
Engineered Human Tissue as A New Platform for Mosquito Bite-Site Biology Investigations
 , , , and
, , , and
Abstract
:Simple Summary
Abstract
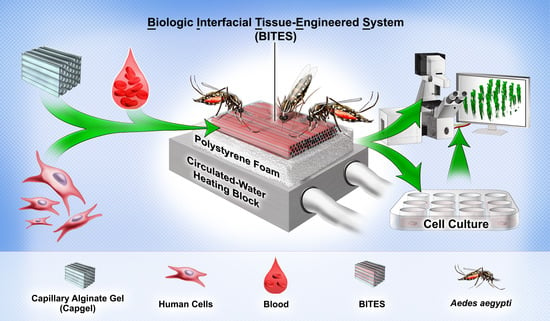
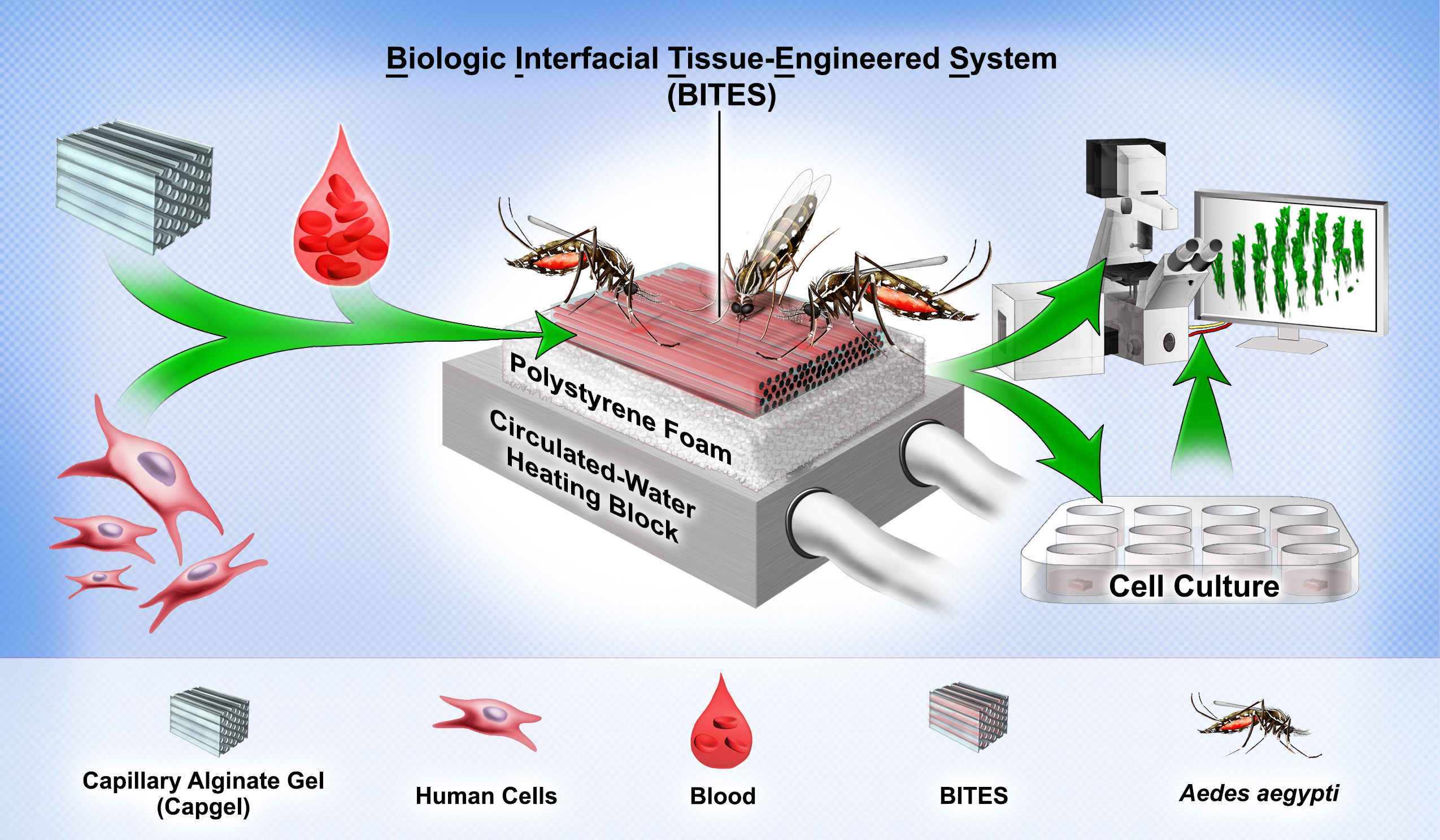{kind=link}
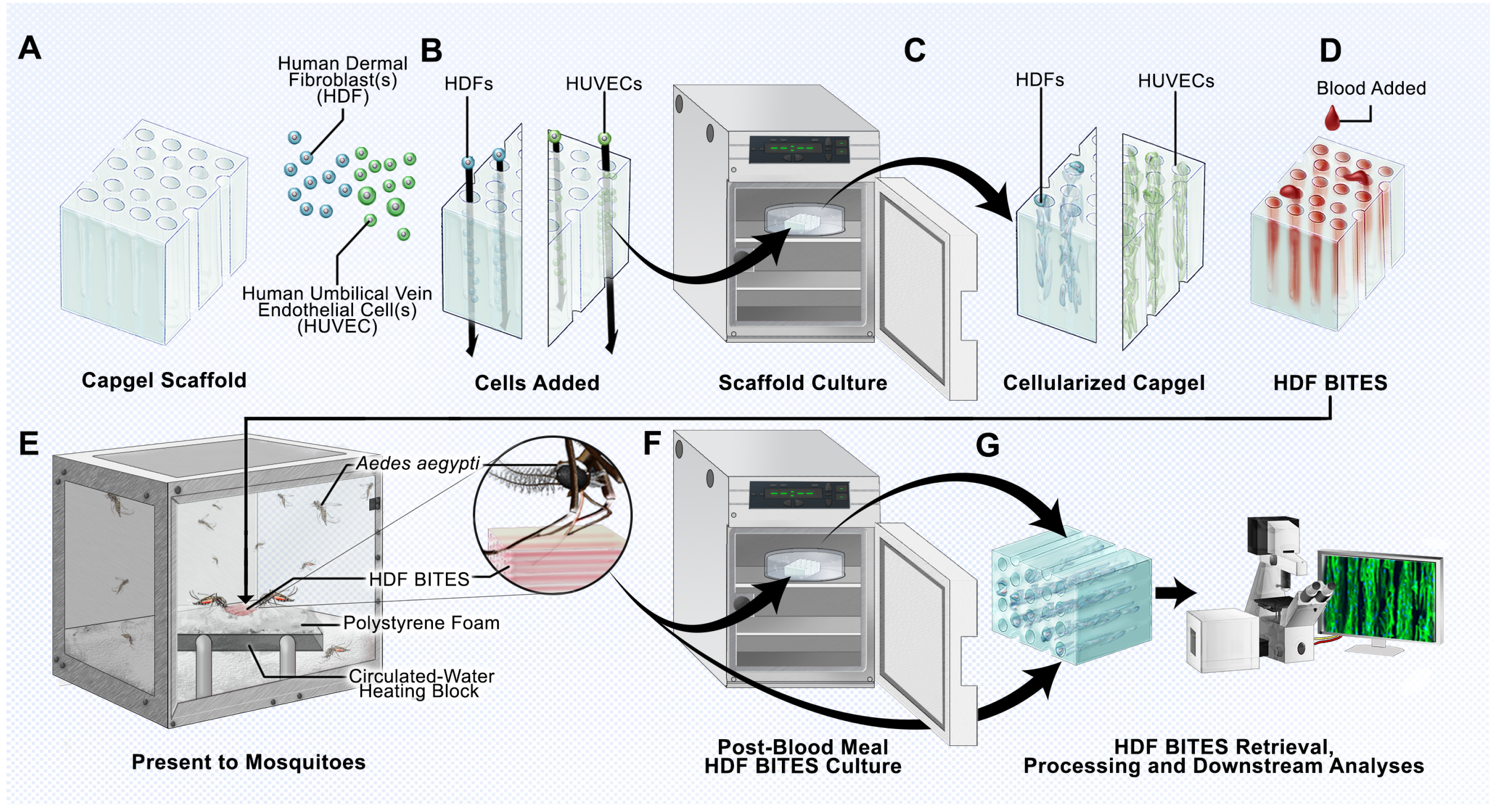{kind=link}
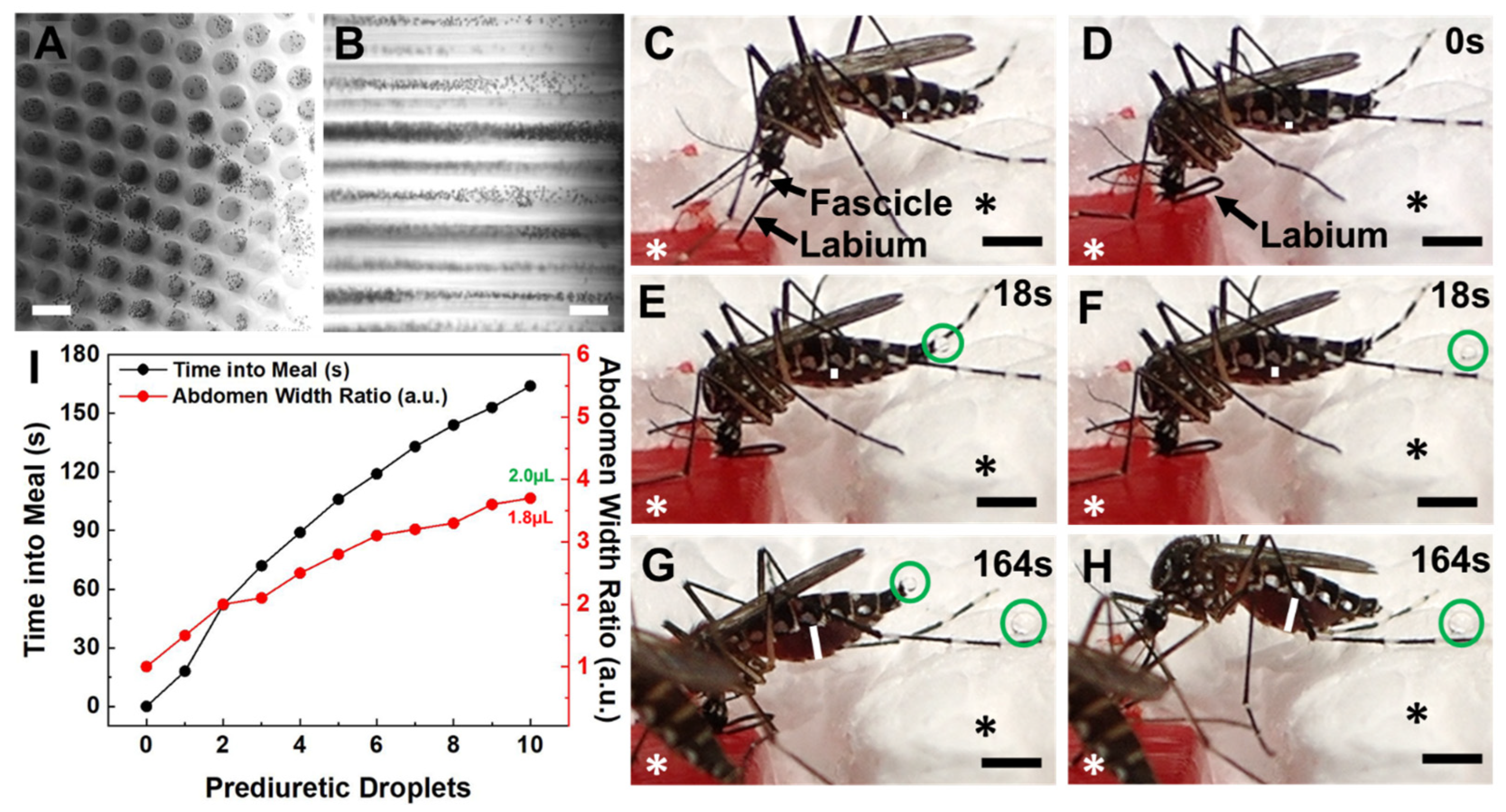{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Mosquito Rearing
2.2. Capgel Scaffold Synthesis and Processing
2.3. Capgel Scaffold Cellularization
2.4. Videography
2.5. Blood Meal Acquisition Experiments
2.6. Fixation, Staining, and Preparation of BITES Constructs
2.7. Confocal Microscopy and Stereomicroscopy
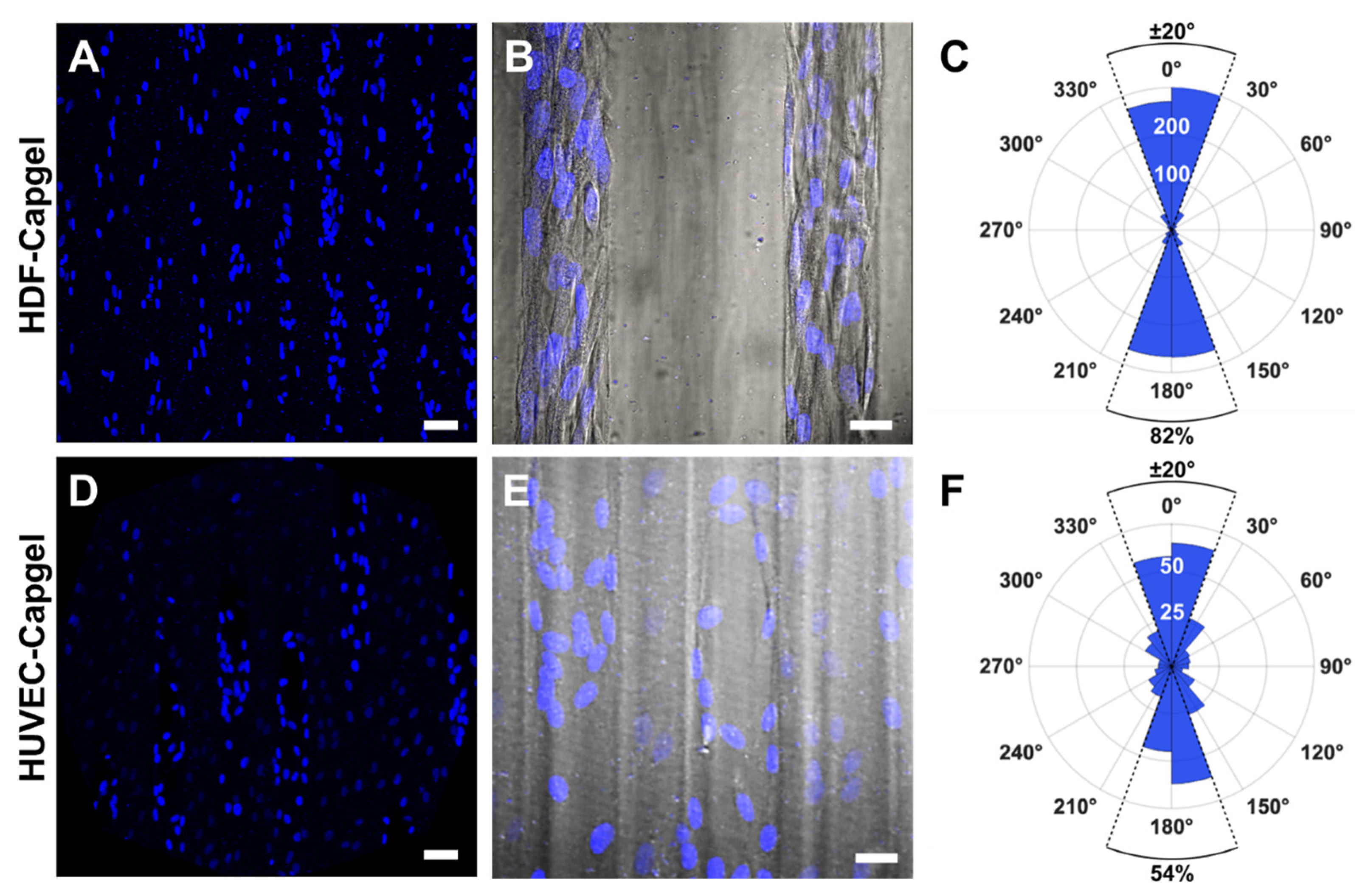
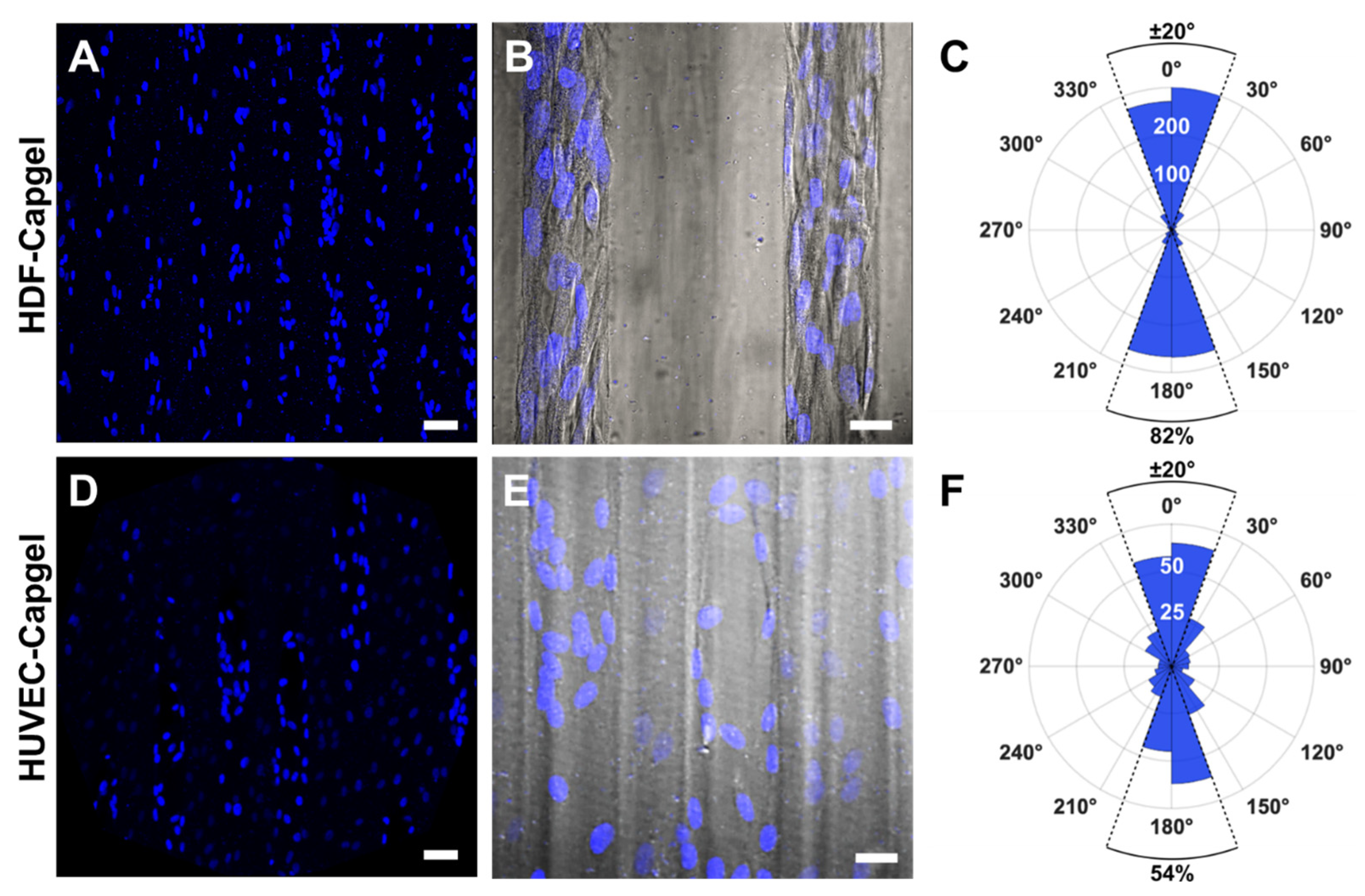
2.8. Evaluating Nuclear Orientation in BITES Microvessels
2.9. Statistical Analysis
3. Results
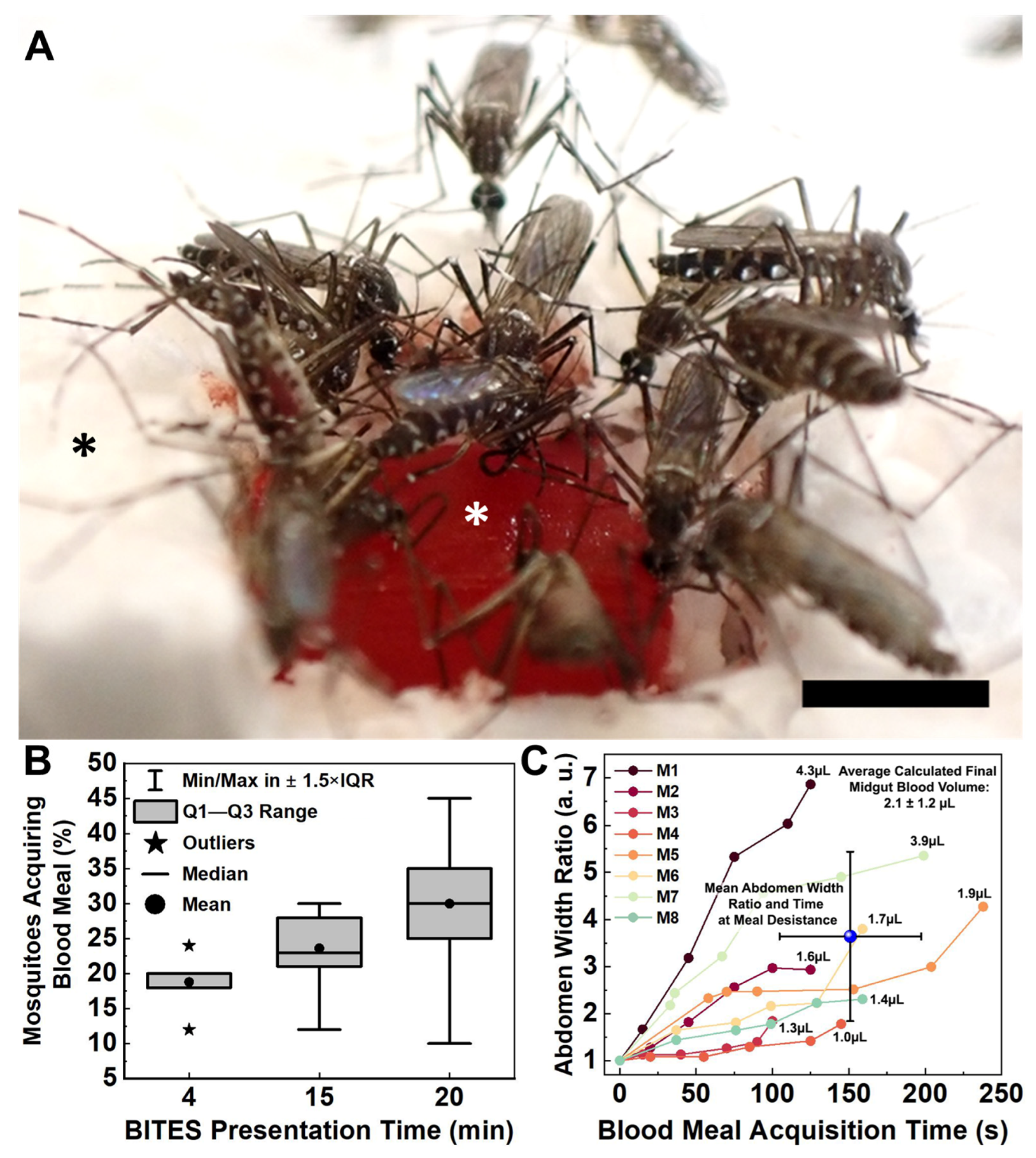
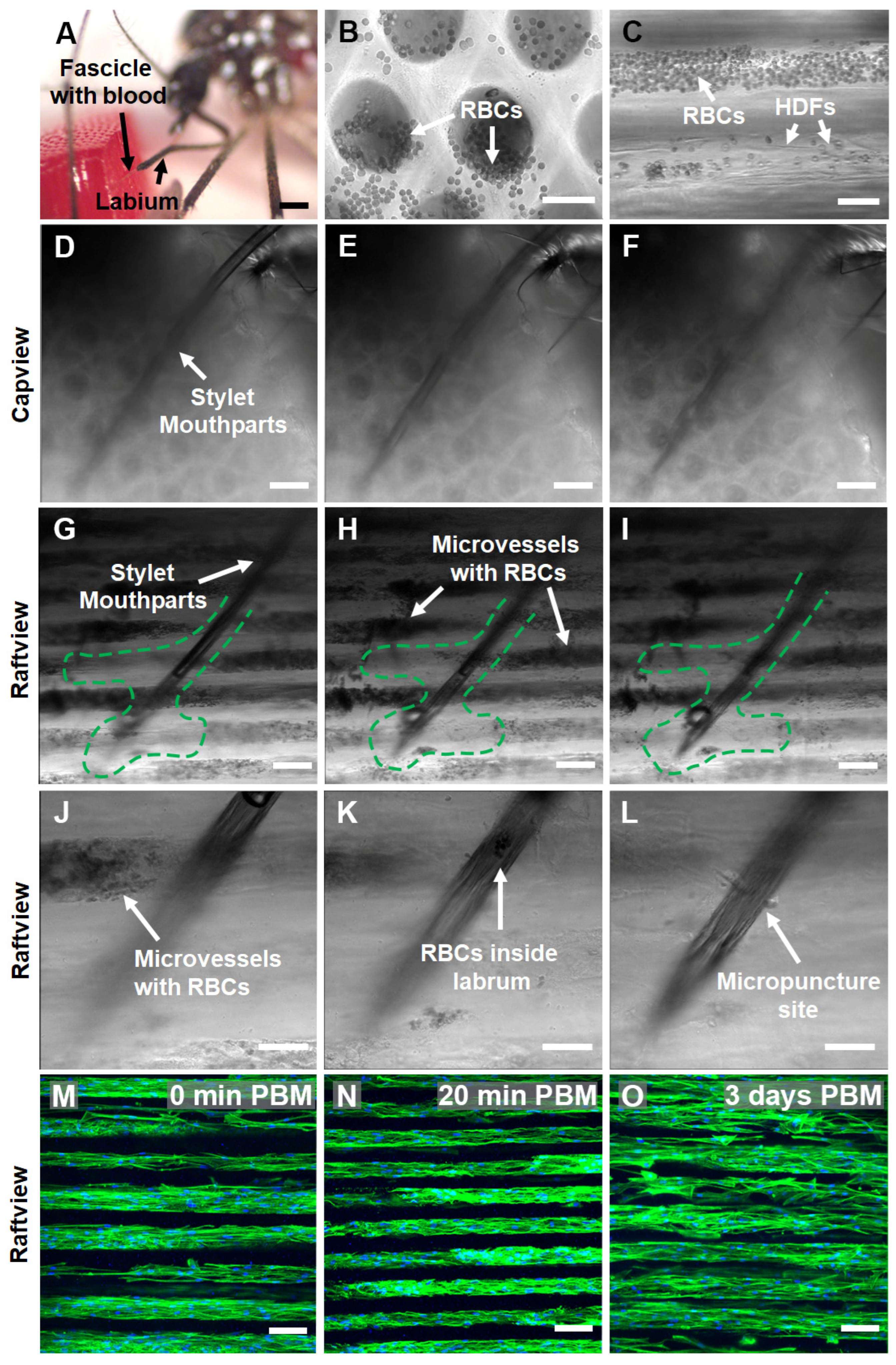
3.1. Blood Meal Acquisition by Ae. aegypti from Blood-Loaded, Non-Cellularized Capgel
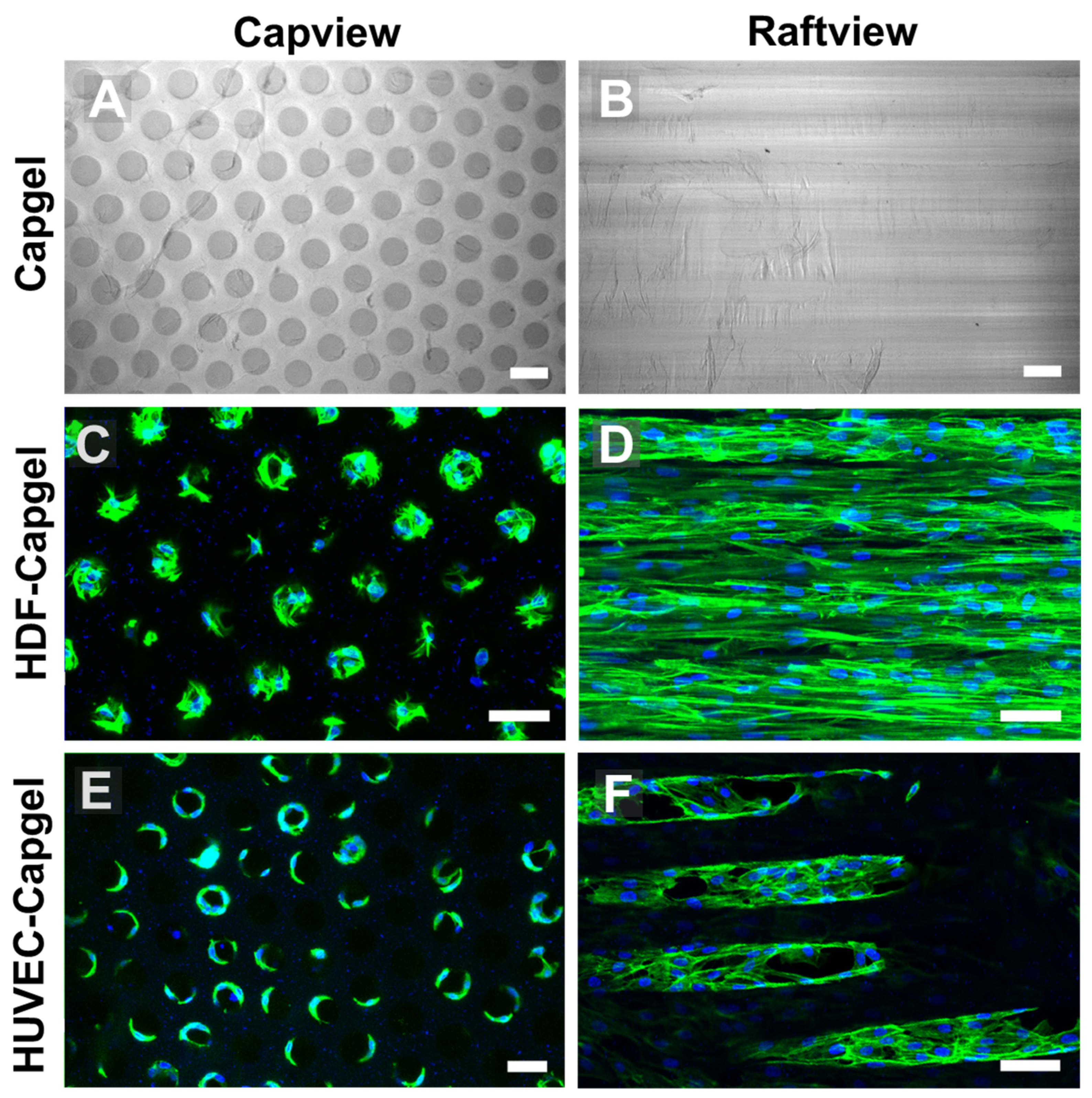
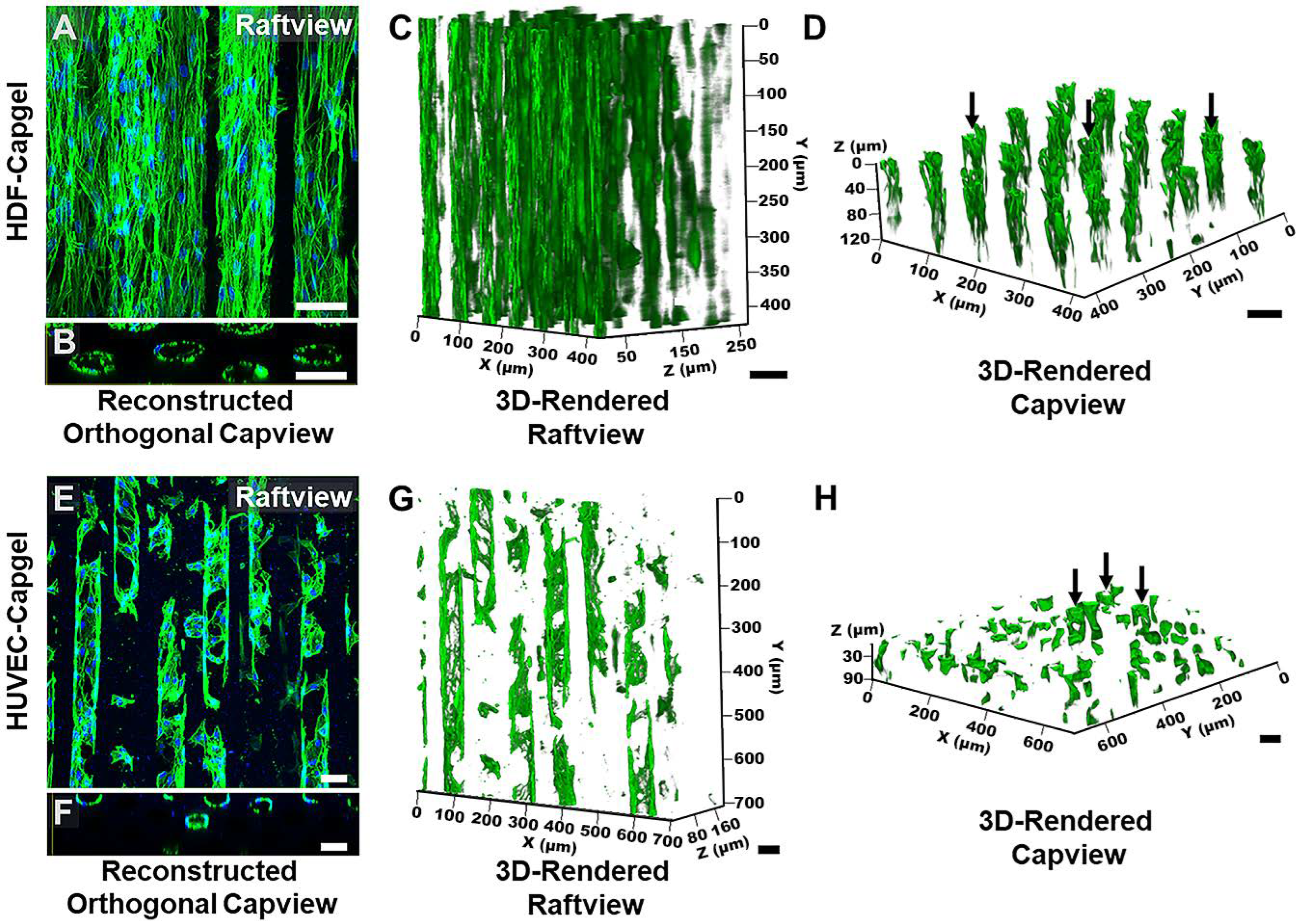
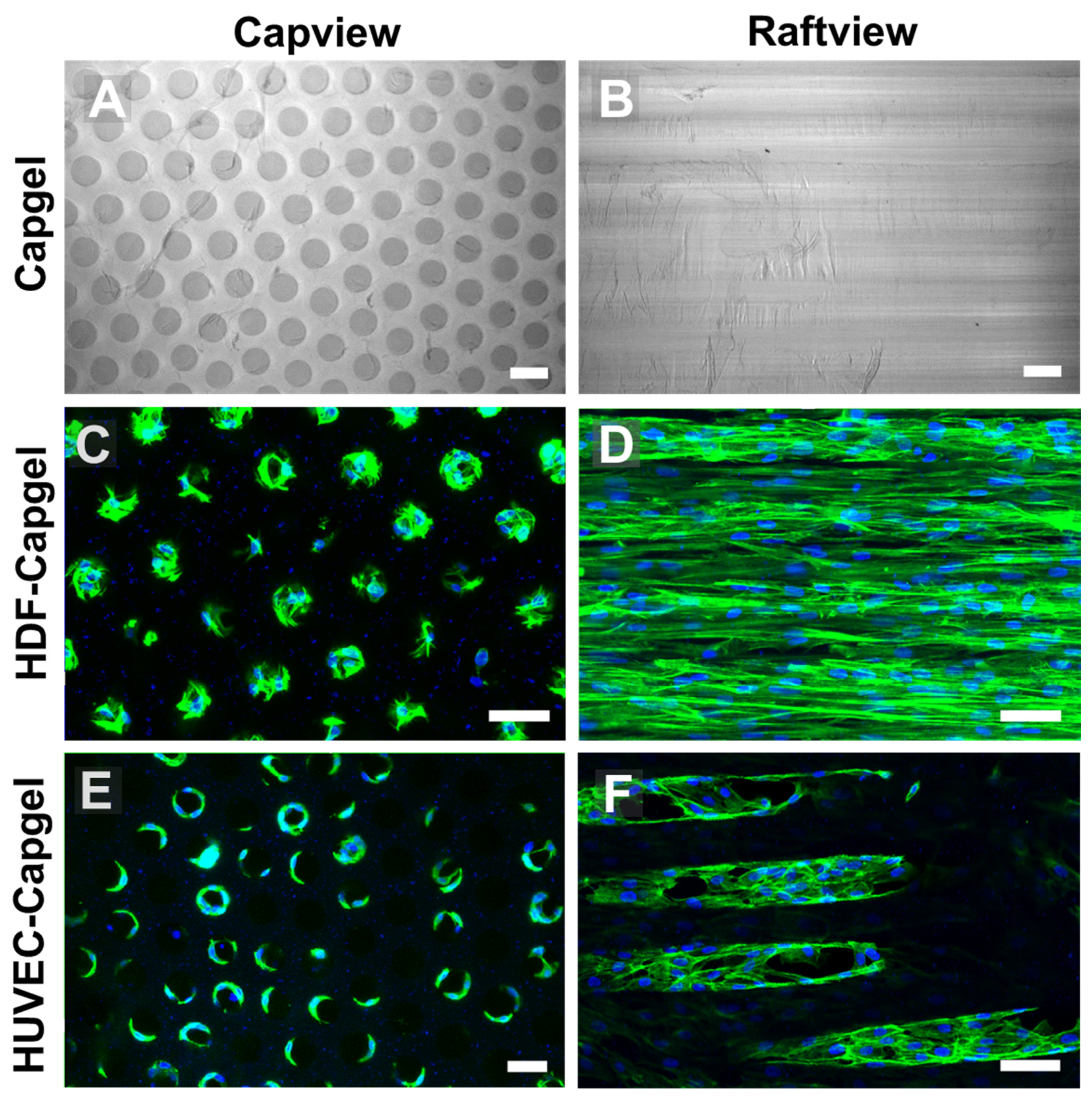
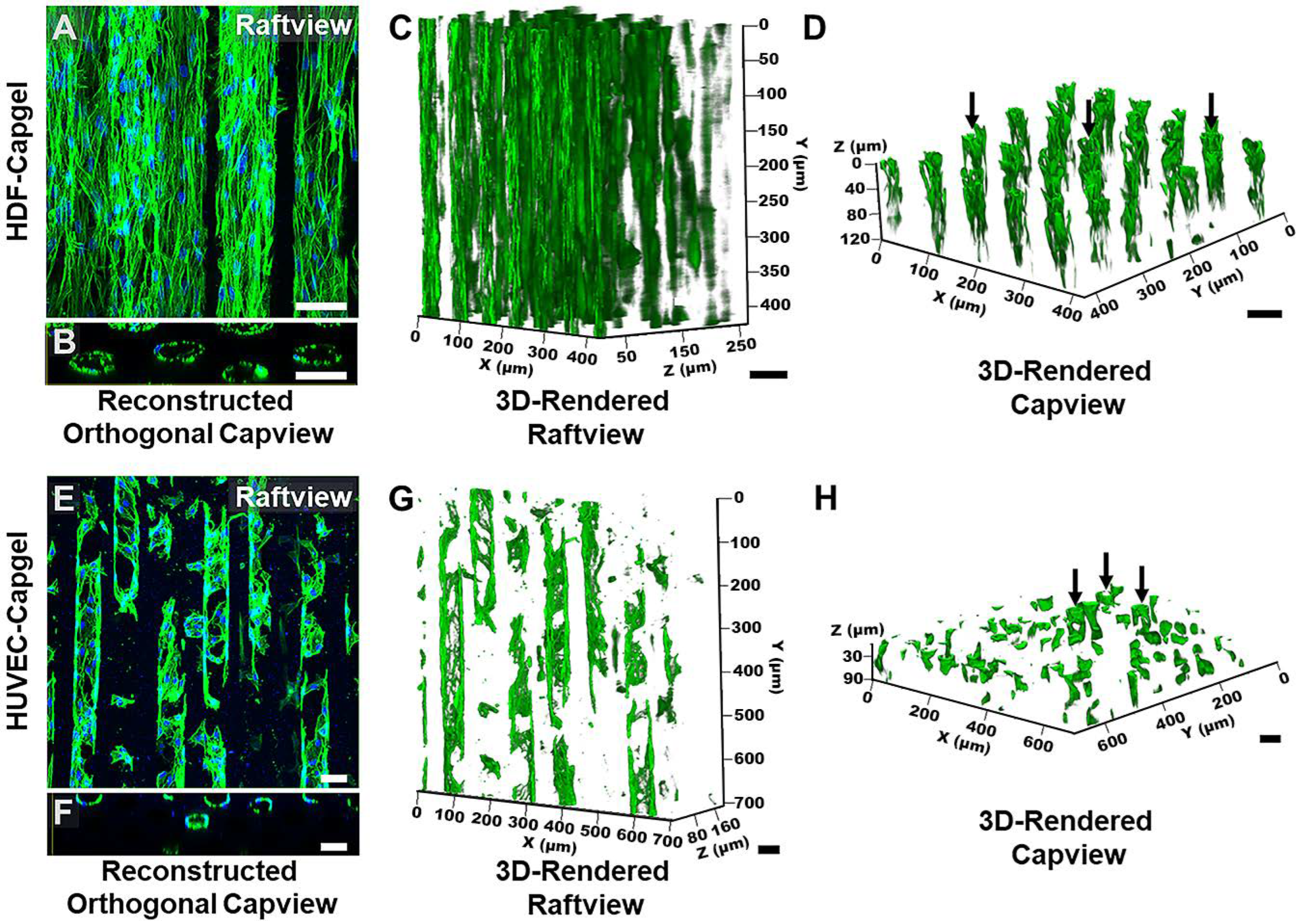
3.2. Cellularization of Capgel with HDFs or HUVECs
3.3. Blood Meal Acquisition by Ae. aegypti from BITES Tissue
3.4. In Situ Characterization of Ae. aegypti Stylet Mouthparts in BITES Microvascular Beds and Post-Blood-Meal BITES Culture
4. Discussion
5. Conclusions
6. Patents
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| Ae. | Aedes |
| BITES | Biologic Interfacial Tissue-Engineered System |
| Capgel | Capillary alginate gel |
| DEET | N,N-diethyl-meta-toluamide |
| DENV | Dengue virus |
| DMEM | Dulbecco’s Modified Eagle Medium |
| DI | Deionized |
| DIC | Differential interference contrast |
| FBS | Fetal bovine serum |
| FTSE | Full-thickness skin equivalents |
| HDFs | Human dermal fibroblasts |
| HUVECs | Human umbilical vein endothelial cells |
| IQR | Interquartile range |
| LVES | Large vessel endothelial supplement |
| PBM | Post-blood-meal |
| PBS | Phosphate-buffered saline |
| PFA | Paraformaldehyde |
| RBC | Red blood cell |
| SEM | Standard error of the mean |
| SD | Standard deviation |
| ZIKV | Zika virus |
References
- WHO. Global Vector Control Response 2017–2030; World Health Organization: Geneva, Switzerland, 2017. [Google Scholar]
- Wheeler, N.J.; Heimark, Z.W.; Airs, P.M.; Mann, A.; Bartholomay, L.C.; Zamanian, M. Genetic and functional diversification of chemosensory pathway receptors in mosquito-borne filarial nematodes. PLoS Biol. 2020, 18, e3000723. [Google Scholar] [CrossRef]
- GBD Disease Injury Incidence and Prevalence Collaborators. Global, regional, and national incidence, prevalence, and years lived with disability for 354 diseases and injuries for 195 countries and territories, 1990–2017: A systematic analysis for the Global Burden of Disease Study 2017. Lancet 2018, 392, 1789–1858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- GBD DALYs and HALE Collaborators. Global, regional, and national disability-adjusted life-years (DALYs) for 315 diseases and injuries and healthy life expectancy (HALE), 1990–2015: A systematic analysis for the Global Burden of Disease Study 2015. Lancet 2016, 388, 1603–1658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agarwal, A.; Joshi, G.; Nagar, D.P.; Sharma, A.K.; Sukumaran, D.; Pant, S.C.; Parida, M.M.; Dash, P.K. Mosquito saliva induced cutaneous events augment Chikungunya virus replication and disease progression. Infect. Genet. Evol. 2016, 40, 126–135. [Google Scholar] [CrossRef] [PubMed]
- Briant, L.; Desprès, P.; Choumet, V.; Missé, D. Role of skin immune cells on the host susceptibility to mosquito-borne viruses. Virology 2014, 464–465, 26–32. [Google Scholar] [CrossRef] [PubMed]
- Novelo, M.; Hall, M.D.; Pak, D.; Young, P.R.; Holmes, E.C.; McGraw, E.A. Intra-host growth kinetics of dengue virus in the mosquito Aedes aegypti. PLoS Pathog. 2019, 15, e1008218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guerrero, D.; Cantaert, T.; Misse, D. Aedes Mosquito Salivary Components and Their Effect on the Immune Response to Arboviruses. Front. Cell Infect. Microbiol. 2020, 10, 407. [Google Scholar] [CrossRef]
- Ryman, K.D.; Klimstra, W.B. Host responses to alphavirus infection. Immunol. Rev. 2008, 225, 27–45. [Google Scholar] [CrossRef]
- Romano, D.; Stefanini, C.; Canale, A.; Benelli, G. Artificial blood feeders for mosquitoes and ticks—Where from, where to? Acta Trop. 2018, 183, 43–56. [Google Scholar] [CrossRef] [PubMed]
- Joint FAO/IAEA Programme. Guidelines for Routine Colony Maintenance of Aedes Mosquito Species; Food and Agriculture Organization of the United Nations, Ed.; International Atomic Energy Agency: Vienna, Austria, 2017; Volume 17. [Google Scholar]
- Luo, Y.P. A novel multiple membrane blood-feeding system for investigating and maintaining Aedes aegypti and Aedes albopictus mosquitoes. J. Vector Ecol. 2014, 39, 271–277. [Google Scholar] [CrossRef]
- Rutledge, L.C.; Moussa, M.A.; Belletti, C.J. An in vitro blood-feeding system for quantitative testing of mosquito repellents. Mosq. News 1976, 36, 283–293. [Google Scholar]
- Ooi, C.-P.; Ahmad, R.; Ismail, Z. Effectiveness of several locally available membranes used for artificial feeding of Aedes albopictus Skuse. Trop. Biomed. 2005, 22, 69–71. [Google Scholar] [PubMed]
- Sri-in, C.; Weng, S.-C.; Shiao, S.-H.; Tu, W.-C. A simplified method for blood feeding, oral infection, and saliva collection of the dengue vector mosquitoes. PLoS ONE 2020, 15, e0233618. [Google Scholar] [CrossRef] [PubMed]
- Hamel, R.; Dejarnac, O.; Wichit, S.; Ekchariyawat, P.; Neyret, A.; Luplertlop, N.; Perera-Lecoin, M.; Surasombatpattana, P.; Talignani, L.; Thomas, F.; et al. Biology of Zika Virus Infection in Human Skin Cells. J. Virol. 2015, 89, 8880–8896. [Google Scholar] [CrossRef] [Green Version]
- Moser, L.A.; Boylan, B.T.; Moreira, F.R.; Myers, L.J.; Svenson, E.L.; Fedorova, N.B.; Pickett, B.E.; Bernard, K.A. Growth and adaptation of Zika virus in mammalian and mosquito cells. PLoS Negl. Trop. Dis. 2018, 12, e0006880. [Google Scholar] [CrossRef]
- Wichit, S.; Hamel, R.; Zanzoni, A.; Diop, F.; Cribier, A.; Talignani, L.; Diack, A.; Ferraris, P.; Liegeois, F.; Urbach, S.; et al. SAMHD1 Enhances Chikungunya and Zika Virus Replication in Human Skin Fibroblasts. Int. J. Mol. Sci. 2019, 20, 1695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Avirutnan, P.; Malasit, P.; Seliger, B.; Bhakdi, S.; Husmann, M. Dengue Virus Infection of Human Endothelial Cells Leads to Chemokine Production, Complement Activation, and Apoptosis. J. Immunol. 1998, 161, 6338–6346. [Google Scholar] [CrossRef] [PubMed]
- Esterly, A.T.; Lloyd, M.G.; Upadhyaya, P.; Moffat, J.F.; Thangamani, S. A Human Skin Model for Assessing Arboviral Infections. JID Innov. 2022, 2, 100128. [Google Scholar] [CrossRef]
- Limon-Flores, A.Y.; Perez-Tapia, M.; Estrada-Garcia, I.; Vaughan, G.; Escobar-Gutierrez, A.; Calderon-Amador, J.; Herrera-Rodriguez, S.E.; Brizuela-Garcia, A.; Heras-Chavarria, M.; Flores-Langarica, A.; et al. Dengue virus inoculation to human skin explants: An effective approach to assess in situ the early infection and the effects on cutaneous dendritic cells. Int. J. Exp. Pathol. 2005, 86, 323–334. [Google Scholar] [CrossRef] [PubMed]
- Janson, K.D.; Carter, B.H.; Jameson, S.B.; de Verges, J.E.; Dalliance, E.S.; Royse, M.K.; Kim, P.; Wesson, D.M.; Veiseh, O. Development of an automated biomaterial platform to study mosquito feeding behavior. Front. Bioeng. Biotechnol. 2023, 11, 1103748. [Google Scholar] [CrossRef]
- Schmid, M.A.; Glasner, D.R.; Shah, S.; Michlmayr, D.; Kramer, L.D.; Harris, E. Mosquito Saliva Increases Endothelial Permeability in the Skin, Immune Cell Migration, and Dengue Pathogenesis during Antibody-Dependent Enhancement. PLoS Pathog. 2016, 12, e1005676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reuter, C.; Walles, H.; Groeber, F. Preparation of a Three-Dimensional Full Thickness Skin Equivalent. Methods Mol. Biol. 2017, 1612, 191–198. [Google Scholar] [CrossRef] [PubMed]
- Willenberg, B.J.; Hamazaki, T.; Meng, F.W.; Terada, N.; Batich, C. Self-assembled copper-capillary alginate gel scaffolds with oligochitosan support embryonic stem cell growth. J. Biomed. Mater. Res. A 2006, 79, 440–450. [Google Scholar] [CrossRef] [PubMed]
- Willenberg, B.J.; Zheng, T.; Meng, F.W.; Meneses, J.C.; Rossignol, C.; Batich, C.D.; Terada, N.; Steindler, D.A.; Weiss, M.D. Gelatinized copper-capillary alginate gel functions as an injectable tissue scaffolding system for stem cell transplants. J. Biomater. Sci. Polym. Ed. 2011, 22, 1621–1637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rocca, D.G.; Willenberg, B.J.; Qi, Y.; Simmons, C.S.; Rubiano, A.; Ferreira, L.F.; Huo, T.; Petersen, J.W.; Ruchaya, P.J.; Wate, P.S.; et al. An injectable capillary-like microstructured alginate hydrogel improves left ventricular function after myocardial infarction in rats. Int. J. Cardiol. 2016, 220, 149–154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bosak, A.; Kwan, M.W.C.; Willenberg, A.; Perle, K.M.D.L.; Weinstein, D.; Hines, R.B.; Schultz, G.S.; Ross, E.A.; Willenberg, B.J. Capillary alginate gel (Capgel™) for the treatment of full-thickness dermal wounds in a hypoxic mouse model. Int. J. Polym. Mater. Polym. Biomater. 2018, 68, 1108–1117. [Google Scholar] [CrossRef]
- Anderson, W.A.; Willenberg, A.R.; Bosak, A.J.; Willenberg, B.J.; Lambert, S. Use of a capillary alginate gel (Capgel™) to study the three-dimensional development of sensory nerves reveals the formation of a rudimentary perineurium. J. Neurosci. Methods 2018, 305, 46–53. [Google Scholar] [CrossRef]
- George, D.S.; Anderson, W.A.; Sommerhage, F.; Willenberg, A.R.; Hines, R.B.; Bosak, A.J.; Willenberg, B.J.; Lambert, S. Bundling of axons through a capillary alginate gel enhances the detection of axonal action potentials using microelectrode arrays. J. Tissue Eng. Regen. Med. 2019, 13, 385–395. [Google Scholar] [CrossRef]
- Panarello, A.P.; Seavey, C.E.; Doshi, M.; Dickerson, A.K.; Kean, T.J.; Willenberg, B.J. Transforming Capillary Alginate Gel (Capgel) into New 3D-Printing Biomaterial Inks. Gels 2022, 8, 376. [Google Scholar] [CrossRef]
- Doshi, M.; Bosak, A.; Neal, C.J.; Isis, N.; Kumar, U.; Jeyaranjan, A.; Sakthivel, T.S.; Singh, S.; Willenberg, A.; Hines, R.B.; et al. Exposure to nanoceria impacts larval survival, life history traits and fecundity of Aedes aegypti. PLoS Negl. Trop. Dis. 2020, 14, e0008654. [Google Scholar] [CrossRef]
- National Institute of Standards and Technology. Chapter 7 Product and Process. Comparisons; Department of Commerce: Washington, DC, USA, 2012. [Google Scholar] [CrossRef]
- Ribeiro, J.M.; Rossignol, P.A.; Spielman, A. Aedes aegypti: Model for Blood Finding Strategy and Prediction of Parasite Manipulation. Exp. Parasitol. 1985, 60, 118–132. [Google Scholar] [CrossRef] [PubMed]
- Friuli, M.; Cafarchia, C.; Lia, R.P.; Otranto, D.; Pombi, M.; Demitri, C. From tissue engineering to mosquitoes: Biopolymers as tools for developing a novel biomimetic approach to pest management/vector control. Parasit. Vectors 2022, 15, 79. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Hol, F.J.H.; Pujhari, S.; Ellington, C.; Narayanan, H.V.; Li, H.; Rasgon, J.L.; Prakash, M. A microfluidic platform for highly parallel bite by bite profiling of mosquito-borne pathogen transmission. Nat. Commun. 2021, 12, 6018. [Google Scholar] [CrossRef]
- Chaves, L.F.; Harrington, L.C.; Keogh, C.L.; Nguyen, A.M.; Kitron, U.D. Blood feeding patterns of mosquitoes: Random or structured? Front. Zool. 2010, 7, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choo, Y.M.; Buss, G.K.; Tan, K.; Leal, W.S. Multitasking roles of mosquito labrum in oviposition and blood feeding. Front. Physiol. 2015, 6, 306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krenn, H.W.; Aspock, H. Form, function and evolution of the mouthparts of blood-feeding Arthropoda. Arthropod Struct. Dev. 2012, 41, 101–118. [Google Scholar] [CrossRef]
- Clement, A.N. Chapter 11: Adult Food and Feeding Mechanisms; The Biology of Mosquitoes; Chapman and Hall: London, UK; Cambridge, MA, USA, 1992; Volume 1, pp. 220–250. [Google Scholar]
- Arca, B.; Lombardo, F.; Valenzuela, J.G.; Francischetti, I.M.; Marinotti, O.; Coluzzi, M.; Ribeiro, J.M. An updated catalogue of salivary gland transcripts in the adult female mosquito, Anopheles gambiae. J. Exp. Biol. 2005, 208 Pt 20, 3971–3986. [Google Scholar] [CrossRef] [Green Version]
- Vogt, M.B.; Lahon, A.; Arya, R.P.; Kneubehl, A.R.; Spencer Clinton, J.L.; Paust, S.; Rico-Hesse, R. Mosquito saliva alone has profound effects on the human immune system. PLoS Negl. Trop. Dis. 2018, 12, e0006439. [Google Scholar] [CrossRef]
- Nayar, J.K.; Sauerman, D.M. The Effects of Nutrition on Survival and Fecundity in Florida Mosquitoes: Part 2. Utilization of a blood meal for survival. J. Med. Entomol. 1975, 12, 99–103. [Google Scholar] [CrossRef]
- Briegel, H.; Rezzonico, L. Concentration of Host Blood Protein During Feeding by Anopheline Mosquitoes (Diptera: Culicidae). J. Med. Entomol. 1985, 22, 612–618. [Google Scholar] [CrossRef] [Green Version]
- Beyenbach, K.W.; Petzel, D.H. Diuresis in Mosquitoes: Role of a Natriuretic Factor. Am. Physiol. Soc. 1987, 2, 171–175. [Google Scholar] [CrossRef]
- Lahondère, C.; Lazzari, C.R. Mosquitoes Cool Down during Blood Feeding to Avoid Overheating. Curr. Biol. 2012, 22, 40–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stobbart, R.H. The control of the diuresis following a blood meal in females of the yellow fever mosquito aedes Aegypti (L.). J. Exp. Biol. 1977, 69, 53–85. [Google Scholar] [CrossRef] [PubMed]
- Braverman, I.M. The Cutaneous Microcirculation. J. Investig. Dermatol. Symp. Proc. 2000, 5, 3–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kosyakova, N.; Kao, D.D.; Figetakis, M.; Lopez-Giraldez, F.; Spindler, S.; Graham, M.; James, K.J.; Won Shin, J.; Liu, X.; Tietjen, G.T.; et al. Differential functional roles of fibroblasts and pericytes in the formation of tissue-engineered microvascular networks in vitro. NPJ Regen. Med. 2020, 5, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hetheridge, C.; Mavria, G.; Mellor, H. Uses of the in vitro endothelial-fibroblast organotypic co-culture assay in angiogenesis research. Biochem. Soc. Trans. 2011, 39, 1597–1600. [Google Scholar] [CrossRef] [PubMed]
- Berthod, F. Chapter 7. Fibroblasts and Endothelial Cells: The Basic Angiogenic Unit; Angiogenesis: Insights from a Systematic Overview; Nova Science Publishers: Hauppauge, NY, USA, 2013; pp. 145–157. [Google Scholar]
- Kunz-Schughart, L.A.; Schroeder, J.A.; Wondrak, M.; van Rey, F.; Lehle, K.; Hofstaedter, F.; Wheatley, D.N. Potential of fibroblasts to regulate the formation of three-dimensional vessel-like structures from endothelial cells in vitro. Am. J. Physiol. Cell Physiol. 2006, 290, C1385–C1398. [Google Scholar] [CrossRef] [Green Version]
- Uwamori, H.; Ono, Y.; Yamashita, T.; Arai, K.; Sudo, R. Comparison of organ-specific endothelial cells in terms of microvascular formation and endothelial barrier functions. Microvasc. Res. 2019, 122, 60–70. [Google Scholar] [CrossRef] [PubMed]
- Langille, B.L.; Adamson, S.L. Relationship between Bloow Flow Direction and Endothelial Cell Orientation at Arterial Branch Sites in Rabbits and Mice. Circ. Res. 1981, 48, 481–488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tkachenko, E.; Gutierrez, E.; Saikin, S.K.; Fogelstrand, P.; Kim, C.; Groisman, A.; Ginsberg, M.H. The nucleus of endothelial cell as a sensor of blood flow direction. Biol. Open 2013, 2, 1007–1012. [Google Scholar] [CrossRef] [Green Version]
- Vion, A.-C.; Perovic, T.; Petit, C.; Hollfinger, I.; Bartels-Klein, E.; Frampton, E.; Gordon, E.; Claesson-Welsh, L.; Gerhardt, H. Endothelial Cell Orientation and Polarity Are Controlled by Shear Stress and VEGF Through Distinct Signaling Pathways. Front. Physiol. 2021, 11, 623769. [Google Scholar] [CrossRef]
- Sinha, R.; Le Gac, S.; Verdonschot, N.; van den Berg, A.; Koopman, B.; Rouwkema, J. Endothelial cell alignment as a result of anisotropic strain and flow induced shear stress combinations. Sci. Rep. 2016, 6, 29510. [Google Scholar] [CrossRef] [Green Version]
- Aubin, H.; Nichol, J.W.; Hutson, C.B.; Bae, H.; Sieminski, A.L.; Cropek, D.M.; Akhyari, P.; Khademhosseini, A. Directed 3D cell alignment and elonga.ation in microengineered hydrogels. Biomaterials 2010, 31, 6941–6951. [Google Scholar] [CrossRef] [Green Version]
- Calcutt, J.J.; Roberts, M.S.; Anissimov, Y.G. Predicting Viable Skin Concentration: Modelling the Subpapillary Plexus. Pharm. Res. 2022, 39, 783–793. [Google Scholar] [CrossRef] [PubMed]
- Le Coupanec, A.; Babin, D.; Fiette, L.; Jouvion, G.; Ave, P.; Misse, D.; Bouloy, M.; Choumet, V. Aedes Mosquito Saliva Modulates Rift Valley Fever Virus Pathogenicity. PLoS Negl. Trop. Dis. 2013, 7, e2237. [Google Scholar] [CrossRef] [PubMed]
- Pingen, M.; Bryden, S.R.; Pondeville, E.; Schnettler, E.; Kohl, A.; Merits, A.; Fazakerley, J.K.; Graham, G.J.; McKimmie, C.S. Host Inflammatory Response to Mosquito Bites Enhances the Severity of Arbovirus Infection. Immunity 2016, 44, 1455–1469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Styer, L.M.; Lim, P.Y.; Louie, K.L.; Albright, R.G.; Kramer, L.D.; Bernard, K.A. Mosquito saliva causes enhancement of West Nile virus infection in mice. J. Virol. 2011, 85, 1517–1527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Surasombatpattana, P.; Ekchariyawat, P.; Hamel, R.; Patramool, S.; Thongrungkiat, S.; Denizot, M.; Delaunay, P.; Thomas, F.; Luplertlop, N.; Yssel, H.; et al. Aedes aegypti saliva contains a prominent 34-kDa protein that strongly enhances dengue virus replication in human keratinocytes. J. Investig. Dermatol. 2014, 134, 281–284. [Google Scholar] [CrossRef] [Green Version]
- Onlamoon, N.; Noisakran, S.; Hsiao, H.M.; Duncan, A.; Villinger, F.; Ansari, A.A.; Perng, G.C. Dengue virus-induced hemorrhage in a nonhuman primate model. Blood 2010, 115, 1823–1834. [Google Scholar] [CrossRef] [Green Version]
- Mores, C.N.; Christofferson, R.C.; Davidson, S.A. The role of the mosquito in a dengue human infection model. J. Infect. Dis. 2014, 209 (Suppl. 2), S71–S78. [Google Scholar] [CrossRef] [Green Version]
- Leitner, W.W.; Costero-Saint Denis, A.; Wali, T. Immunological consequences of arthropod vector-derived salivary factors. Eur. J. Immunol. 2011, 41, 3396–3400. [Google Scholar] [CrossRef] [PubMed]
- Cevc, G.; Vierl, U. Spatial distribution of cutaneous microvasculature and local drug clearance after drug application on the skin. J. Control Release 2007, 118, 18–26. [Google Scholar] [CrossRef] [PubMed]
- Sorrell, J.M.; Caplan, A.I. Fibroblast heterogeneity: More than skin deep. J. Cell Sci. 2004, 117 Pt 5, 667–675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva, T.L.A.; Radtke, A.; Balaban, A.; Pascini, T.V.; Pala, Z.R.; Roth, A.; Alvarenga, P.H.; Jeong, Y.J.; Olivas, J.; Ghosh, A.K.; et al. The fibrinolytic system enables the onset of Plasmodiuminfection in the mosquito vector and the mammalian host. Sci. Adv. 2021, 7, eabe3362. [Google Scholar] [CrossRef] [PubMed]
- Dormont, L.; Mulatier, M.; Carrasco, D.; Cohuet, A. Mosquito Attractants. J. Chem. Ecol. 2021, 47, 351–393. [Google Scholar] [CrossRef] [PubMed]
- Seavey, C.E.; Doshi, M.; Colamarino, A.; Kim, B.N.; Dickerson, A.K.; Willenberg, B.J. Graded Atmospheres of Volatile Pyrethroid Overlaid on Host Cues can be Established and Quantified within a Novel Flight Chamber for Mosquito Behavior Studies. Environ. Entomol. 2023, 52, 197–209. [Google Scholar] [CrossRef] [PubMed]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Seavey, C.E.; Doshi, M.; Panarello, A.P.; Felice, M.A.; Dickerson, A.K.; Jewett, M.W.; Willenberg, B.J. Engineered Human Tissue as A New Platform for Mosquito Bite-Site Biology Investigations. Insects 2023, 14, 514. https://doi.org/10.3390/insects14060514
Seavey CE, Doshi M, Panarello AP, Felice MA, Dickerson AK, Jewett MW, Willenberg BJ. Engineered Human Tissue as A New Platform for Mosquito Bite-Site Biology Investigations. Insects. 2023; 14(6):514. https://doi.org/10.3390/insects14060514
Chicago/Turabian StyleSeavey, Corey E., Mona Doshi, Andrew P. Panarello, Michael A. Felice, Andrew K. Dickerson, Mollie W. Jewett, and Bradley J. Willenberg. 2023. "Engineered Human Tissue as A New Platform for Mosquito Bite-Site Biology Investigations" Insects 14, no. 6: 514. https://doi.org/10.3390/insects14060514