
Fatal Attraction: Argiope Spiders Lure Male Hemileuca Moth Prey with the Promise of Sex


1
McGuire Center for Lepidoptera and Biodiversity, Research Associate, Florida Museum of Natural History, University of Florida, 3215 Hull Rd., UF Cultural Plaza, Gainesville, FL 32611-2710, USA
2
Department of Plant Pathology, University of Georgia, 2315 Miller Plant Sciences, Athens, GA 30602, USA
*
Author to whom correspondence should be addressed.
Insects 2024, 15(1), 53; https://doi.org/10.3390/insects15010053
Submission received: 14 December 2023
/
Revised: 27 December 2023
/
Accepted: 9 January 2024
/
Published: 12 January 2024
(This article belongs to the Section Other Arthropods and General Topics)
{kind=link}
Abstract
:Simple Summary
Predators and prey have direct interactions that influence their short-term behaviors, including resource allocation and strategies for moving through habitats. However, the presently observed behaviors are the products of coevolutionary interactions, posited to be a history of measures and countermeasures between the predator and prey. We found that Argiope (orb-weaver) spiders in the continental USA appear to use a pheromone lure that mimics the mating pheromone of the day-flying Hemileuca moth (buck moth) to entice male moths into their webs. We found evidence that different phylogenetic groups of Hemileuca moths respond to the Argiope pheromone lure with a broad range of responses, ranging from indifferent to acutely strongly attracted, suggesting a coevolutionary history of predator–prey countermeasures. One of these countermeasures may be the potential evolution of moth developmental timing (adult emergence) to avoid Argiope predation in areas where the ranges of the moths and spiders overlap.
Abstract
Predator–prey coevolution, particularly chemo-ecological arms races, is challenging to study as it requires the integration of behavioral, chemical ecology, and phylogenetic studies in an amenable system. Moths of the genus Hemileuca (Saturniidae) are colorful, diurnal, and fast and often fly well above the vegetation canopy layer. However, several Hemileuca species have been reported as being captured in spider webs, specifically Argiope species (Araneidae). Female Hemileuca are known to produce mating pheromones and spiders of the Araneidae family are known to use pheromone lures to attract lepidopteran prey. We presented primarily female Argiope aurantia, which are attractive to male Anisota pellucida (Saturniidae), to different populations of Hemileuca species across the southern and western United States to categorize the homing response strength of different species of male Hemileuca. When we mapped these Argiope lure attraction strength categories onto the most recently published Hemileuca phylogeny, the behavioral patterns suggested a potential co-evolutionary arms race between predators and prey. Males of Hemileuca maia, H. grotei, and H. nevadensis (all in the same clade) appeared to have no attraction to A. aurantia, while H. magnifica and H. hera (within a different, separate clade) appeared to be strongly attracted to A. aurantia, but H. nuttalli (also within the H. hera and H. magnifica clade) displayed no attraction. Furthermore, Hemileuca eglanterina (yet a different clade) displayed strong, weak, and no attraction to A. aurantia, depending on the population. These apparent clade partitioning patterns of Argiope lure effectiveness and within-species variation in Hemileuca lure responses suggest a predator–prey coevolutionary history of measures and countermeasures.
1. Introduction
Coevolution, the reciprocal adaptation or counter-adaptation between interacting species, can drive organism diversification, population differentiation, and ecological specialization [1,2,3]. Initially conceived as an explanation for the tradeoffs between host plant specialization and host plant defenses [1], coevolution may also influence predator–prey interactions through repeated countermeasures of predator tactics and prey responses [4,5,6,7,8]. Although reciprocal behavioral adaptation by predators and prey may be rapid and relatively short-lived when compared with speciation rates [9], the signatures of predator–prey coevolution occur throughout nature, appearing as highly derived and specialized behavioral interactions. Studies of coevolutionary predator–prey behavioral interactions can only represent brief snapshots in evolutionary time. However, these studies imply that predator–prey systems with highly specialized interactions have most likely evolved to their present states over successive generations of predator specialization and prey countermeasures [10,11].
Much of the evidence for predator–prey coevolution arises from the study of chemo-ecological interactions. Along one axis of chemo-ecological interactions, arms races between toxic prey and toxin-resistant predators appear to generate a landscape mosaic of semi-independently evolving predator–prey populations that vary in their degree of toxicity, resistance, and predation/avoidance behaviors [12]. Over many generations, the landscape mosaic of different selective pressures and species’ interactions can diversify predator tactics and prey responses, potentially even facilitating the radiation of Müllerian and Batesian mimicry systems [13,14]. Along another axis of chemo-ecological evolutionary interactions, olfactory cues (scents and lures) emerge as potentially prominent drivers in predator–prey coevolution [11]. For species interactions that are driven by chemical cues, the prey species of ambush predators appear capable of detecting and modifying their behavior to reduce predation risk or avoid the predator altogether, while naïve prey species, or those with a new evolutionary history with a predator, are insensitive to predator chemical cues and experience a higher predation rate [15,16]. Such systems set up spatiotemporal metapopulation dynamics of predator–prey colonization and extinctions that can be labile and modified over evolutionary time scales.
Spiders are well-known for their use of chemical attractants to bait prey to their location [17]. As spiders tend to be mostly sessile, central-place foragers [17,18,19], the deployment of volatile compounds to bait and concentrate potential prey increases the chances of successful capture, compared to a passive approach of simple web placement. In one of the most thoroughly studied of these chemical lure systems, large immature and adult female bolas spiders of the genera Mastaphora, Cladomelea, and Ordgarius produce a palette of sex pheromone mimics to attract male moth prey. When baited into the spider’s capture range, the bolas spider lassos the moth with a highly modified, single-stranded web [20,21,22,23]. Juvenile bolas spiders of both sexes and adult male bolas spiders, on the other hand, emit pheromones to attract moth flies (Psychodidae), which are captured by the spiders at close range with their forelegs [24,25]. The secondary roles of spider pheromone volatiles appear to be diverse and widespread [17,26], suggesting the potential for predator–prey coevolutionary geographic mosaics of population and taxonomic differentiation similar to those observed in other chemo-ecological systems (e.g., garter snakes and their toxic amphibian prey [12]).
The degree to which pheromone lure systems have become specialized for prey attraction and how coevolution may be shaping these interactions is not well understood [26]. In North America, there are anecdotal accounts detailing the capture of large (8 to 10 cm), day-flying, male Hemileuca species (buck moths) (Saturniidae: Hemileucinae) in the webs of Argiope species (Araneidae) [27,28,29]. Male Hemileuca are vagile, strong-flying moths that typically fly well above vegetation canopies in grassland and shrub-dominated habitats [27,29,30]. However, the webs of Argiope in moth habitats are spun between plants, often within 1.5 m of the ground, where male Hemileuca rarely fly except to court females (adult Hemileuca have incomplete digestive systems so they do not seek food resources). Unmated female Hemileuca species emit a mixture of volatile pheromones that aid in mate location [31,32,33,34,35]. These pheromones trigger circular locating flights (often 500 m to 1 km in diameter) by males, in which, when the pheromone becomes concentrated enough, they then search up a pheromone gradient (~100 m) to precisely locate the calling female [27,29,30,35]. Observations of Argiope web locations and the capture of strictly male Hemileuca moths strongly suggest that female Argiope use a chemical lure/pheromone trap to bait male Hemileuca prey.
Chemically variable pheromones are emitted within and between different Hemileuca species, and they appear to be relatively diverse in mixture composition as well as in stereochemical structure [31,32,33,34,35]. Each Hemileuca species appears to have a potentially unique suite and mixture of pheromones, but even different populations of the same species can vary in their pheromone profiles [31,32,35]. Due to the standing diversity and variation of Hemileuca pheromones, it is possible that Hemileuca–Argiope predator–prey relationships are shaped by coevolutionary chemo-ecological interactions that vary over space and time—a predator–prey arms race. To evaluate this possibility, we characterized the relative attractiveness of female Argiope spiders regarding male Hemileuca homing for different Hemileuca species across North America and overlaid these interactions on the most recent and complete published Hemileuca phylogeny. A coevolutionary predator–prey arms race should generate a mosaic of species–species interactions across the phylogeny where the pheromone lure is effective and is not countered by the prey, along with other instances where the pheromone lure is ineffective. In a phylogenetic framework, evidence of coevolutionary interactions between Hemileuca and Argiope could be manifested as clade-specific patterns of lure effectiveness and/or a wide range of lure responses within a clade or a single Hemileuca species.
2. Materials and Methods
2.1. Study Species
Argiope are large (~3–7 cm), colorful, orb-weaving spiders (Araneidae), which are distributed throughout the world. Argiope aurantia, commonly known as the black-and-yellow garden spider, is frequently observed in gardens, fields, and along lake edges across the United States, although it is less common in the western Great Plains, the Rocky Mountain region, and in the western deserts [36]. Adults of A. aurantia mature in late summer through autumn in most of its range [37], although they mature as early as June in Florida (Warren pers. obs.). Argiope are general insect feeders and accept a very wide range of prey items. Mature A. aurantia females may live as long as 6 months (Warren pers. obs.) Throughout North America, the ranges of A. aurantia and Hemileuca (see below) broadly overlap.
Hemileuca (buck moths) are relatively large (8–10 cm), colorful, day-flying Saturniidae (silk moths) that are restricted to North America and Mexico [29]. Like most adult saturniid moths, Hemileuca adults lack functional feeding mouthparts, and their digestive systems are incomplete [27,29]; thus, the adults do not feed and are short-lived [27,29,30]. Hemileuca mate location, at the broader landscape level, is accomplished through volatile sex pheromones. These pheromones can occur as a dominant volatile or as a mixture of multiple volatiles [31,32,33,34,35,38]. Hemileuca pheromones are known to be species-specific, but they may be cross-compatible as the pheromone components of one species may attract the males of another [34,39]. During mate location, male Hemileuca often fly in large circles through a habitat where females are likely to occur. When a pheromone plume is detected, the flight paths become smaller in diameter, enabling the males to geospatially map where the pheromone concentrations are greatest, ultimately following the pheromone concentration gradient to the emitting female [27,40].
2.2. Evaluating Argiope Lure Attraction to Male Hemileuca
Mature or last-instar immature female Agriope aurantia (hereafter Argiope) that were observed to attract diurnal male Anisota pellucida (Saturniidae) (up to 20 individual A. pellucida moths each day; see Video S1 in the Supplementary Materials) were gathered from the vicinity of Gainesville, FL, USA, and kept in 0.75 m × 0.75 m rectangular mesh collapsible cages. The captive Argiope were fed crickets purchased from PetSmart every 2–3 days, a diet occasionally supplemented with wild-harvested grasshoppers; at no point were the spiders offered Hemileuca prey. Caged Argiope were kept in a manner that would maximize their health and welfare throughout the duration of the experiment, including regular misting with water and exposure to sunlight. During field assays from 2016–2018, immature female Argiope were also gathered and presented at field sites to determine which instars attract moths and it was determined that only mature or last-instar Argiope appeared capable of emitting volatile Hemileuca lures.
At the study sites, side-by-side cages (one cage containing a calling Argiope and one cage without a spider) were deployed ~5 m apart from each other, from the late morning to mid-afternoon when Hemileuca were actively flying. Cages were placed in habitats with low vegetation and ample exposure, such as ridge tops, hilltops, and elevated roads, so that incoming male Hemileuca moths could be clearly observed and to enhance the dispersal of emitted pheromones. Because spider webs themselves may visually attract prey [41,42,43], we presented Argiope in their cages to counter any signal based on the webs and to standardize their presentation to Hemileuca.
We developed a relatively straightforward set of behavioral criteria for categorizing the degree of attraction to cages (with and without Argiope) by searching male Hemileuca moths within 10 m of the cages. Although this 10-meter distance may or may not be the male moths’ true perceptual range, there was a clear behavioral response to Argiope spiders at this distance that was straightforward to identify and record for individual males. To be categorized as being strongly attracted to Argiope, Hemileuca males had to deviate from their flight path, either contact the ground immediately in front of the cage or contact the cage itself, and remain near the cage location (<2.5 m) for more than two minutes. We considered a deviation from the original flight path and movement towards the cage without stopping or contacting the cage as “weak attraction”. No attraction was assigned to a male that did not deviate from its flight path (within a 10-meter radius of Argiope) and orient towards Argiope. For each male that came within 10 m of the experimental Argiope, we recorded whether it was strongly attracted, weakly attracted, or not attracted to the “calling Argiope”.
In our behavioral assays, indexing moth attraction was important, and we were also concerned with other potential sources of attraction besides the Argiope. We directly assessed potential alternative sources of attraction/repulsion/indifference prior to formal data collection. First, the cages alone may either attract or repel Hemileuca, due to unknown and uncharacterized properties. After placing unoccupied cages near Argiope-occupied cages and unoccupied cages in isolation, it was immediately apparent that the unoccupied cages were not attractive to male Hemileuca. Although the cages did not appear to be repulsive to male Hemileuca (e.g., turning away from the cages once a perceptual range threshold was crossed), it is possible that the cages produce repulsive signals at close distances that are overwhelmed by the Argiope volatile. Second, not all Argiope appeared to emit the lure at all times, but some individuals reliably called for longer periods of time (daily, for weeks) while others only called on some days. Therefore, we frequently presented multiple Argiope aurantia (up to 12 individuals in separate cages), to ensure a high likelihood that at least one individual was emitting. Occasionally, we also evaluated two additional Argiope species (A. florida and A. trifasciata). Although we omit these species from formal statistical analysis due to limited sampling and experimental effort, we consider them as potentially relevant biological observations. Last, after we established that a caged Argiope putatively called and attracted a number of male Hemileuca, we moved the cage to another location >5 m away as a manipulation test of lure attractiveness. In these instances, some males remained for 5 to 10 min at the location where the cage had previously been positioned, but most males reoriented to the new location of the calling spider within 1 min. We also noted individual variations in the Argiope attractiveness of lures that varied over space and time. Multiple caged Argiope may call simultaneously at the same site but there appeared to be obvious individual differences in lure effectiveness (Video S2 in the Supplementary Materials). In other instances, an individual spider would be an effective caller one day but not the next. Together, these observations strongly suggest that relative lure attractiveness was estimated with our methodology and that the lure-emitting system is under some degree of individual control.
2.3. Lure Attraction and Overlay on a Published Hemileuca Phylogeny
We compared the percentage of Hemileuca moths displaying evidence of strong attraction to the caged Argiope to the percentage of those that did not display strong attraction (with weak and no-attraction combined) via a two-tailed Z-proportions test (α < 0.05) to assign each population a categorical lure response. In opting for a proportions test, we assumed that the number of spiders deployed at any site on any date would not impact the homing behavior to a degree that would overcome the broader (more conservative) properties of the proportions test based on a Z-statistic. For Hemileuca species containing more than one study population, if >80% of the aggregate populations were categorized as strongly attracted, we designated that species with a strong lure attraction response (the same for the no-lure response category). If the percentage of populations did not statistically differ from a 50% ratio of no attraction (and weak attraction) to strong attraction, we considered that particular Hemileuca taxon to have a mixed lure response. We then mapped these three lure attraction categories, namely, strong attraction, no attraction, and mixed attraction, for each Hemileuca species with behavioral lure assays (H. eglanterina, H. grotei, H. hera, H. magnifica, H. maia, H. nevadensis, and H. nuttalli) (Supplementary Table S1) onto the most recent published Hemileuca phylogeny, as presented by Rubinoff and Sperling [36]. The phylogeny we selected was the maximum likelihood tree produced from combined cytochrome oxidase I (COI—mtDNA) and elongation factor 1 alpha (EF1α—nuclear DNA) ([36] Figure 6) because that tree was constructed from the greatest amount of sequence information. By overlaying these assayed Argiope lure categories onto the published Hemileuca phylogeny, we could evaluate whether there may be a phylogenetic signal associated with Argiope lure attraction.
3. Results
Seven Hemileuca species, namely, H. eglanterina, H. grotei, H. hera, H. magnifica, H. maia, H. nevadensis and H. nuttalli, were assayed for Argiope lure attraction across the western, central, and southern USA from populations in the states of New Mexico, Utah, Colorado, Wyoming, Texas, Georgia, and Florida (Supplementary Table S1). A total of 33 populations were presented with caged Argiope for an aggregate observational time of ~127 h (Supplementary Table S1).
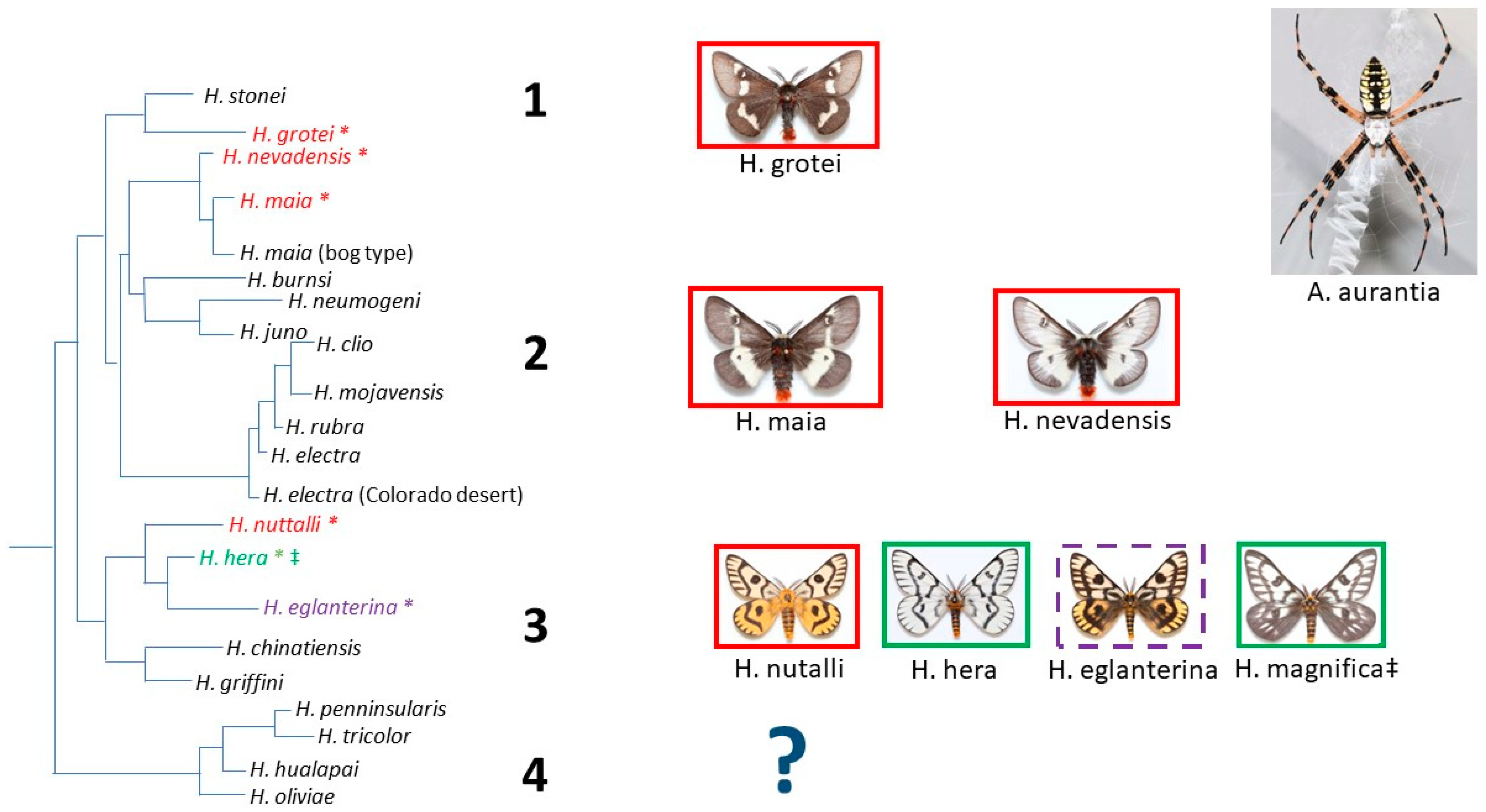
When we overlaid the species categories of Argiope lure attraction on the published Hemileuca phylogeny, there appeared to be evidence of phylogenetic association related to lure effectiveness. All sampled populations of Hemileuca grotei, H. maia, and H. nevadensis consistently showed no homing responses by any individual male moths to caged Argiope (Supplementary Table S1); these taxa occurred in clades 1 and 2 (Figure 1). Likewise, strong lure attractiveness appeared in the Hemileuca taxa associated with clade 3 (H. hera and H. magnifica), but this clade also contained species without any evidence of attraction (H. nuttalli), along with one species containing populations that were either strongly attracted, weakly attracted, or showed no evidence of attraction (H. eglanterina) (Figure 1, Supplementary Table S1).
For H. hera and H. magnifica, there was one population of each species where Argiope lures appeared to be weakly or non-attractive to most male moths (Supplementary Table S1). It is likely that in these instances the Argiope had ceased lure emission as moths were attracted to the same Argiope on a later date or later that day in a close-proximity subpopulation (Supplementary Table S1). We also noted that Arigiope florida and A. trifasciata did not appear to emit a lure that elicited a homing response from male H. grotei, H. maia, and H. nevadensis. However, these three Hemileuca species were not documented as being attracted to A. aurantia either (Figure 1), and the presentation of A. florida and A. trifasciata was an infrequent, late-season occurrence (Supplementary Table S1).
4. Discussion
In our research, there was evidence of varied Argiope lure effectiveness, suggesting that a chemical coevolutionary arms race may be occurring between Argiope aurantia and moths of the genus Hemileuca. First, although we performed lure behavioral bioassays on seven Hemileuca species occurring throughout the western, central, and southern USA, we have representatives from 3 of the 4 primary Hemileuca clades defined by Rubinoff and Sperling [36]. We observed a variation in apparent Argiope lure effectiveness among taxa between clades and there may be a phylogenetic association with whether or not the Argiope lure attracts male Hemileuca (Figure 1). None of the clades were fully represented in our behavioral assays, but members of clades 1 and 2 (H. grotei, H. maia, and H. nevadensis) showed no evidence of lure attraction in the species and populations evaluated (Figure 1, Supplementary Table S1). Not a single instance of weak attraction was observed in any H. grotei, H. maia, or H. nevadensis individual, suggesting that although the spiders were likely emitting pheromones, those volatile chemicals were not capable of inducing a homing response towards the Argiope. Counter to our results, Horton [28] reported the finding of Hemileuca lucina, which is closely related to H. maia and H. nevadensis (though not included in the phylogeny of Rubinoff and Sperling [36]) in the webs of Argiope. There are also anecdotal reports of male Hemileuca clio (placed within clade 2 as defined by Rubinoff and Sperling [36]), which is distantly related to H. maia and H. nevadensis, being attracted to Argiope in Arizona which was verbally communicated to the first author by M. Collins and S. McElfresh. Thus, the distribution and intensity of Argiope lure attractiveness across clade 2 appears to be more variable and is perhaps even biogeographically partitioned in a way not revealed by our study. The clade-specific sorting of traits may emerge as a consequence of a relatively long-standing coevolutionary history between Argiope and Hemileuca (as opposed to a recently evolved interaction), but we cannot validate this evolutionary history with the data we present.
Although the taxa in Hemileuca clades 1 and 2 are sparsely represented, the Hemileuca spp. in clade 3 (Figure 1) were more thoroughly assayed for Argiope lure responses. The patterns of Argiope lure responses by male Hemileuca in clade 3 suggest the potential for an ongoing evolutionary chemical arms race between predator and prey. We found clear evidence of strong lure attraction to Argiope by H. hera and H. magnifica. In some populations, Argiope attracted males in the high hundreds to their cages (Supplementary Table S1). Yet, within this same clade, there was no evidence of attraction, even weak attraction, to Argiope by any individuals of H. nuttalli from multiple different populations (Supplementary Table S1). This suggests the potential finding that either Argiope pheromones may have recently evolved to lure H. hera and H. magnifica, that H. nuttalli has recently evolved with an indifference to the lures, or that the lures were potentially never effective with H. nuttalli. We do not yet have the resolved information that would enable a strongly supported answer to these three scenarios, but the patterns of strong lure effectiveness and lure ineffectiveness for baiting Hemileuca prey is another expected consequence of predator–prey coevolutionary arms races [1,2,3,4,5,6].
Perhaps the most interesting and biologically revealing patterns of predator–prey coevolutionary responses in lure attraction reside with the species H. eglanterina. We had evidence that the Argiope lure spans the range of male moths’ homing responses between H. eglanterina populations, including no attraction, strong attraction, and weak attraction to the caged spiders (Supplementary Table S1). Interestingly, H. eglanterina is known to have a mixture of female moth pheromone volatile constituents that, when mixed in certain proportions, are effective at attracting male H. eglanterina in some populations and not in others [32,34]. It is possible that the pheromone constituent diversity in H. eglanterina either predisposes it to Argiope lure attraction or to indifference, or that perhaps the pheromone mixtures are evolving in response to Argiope lures. Although we do not know the Argiope lure’s chemical composition, it is also possible that they too may be capable of producing different volatile mixtures since bolas spiders, which are also Araneidae, are known to do this to target specific prey [23,24,25].
Argiope predation on male Hemileuca in nature remains anecdotal, unquantified, and infrequently reported in the literature [27,28,29]. There are two speculative but potentially revealing observations we present, based on the qualitative knowledge gained during our study. First, while Argiope occur in the majority of the Hemileuca habitats assayed during this study, and their ranges in North America broadly overlap [29,37], locally, Argiope usually mature (the calling life stage of Argiope) following the main flights of summer-flying Hemileuca (H. eglanterina, H. nuttalli, H. hera, and H. magnifica), temporally separating predator from prey. Our experimental introduction of mature female Argiope into Hemileuca populations during their peak flight represents a scenario that is unlikely to occur naturally in these populations. Given the intensity of the attraction of male H. hera and H. magnifica to mature, calling Argiope, it is possible that the chemical arms race between these taxa has helped shape the mid-summer flight times of these species, as opposed to the late fall flight times of species in clades 1 and 2. In clades 1 and 2, these taxa tend to have a long pupal diapause stage that extends into the late fall and early winter months [29], wherein Argiope may or may not overlap with Hemileuca flights. While the following proposal is speculative, and while there are birds and large dragonflies (Aeshnidae) that are adult moth predators, it is possible that either the flight times of Hemileuca coincidentally do not overlap with the Argiope life stages that are capable of predating adult Hemileuca or that Hemileuca development has evolved to avoid Argiope predation. An example of this potentially resides in the H. maia group, where the adult moths emerge from late October through early January (depending on latitude), flying after adult Argiope have died. Second, the Argiope lure may not be restricted to Hemileuca nor to Argiope aurantia, as we have evidence of strong lure attraction to another saturniid moth genus, Anisota (Saturniidae: Ceratocampinae), and evidence that other Argiope species likely produce a moth pheromone lure as well (Supplementary Table S1). Third, there may be asymmetric sexual selection for sensitivity to the Argiope lure as we never recorded 100% of the observed males homing to caged Argiope, nor were any female Hemileuca observed to be baited by the spiders (Supplementary Table S1). The skewed sex ratio suggests that it is likely that the predator–prey coevolutionary interactions differ for male and female moths. Finally, a mating pheromone of Argiope bruennichi has been published [44] but it remains to be tested whether the Argiope mating pheromone can also bind and elicit the homing response in Hemileuca or if the Argiope lure is produced independently of the spider mating pheromone.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/insects15010053/s1, Table S1: Hemileuca behavior data. Video S1: Anisota pellucida at Argiope aurantia, Video S2: Hemileuca magnifica at Argiope aurantia.
Author Contributions
Conceptualization, A.D.W. and P.M.S.; methodology, A.D.W. and P.M.S.; formal analysis, P.M.S. and A.D.W.; investigation, A.D.W. and P.M.S.; data curation, A.D.W. and P.M.S.; writing—original draft preparation, P.M.S. and A.D.W.; writing—review and editing, P.M.S. and A.D.W. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
Behavioral assay data, site locations, dates, and proportion Z-test results are presented in the Supplementary Materials.
Acknowledgments
We thank S. Warren, W. Bernstein, J. Cox, T. Ortenberger, T. Lawyer, E. Anderson, and R. Peigler for their assistance during field experiments, help with spider acquisition and maintenance, and for discussions. We also thank J. Adams for accommodations during fieldwork, and M. Collins for literature and discussions. Two anonymous reviewers provided thoughtful comments that helped improve this manuscript.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Ehrlich, P.R.; Raven, P.H. Butterflies and plants: A study in coevolution. Evolution 1964, 18, 586–608. [Google Scholar] [CrossRef]
- Pellmyr, O. Yuccas, yucca moths, and coevolution: A review. Ann. Mo. Bot. Gard. 2003, 90, 35–55. [Google Scholar] [CrossRef]
- Yoder, J.B.; Nuismer, S.L. When does coevolution promote diversification? Am. Nat. 2010, 176, 802–817. [Google Scholar] [CrossRef]
- West, K.; Cohen, A.; Baron, M. Morphology and behavior of crabs and gastropods from Lake Tanganyika, Africa: Implications for lacustrine predator-prey coevolution. Evolution 1991, 45, 589–607. [Google Scholar] [CrossRef]
- Vermeij, G.J. The evolutionary interaction among species: Selection, escalation, and coevolution. Ann. Rev. Ecol. Syst. 1994, 25, 219–236. [Google Scholar] [CrossRef]
- Abrams, P.A. The evolution of predator-prey interactions: Theory and evidence. Ann. Rev. Ecol. Syst. 2000, 31, 79–105. [Google Scholar] [CrossRef]
- Hieber, C.S.; Wilcox, S.R.; Boyle, J.; Uetz, G.W. The spider and fly revisited: Ploy–counterploy behavior in a unique predator–prey system. Behav. Ecol. Sociobiol. 2002, 53, 51–60. [Google Scholar] [CrossRef]
- Kopp, M.; Tollrian, R. Reciprocal phenotypic plasticity in a predator–prey system: Inducible offences against inducible defences? Ecol. Lett. 2003, 6, 742–748. [Google Scholar] [CrossRef]
- Hairston, N.G., Jr.; Ellner, S.P.; Geber, M.A.; Yoshida, T.; Fox, J.A. Rapid evolution and the convergence of ecological and evolutionary time. Ecol. Lett. 2005, 8, 1114–1127. [Google Scholar] [CrossRef]
- Lehtonen, J.; Whitehead, M.R. Sexual deception: Coevolution or inescapable exploitation. Curr. Zool. 2014, 60, 52–61. [Google Scholar] [CrossRef]
- Lindstedt, C.; Mokkonen, M. The evolutionary strategy of deception. Curr. Zool. 2014, 60, 1–5. [Google Scholar] [CrossRef]
- Brodie, E.D.; Ridenhour, B.J.; Brodie, E.D., III. The evolutionary response of predators to dangerous prey: Hotspots and coldspots in the geographic mosaic of coevolution between garter snakes and newts. Evolution 2002, 56, 2067–2082. [Google Scholar] [CrossRef]
- Sherratt, T.N. The evolution of warning signals. Proc. R. Soc. Lond. B 2002, 269, 741–746. [Google Scholar] [CrossRef]
- Speed, M.P.; Ruxton, G.D.; Blount, J.D.; Stephens, P.A. Diversification of honest signals in a predator–prey system. Ecol. Lett. 2010, 13, 744–753. [Google Scholar] [CrossRef]
- Downes, S.; Shine, R. Sedentary snakes and gullible geckos: Predator–prey coevolution in nocturnal rock-dwelling reptiles. Anim. Behav. 1998, 55, 1373–1385. [Google Scholar] [CrossRef]
- Heiling, A.M.; Herberstein, M.E. Predator–prey coevolution: Australian native bees avoid their spider predators. Proc. R. Soc. Lond. B 2004, 271, S196–S198. [Google Scholar] [CrossRef]
- Pekár, S.; Toft, S. Trophic specialization in a predatory group: The case of prey-specialized spiders (Araneae). Biol. Rev. 2015, 90, 744–761. [Google Scholar] [CrossRef]
- Nyffeler, M.; Sterling, W.L.; Dean, D.A. How spiders make a living. Environ. Entomol. 1994, 23, 1357–1367. [Google Scholar] [CrossRef]
- Seer, F.K.; ElBalti, N.; Schrautzer, J.; Irmler, U. How much space is needed for spider conservation? Home range and movement patterns of wolf spiders (Aranea, Lycosidae) at Baltic Sea beaches. J. Insect Conserv. 2015, 19, 791–800. [Google Scholar] [CrossRef]
- Yeargan, K.V. Ecology of a bolas spider, Mastophora hutchinsoni: Phenology, hunting tactics, and evidence for aggressive chemical mimicry. Oecologia 1988, 74, 524–530. [Google Scholar] [CrossRef]
- Yeargan, K.V. Biology of Bolas spiders. Ann. Rev. Entomol. 1994, 39, 81–99. [Google Scholar] [CrossRef]
- Gemeno, C.; Yeargan, K.V.; Haynes, K.F. Aggressive chemical mimicry by the bolas spider Mastaphora hutchinsoni: Identification and quantification of a major prey’s sex pheromone components in the spider’s volatile emissions. J. Chem. Ecol. 2000, 26, 1235–1243. [Google Scholar] [CrossRef]
- Haynes, K.F.; Gemeno, C.; Yeargan, K.V.; Millar, J.G.; Johnson, K.M. Aggressive chemical mimicry of moth pheromones by a bolas spider: How does this specialist predator attract more than one species of prey? Chemoecology 2002, 12, 99–105. [Google Scholar] [CrossRef]
- Yeargan, K.V.; Quate, L.W. Juvenile bolas spiders attract psychodid flies. Oecologia 1996, 106, 266–271. [Google Scholar] [CrossRef]
- Yeargan, K.V.; Quate, L.W. Adult male bolas spiders retain juvenile hunting tactics. Oecologia 1997, 112, 572–576. [Google Scholar] [CrossRef]
- Schulz, S. Spider pheromones—A structural perspective. J. Chem. Ecol. 2013, 39, 1–14. [Google Scholar] [CrossRef]
- Ferguson, D.C. The Moths of America North of Mexico: Fascicle 20.2 A Bombycoidea Saturniidae (Part); EW Classey Limited and RBD Publications: London, UK, 1971; 176p. [Google Scholar]
- Horton, C.C. Apparent attraction of moths by the webs of araneid spiders. J. Arachnol. 1979, 7, 88. [Google Scholar]
- Tuskes, P.M.; Tuttle, J.P.; Collins, M.M. The Wild Silk Moths of North America: A Natural History of the Saturniidae of the United States and Canada; Cornell University Press: Ithaca, NY, USA, 1996; 250p. [Google Scholar]
- Severns, P.M. The effects of a fall prescribed burn on Hemileuca eglanterina Boisduval (Saturniidae). J. Lepid. Soc. 2003, 57, 137–143. [Google Scholar]
- McElfresh, J.S.; Millar, J.G. Geographic variation in the sex pheromone blend of Hemileuca electra from southern California. J. Chem. Ecol. 1999, 25, 2505–2525. [Google Scholar] [CrossRef]
- McElfresh, J.S.; Millar, J.G. Geographic variation in the pheromone system of the saturniid moth Hemileuca eglanterina. Ecology 2001, 82, 3505–3518. [Google Scholar] [CrossRef]
- McElfresh, J.S.; Millar, J.G. Sex pheromone of the saturniid moth, Hemileuca burnsi, from the western Mohave Desert of southern California. J. Chem. Ecol. 2008, 34, 1115–1124. [Google Scholar] [CrossRef] [PubMed]
- McElfresh, J.S.; Millar, J.G. Possible reproductive character displacement in saturniid moths in the genus Hemileuca. In Pheromone Communication in Moths. Evolution, Behavior, and Application; Allison, J.D., Cardé, R.T., Eds.; University of California Press: Oakland, CA, USA, 2016; pp. 225–232. [Google Scholar]
- McElfresh, J.S.; Hammond, A.M.; Millar, J.G. Sex pheromone components of the buck moth Hemileuca maia. J. Chem. Ecol. 2001, 27, 1409–1422. [Google Scholar] [CrossRef] [PubMed]
- Rubinoff, D.; Sperling, F.A.H. Evolution of ecological traits and wing morphology in Hemileuca (Saturniidae) based on a two-gene phylogeny. Mol. Phylogenet. Evol. 2002, 25, 70–86. [Google Scholar] [CrossRef] [PubMed]
- Bradley, R.A. Common Spiders of North America; University of California Press: Oakland, CA, USA, 2012; 288p. [Google Scholar]
- Collins, M.M.; Tuskes, P.M. Reproduction isolation in sympatric species of day-flying moths (Hemileuca: Saturniidae). Evolution 1979, 33, 728–733. [Google Scholar] [CrossRef]
- Tuskes, P.M. The biology and distribution of California Hemileucinae (Saturniidae). J. Lepid. Soc. 1984, 38, 281–309. [Google Scholar]
- Cardé, R.T. Moth navigation along pheromone plumes. In Pheromone Communication in Moths. Evolution, Behavior, and Application; Allison, J.D., Cardé, R.T., Eds.; University of California Press: Oakland, CA, USA, 2016; pp. 173–189. [Google Scholar]
- Craig, C.L.; Bernard, G.D. Insect attraction to ultraviolet-reflecting spider webs and web decorations. Ecology 1990, 71, 616–623. [Google Scholar] [CrossRef]
- Li, D. Spiders that decorate their webs at higher frequency intercept more prey and grow faster. Proc. R. Soc. B 2005, 272, 1753–1757. [Google Scholar] [CrossRef]
- Blamires, S.J.; Hochuli, D.F.; Thompson, M.B. Why cross the web: Decoration spectral properties and prey capture in an orb spider (Argiope keyserlingi) web. Biol. J. Linnean Soc. 2008, 94, 221–229. [Google Scholar] [CrossRef]
- Chinta, S.P.; Goller, S.; Lux, J.; Funke, S.; Uhl, G.; Schulz, S. The sex pheromone of the wasp spider Argiope bruennichi. Angew. Chem. Int. Ed. 2010, 49, 2033–2036. [Google Scholar] [CrossRef]
Figure 1.
Hemileuca phylogeny, as presented by Rubinoff and Sperling [36] (Figure 6), including the proposed clades (1–4) and position of the Hemileuca taxa (pictured) evaluated for male attraction to caged Argiope aurantia. Boxes around each Hemileuca taxon signify whether there was no attraction (red box, and red taxon name with an *), strong attraction (green box with a green taxon name and an *), or a mixture of populations with no or strong attraction (purple hashed border with a purple taxon and an *) to caged A. aurantia (Supplementary Table S1). We did not evaluate Hemileuca attraction from any members of clade 4 and H. magnifica (‡) was originally considered a subspecies of H. hera, so we assume that its position would be in clade 2 with H. hera.
Figure 1.
Hemileuca phylogeny, as presented by Rubinoff and Sperling [36] (Figure 6), including the proposed clades (1–4) and position of the Hemileuca taxa (pictured) evaluated for male attraction to caged Argiope aurantia. Boxes around each Hemileuca taxon signify whether there was no attraction (red box, and red taxon name with an *), strong attraction (green box with a green taxon name and an *), or a mixture of populations with no or strong attraction (purple hashed border with a purple taxon and an *) to caged A. aurantia (Supplementary Table S1). We did not evaluate Hemileuca attraction from any members of clade 4 and H. magnifica (‡) was originally considered a subspecies of H. hera, so we assume that its position would be in clade 2 with H. hera.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Warren, A.D.; Severns, P.M. Fatal Attraction: Argiope Spiders Lure Male Hemileuca Moth Prey with the Promise of Sex. Insects 2024, 15, 53. https://doi.org/10.3390/insects15010053
AMA Style
Warren AD, Severns PM. Fatal Attraction: Argiope Spiders Lure Male Hemileuca Moth Prey with the Promise of Sex. Insects. 2024; 15(1):53. https://doi.org/10.3390/insects15010053
Chicago/Turabian StyleWarren, Andrew D., and Paul M. Severns. 2024. "Fatal Attraction: Argiope Spiders Lure Male Hemileuca Moth Prey with the Promise of Sex" Insects 15, no. 1: 53. https://doi.org/10.3390/insects15010053
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.