Pers.) Link, 1809 | EPF, PP, MP | Batta, 2020 [69]; Götz and Karbowy-Thongbai, 2023 [70]; Zhu et al., 2022a [71] |
1 EPF = entomopathogenic fungi; PP = plant pathogen; PEP = plant endophyte; MP = mycoparasite. ‘This study’ indicates that this is the first report of this pathogen being an EPF.
Table 4.
Voucher and GenBank accession numbers for exemplar isolates of entomopathogenic species collected and identified in Pennsylvania and New York, from 2021–2023 1.
Table 4.
Voucher and GenBank accession numbers for exemplar isolates of entomopathogenic species collected and identified in Pennsylvania and New York, from 2021–2023 1.
| Fungal Species | ARSEF Number | ITS | LSU | RPB1 | RPB2 | TEF1-α | TUB2 |
|---|---|---|---|---|---|---|---|
| Akanthomyces muscarius | 14661 | OR577160 2 | OR575222 | OR593718 | OR593721 | OR593725 | |
| Clonostachys eriocamporesii | 14696 | OR582991 2 | OR602799 | ||||
| Clonostachys rosea | 14682 | OR577161 | OR593726 | ||||
| Colletotrichum fioriniae | 14695 | OR583017 | OR672136 | ||||
| Cordyceps cateniannulata | 14662 | OR577162 2 | OR575223 | OR593719 | OR593727 | ||
| Cordyceps javanica | 14690 | OR577169 | OR672137 | ||||
| Flavocillium bifurcatum | 14694 | OR582994 2 | OR577040 | OR602848 | OR602852 | OR602802 | |
| Fusarium avenaceum | 14691 | OR577163 | OR593728 | ||||
| Fusarium concentricum | 14687 | OR652385 | OR593729 | ||||
| Fusarium falsibabinda | 14667 | OR577164 2 | OR593730 | ||||
| Fusarium fujikuroi | 14677 | OR577165 2 | OR593722 | OR593731 | |||
| Fusarium graminearum | 14692 | OR577166 2 | OR593723 | OR593732 | |||
| Samsoniella sp. | 14689 | OR577167 | OR593720 | OR593724 | OR672131 | OR672138 | |
| Sarocladium strictum | 14693 | OR577168 2 | OR672132 | ||||
| Trichothecium roseum | 14697 | OR583015 2 | OR672134 |
Table 5.
Distribution of 16 entomopathogenic fungal species across sites and life stages, isolated from Lycorma delicatula in 2021–2023.
Table 5.
Distribution of 16 entomopathogenic fungal species across sites and life stages, isolated from Lycorma delicatula in 2021–2023.
| Fungal Species | # Sites with Fungus | Instars Infected | |||||||
|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | A1 | A2 | A3 | Total by Row | ||
| Akanthomyces muscarius | 3 | 2 | 1 | 1 | 5 | 9 | |||
| Beauveria bassiana | 5 | 1 | 1 | 4 | 32 | 50 | 88 | ||
| Clonostachys eriocamporesii | 1 | 1 | 1 | ||||||
| Clonostachys rosea | 2 | 1 | 2 | 3 | |||||
| Colletotrichum fioriniae | 1 | 1 | 1 | ||||||
| Cordyceps cateniannulata | 1 | 1 | 1 | ||||||
| Cordyceps javanica | 1 | 2 | 2 | ||||||
| Flavocillium bifurcatum | 1 | 1 | 1 | ||||||
| Fusarium avenaceum | 4 | 2 | 1 | 1 | 4 | ||||
| Fusarium concentricum | 3 | 1 | 2 | 1 | 4 | ||||
| Fusarium falsibabinda | 2 | 1 | 4 | 5 | |||||
| Fusarium fujikuroi | 6 | 1 | 6 | 14 | 9 | 30 | |||
| Fusarium graminearum | 3 | 3 | 2 | 5 | |||||
| Samsoniella sp. | 1 | 1 | 1 | ||||||
| Sarocladium strictum | 5 | 3 | 6 | 10 | 3 | 22 | |||
| Trichothecium roseum | 1 | 2 | 2 | ||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Hajek, A.E.; Everest, T.A.; Clifton, E.H. Accumulation of Fungal Pathogens Infecting the Invasive Spotted Lanternfly, Lycorma delicatula. Insects 2023, 14, 912. https://doi.org/10.3390/insects14120912
AMA Style
Hajek AE, Everest TA, Clifton EH. Accumulation of Fungal Pathogens Infecting the Invasive Spotted Lanternfly, Lycorma delicatula. Insects. 2023; 14(12):912. https://doi.org/10.3390/insects14120912
Chicago/Turabian StyleHajek, Ann E., Thomas A. Everest, and Eric H. Clifton. 2023. "Accumulation of Fungal Pathogens Infecting the Invasive Spotted Lanternfly, Lycorma delicatula" Insects 14, no. 12: 912. https://doi.org/10.3390/insects14120912
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.