
Factors Affecting Pollination and Pollinators in Oil Palm Plantations: A Review with an Emphasis on the Elaeidobius kamerunicus Weevil (Coleoptera: Curculionidae)
 , , and
, , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Oil Palm Pollination Agents
2.1. Biology of Elaeidobius kamerunicus
2.2. Roles of VOCs in Oil Palm Pollination
2.3. Key Characteristics of E. kamerunicus as the Main Pollinator
2.3.1. Adapted Mouthparts
2.3.2. Complementarity with Native Pollinators
2.3.3. High Pollen Carrying Capacity
2.3.4. Ability to Select High-Quality Pollen
2.4. Pollinator Health
2.4.1. Endogamy of Weevils and Genetic Bottleneck
2.4.2. Nematodes Parasitism
2.4.3. Predatory Threats
2.4.4. Use of Pesticides
3. Inadequate Pollen Deposition and Pollen Quality
4. Habitat Loss
5. Proximity of Oil Palms to Natural Forests
6. Climate and Weather Driver
7. Conclusions and Recommendations
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- F.A.O. of the U.N. (FAO). Why Bees Matter. Available online: https://www.fao.org/3/I9527EN/i9527en.PDF (accessed on 20 February 2023).
- Butler, R. Bees and Other Pollinators Are in Danger of Disappearing for Good. Natl. Geogr. 2019. Available online: https://www.nationalgeographic.com/environment/2019/06/bees-other-pollinators-disappearing/ (accessed on 20 June 2019).
- O.R.S. The Buzz About Colony Collapse Disorder: Can Science Save Bees? Sci. Am. 2021. Available online: https://www.scientificamerican.com/article/the-buzz-about-colony-collapse-disorder-can-science-save-bees/ (accessed on 1 January 2021).
- Reis, E.; Silva-Mann, R.; Damião, M.; de Souza, E.; Franco, D.F. Pollen Quality and Quantity Effect on Fruit Set and Fruit Quality in Coffee Trees. J. Plant Growth Regul. 2020, 39, 919–928. Available online: http://journal.um-surabaya.ac.id/index.php/JKM/article/view/2203 (accessed on 22 December 2022).
- Swaray, S.; Amirudin, M.; Rafii, M.; Jamian, S.; Ismail, M.; Jalloh, M.; Eswa, M.; Marjuni, M.; Akos, I.; Yusuff, O. Oil palm inflorescence sex ratio and fruit set assessment in dura × pisifera biparental progenies on fibric peat soil. Agronomy 2021, 11, 1380. [Google Scholar] [CrossRef]
- Kushairi, A.; Ong-Abdullah, M.; Nambiappan, B.; Hishamuddin, E.; Bidin, M.; Ghazali, R.; Subramaniam, V.; Sundram, S.; Parveez, G. Oil palm economic performance in Malaysia and R&D progress in 2018. J. Oil Palm Res. 2019, 31, 165–194. [Google Scholar]
- Hartley, C. Oil palm breeding: Genetics and genomics. Ann. Appl. Biol. 2003, 142, 107–120. [Google Scholar] [CrossRef]
- Hardy, O.J.; Fernandez, E.; Nordine, O. Elaeis guineensis (oil palm). In Fruit Trees and Oil-Producing Trees; Springer: Berlin/Heidelberg, Germany, 2020; pp. 17–27. [Google Scholar]
- Rajamohanan, P.R.; Kushairi, A.; Chee, S.; Ong-Abdullah, M.; Leslie, C.; Singh, R. Effect of pollination on oil palm yield. J. Oil Palm Res. 2017, 29, 220–227. [Google Scholar]
- Klein, A.M.; Vaissie, B.; Cane, J.; Steffan-Dewenter, I.; Cunningham, S.; Kremen, C.; Tscharntke, T. Importance of pollinators in changing landscapes for world crops. Proc. R. Soc. B Biol. Sci. 2007, 274, 303–313. [Google Scholar] [CrossRef]
- Prasetyo, A.E.; Purba, W.; Susanto, A. Elaeidobius kamerunicus: Application of hatch and carry technique for increasing oil palm fruit set. J. Oil Palm Res. 2014, 26, 195–202. [Google Scholar]
- Zulkefli, M.H.H.; Jamian, S.; Adam, N.; Jalinas, J.; Mohamad, S.; Masri, M.M.M. Beyond four decades of Elaeidobius kamerunicus Faust (Coleoptera: Curculionidae) in the Malaysian oil palm industry: A review. J. Trop. Ecol. 2020, 36, 282–292. [Google Scholar] [CrossRef]
- Wahyuni, M.; Wagino; Manurung, H. The Effect of Palm Oil Planting Material Characteristics on the Population of Weevil Pollinator (Elaeidobius kamerunicus). J. Phys. Conf. Ser. 2019, 1542, 12073. [Google Scholar] [CrossRef]
- Harun, M.; Noor, M.R.M. Fruit Set and Oil Palm Bunch Components. J. Oil Palm Res. 2002, 14, 24–33. [Google Scholar]
- Dhileepan, K.; Nampoothiri, K.U.K. Pollination potential of the introduced weevil Elaeidobius kamerunicus Faust in the oil palm (Elaeis guineensis Jacq.) plantations. Indian J. Agric. Sci. 1989, 59, 517–521. [Google Scholar]
- Ruiz-Alvarez, E.; Daza, E.; Caballero-Blanco, K.; Mosquera-Montoya, M. Complementing assisted pollination with artificial pollination in oil palm crops planted with interspecific hybrids O×G (Elaeis guineensis × Elaeis oleifera): Is it profitable? OCL 2021, 28, 27. [Google Scholar] [CrossRef]
- Appiah, S.O.; Agyei-Dwarko, D. Studies on Entomophil pollination towards Sustainable Production and Increased Profitability in the Oil Palm: A Review. Elixir Agric. 2013, 55, 56126660. [Google Scholar]
- Kouakou, M.; Hala, N.; Akpesse, A.; Tuo, Y.; Dagnogo, M.; Konan, K.; Koua, H.K. Comparative Efficacy of Elaeidobius kamerunicus, E. plagiatus, E. subvittatus (Coleoptera: Curculionidae) and Microporum spp. (Coleoptera: Nitidulidae) in the Pollination of Oil Palm (Elaeis guineensis). J. Exp. Biol. Agric. Sci. 2014, 2, 538–545. Available online: http://www.jebas.org (accessed on 4 February 2021).
- Meléndez, M.R.; Ponce, W.P. Pollination in the oil palms Elaeis guineensis, E. oleifera and their hybrids (O×G), in tropical America. Pesqui. Agropecuária Trop. 2016, 46, 102–110. [Google Scholar] [CrossRef]
- Auffray, T.; Frérot, B.; Poveda, R.; Louise, C.; Beaudoin-Ollivier, L. Diel patterns of activity for insect pollinators of two oil palm species (Arecales: Arecaceae). J. Insect Sci. 2017, 17, 4–9. [Google Scholar] [CrossRef]
- Tambunan, V.B.; Apriyanto, A.; Ajambang, W.; Etta, C.; Sahari, B.; Damayanti, B.; Hidayat, P. Molecular identification and population genetic study of Elaeidobius kamerunicus Faust. (Coleoptera: Curculionidae) from Indonesia, Malaysia and Cameroon based on mitochondrial gene. Biodiversitas 2020, 21, 3263–3270. [Google Scholar] [CrossRef]
- Syed, R.A. The latest developments of Elaeidobius’s role in pollination: Feasibility of introducing another species. In Proceedings of the Symposium on Impact of Pollinating Weevil on the Malaysian Oil Palm Industry, Kuala Lumpur, Malaysia, 21–22 February 1984; pp. 58–81. [Google Scholar]
- Labarca, M.V.; Narváez, Z. Identification and pollinating insect’s population fluctuation in oil palm (Elaeis guineensis Jacquin) in south of Maracaibo Lake, Zulia state, Venezuela. Rev. Fac. Agron. Univ. Zulia 2009, 26, 305–324. [Google Scholar]
- Sambathkumar, S.; Ranjith, A.M. Insect pollinators of oil palm in Kerala with special reference to African weevil, Elaeidobius kamerunicus Faust. Pest Manag. Hortic. Ecosyst. 2011, 17, 14–18. [Google Scholar]
- Hala, N.; Tuo, Y.; HK, A.K.; Tano, Y. Entomofauna of oil palm tree inflorescences at La Mé experimental station (Côte d’Ivoire). Am. J. Exp. Agric. 2012, 2, 306–319. [Google Scholar]
- De Chenon, R.D. Entomophile pollination of oil palm in West Africa. Preliminary research. In The Oil Palm in Agriculture in the Eighties; Incorporated Society of Planters: Petaling Jaya, Malaysia, 1981; pp. 239–291. [Google Scholar]
- Li, K.; Tscharntke, T.; Saintes, B.; Buchori, D.; Grass, I. Critical factors limiting pollination success in oil palm: A systematic review. Agric. Ecosyst. Environ. 2019, 280, 152–160. [Google Scholar] [CrossRef]
- Mohamad, S.A.; Masri, M.M.; Kamarudin, N.; Sulaiman, M.; Costa, A.; Ong-Abdullah, M.; Othman, H.; Ahmad, S.; Syarif, M.; Karim, Z.; et al. Impact of Elaeidobius kamerunicus (Faust) Introduction on Oil Palm Fruit Formation in Malaysia and Factors Affecting its Pollination Efficiency: A Review. J. Oil Palm Res. 2022, 35, 1–12. [Google Scholar] [CrossRef]
- Tan, Y.P.; Basri, M.W. Another pollinating insect of oil palm—Pyroderces spp. In Proceedings of the Seminar on the Impact of the Pollinating Weevil on the Malaysian Industry, Kuala Lumpur, Malaysia, 21–24 February 1984; Palm Oil Research Institute of Malaysia: Kuala Lumpur, Malaysia, 1985; pp. 50–57. [Google Scholar]
- Corley, R.H.V.; Tinker, P.B. The Oil Palm, 4th ed.; John Wiley & Sons: Hoboken, NJ, USA, 2016; pp. 495–506. [Google Scholar]
- Anggraeni, T.; Rahayu, S.; Ahmad, I.; Esyanti, R.; Putra, R.E. Resources partitioning and different foraging behavior is the basis for the coexistence of Thrips hawaiiensis (Thysanoptera: Tripidae) and Elaeidobius kamerunicus (Coleoptera: Curculionidae) on oil palm (Elaeis guineensis Jacq) flower. J. Entomol. Nematol. 2013, 5, 59–63. [Google Scholar] [CrossRef]
- Mohamad, S.A.; Ahmad, S.N.; Syarif, M.N.Y.; Sulaiman, M.R.; Hung, K.J.; Mohammed, M.A.; Masri, M.M.M. Population Abundance of Oil Palm Pollinating Weevil, Elaeidobius kamerunicus Faust and its Relation to Fruit Set Formation in Mineral and Peat Soil Areas in Peninsular Malaysia. Serangga 2021, 26, 107–117. [Google Scholar]
- Teo, T.M. Effectiveness of the oil palm pollinating weevil, Elaeidobius kamerunicus, in Malaysia. Utar Agric. Sci. J. 2015, 1, 40–43. [Google Scholar]
- Wahid, M.B.; Kamarudin, N.H. Role and effectiveness of Elaeidobius kamerunicus, Thrips hawaiiensis and Pyroderces sp. in pollination of mature oil palm in Peninsular Malaysia. Elaeis 1997, 9, 1–16. [Google Scholar]
- Aminah, H.; Salim, M.; Abdul-Razak, M. Feeding behavior and host specificity of Elaeidobius kamerunicus (Coleoptera: Curculionidae), pollinator of oil palm flowers. J. Econ. Entomol. 1998, 91, 140–147. [Google Scholar]
- Kartika, J.G.; Hidayat, P.; Wibowo, A.; Suryobroto, B. Oviposition preference of Elaeidobius kamerunicus for pollinating oil palm flowers in a laboratory experiment. Hayati J. Biosci. 2018, 25, 74–79. [Google Scholar]
- Ong, K.H.; Norhadi, I.; Kushairi, A. Life cycle of the oil palm pollinating weevil, Elaeidobius kamerunicus in the laboratory. J. Oil Palm Res. 2016, 28, 352–360. [Google Scholar]
- Rajamohan, A.; Mastura, M.; Wong, Y.; Wahizatul, A.A. Pollination and breeding behavior of the oil palm pollinating weevil, Elaeidobius kamerunicus. J. Oil Palm Res. 2015, 27, 41–50. [Google Scholar]
- Tuo, Y.; Koua, H.; Hala, N. Biology of Elaeidobius kamerunicus and Elaeidobius plagiatus (Coleoptera: Curculionidae) main pollinators of oil palm in west Africa. Eur. J. Sci. Res. 2011, 49, 426–432. [Google Scholar]
- Siswanto, D.S. Population of oil palm pollinator insect (Elaeidobius kamerunicus Faust.) at PTP Nusantara VIII Cisalak Baru, Rangkasbitung-Banten. IOP Conf. Ser. Earth Environ. Sci. 2020, 418, 012045. [Google Scholar] [CrossRef]
- Girsang, R.J.; Tobing, M.; Pangestuningsih, Y. Biology of Insect Pollinator Elaeidobius kamerunicus (Coleoptera: Curculionidae) after 33 Years being Introduced in North Sumatera. J. Agroekoteknologi FP USU 2017, 5, 348–354. [Google Scholar]
- Haran, J.M.; Beaudoin-Ollivier, L.; Benoit, L.; Kuschel, G. Revision of the palm-pollinating weevil genus Elaeidobius Kuschel, 1952 (Curculionidae, Curculioninae, Derelomini) with descriptions of two new species. Eur. J. Taxon. 2020, 2020, 1–32. [Google Scholar] [CrossRef]
- Fahmi-Halil, M.; Haris-Hussain, M.; Mirad, R.; Ab, I.; Jalinas, J. Responses of Oil Palm Pollinator, Elaeidobius kamerunicus to Different Concentrations of Estragoles 2 3. 2021. Available online: https://www.biorxiv.org/content/10.1101/2021.03.24.436726v1 (accessed on 6 December 2022). [CrossRef]
- Holopainen, J.K.; Gershenzon, J. Multiple stress factors and the emission of plant VOCs. Trends Plant Sci. 2010, 15, 176–184. [Google Scholar] [CrossRef]
- Yousefi, M.; Rafie, A.M.; Aziz, S.A.; Azrad, S.; Razak, A.A. Introduction of current pollination techniques and factors affecting pollination effectiveness by Elaeidobius kamerunicus in oil palm plantations on regional and global scale: A review. S. Afr. J. Bot. 2020, 132, 171–179. [Google Scholar] [CrossRef]
- Barrett, S.C.H.; Hough, J. Sexual Dimorphism in Flowering Plants. J. Exp. Bot. 2013, 64, 67–82. [Google Scholar] [CrossRef]
- Fahmi, M.H.M.; Bukhary, A.A.; Norma, H.; Idris, A.B. Analysis of volatile organic compound from Elaeis guineensis inflorescences planted on different soil types in Malaysia. AIP Conf. Proc. 2016, 1784, 060020. [Google Scholar] [CrossRef]
- Ashman, T.-L. Sniffing out patterns of sexual dimorphism in floral scent. Funct. Ecol. 2009, 23, 852–862. [Google Scholar] [CrossRef]
- Raja, M.A.; Uma, S.; Vikram, P.; Ananthakrishnan, T.N. A molecular phylogenetic assessment of the Elaeidobius kamerunicus pollinator weevils (Coleoptera: Curculionidae) from India, Indonesia and Africa. J. Genet. 2016, 95, 481–487. [Google Scholar]
- Lodhi, G.A.; Ahmad, S. Host specificity of the pollinating weevil, Elaedobius kamerunicus (Coleoptera: Curculionidae), of the oil palm. Bull. Entomol. Res. 1976, 66, 247–258. [Google Scholar]
- Nur, S.F.H.; Sulaiman, Z.; Zakaria, R. Characterization of volatiles released by oil palm flowers and selection of an optimal lure for Elaeidobius kamerunicus (Coleoptera: Curculionidae) using behavioral assays. Environ. Entomol. 2017, 46, 707–714. [Google Scholar]
- Yue, J.; Yan, Z.; Bai, C.; Chen, Z.; Lin, W.; Jiao, F. Pollination Activity of Elaeidobius kamerunicus (Coleoptera: Curculionoidea) on Oil Palm on Hainan Island. Fla. Entomol. 2015, 98, 499–505. [Google Scholar] [CrossRef]
- Asib, N.; Musli, N.N. Effect of six insecticides on oil palm pollinating weevil, Elaeidobius kamerunicus (Coleoptera: Curculionidae). Serangga 2020, 25, 1–9. [Google Scholar]
- Egonyu, J.P.; Stephen, E.; Baguma, J.; Otema, M.; Ddamulira, G. Insect flower-visitors of African oil palm Elaeis guineensis at different sites and distances from natural vegetation in Uganda. Int. J. Trop. Insect Sci. 2021, 41, 2477–2487. [Google Scholar] [CrossRef]
- Tambunan, V.B.; Sahari, B.; Buchori, D.; Hidayat, P. Pollen load and distribution on the body of Elaeidobius kamerunicus Faust. (Coleoptera: Curculionidae) within oil palm plantations. J. Entomol. Indones. 2020, 17, 81. [Google Scholar] [CrossRef]
- Dhileepan, K. Pollen carrying capacity, pollen load and pollen transferring ability of the oil palm pollinating weevil Elaeidobius kamerunicus Faust in India. Oléagineux 1992, 47, 55–61. [Google Scholar]
- Fatihah, A.L.N.; Fahmi, M.M.; Luqman, H.; Nadiah, S.S.; Teo, T.; Riza, H.I.; Idris, A.B. Effects of Rainfall, Number of Male Inflorescences and Spikelets on the Population Abundance of Elaeidobius kamerunicus (Coleoptera: Curculionidae). Sains Malays. 2019, 48, 15–21. [Google Scholar] [CrossRef]
- Caudwell, R.W.; Reid, A.; Mensah, B.; Chinchilla, C. Insect pollination of oil palm-a comparison of the long term viability and sustainability of Elaeidobious kamerunicus in Papua New Guinea, Indonesia, Costa Rica, and Ghana. ASD Oil Palm Pap. 2003, 25, 1–16. [Google Scholar]
- Kuriwada, T.; Kumano, N.; Shiromoto, K.; Haraguchi, D. The effect of inbreeding on mating behaviour of West Indian sweet potato weevil Euscepes postfasciatus. Ethology 2011, 117, 822–828. [Google Scholar] [CrossRef]
- Facon, B.; Hufbauer, R.; Tayeh, A.; Loiseau, A.; Lombaert, E.; Vitalis, R.; Guillemaud, T.; Lundgren, J.; Estoup, A. Inbreeding depression is purged in the invasive insect Harmonia axyridis. Curr. Biol. 2011, 21, 424–427. [Google Scholar] [CrossRef] [PubMed]
- Müller, T.; Lamprecht, T.; Schrieber, K. Lifetime inbreeding depression in a leaf beetle. Ecol. Evol. 2018, 8, 6889–6898. [Google Scholar] [CrossRef] [PubMed]
- Vayssade, C.; De Fazio, C.; Quaglietti, B.; Auguste, A.; Ris, N.; Fauvergue, X. Inbreeding depression in a parasitoid wasp with single-locus complementary sex determination. PLoS ONE 2014, 9, e97733. [Google Scholar] [CrossRef] [PubMed]
- Latip, N.F.A.; Ghani, I.; Hazmi, I.; Abidin, C.; Sartiami, D.; Yacob, S.; Nasir, D.M. Morphometric Comparison of the Oil Palm Pollinator Elaeidobius kamerunicus Faust (Coleoptera: Curculionidae) from Malaysia, Indonesia, and Liberia. Coleopt. Bull. 2019, 73, 746–756. [Google Scholar] [CrossRef]
- Nasir, D.M.; Mamat, N.; Muneim, N.A.; Ong-Abdullah, M.; Latip, N.A.; Su, S.; Hazmi, I.R. Morphometric Analysis of the Oil Palm Pollinating Weevil, Elaeidobius kamerunicus (Faust, 1878) (Coleoptera: Curculionidae) from Oil Palm Plantations in Malaysia. J. Entomol. Res. Soc. 2020, 22, 275–291. [Google Scholar] [CrossRef]
- Haran, J.; Abanda, R.N.; Benoit, L.; Bakoumé, C.; Beaudoin-Ollivier, L. Multilocus phylogeography of the world populations of Elaeidobius kamerunicus (Coleoptera, Curculionidae), pollinator of the palm Elaeis guineensis. Bull. Entomol. Res. 2020, 110, 654–662. [Google Scholar] [CrossRef]
- Mohamad, R.; Ahmad, A.; Zulkifli, N.; Rosli, M.A. Genetic diversity and structure of the oil palm pollinator Elaeidobius kamerunicus (Coleoptera: Curculionidae) in Malaysia. Sci. Rep. 2019, 9, 1–10. [Google Scholar]
- Baladouni, Y.; Kekeunou, S.; Tindo, M.; Tchoumboué, J. Genetic diversity of the oil palm pest Elaeidobius kamerunicus (Coleoptera, Curculionidae) in Cameroon. J. Appl. Entomol. 2015, 139, 520–529. [Google Scholar] [CrossRef]
- Bakara, R.D.J.; Buchori, D.; Kusumah, R.; Sahari, B.; Apriyanto, A. Genetic diversity of Elaeidobius kamerunicus Faust. (Coleoptera: Curculionidae) across geographic areas in Indonesia: Implications for oil palm sustainability. IOP Conf. Ser. Earth Environ. Sci. 2019, 325, 12003. [Google Scholar] [CrossRef]
- Fernandez-Conradi, P.; Smith, C.; Girling, R.; Staley, J.T. Effects of parasitic infection on the fitness of the oil palm weevil, Eleschus kamerunicus. Entomol. Exp. Appl. 2016, 161, 221–229. [Google Scholar]
- Laumann, R.A.; Guimarães, J.H. Nematóides parasitas de Rhynchophorus palmarum L. (Coleoptera: Curculionidae). An. Soc. Entomológica Bras. 1968, 37, 391–401. [Google Scholar]
- Sugau, J.B.; Potts, S.; Wright, D.J. Eleschus kamerunicus Faust (Coleoptera: Curculionidae) as a vector of the nematode Cosmocercoides sp. in oil palm plantations in Sabah, Malaysia. J. Oil Palm Res. 2011, 23, 100–107. [Google Scholar]
- Campos-Herrera, R. Nematode parasites of insects: Phylogeny, evolution and ecophysiology. In Insect Pathology; Academic Press: Cambridge, MA, USA, 2015; pp. 107–139. [Google Scholar]
- Husain, N.H.N.; Ghani, I.; Ramle, M.; Daud, S.K. Identification and screening of oil palm (Elaeis guineensis) for resistance against basal stem rot in Malaysia. J. Oil Palm Res. 2017, 29, 528–535. [Google Scholar]
- Jaffuel, G.; Hiltpold, I.; Turlings, T.; Newsham, K.K. The role of soil-dwelling insects and nematodes in ecosystem functioning. J. Ecol. 2019, 107, 2496–2510. [Google Scholar]
- Lee, H.L.; Mohd-Azmi, M.; Lim, G.; Yusuf, S.; Salleh, F.; Md-Zain, B.; Yaakop, S. Entomopathogenic nematodes, Heterorhabditis indica and Steinernema scapterisci reduce populations of Elaeidobius kamerunicus (Coleoptera: Curculionidae) in oil palm plantations. J. Appl. Entomol. 2018, 142, 87–96. [Google Scholar] [CrossRef]
- Swaray, S.; Rafii, M.; Amiruddin, M.; Ismail, M.; Jamian, S.; Jalloh, M.; Oladosu, Y.; Mohamad, M.; Marjuni, M.; Kolapo, O.; et al. Assessment of Oil Palm Pollinating Weevil (Elaeidobius kamerunicus) Population Density in Biparental dura × pisifera Hybrids on Deep Peat-Soil in Perak State, Malaysia. Insects 2021, 12, 221. [Google Scholar] [CrossRef]
- Mazza, G.; Francardi, V.; Simoni, S.; Benvenuti, C.; Cervo, R.; Faleiro, J.; Llacer, E.; Longo, S.; Nanneli, R.; Tarasco, E.; et al. An overview on the natural enemies of Rhynchophorus palm weevils, with focus on R. ferrugineus. Biol. Control 2014, 77, 83–92. [Google Scholar] [CrossRef]
- Luqman, H.A.M.; Dzulhelmi, M.; Idris, A.; Riza, H.I. The potential natural predators of Elaeidobius kamerunicus Faust, 1878 (coleoptera: Curculionidae) in Malaysia. Serangga 2017, 22, 239–252. [Google Scholar]
- De Chenon, R.D.; Agus, S. Ecological observations on diurnal birds in Indonesian oil palm plantations. J. Oil Palm Res. 2006, 18, 122–143. [Google Scholar]
- Van der Sluijs, J.P.; Simon-Delso, N.; Goulson, D.; Maxim, L.; Bonmatin, J.; Belzunces, L.P. A behavioral perspective on the governance of common resources. Curr. Opin. Environ. Sustain. 2015, 12, 1–5. [Google Scholar] [CrossRef]
- Syari, J.; Ahmad, N.; Amal, G.; Azlina, Z.; Badrul, A. Impacts of 2 species of predatory Reduviidae on bagworms in oil palm plantations. Insect Sci. 2016, 24, 285–294. [Google Scholar]
- Su, Y.J.; Chen, C.; Tseng, Y.; Hwang, S.Y. Susceptibility of Elaeidobius kamerunicus and Oryctes rhinoceros to chlorantraniliprole and cypermethrin insecticides. J. Asia. Pac. Entomol. 2017, 20, 1195–1200. [Google Scholar] [CrossRef]
- Ismail, N.F.; Ghani, I.; Othman, N.W. Detrimental effects of commonly used insecticides in oil palm to pollinating weevil, Elaeidobius kamerunicus Faust. (Coleoptera: Curculionidae). J. Oil Palm Res. 2020, 32, 439–452. [Google Scholar] [CrossRef]
- Sulistyawati, E.; Kuswadi, A.; Widiastuti, R.; Syahputra, E. Differential Susceptibility of Male and Female Elaeidobius kamerunicus to Insecticides. Hayati J. Biosci. 2020, 27, 141–146. Available online: http://journal.um-surabaya.ac.id/index.php/JKM/article/view/22030 (accessed on 21 December 2022).
- Ooi, L.H.; Sambanthamurthi, R.; Tan, Y.H. Pollen viability and in vitro pollen germination of oil palm (Elaeis guineensis Jacq.). J. Oil Palm Res. 2010, 22, 762–771. [Google Scholar]
- Kushairi, A.; Henson, I.; Rajanaidu, N. Pollination in oil palm (Elaeis guineensis Jacq.). J. Oil Palm Res. 2012, 24, 1325–1340. [Google Scholar]
- Tarmizi, A.H.A.; Yusof, M.; Salleh, S.; Fadzil, A.A. The Relationship between Oil Palm Pollen Quantity and Fruit Set. J. Oil Palm Res. 2014, 26, 49–58. Available online: https://eje.bioscientifica.com/view/journals/eje/171/6/727.xml0 (accessed on 23 December 2022).
- Haniff, H.A.; Yusoff, M.; Tan, C.; Arif, S.; Zamzuri, I. Effect of pollination on the yield and quality of oil palm fruit. J. Oil Palm Res. 2016, 28, 18–29. [Google Scholar]
- Woittiez, L.S.; van Wijk, M.; Slingerland, M.; van Noordwijk, M.; Giller, K.E. Yield gaps in oil palm: A quantitative review of contributing factors. Eur. J. Agron. 2017, 83, 57–77. [Google Scholar] [CrossRef]
- Kushairi, A.; Rajanaidu, N. Breeding for high oil quality and yield in oil palm. In Oil Palm Breeding; Springer: Dordrecht, The Netherlands, 2001; pp. 159–170. [Google Scholar]
- Jalani, B.S.; Wickneswari, R.; Rajanaidu, N. Yield performance of Pisifera palms in the Malaysian oil palm breeding programme. Planter 1997, 73, 95–101. [Google Scholar]
- Mohamad, S.A.; Syarif, M.; Ahmad, S.; Masri, M.; King, J.; Kamarudin, N. Population Density of Elaeidobius kamerunicus Faust in Different Spikelet Position at Anthesising Male Inflorescence of Elaeis guineensis Jacq. in Sabah And Sarawak, Malaysia. J. Oil Palm Res. 2021, 33, 21–36. [Google Scholar] [CrossRef]
- Rajanaidu, N.; Kushairi, A. Tenera breeding and selection for high yield in Malaysia. In Oil Palm Breeding; Springer: Dordrecht, The Netherlands, 2001; pp. 189–197. [Google Scholar]
- Singh, R.; Harikrishna, K.; Kushairi, A. Hybrid oil palm development in Malaysia. In Oil Palm Genetic Improvement; Springer: Dordrecht, The Netherlands, 2005; pp. 219–244. [Google Scholar]
- Jalani, B.S.; Rajanaidu, N. Reproductive development and yield of Tenera oil palms. Planter 1991, 67, 513–524. [Google Scholar]
- Rajanaidu, N.; Kushairi, A. Oil palm breeding—Achievements and potential. In Proceedings of the International Symposium on Recent Developments in Oil Palm Tissue Culture and Biotechnology, Kuala Lumpur, Malaysia, 24–25 September 2002; pp. 117–140. [Google Scholar]
- MPOB. Insect Pollination Efficiency in Oil Palm. 2018. Available online: https://scholar.google.com/scholar_lookup?title=Insect+pollination+efficiency+in+oil+palm&author=MPOB&publication_year=2018 (accessed on 24 April 2019).
- ISOPA. Current Status of Oil Palm Fruit Set in Malaysia; ISOPA Mini Seminar; Kuala Lumpur Convention Centre: Kuala Lumpur, Malaysia, 2019. [Google Scholar]
- Noh, A.; Rafii, M.; Saleh, G.; Kushairi, A. Genetic performance of 40 Deli dura × AVROS pisifera full-sib families. J. Oil Palm Res. 2010, 22, 781–795. [Google Scholar]
- Norman, K.; Ramle, M.; Saharul, A.; Mohd, R.S. Fruit set and weevil pollination issues in oil palm. Planter 2018, 94, 565–578. [Google Scholar]
- Corley, H.R.V.; Gray, B.S. Growth and Morphology. In Oil Palm Research; Corley, R.H.V., Hardon, J.J., Wood, B.J., Eds.; Elsevier: Amsterdam, The Netherlands, 1976; pp. 7–21. [Google Scholar]
- Donough, C.R.; Chew, K.; Law, I.H. Effect of Fruit Set on OER and KER: Results from Studies at Pamol Estates (Sabah) Sdn Bhd. Planter 1996, 72, 203–219. [Google Scholar]
- Potts, S.G.; Imperatriz-Fonseca, V.; Ngo, H.; Aizen, M.; Biesmeijer, J.; Breeze, T.; Garibaldi, L.; Hill, R.; Setele, J.; Vanbergen, A.J. Safeguarding pollinators and their values to human well-being. Nature 2016, 540, 220–229. [Google Scholar] [CrossRef]
- Clough, Y.; Krishna, V.; Corre, M.; Darras, K.; Denmead, L.; Meijide, A.; Moser, S.; Musshoff, O.; Steinebach, S.; Veldkamp, E.; et al. Land-use choices follow profitability at the expense of ecological functions in Indonesian smallholder landscapes. Nat. Commun. 2016, 7, 13137. [Google Scholar] [CrossRef]
- Kormann, U.G.; Grattarola, F.; Pacheco, A.; Tscharntke, T.; Clough, Y. Deforestation and habitat fragmentation differentially impact the richness and functional diversity of bees and wasps in tropical agricultural landscapes. J. Appl. Ecol. 2019, 56, 2671–2681. [Google Scholar]
- Azhar, B.; Saadun, N.; Puan, C.; Kamarudin, N.; Aziz, N.; Nurhidayu, S.; Fischer, J. Promoting landscape heterogeneity to improve the biodiversity benefits of certified palm oil production: Evidence from Peninsular Malaysia. Glob. Ecol. Conserv. 2015, 3, 553–561. [Google Scholar] [CrossRef]
- MPOB. Overview of Malaysian Palm Oil Industry. 2020. Available online: https://bepi.mpob.gov.my/images/overview/Overview_of_Industry_2020.pdf (accessed on 2 February 2023).
- Budiharta, S.; Erik, M.; Gaveau, D.; Struebig, M.; Wilting, A.; Kramer-Schadt, S.; Niedballa, J.; Raes, N.; Maron, M.; Wilson, K.A. Restoration to offset the impacts of developments at a landscape scale reveals opportunities, challenges and tough choices. Glob. Environ. Chang. 2018, 52, 152–161. [Google Scholar] [CrossRef]
- Lucey, J.M.; Palmer, G.; Yeong, K.; Edwards, D.; Senior, M.; Scriven, S.; Reynolds, G.; Hill, J.K. Reframing the evidence base for policy-relevance to increase impact: A case study on forest fragmentation in the oil palm sector. J. Appl. Ecol. 2017, 54, 731–736. [Google Scholar] [CrossRef]
- Viana, B.F.; Boscolo, D.; Neto, E.; Lopes, L.; Lopes, A.; Ferreira, P.; Pigozzo, C.; Primo, L.M. How well do we understand landscape effects on pollinators and pollination services? J. Pollinat. Ecol. 2012, 7, 31–40. [Google Scholar] [CrossRef]
- Mayfield, M.M. The importance of nearby forest to known and potential pollinators of oil palm (Elaeis guineënsis Jacq.; Areceaceae) in Southern Costa Rica. Econ. Bot. 2005, 59, 190–196. [Google Scholar] [CrossRef]
- Wisz, M.S.; Pottier, J.; Kissling, W.; Pellissier, L.; Lenoir, J.; Damgaard, C.; Dormann, C.; Forchhammer, M.; Grytnes, J.; Guisan, A.; et al. The role of biotic interactions in shaping distributions and realised assemblages of species: Implications for species distribution modelling. Biol. Rev. 2013, 88, 15–30. [Google Scholar] [CrossRef] [PubMed]
- González-Varo, J.P.; Biesmeijer, J.; Bommarco, R.; Potts, S.; Schweiger, O.; Smith, H.; Steffan-Dewenter, I.; Szentgyo, H.; Woyciechowski, M.; Vila, M. Combined effects of global change pressures on animal-mediated pollination. Trends Ecol. Evol. 2013, 28, 524–530. [Google Scholar] [CrossRef] [PubMed]
- Chinchilla, C.M.; Richardson, D.L. Pollinating insects and the pollination of oil palms in Central America. ASD Oil Palm Pap. (Costa Rica) 1991, 2, 1–18. [Google Scholar]
- Noor, W.N.W.M.; Nawi, N.; Seng, K.; Buda, M. Supply Response Analysis on the Impact of Climate Change on Oil Palm Production in Malaysia. in IOP Conf. Ser. Earth Environ. Sci. 2021, 757, 12009. [Google Scholar] [CrossRef]
- Kato, M.; Kosaka, Y.; Kawakita, A.; Okuyama, Y.; Kobayashi, C.; Phimminith, T.; Thongphan, D. Plant–pollinator interactions in tropical monsoon forests in Southeast Asia. Am. J. Bot. 2008, 95, 1375–1394. [Google Scholar] [CrossRef]
- Lumentut, N.; Meldy, H. The Demographic of Elaeidobius kamerunicus Faust Beetle (Coleoptera: Curculionidae) as Insect Pollinators on Oil Palm Plant. Bul. Palma 2016, 17, 89–95. [Google Scholar] [CrossRef]
- Rizuan, Z.A.C.M.; Hisham, H.N.; Samsudin, A. Role of Pollinating Weevil (Elaeidobius kamerunicus), Seasonal Effect and Its Relation to Fruit Set in Oil Palm Area of FELDA. In Proceedings of the PIPOC 2013 Conference, Kuala Lumpur, Malaysia, 19–21 November 2013; pp. 19–22. [Google Scholar]


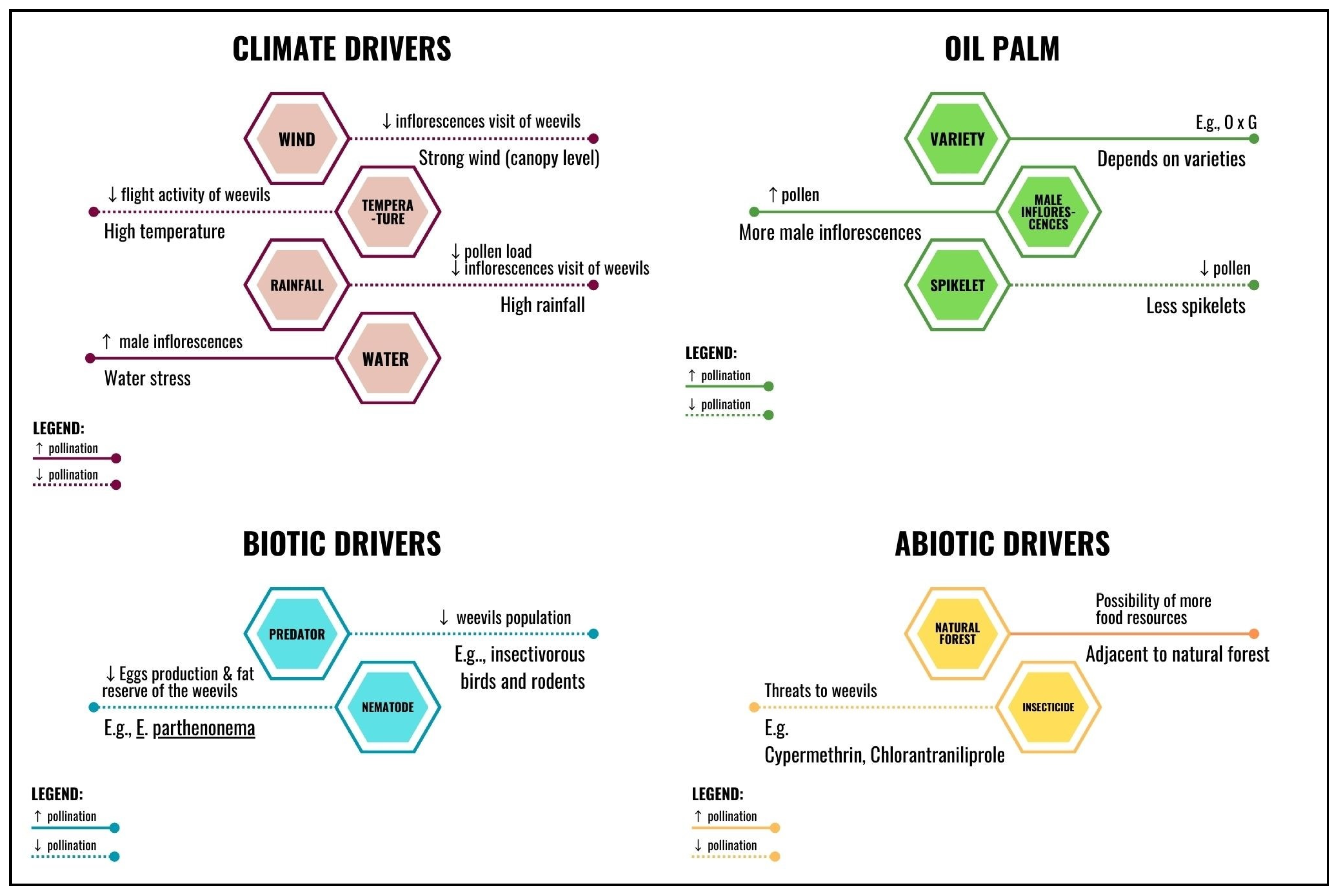{kind=link}
| Technique | Methods | Pollination Efficiency | Limitations |
|---|---|---|---|
| Hatch and Carry application [11,12,13] |
|
|
|
|
| ||
| Hand pollination [14] |
|
| |
|
| ||
| Wind- and weevil-assisted pollination [15] | Wind pollination—inflorescences were covered with fine nylon bags to prevent the entry of the pollinating weevils. Weevil pollination—inflorescences were covered with transparent plastic bags with openings at the bottom to facilitate the entry of the weevils. Wind + Weevil pollination—inflorescences were left open where the pollination was affected by both wind and weevils. Wind + Assisted pollination—inflorescences were covered with nylon-mesh bags and were given three rounds of assisted pollination from the first to the third day of anthesis. Wind + Weevil + Assisted pollination—inflorescences were left open to facilitate pollination by wind and weevils and were supplemented with two rounds of assisted pollination. |
| |
| Complementing-assisted pollination with artificial pollination [16] | Assisted pollination was carried out during anthesis by applying the pollen to the receptive female inflorescences. Subsequently, on the 7th and 14th days after anthesis, 1-naphthaleneacetic acid (NAA) was applied. In total, there were three applications for the inflorescences. |
|
|
|
| Characteristics |
|---|
|
| Characteristics | Elaeidobius kamerunicus | Thrips hawaiiensis | Pyroderces sp. |
|---|---|---|---|
| Active time | Morning to mid-day a | Early morning to late afternoon | Sunset d |
| Pupation habitats | Male inflorescence d | Soil c | Male inflorescence d |
| Most efficient period | Wet d | Dry d | ND |
| Activity in the female inflorescences | Feeding and ovipositing c | Feeding c | Ovipositing c |
| Activity in the male inflorescences | Incidental c | Inhabits b | Visits c |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gintoron, C.S.; Mohammed, M.A.; Sazali, S.N.; Deka, E.Q.; Ong, K.H.; Shamsi, I.H.; King, P.J.H. Factors Affecting Pollination and Pollinators in Oil Palm Plantations: A Review with an Emphasis on the Elaeidobius kamerunicus Weevil (Coleoptera: Curculionidae). Insects 2023, 14, 454. https://doi.org/10.3390/insects14050454
Gintoron CS, Mohammed MA, Sazali SN, Deka EQ, Ong KH, Shamsi IH, King PJH. Factors Affecting Pollination and Pollinators in Oil Palm Plantations: A Review with an Emphasis on the Elaeidobius kamerunicus Weevil (Coleoptera: Curculionidae). Insects. 2023; 14(5):454. https://doi.org/10.3390/insects14050454
Chicago/Turabian StyleGintoron, Christharina S., Muhamad Azmi Mohammed, Siti Nurlydia Sazali, Elvy Quatrin Deka, Kian Huat Ong, Imran Haider Shamsi, and Patricia Jie Hung King. 2023. "Factors Affecting Pollination and Pollinators in Oil Palm Plantations: A Review with an Emphasis on the Elaeidobius kamerunicus Weevil (Coleoptera: Curculionidae)" Insects 14, no. 5: 454. https://doi.org/10.3390/insects14050454