

Stability and Assembly Mechanisms of Butterfly Communities across Environmental Gradients of a Subtropical Mountain
 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area, Butterfly Survey and Environmental Factors
2.2. Extraction and Sequencing of COI DNA
2.3. Phylogenetic Placement Tree Reconstruction
2.4. Diversity Patterns
2.5. The Effect of the Complexity and Diversity on Stability under Environmental Changes
2.6. Assembly Mechanisms of Butterfly Communities
2.7. Phylogenetic Barcode Tree Reconstruction
3. Results
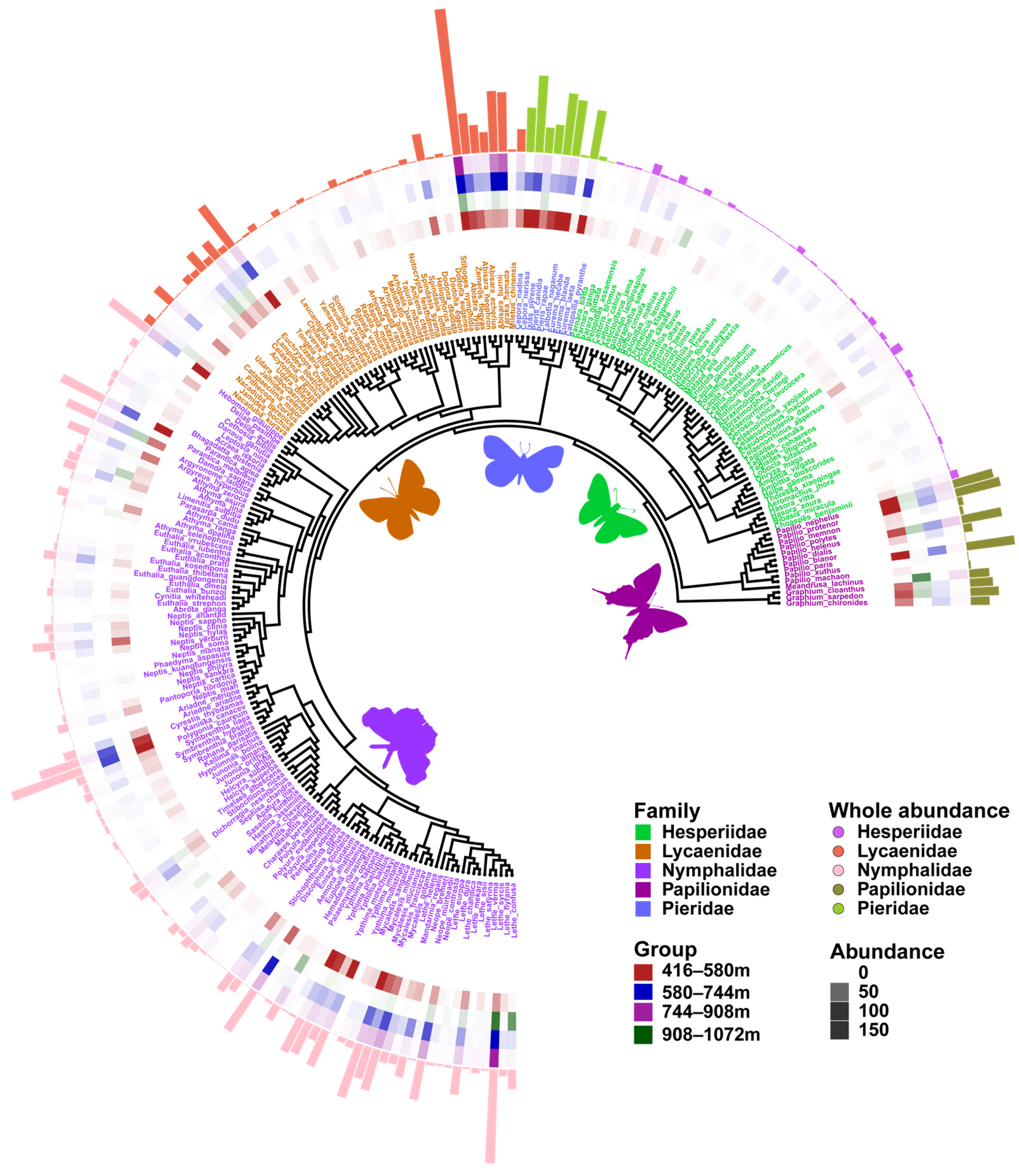
3.1. Diversity Patterns
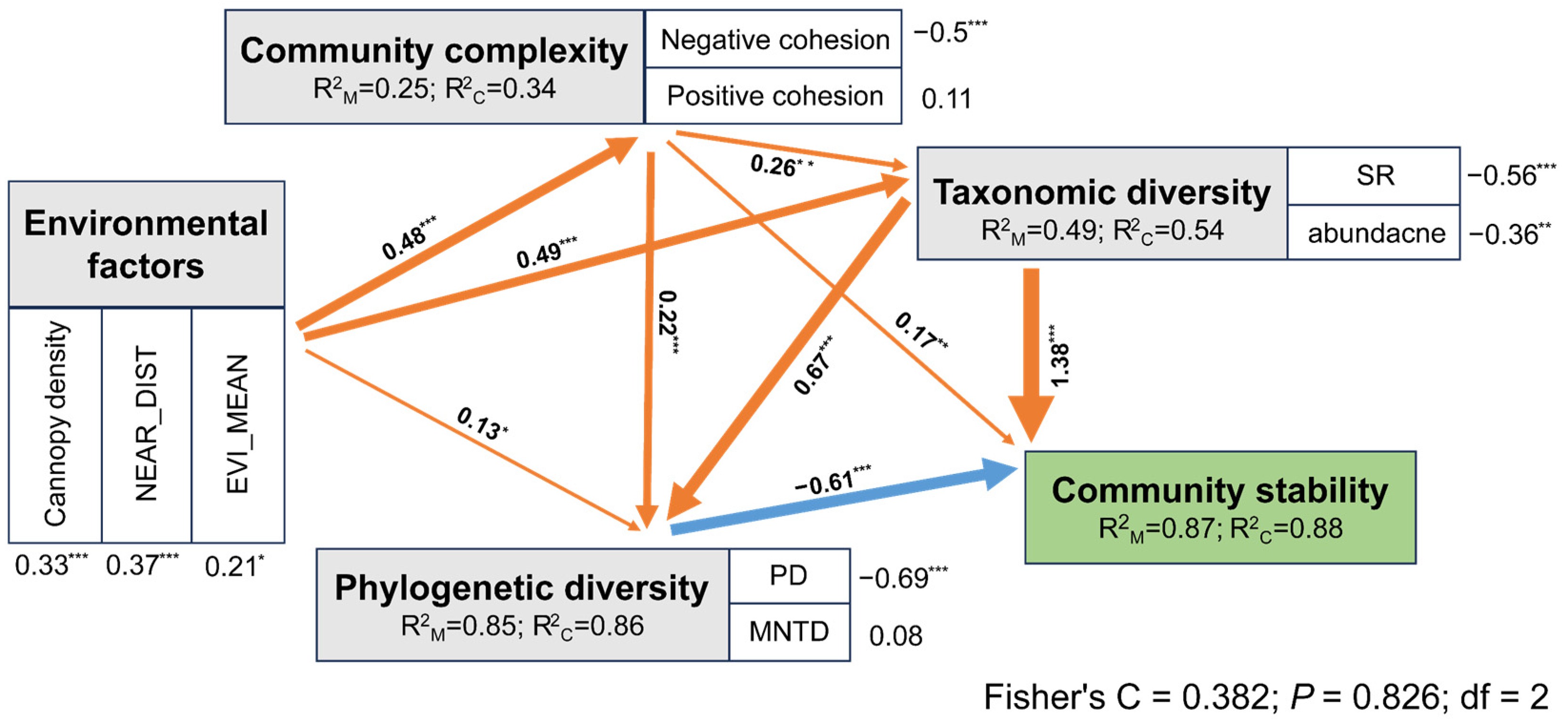
3.2. The Effect of Complexity and Diversity on Stability under Environmental Changes
3.3. Assembly Mechanisms of Butterfly Communities
3.4. Phylogenetic Analyses Using Barcode and Placement Trees
4. Discussion
4.1. Diversity Patterns
4.2. The Effect of Complexity and Diversity on Stability under Environmental Changes
4.3. Assembly Mechanisms of Butterfly Communities
4.4. Phylogenetic Analyses Using Barcode and Placement Trees
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mittermeier, R.A.; Turner, W.R.; Larsen, F.W.; Brooks, T.M.; Gascon, C. Global biodiversity conservation: The critical role of hotspots. In Biodiversity Hotspots; Zachos, F.E., Habel, J.C., Eds.; Springer Publishers: London, UK, 2011; pp. 3–22. [Google Scholar]
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; Da Fonseca, G.A.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef]
- Noss, R.F.; Platt, W.J.; Sorrie, B.A.; Weakley, A.S.; Means, D.B.; Costanza, J.; Peet, R.K. How global biodiversity hotspots may go unrecognized: Lessons from the North American Coastal Plain. Divers. Distrib. 2015, 21, 236–244. [Google Scholar] [CrossRef]
- Wang, S.; Girardello, M.; Zhang, W. Potential and progress of studying mountain biodiversity by means of butterfly genetics and genomics. J. Genet. Genom. 2023, 51, 1673–8527. [Google Scholar] [CrossRef]
- Ashton, L.A.; Nakamura, A.; Basset, Y.; Burwell, C.J.; Cao, M.; Eastwood, R.; Odell, E.; de Oliveira, E.G.; Hurley, K.; Katabuchi, M.; et al. Vertical stratification of moths across elevation and latitude. J. Biogeogr. 2016, 43, 59–69. [Google Scholar] [CrossRef]
- Beck, J.; McCain, C.M.; Axmacher, J.C.; Ashton, L.A.; Bärtschi, F.; Brehm, G.; Choi, S.W.; Cizek, O.; Colwell, R.K.; Fiedler, K.; et al. Elevational species richness gradients in a hyperdiverse insect taxon: A global meta-study on geometrid moths. Glob. Ecol. Biogeogr. 2017, 26, 412–424. [Google Scholar] [CrossRef]
- Szewczyk, T.; McCain, C.M. A systematic review of global drivers of ant elevational diversity. PLoS ONE 2016, 11, e0155404. [Google Scholar] [CrossRef]
- Barberán, A.; Bates, S.T.; Casamayor, E.O.; Fierer, N. Using network analysis to explore co-occurrence patterns in soil microbial communities. ISME J. 2012, 6, 343–351. [Google Scholar] [CrossRef]
- Yuan, M.; Guo, X.; Wu, L.; Zhang, Y.; Xiao, N.; Ning, D.; Shi, Z.; Zhou, X.; Wu, L.; Yang, Y.; et al. Climate warming enhances microbial network complexity and stability. Nat. Clim. Chang. 2021, 11, 343–348. [Google Scholar] [CrossRef]
- Landi, P.; Minoarivelo, H.O.; Brännström, Å.; Hui, C.; Dieckmann, U. Complexity and stability of ecological networks: A review of the theory. Popul. Ecol. 2018, 60, 319–345. [Google Scholar] [CrossRef]
- Okuyama, T.; Holland, J.N. Network structural properties mediate the stability of mutualistic communities. Ecol. Lett. 2008, 11, 208–216. [Google Scholar] [CrossRef]
- Ives, A.R.; Klug, J.L.; Gross, K. Stability and species richness in complex communities. Ecol. Lett. 2000, 3, 399–411. [Google Scholar] [CrossRef]
- Grossiord, C.; Granier, A.; Ratcliffe, S.; Bouriaud, O.; Bruelheide, H.; Chećko, E.; Forrester, D.I.; Dawud, S.M.; Finér, L.; Pollastrini, M.; et al. Tree diversity does not always improve resistance of forest ecosystems to drought. Proc. Natl. Acad. Sci. USA 2014, 111, 14812–14815. [Google Scholar] [CrossRef]
- de la Riva, E.G.; Lloret, F.; Pérez-Ramos, I.M.; Marañón, T.; Saura-Mas, S.; Díaz-Delgado, R.; Villar, R. The importance of functional diversity in the stability of Mediterranean shrubland communities after the impact of extreme climatic events. J. Plant Ecol. 2017, 10, 281–293. [Google Scholar] [CrossRef]
- Ma, B.; Zhao, K.; Lv, X.; Su, W.; Dai, Z.; Gilbert, J.A.; Brookes, P.C.; Faust, K.; Xu, J. Genetic correlation network prediction of forest soil microbial functional organization. ISME J. 2018, 12, 2492–2505. [Google Scholar] [CrossRef]
- Stegen, J.C.; Lin, X.; Konopka, A.E.; Fredrickson, J.K. Stochastic and deterministic assembly processes in subsurface microbial communities. ISME J. 2012, 6, 1653–1664. [Google Scholar] [CrossRef]
- Liu, W.; Liu, L.; Yang, X.; Deng, M.; Wang, Z.; Wang, P.; Yang, S.; Li, P.; Peng, Z.; Yang, L. Long-term nitrogen input alters plant and soil bacterial, but not fungal beta diversity in a semiarid grassland. Glob. Chang. Biol. 2021, 27, 3939–3950. [Google Scholar] [CrossRef]
- Zhong, Y.; Sorensen, P.O.; Zhu, G.; Jia, X.; Liu, J.; Shangguan, Z.; Wang, R.; Yan, W. Differential Microbial Assembly Processes and Co-Occurrence Networks in the Soil-root Continuum Along an Environmental Gradient. iMeta 2022, 1, e18. [Google Scholar] [CrossRef]
- Xu, M.J.; Zhu, X.Z.; Chen, S.P.; Pang, S.; Liu, W.; Gao, L.L.; Yang, W.; Li, T.T.; Zhang, Y.H.; Luo, C.; et al. Distinctive Pattern and Mechanism of Precipitation Changes Affecting Soil Microbial Assemblages in the Eurasian Steppe. iScience 2022, 25, 103893. [Google Scholar] [CrossRef]
- Webb, C.O. Exploring the phylogenetic structure of ecological communities: An example for rain forest trees. Am. Nat. 2000, 156, 145–155. [Google Scholar] [CrossRef]
- Hao, M.; Jin, Q.; Meng, G.; Yang, C.; Yang, S.; Shi, Z.; Tang, M.; Liu, S.; Li, Y.; Li, J. Using full-length metabarcoding and DNA barcoding to infer community assembly for speciose taxonomic groups: A case study. Evol. Ecol. 2020, 34, 1063–1088. [Google Scholar] [CrossRef]
- Liu, J.; Liu, J.; Ge, X.; Huang, G.; Zhou, Z.; Wan, S. The use of DNA barcoding to assess phylogenetic β-diversity in mid-subtropical evergreen broad-leaved forests of China. Forests 2019, 10, 923. [Google Scholar] [CrossRef]
- Qian, H.; Jin, Y. Are phylogenies resolved at the genus level appropriate for studies on phylogenetic structure of species assemblages? Plant Divers. 2021, 43, 255–263. [Google Scholar] [CrossRef]
- Li, Z.; Linard, B.; Vogler, A.P.; Yu, D.W.; Wang, Z. Phylogenetic diversity only weakly mitigates climate-change-driven biodiversity loss in insect communities. Mol. Ecol. 2023, 32, 6147–6160. [Google Scholar] [CrossRef]
- Jin, L.; Liu, J.J.; Xiao, T.W.; Li, Q.M.; Lin, L.X.; Shao, X.N.; Ma, C.X.; Li, B.H.; Mi, X.C.; Ren, H.B. Plastome-based phylogeny improves community phylogenetics of subtropical forests in China. Mol. Ecol. Resour. 2022, 22, 319–333. [Google Scholar] [CrossRef]
- Ivushkin, K.; Bartholomeus, H.; Bregt, A.K.; Pulatov, A.; Kempen, B.; De Sousa, L. Global mapping of soil salinity change. Remote Sens. Environ. 2019, 231, 111260. [Google Scholar] [CrossRef]
- Abrego, N.; Roslin, T.; Huotari, T.; Ji, Y.; Schmidt, N.M.; Wang, J.; Yu, D.W.; Ovaskainen, O. Accounting for species interactions is necessary for predicting how arctic arthropod communities respond to climate change. Ecography 2021, 44, 885–896. [Google Scholar] [CrossRef]
- Srivathsan, A.; Ang, Y.; Heraty, J.M.; Hwang, W.S.; Jusoh, W.F.A.; Kutty, S.N.; Puniamoorthy, J.; Yeo, D.; Roslin, T.; Meier, R. Convergence of dominance and neglect in flying insect diversity. Nat. Ecol. Evol. 2023, 7, 1012–1021. [Google Scholar] [CrossRef]
- Lawton, J.H.; Bignell, D.E.; Bolton, B.; Bloemers, G.F.; Eggleton, P.; Hammond, P.M.; Hodda, M.; Holt, R.D.; Larsen, T.B.; Mawdsley, N.A.; et al. Biodiversity inventories, indicator taxa and effects of habitat modification in tropical forest. Nature 1998, 391, 72–76. [Google Scholar] [CrossRef]
- Oliver, T.H.; Marshall, H.H.; Morecroft, M.D.; Brereton, T.; Prudhomme, C.; Huntingford, C. Interacting effects of climate change and habitat fragmentation on drought-sensitive butterflies. Nat. Clim. Chang. 2015, 5, 941–945. [Google Scholar] [CrossRef]
- Wu, C.S.; Hsu, Y.F. Butterflies of China; Straits Publishing House: Fujian, China, 2017. [Google Scholar]
- Xiao, Z. Inventory and Assessment of Wildlife and Its Habitat in Protected Areas—An Example from Chebaling National Nature Reserve, Guangdong, China; China Forestry Publishing House: Beijing, China, 2019. [Google Scholar]
- Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar]
- Hebert, P.D.; Penton, E.H.; Burns, J.M.; Janzen, D.H.; Hallwachs, W. Ten species in one: DNA barcoding reveals cryptic species in the neotropical skipper butterfly Astraptes fulgerator. Proc. Natl. Acad. Sci. USA 2004, 101, 14812–14817. [Google Scholar] [CrossRef]
- Wang, M.Q.; Li, Y.; Chesters, D.; Anttonen, P.; Bruelheide, H.; Chen, J.T.; Durka, W.; Guo, P.F.; Härdtle, W.; Ma, K.P.; et al. Multiple components of plant diversity loss determine herbivore phylogenetic diversity in a subtropical forest experiment. J. Ecol. 2019, 107, 2697–2712. [Google Scholar] [CrossRef]
- Swindell, S.R.; Plasterer, T.N. SEQMAN. Contig Assembly. Methods Mol. Biol. 1997, 70, 75–89. [Google Scholar]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Chesters, D. The phylogeny of insects in the data-driven era. Syst. Entomol. 2020, 45, 540–551. [Google Scholar] [CrossRef]
- Chesters, D.; Ferrari, R.R.; Lin, X.; Orr, M.C.; Staab, M.; Zhu, C.D. Launching insectphylo. org; a new hub facilitating construction and use of synthesis molecular phylogenies of insects. Mol. Ecol. Resour. 2023, 23, 1556–1573. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Magurran, A.E. Biological diversity. Curr. Biol. 2005, 15, R116–R118. [Google Scholar] [CrossRef]
- Faith, D.P. Conservation evaluation and phylogenetic diversity. Biol. Conserv. 1992, 61, 1–10. [Google Scholar] [CrossRef]
- Webb, C.O.; Ackerly, D.D.; McPeek, M.A.; Donoghue, M.J. Phylogenies and community ecology. Annu. Rev. Ecol. Syst. 2002, 33, 475–505. [Google Scholar] [CrossRef]
- Breiman, L. Random forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef]
- Chao, A.; Gotelli, N.J.; Hsieh, T.C.; Sander, E.L.; Ma, K.H.; Colwell, R.K.; Ellison, A.M. Rarefaction and extrapolation with Hill numbers: A framework for sampling and estimation in species diversity studies. Ecol. Monogr. 2014, 84, 45–67. [Google Scholar] [CrossRef]
- Baselga, A. Partitioning the turnover and nestedness components of beta diversity. Glob. Ecol. Biogeogr. 2010, 19, 134–143. [Google Scholar] [CrossRef]
- Leprieur, F.; Albouy, C.; De Bortoli, J.; Cowman, P.F.; Bellwood, D.R.; Mouillot, D. Quantifying phylogenetic beta diversity: Distinguishing between ‘true’turnover of lineages and phylogenetic diversity gradients. PLoS ONE 2012, 7, e42760. [Google Scholar] [CrossRef]
- Cardoso, P.; Rigal, F.; Carvalho, J.C.; Fortelius, M.; Borges, P.A.V.; Podani, J.; Schmera, D. Partitioning taxon, phylogenetic and functional beta diversity into replacement and richness difference components. J. Biogeogr. 2014, 41, 749–761. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022. [Google Scholar]
- Herren, C.M.; McMahon, K.D. Cohesion: A method for quantifying the connectivity of microbial communities. ISME J. 2017, 11, 2426–2438. [Google Scholar] [CrossRef]
- Xun, W.; Liu, Y.; Li, W.; Ren, Y.; Xiong, W.; Xu, Z.; Zhang, N.; Miao, Y.; Shen, Q.; Zhang, R. Specialized metabolic functions of keystone taxa sustain soil microbiome stability. Microbiome 2021, 9, 35. [Google Scholar] [CrossRef]
- Lefcheck, J.S. piecewiseSEM: Piecewise structural equation modelling in r for ecology, evolution, and systematics. Methods Ecol. Evol. 2016, 7, 573–579. [Google Scholar] [CrossRef]
- MacNally, R.; Walsh, C.J. Hierarchical partitioning public-domain software. Biol. Conserv. 2004, 13, 659–660. [Google Scholar]
- Sloan, W.T.; Lunn, M.; Woodcock, S.; Head, I.M.; Nee, S.; Curtis, T.P. Quantifying the roles of immigration and chance in shaping prokaryote community structure. Environ. Microbiol. 2006, 8, 732–740. [Google Scholar] [CrossRef]
- Burns, A.R.; Stephens, W.Z.; Stagaman, K.; Wong, S.; Rawls, J.F.; Guillemin, K.; Bohannan, B.J. Contribution of neutral processes to the assembly of gut microbial communities in the zebrafish over host development. ISME J. 2016, 10, 655–664. [Google Scholar] [CrossRef]
- Stegen, J.C.; Lin, X.; Fredrickson, J.K.; Chen, X.; Kennedy, D.W.; Murray, C.J.; Rockhold, M.L.; Konopka, A. Quantifying community assembly processes and identifying features that impose them. ISME J. 2013, 7, 2069–2079. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Castresana, J. Selection of conserved blocks from multiple alignments for their use in phylogenetic analysis. Mol. Biol. Evol. 2000, 17, 540–552. [Google Scholar] [CrossRef]
- Hall, T.; Biosciences, I.; Carlsbad, C. BioEdit: An important software for molecular biology. GERF Bull. Biosci. 2011, 2, 60–61. [Google Scholar]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.; Von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef]
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; Von Haeseler, A.; Lanfear, R. IQ-TREE 2: New models and efficient methods for phylogenetic inference in the genomic era. Mol. Biol. Evol. 2020, 37, 1530–1534. [Google Scholar] [CrossRef]
- Hoang, D.T.; Chernomor, O.; Von Haeseler, A.; Minh, B.Q.; Vinh, L.S. UFBoot2: Improving the ultrafast bootstrap approximation. Mol. Biol. Evol. 2018, 35, 518–522. [Google Scholar] [CrossRef]
- Dove, A.D.; Cribb, T.H. Species accumulation curves and their applications in parasite ecology. Trends Parasitol. 2006, 22, 568–574. [Google Scholar] [CrossRef]
- Hsieh, T.C.; Ma, K.H.; Chao, A. iNEXT: An R package for rarefaction and extrapolation of species diversity (Hill numbers). Methods Ecol. Evol. 2016, 7, 1451–1456. [Google Scholar] [CrossRef]
- Camero, É.; Calderón, A.M. Comunidad de mariposas diurnas (Lepidoptera: Rhopalocera) en un gradiente altitudinal del cañón del río Combeimatolima, Colombia. Acta Biolo Colomb. 2007, 12, 95–110. [Google Scholar]
- Carneiro, E.; Mielke, O.; Casagrande, M.; Fiedler, K. Skipper richness (Hesperiidae) along elevational gradients in Brazilian Atlantic Forest. Neotrop. Entomol. 2014, 43, 27–38. [Google Scholar] [CrossRef]
- Lai, Y.; Liu, Y.; Liu, X. Elevational diversity patterns of green lacewings (Neuroptera: Chrysopidae) uncovered with DNA barcoding in a biodiversity hotspot of Southwest China. Front. Ecol. Evol. 2021, 9, 778686. [Google Scholar] [CrossRef]
- Liu, S.; Meng, Q.; Guo, W. Moth diversity in the northern slope of Changbai Mountain Nature Reserve. J. Northeast. For. Univ. 2007, 35, 51. [Google Scholar]
- Zou, Y.; Sang, W.; Zhou, H.; Huang, L.; Axmacher, J.C. Altitudinal diversity patterns of ground beetles (Coleoptera: Carabidae) in the forests of Changbai Mountain, Northeast China. Insect Conserv. Divers. 2014, 7, 161–171. [Google Scholar] [CrossRef]
- Lyu, H.R.; Liu, S.S.; Zhu, J.Y.; Ye, Y.C.; Chen, H.Y.; Mo, L.J. Effects of human disturbance on understory woody species composition and diversity in fengshui forests. Sheng Wu Duo Yang Xing 2009, 17, 458–467. [Google Scholar]
- Chowdhury, S.; Dubey, V.K.; Choudhury, S.; Das, A.; Jeengar, D.; Sujatha, B.; Kumar, A.; Kumar, N.; Semwal, A.; Kumar, V. Insects as bioindicator: A hidden gem for environmental monitoring. Front. Environ. Sci. 2023, 11, 273. [Google Scholar] [CrossRef]
- Chamberlain, D.; Brambilla, M.; Caprio, E.; Pedrini, P.; Rolando, A. Alpine bird distributions along elevation gradients: The consistency of climate and habitat effects across geographic regions. Oecologia 2016, 181, 1139–1150. [Google Scholar] [CrossRef]
- McCain, C.M. Could temperature and water availability drive elevational species richness patterns? A global case study for bats. Glob. Ecol. Biogeogr. 2007, 16, 1–13. [Google Scholar] [CrossRef]
- Montaño-Centellas, F.A.; Loiselle, B.A.; Tingley, M.W. Ecological drivers of avian community assembly along a tropical elevation gradient. Ecography 2021, 44, 574–588. [Google Scholar] [CrossRef]
- Nunes, C.A.; Braga, R.F.; Figueira, J.E.; Neves, F.d.S.; Fernandes, G.W. Dung beetles along a tropical altitudinal gradient: Environmental filtering on taxonomic and functional diversity. PLoS ONE 2016, 11, e0157442. [Google Scholar] [CrossRef]
- Pires, A.C.V.; Barbosa, M.; Beiroz, W.; BeirÃo, M.V.; Marini-Filho, O.J.; Duarte, M.; Mielke, O.H.; Ladeira, F.A.; Nunes, Y.R.; Negreiros, D.; et al. Altitudinal variation in butterfly community associated with climate and vegetation. An. Acad. Bras. Cienc. 2020, 92, e20190058. [Google Scholar] [CrossRef]
- Wang, X.; Wiegand, T.; Anderson-Teixeira, K.J.; Bourg, N.A.; Hao, Z.; Howe, R.; Jin, G.; Orwig, D.A.; Spasojevic, M.J.; Wang, S.; et al. Ecological drivers of spatial community dissimilarity, species replacement and species nestedness across temperate forests. Glob. Ecol. Biogeogr. 2018, 27, 581–592. [Google Scholar] [CrossRef]
- Liu, C.; Dudley, K.L.; Xu, Z.H.; Economo, E.P. Mountain metacommunities: Climate and spatial connectivity shape ant diversity in a complex landscape. Ecography 2018, 41, 101–112. [Google Scholar] [CrossRef]
- García-Palacios, P.; Gross, N.; Gaitán, J.; Maestre, F.T. Climate mediates the biodiversity–ecosystem stability relationship globally. Proc. Natl. Acad. Sci. USA 2018, 115, 8400–8405. [Google Scholar] [CrossRef]
- Loreau, M.; De Mazancourt, C. Biodiversity and ecosystem stability: A synthesis of underlying mechanisms. Ecol. Lett. 2013, 16, 106–115. [Google Scholar] [CrossRef]
- Ratzke, C.; Barrere, J.; Gore, J. Strength of species interactions determines biodiversity and stability in microbial communities. Nat. Ecol. Evol. 2020, 4, 376–383. [Google Scholar] [CrossRef]
- Shen, Z.; Yu, B.; Shao, K.; Gao, G.; Tang, X. Warming reduces microeukaryotic diversity, network complexity and stability. Environ. Res. 2023, 238, 117235. [Google Scholar] [CrossRef]
- Wagg, C.; Dudenhöffer, J.H.; Widmer, F.; Van Der Heijden, M.G. Linking diversity, synchrony and stability in soil microbial communities. Funct. Ecol. 2018, 32, 1280–1292. [Google Scholar] [CrossRef]
- Cadotte, M.W.; Dinnage, R.; Tilman, D. Phylogenetic diversity promotes ecosystem stability. Ecology 2012, 93, S223–S233. [Google Scholar] [CrossRef]
- Kelly, S.; Grenyer, R.; Scotland, R.W. Phylogenetic trees do not reliably predict feature diversity. Divers. Distrib. 2014, 20, 600–612. [Google Scholar] [CrossRef]
- Craven, D.; Eisenhauer, N.; Pearse, W.D.; Hautier, Y.; Isbell, F.; Roscher, C.; Bahn, M.; Beierkuhnlein, C.; Bönisch, G.; Buchmann, N.; et al. Multiple facets of biodiversity drive the diversity–stability relationship. Nat. Ecol. Evol. 2018, 2, 1579–1587. [Google Scholar] [CrossRef]
- Fernandez-Going, B.M.; Anacker, B.L.; Harrison, S.P. Temporal variability in California grasslands: Soil type and species functional traits mediate response to precipitation. Ecology 2012, 93, 2104–2114. [Google Scholar] [CrossRef]
- Polley, H.W.; Isbell, F.I.; Wilsey, B.J. Plant functional traits improve diversity-based predictions of temporal stability of grassland productivity. Oikos 2013, 122, 1275–1282. [Google Scholar] [CrossRef]
- Montoya, J.M.; Pimm, S.L.; Solé, R.V. Ecological networks and their fragility. Nature 2006, 442, 259–264. [Google Scholar] [CrossRef]
- Shen, C.; Wang, J.; Jing, Z.; Qiao, N.H.; Xiong, C.; Ge, Y. Plant diversity enhances soil fungal network stability indirectly through the increase of soil carbon and fungal keystone taxa richness. Sci. Total Environ. 2022, 818, 151737. [Google Scholar] [CrossRef]
- Hernandez, D.J.; David, A.S.; Menges, E.S.; Searcy, C.A.; Afkhami, M.E. Environmental stress destabilizes microbial networks. ISME J. 2021, 15, 1722–1734. [Google Scholar] [CrossRef]
- Hautier, Y.; Tilman, D.; Isbell, F.; Seabloom, E.W.; Borer, E.T.; Reich, P.B. Anthropogenic environmental changes affect ecosystem stability via biodiversity. Science 2015, 348, 336–340. [Google Scholar] [CrossRef]
- Lepš, J.; Májeková, M.; Vítová, A.; Doležal, J.; de Bello, F. Stabilizing effects in temporal fluctuations: Management, traits, and species richness in high-diversity communities. Ecology 2018, 99, 360–371. [Google Scholar] [CrossRef]
- Dini-Andreote, F.; Stegen, J.C.; Van Elsas, J.D.; Salles, J.F. Disentangling mechanisms that mediate the balance between stochastic and deterministic processes in microbial succession. Proc. Natl. Acad. Sci. USA 2015, 112, E1326–E1332. [Google Scholar] [CrossRef]
- Chen, W.; Jiao, S.; Li, Q.; Du, N. Dispersal limitation relative to environmental filtering governs the vertical small-scale assembly of soil microbiomes during restoration. J. Appl. Ecol. 2020, 57, 402–412. [Google Scholar] [CrossRef]
- Kerfahi, D.; Tripathi, B.M.; Slik, J.F.; Sukri, R.S.; Jaafar, S.; Adams, J.M. Distinctive soil archaeal communities in different variants of tropical equatorial forest. Microb. Ecol. 2018, 76, 215–225. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, B.; Liu, Y.; Guo, Y.; Shi, P.; Wei, G. Distinct large-scale biogeographic patterns of fungal communities in bulk soil and soybean rhizosphere in China. Sci. Total Environ. 2018, 644, 791–800. [Google Scholar] [CrossRef]
- DeLong, J.P.; Okie, J.G.; Moses, M.E.; Sibly, R.M.; Brown, J.H. Shifts in metabolic scaling, production, and efficiency across major evolutionary transitions of life. Proc. Natl. Acad. Sci. USA 2010, 107, 12941–12945. [Google Scholar] [CrossRef]
- Gu, S.; Xiong, X.; Tan, L.; Deng, Y.; Du, X.; Yang, X.; Hu, Q. Soil microbial community assembly and stability are associated with potato (Solanum tuberosum L.) fitness under continuous cropping regime. Front. Plant Sci. 2022, 13, 1000045. [Google Scholar] [CrossRef]
- Shi, Y.; Li, Y.; Xiang, X.; Sun, R.; Yang, T.; He, D.; Zhang, K.; Ni, Y.; Zhu, Y.-G.; Adams, J.M.; et al. Spatial scale affects the relative role of stochasticity versus determinism in soil bacterial communities in wheat fields across the North China Plain. Microbiome 2018, 6, 27. [Google Scholar] [CrossRef]
- Bryant, J.A.; Lamanna, C.; Morlon, H.; Kerkhoff, A.J.; Enquist, B.J.; Green, J.L. Microbes on mountainsides: Contrasting elevational patterns of bacterial and plant diversity. Proc. Natl. Acad. Sci. USA 2008, 105, 11505–11511. [Google Scholar] [CrossRef]
- Li, W.; Kuzyakov, Y.; Zheng, Y.; Li, P.; Li, G.; Liu, M.; Alharbi, H.A.; Li, Z. Depth effects on bacterial community assembly processes in paddy soils. Soil. Biol. Biochem. 2022, 165, 108517. [Google Scholar] [CrossRef]
- Zhou, J.; Ning, D. Stochastic community assembly: Does it matter in microbial ecology? Microbiol. Mol. Biol. Rev. 2017, 81, 10–1128. [Google Scholar] [CrossRef]
- Sun, W.; Xiao, E.; Pu, Z.; Krumins, V.; Dong, Y.; Li, B.; Hu, M. Paddy soil microbial communities driven by environment-and microbe-microbe interactions: A case study of elevation-resolved microbial communities in a rice terrace. Sci. Total Environ. 2018, 612, 884–893. [Google Scholar] [CrossRef]
- Henriques, N.R.; Lourenço, G.M.; Diniz, E.S.; Cornelissen, T. Is elevation a strong environmental filter? Combining taxonomy, functional traits and phylogeny of butterflies in a tropical mountain. Ecol. Entomol. 2022, 47, 613–625. [Google Scholar] [CrossRef]
- Caruso, T.; Chan, Y.; Lacap, D.C.; Lau, M.C.; McKay, C.P.; Pointing, S.B. Stochastic and deterministic processes interact in the assembly of desert microbial communities on a global scale. ISME J. 2011, 5, 1406–1413. [Google Scholar] [CrossRef]
- Chen, W.; Ren, K.; Isabwe, A.; Chen, H.; Liu, M.; Yang, J. Stochastic processes shape microeukaryotic community assembly in a subtropical river across wet and dry seasons. Microbiome 2019, 7, 138. [Google Scholar]
- Hu, A.; Ju, F.; Hou, L.; Li, J.; Yang, X.; Wang, H.; Mulla, S.I.; Sun, Q.; Bürgmann, H.; Yu, C.P. Strong impact of anthropogenic contamination on the co-occurrence patterns of a riverine microbial community. Environ. Microbiol. 2017, 19, 4993–5009. [Google Scholar] [CrossRef]
- Pandit, S.N.; Kolasa, J.; Cottenie, K. Contrasts between habitat generalists and specialists: An empirical extension to the basic metacommunity framework. Ecology 2009, 90, 2253–2262. [Google Scholar] [CrossRef]
- Xu, Y.; Liu, J.J.; Li, H.N.; Liu, J.; Burgess, K.S.; Ge, X.J. The effects of evolutionary and environmental variance on estimates of phylogenetic diversity in temperate forest plots. J. Plant Ecol. 2021, 14, 96–107. [Google Scholar] [CrossRef]
- Jantzen, J.R.; Whitten, W.M.; Neubig, K.M.; Majure, L.C.; Soltis, D.E.; Soltis, P.S. Effects of taxon sampling and tree reconstruction methods on phylodiversity metrics. Ecol. Evol. 2019, 9, 9479–9499. [Google Scholar] [CrossRef]
- Forest, F.; Grenyer, R.; Rouget, M.; Davies, T.J.; Cowling, R.M.; Faith, D.P.; Balmford, A.; Manning, J.C.; Procheş, Ş.; van der Bank, M.; et al. Preserving the evolutionary potential of floras in biodiversity hotspots. Nature 2007, 445, 757–760. [Google Scholar] [CrossRef]
- Hollingsworth, P.M.; Li, D.-Z.; van der Bank, M.; Twyford, A.D. Telling plant species apart with DNA: From barcodes to genomes. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2016, 371, 20150338. [Google Scholar] [CrossRef]
- Liu, J.; Yan, H.F.; Newmaster, S.G.; Pei, N.; Ragupathy, S.; Ge, X.J. The use of DNA barcoding as a tool for the conservation biogeography of subtropical forests in China. Divers. Distrib. 2015, 21, 188–199. [Google Scholar] [CrossRef]
- Webster, A.J.; Payne, R.J.; Pagel, M. Molecular phylogenies link rates of evolution and speciation. Science 2003, 301, 478. [Google Scholar] [CrossRef]
- Münkemüller, T.; Gallien, L.; Pollock, L.J.; Barros, C.; Carboni, M.; Chalmandrier, L.; Mazel, F.; Mokany, K.; Roquet, C.; Smyčka, J.; et al. Dos and don’ts when inferring assembly rules from diversity patterns. Glob. Ecol. Biogeogr. 2020, 29, 1212–1229. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wei, F.; Xie, T.; Su, C.; He, B.; Shu, Z.; Zhang, Y.; Xiao, Z.; Hao, J. Stability and Assembly Mechanisms of Butterfly Communities across Environmental Gradients of a Subtropical Mountain. Insects 2024, 15, 230. https://doi.org/10.3390/insects15040230
Wei F, Xie T, Su C, He B, Shu Z, Zhang Y, Xiao Z, Hao J. Stability and Assembly Mechanisms of Butterfly Communities across Environmental Gradients of a Subtropical Mountain. Insects. 2024; 15(4):230. https://doi.org/10.3390/insects15040230
Chicago/Turabian StyleWei, Fanyu, Tingting Xie, Chengyong Su, Bo He, Zufei Shu, Yingming Zhang, Zhishu Xiao, and Jiasheng Hao. 2024. "Stability and Assembly Mechanisms of Butterfly Communities across Environmental Gradients of a Subtropical Mountain" Insects 15, no. 4: 230. https://doi.org/10.3390/insects15040230