
Genetic Diversity and Population Structure of the Invasive Oriental Fruit Fly, Bactrocera dorsalis (Diptera: Tephritidae) in Burkina Faso

 , , ,
, , ,  ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Molecular Analyses
2.3. Data analysis
2.3.1. Flies Abundance, Genetic Diversity and Genetic Relations Analysis
2.3.2. Analysis of Genetic Structure
3. Results
3.1. Abundance and Diversity
3.1.1. Abundance of Bactrocera dorsalis Pests in Burkina Faso
3.1.2. Polymorphism of Microsatellite Markers and Genetic Diversity
3.2. Population Structure
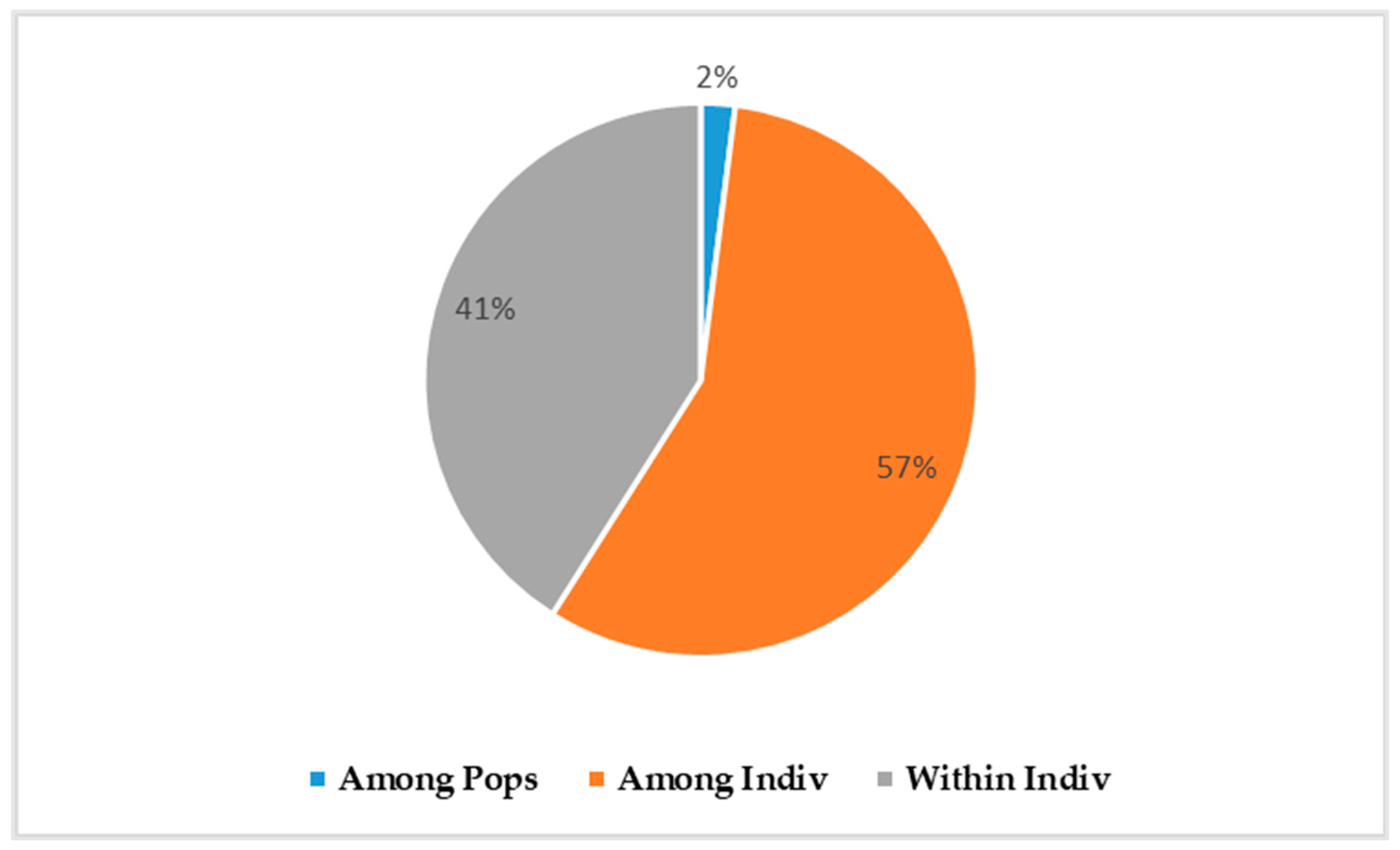
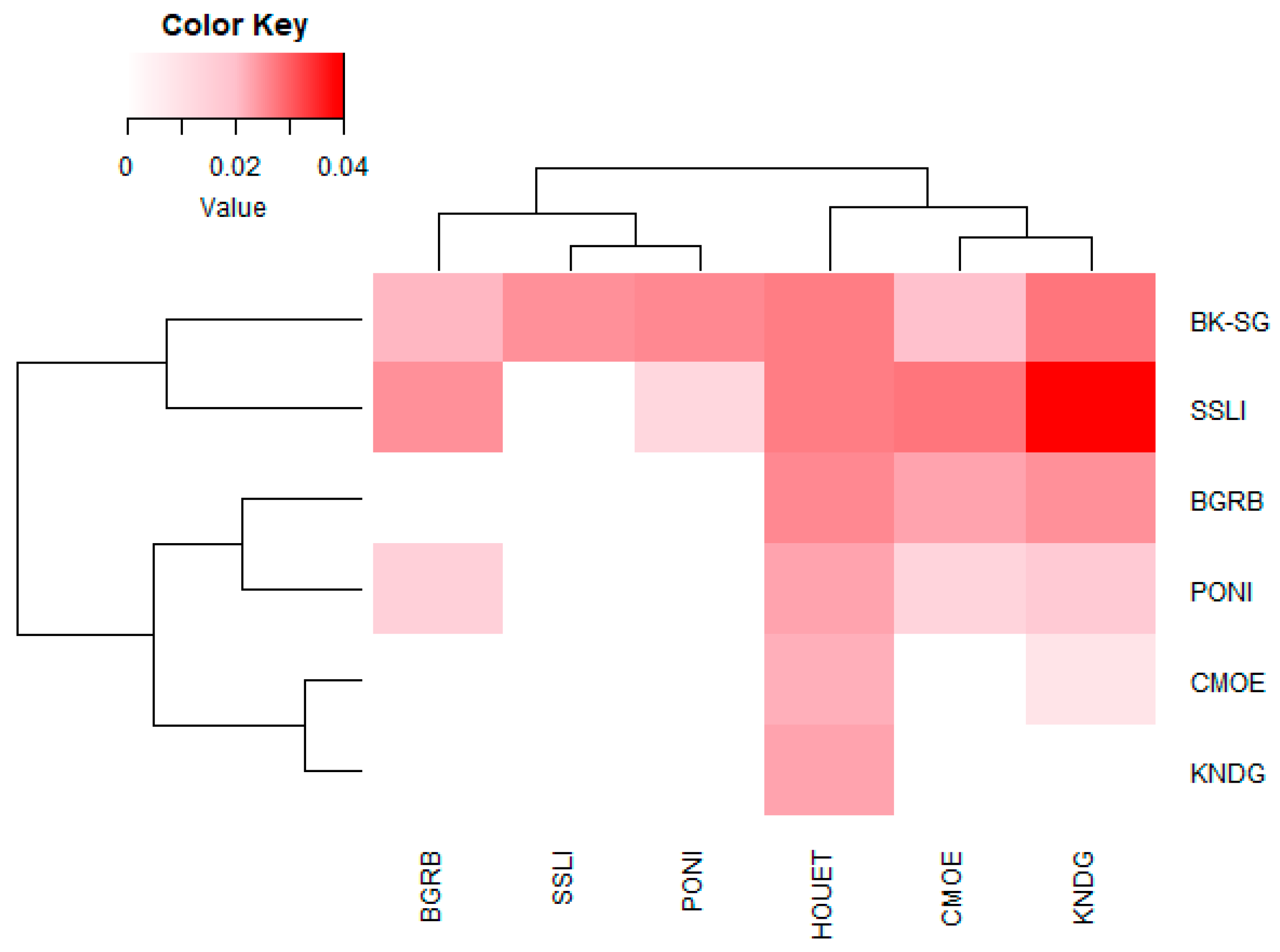
3.2.1. Genetic Differentiation
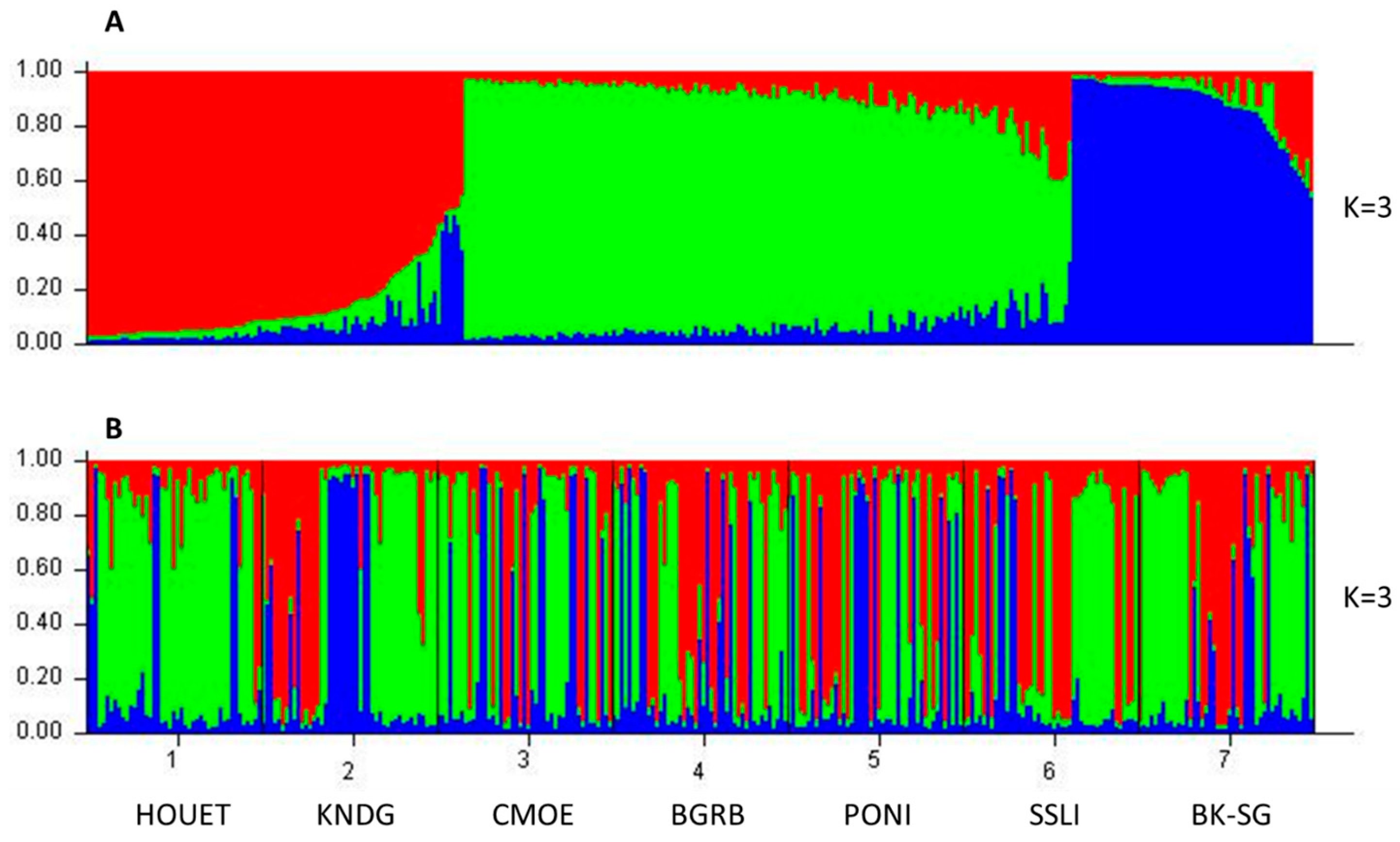
3.2.2. Multivariate Analysis and Genetic Clustering
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zida, I.; Nacro, S.; Dabiré, R.; Moquet, L.; Delatte, H.; Somda, I. Host range and species diversity of Tephritidae of three plant formations in Western Burkina Faso. Bull. Èntomol. Res. 2020, 110, 732–742. [Google Scholar] [CrossRef] [PubMed]
- IPPC. ISPM 27 Diagnostic Protocols for Regulated Pests DP29: Bactrocera dorsalis; International Plant Protection Convention: Rome, Italy, 2019; Volume ISPM 27, p. 34. [Google Scholar]
- Liu, H.; Zhang, D.; Xu, Y.; Wang, L.; Cheng, D.; Qi, Y.; Zeng, L.; Lu, Y. Invasion, expansion, and control of Bactrocera dorsalis (Hendel) in China. J. Integr. Agric. 2019, 18, 771–787. [Google Scholar] [CrossRef]
- Dohino, T.; Hallman, G.J.; Grout, T.G.; Clarke, A.R.; Follett, P.A.; Cugala, D.R.; Tu, D.M.; Murdita, W.; Hernandez, E.; Pereira, R.; et al. Phytosanitary Treatments against Bactrocera dorsalis (Diptera: Tephritidae): Current Situation and Future Prospects. J. Econ. Èntomol. 2016, 110, 67–79. [Google Scholar] [CrossRef]
- Qin, Y.; Krosch, M.N.; Schutze, M.K.; Zhang, Y.; Wang, X.; Prabhakar, C.S.; Susanto, A.; Hee, A.K.W.; Ekesi, S.; Badji, K.; et al. Population structure of a global agricultural invasive pest, Bactrocera dorsalis (Diptera: Tephritidae). Evol. Appl. 2018, 11, 1990–2003. [Google Scholar] [CrossRef] [PubMed]
- Ekesi, S.; De Meyer, M.; Malacrida, A. Taxonomy, Bioecology, and Management of Native and Exotic Fruit Fly Species in Africa. Annu. Rev. Entomol. 2015, 61, 219–238. [Google Scholar] [CrossRef] [PubMed]
- Loomans, A.; Diakaki, M.; Kinkar, M.; Schenk, M.; Vos, S. Pest survey card on Bactrocera dorsalis. EFSA Support. Publ. 2019, 16, 1714E. [Google Scholar] [CrossRef]
- Vargas, R.I.; Piñero, J.C.; Leblanc, L. An Overview of Pest Species of Bactrocera Fruit Flies (Diptera: Tephritidae) and the Integration of Biopesticides with Other Biological Approaches for Their Management with a Focus on the Pacific Region. Insects 2015, 6, 297–318. [Google Scholar] [CrossRef] [PubMed]
- Schutze, M.K.; Aketarawong, N.; Amornsak, W.; Armstrong, K.F.; Augustinos, A.A.; Barr, N.; Bo, W.; Bourtzis, K.; Boykin, L.M.; Cáceres, C.; et al. Synonymization of key pest species within the Bactrocera dorsalis species complex (Diptera: Tephritidae): Taxonomic changes based on a review of 20 years of integrative morphological, molecular, cytogenetic, behavioural and chemoecological data. Syst. Èntomol. 2014, 40, 456–471. [Google Scholar] [CrossRef]
- Zhao, S.; Xing, Z.; Liu, Z.; Liu, Y.; Liu, X.; Chen, Z.; Li, J.; Yan, R. Efficient somatic and germline genome engineering of Bactrocera dorsalis by the CRISPR/Cas9 system. Pest Manag. Sci. 2018, 75, 1921–1932. [Google Scholar] [CrossRef]
- Jaffar, S.; Rizvi, S.A.H.; Lu, Y. Understanding the Invasion, Ecological Adaptations, and Management Strategies of Bactrocera dorsalis in China: A Review. Horticulturae 2023, 9, 1004. [Google Scholar] [CrossRef]
- Lux, S.A.; Copeland, R.S.; White, I.M.; Manrakhan, A.; Billah, M.K. A New Invasive Fruit Fly Species from the Bactrocera dorsalis (Hendel) Group Detected in East Africa. Int. J. Trop. Insect Sci. 2003, 23, 355–361. [Google Scholar] [CrossRef]
- Drew, R.A.I.; Tsuruta, K.; White, I.M. A new species of pest fruit fly (Diptera: Tephritidae: Dacinae) from Sri Lanka and Africa. Afr. Entomol. 2005, 13, 149–154. [Google Scholar]
- Badii, K.; Billah, M.; Afreh-Nuamah, K.; Obeng-Ofori, D.; Nyarko, G. Review of the pest status, economic impact and management of fruit-infesting flies (Diptera: Tephritidae) in Africa. Afr. J. Agric. Res. 2015, 10, 1488–1498. [Google Scholar] [CrossRef]
- CABI. Bactrocera dorsalis (Oriental Fruit Fly). Invasive Species Compendium. 2021. Available online: https://www.cabi.org/isc/datasheet/17685 (accessed on 25 May 2022).
- Zida, I.; Nacro, S.; Dabiré, R.; Somda, I. Co-Existence of Bactrocera dorsalis Hendel (Diptera: Tephritidae) and Ceratitis cosyra Walker (Diptera: Tephritidae) in the Mango Orchards in Western Burkina Faso. Adv. Èntomol. 2020, 8, 46–55. [Google Scholar] [CrossRef]
- Heve, W.K.; Adjadeh, T.A.; Billah, M.K. Overview and future research needs for development of effective biocontrol strategies for management of Bactrocera dorsalis Hendel (Diptera: Tephritidae) in sub-Saharan Africa. Pest Manag. Sci. 2021, 77, 4224–4237. [Google Scholar] [CrossRef] [PubMed]
- Ekesi, S.; Billah, M.K.; Nderitu, P.W.; Lux, S.A.; Rwomushana, I. Evidence for Competitive Displacement of Ceratitis cosyra by the Invasive Fruit Fly Bactrocera invadens (Diptera: Tephritidae) on Mango and Mechanisms Contributing to the Displacement. J. Econ. Èntomol. 2009, 102, 981–991. [Google Scholar] [CrossRef] [PubMed]
- Mutamiswa, R.; Nyamukondiwa, C.; Chikowore, G.; Chidawanyika, F. Overview of oriental fruit fly, Bactrocera dorsalis (Hendel) (Diptera: Tephritidae) in Africa: From invasion, bio-ecology to sustainable management. Crop Prot. 2020, 141, 105492. [Google Scholar] [CrossRef]
- Ouédraogo, S.N.; Vayssières, J.-F.; Dabiré, A.R.; Rouland-Lefèvre, C. Biodiversité des mouches des fruits (Diptera: Tephritidae) en vergers de manguiers de l’ouest du Burkina Faso: Structure et comparaison des communautés de différents sites. Fruits 2011, 66, 393–404. [Google Scholar] [CrossRef]
- Isasawin, S.; Aketarawong, N.; Thanaphum, S. Characterization and evaluation of microsatellite markers in a strain of the oriental fruit fly, Bactrocera dorsalis (Diptera: Tephritidae), with a genetic sexing character used in sterile insect population control. Eur. J. Èntomol. 2012, 109, 331–338. [Google Scholar] [CrossRef]
- Sutantawong, M.; Orankanok, W.; Enkerlin, W.R.; Wornoayporn, V.; Caceres, C. The sterile insect technique for control of the oriental fruit fly, Bactrocera dorsalis (Hendel), in mango orchards in Ratchaburi Province, Thailand. In Proceedings of the 6th International Fruit Fly Symposium, Stellenbosch, South Africa, 6–10 May 2002; pp. 223–232. Available online: https://nucleus.iaea.org/sites/naipc/twd/Documents/6thISFFEI_Proceedings/SUTANTAWONG.pdf (accessed on 23 January 2024).
- Leftwich, P.T.; Spurgin, L.G.; Harvey-Samuel, T.; Thomas, C.J.E.; Paladino, L.C.; Edgington, M.P.; Alphey, L. Genetic pest management and the background genetics of release strains. Philos. Trans. R. Soc. B Biol. Sci. 2020, 376, 20190805. [Google Scholar] [CrossRef]
- Legros, M.; Marshall, J.M.; Macfadyen, S.; Hayes, K.R.; Sheppard, A.; Barrett, L.G. Gene drive strategies of pest control in agricultural systems: Challenges and opportunities. Evol. Appl. 2021, 14, 2162–2178. [Google Scholar] [CrossRef] [PubMed]
- Medina, R.F. Gene drives and the management of agricultural pests. J. Responsible Innov. 2017, 5, S255–S262. [Google Scholar] [CrossRef]
- Choudhary, J.S.; Naaz, N.; Prabhakar, C.S.; Lemtur, M. Genetic analysis of oriental fruit fly, Bactrocera dorsalis (Diptera: Tephritidae) populations based on mitochondrial cox1 and nad1 gene sequences from India and other Asian countries. Genetica 2016, 144, 611–623. [Google Scholar] [CrossRef] [PubMed]
- National Academy of Sciences. Gene Drives on the Horizon; National Academies Press: Washington, DC, USA, 2016. [Google Scholar]
- Shi, W.; Kerdelhué, C.; Ye, H. Genetic Structure and Inferences on Potential Source Areas for Bactrocera dorsalis (Hendel) Based on Mitochondrial and Microsatellite Markers. PLoS ONE 2012, 7, e37083. [Google Scholar] [CrossRef] [PubMed]
- Zida, I.; Nacro, S.; Dabiré, R.; Ouédraogo, S.N.; Somda, I. Shea fruit-infesting fruit flies (Diptera: Tephritidae) and evaluation of infestation level according to the ethno-varieties in Western Burkina Faso. Int. J. Trop. Insect Sci. 2020, 40, 493–501. [Google Scholar] [CrossRef]
- McQuate, G.T.; Vargas, R.I. Assessment of Attractiveness of Plants as Roosting Sites for the Melon Fly, Bactrocera cucurbitae, and Oriental Fruit Fly, Bactrocera dorsalis. J. Insect Sci. 2007, 7, 57. [Google Scholar] [CrossRef] [PubMed]
- Aketarawong, N.; Bonizzoni, M.; Malacrida, A.R.; Gasperi, G.; Thanaphum, S. Seventeen novel microsatellite markers from an enriched library of the pest species Bactrocera dorsalis sensu stricto. Mol. Ecol. Notes 2006, 6, 1138–1140. [Google Scholar] [CrossRef]
- Khamis, F.M.; Karam, N.; Ekesi, S.; DE Meyer, M.; Bonomi, A.; Gomulski, L.M.; Scolari, F.; Gabrieli, P.; Siciliano, P.; Masiga, D.; et al. Uncovering the tracks of a recent and rapid invasion: The case of the fruit fly pest Bactrocera invadens (Diptera: Tephritidae) in Africa. Mol. Ecol. 2009, 18, 4798–4810. [Google Scholar] [CrossRef] [PubMed]
- Yi, C.; Zheng, C.; Zeng, L.; Xu, Y. High genetic diversity in the offshore island populations of the tephritid fruit fly Bactrocera dorsalis. BMC Ecol. 2016, 16, 46. [Google Scholar] [CrossRef]
- Peakall, R.; Smouse, P.E. GenAlEx 6.5: Genetic analysis in Excel. Population genetic software for teaching and research—An update. Bioinformatics 2012, 28, 2537–2539. [Google Scholar] [CrossRef]
- Konopiński, M.K. Shannon diversity index: A call to replace the original Shannon’s formula with unbiased estimator in the population genetics studies. PeerJ 2020, 8, e9391. [Google Scholar] [CrossRef] [PubMed]
- Nei, M. Chapter 9: Genetic Distance Between Populations. In The American Naturalist; Columbia University Press: New York, NY, USA, 1972; Volume 106, pp. 208–253. [Google Scholar]
- Jombart, T.; Devillard, S.; Balloux, F. Discriminant analysis of principal components: A new method for the analysis of genetically structured populations. BMC Genet. 2010, 11, 94. [Google Scholar] [CrossRef] [PubMed]
- Earl, D.A.; vonHoldt, B.M. STRUCTURE HARVESTER: A website and program for visualizing STRUCTURE output and implementing the Evanno method. Conserv. Genet. Resour. 2012, 4, 359–361. [Google Scholar] [CrossRef]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software structure: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef] [PubMed]
- Ramasamy, R.K.; Ramasamy, S.; Bindroo, B.B.; Naik, V.G. STRUCTURE PLOT: A program for drawing elegant STRUCTURE bar plots in user friendly interface. SpringerPlus 2014, 3, 431. [Google Scholar] [CrossRef] [PubMed]
- Zida, I.; Nacro, S.; Dabiré, R.; Somda, I. Seasonal Abundance and Diversity of Fruit Flies (Diptera: Tephritidae) in Three Types of Plant Formations in Western Burkina Faso, West Africa. Ann. Èntomol. Soc. Am. 2020, 113, 343–354. [Google Scholar] [CrossRef]
- Faye, M.; Ndiaye, A.; Diallo, I.; Sembene, P.M. Genetic identification of populations of Bactrocera dorsalis (Diptera Tephritidae) in the Niayes and Lower Casamance areas in Senegal. J. Appl. Biosci. 2021, 158, 16351–16362. [Google Scholar] [CrossRef]
- Diallo, I.; Ndiaye, A.; Faye, E.; Faye, M.; Sembene, M. Does landscape factors drive the genetic diversity and structure of populations of fruit flies, Senegal? Exploratory study of the case of Bactrocera dorsalis (Hendel, 1912) in the Niayes, Senegal. J. Entomol. Zool. Stud. 2021, 9, 47–52. [Google Scholar] [CrossRef]
- Gnanvossou, D.; Hanna, R.; Goergen, G.; Salifu, D.; Tanga, C.M.; Mohamed, S.A.; Ekesi, S. Diversity and seasonal abundance of tephritid fruit flies in three agro-ecosystems in Benin, West Africa. J. Appl. Èntomol. 2017, 141, 798–809. [Google Scholar] [CrossRef]
- Vayssières, J.-F.; Korie, S.; Ayegnon, D. Correlation of fruit fly (Diptera Tephritidae) infestation of major mango cultivars in Borgou (Benin) with abiotic and biotic factors and assessment of damage. Crop Prot. 2009, 28, 477–488. [Google Scholar] [CrossRef]
- De Meeûs, T. Initiation à La Génétique des Populations Naturelles: Applications aux Parasites et à Leurs Vecteurs; IRD Éditio; IRD: Marseille, France, 2012. [Google Scholar]
- Traore, N.; Galizi, R.; Kientega, M.; Ma, H. Developing genetic tools to control the Oriental Fruit Fly Bactrocera dorsalis [Diptera: Tephritidae]: Potential strategies and molecular tools. Authorea Prepr. 2023, 1–22. [Google Scholar] [CrossRef]
- Wright, S. Variability Within and Among Natural Populations. In Evolution and the Genetics of Populations; University of Chicago Press: Chicago, IL, USA, 1978; Volume 4. [Google Scholar]
- Stinner, R.E.; Barfield, C.S.; Stimac, J.L.; Dohse, L. Dispersal and Movement of Insect Pests. Annu. Rev. Èntomol. 1983, 28, 319–335. [Google Scholar] [CrossRef]
- Lanzaro, G.C.; Campos, M.; Crepeau, M.; Cornel, A.; Estrada, A.; Gripkey, H.; Haddad, Z.; Kormos, A.; Palomares, S. Selection of sites for field trials of genetically engineered mosquitoes with gene drive. Evol. Appl. 2021, 14, 2147–2161. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Li, B.; Chen, N.; Yang, D.; Li, L.; Liu, T. Evaluation of Reference Genes for Quantitative Reverse Transcription Polymerase Chain Reaction in Bactrocera dorsalis (Diptera: Tephritidae) Subjected to Various Phytosanitary Treatments. Insects 2021, 12, 945. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sampling Sites | Plant Formations | Density (FTD)/Plant Formation | Proportions (%) [95%Cis]/Plant Formation/Site | Density (FTD)/Site | Proportions (%)/Site |
|---|---|---|---|---|---|
| BGRB | MO | 38 | 24.36 [17.62, 31.00] | 156 | 2.97 [02.51, 03.43] |
| SAP | 50 | 32.05 [24.73, 39.37] | |||
| WLD | 68 | 43.59 [35.76, 51.42] | |||
| BK-SG | MO | 17 | 19.10 [10.95, 27.25] | 89 | 1.69 [01.35, 02.04] |
| SAP | 09 | 10.11 [03.86, 16.36] | |||
| WLD | 63 | 70.79 [61.33, 80.25] | |||
| CMOE | MO | 504 | 79.18 [76.03, 82.31] | 637 | 12.14 [11.25, 13.02] |
| SAP | 92 | 14.45 [11.71, 17.17] | |||
| WLD | 41 | 06.43 [04.53, 08.34] | |||
| HOUET | MO | 149 | 83.71 [78.27, 89.15] | 178 | 3.39 [02.99, 03.78] |
| SAP | 24 | 13.48 [08.48, 18.48] | |||
| WLD | 05 | 02.82 [00.39, 05.23] | |||
| KNDG | MO | 1124 | 37.64 [35.99, 39.28] | 2986 | 56.88 [55.55, 58.21] |
| SAP | 1313 | 43.97 [42.19, 45.75] | |||
| WLD | 549 | 18.39 [16.99, 19.76] | |||
| PONI | MO | 645 | 57.69 [54.87, 60.63] | 1118 | 21.29 [20.18, 22.40] |
| SAP | 92 | 8.23 [06.62, 09.85] | |||
| WLD | 381 | 34.08 [31.32, 36.86] | |||
| SSLI | MO | 45 | 51.72 [41.20, 62.24] | 87 | 1.66 [01.31, 01.90] |
| SAP | 31 | 35.63 [25.61, 45.64] | |||
| WLD | 11 | 12.64 [05.66, 19.61] |
| Locus | N | Na | Ne | I | Ho | He | Ht | Fis | Fit | Fst | Nm |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Bd15 | 44.286 | 1.286 | 1.007 | 0.018 | 0.003 | 0.006 | 0.006 | 0.491 | 0.499 | 0.015 | 16.819 |
| Bd19 | 44.286 | 4 | 3.131 | 1.239 | 0.622 | 0.673 | 0.7 | 0.075 | 0.111 | 0.039 | 6.139 |
| Bi1 | 44.143 | 3 | 1.527 | 0.611 | 0.356 | 0.34 | 0.345 | −0.044 | −0.03 | 0.014 | 17.553 |
| Bi5 | 44.429 | 3.571 | 1.454 | 0.594 | 0.276 | 0.304 | 0.31 | 0.092 | 0.109 | 0.019 | 12.915 |
| Bi8 | 44.714 | 2 | 1.147 | 0.223 | 0.035 | 0.118 | 0.125 | 0.704 | 0.719 | 0.052 | 4.561 |
| Bi10 | 44.143 | 3 | 1.834 | 0.773 | 0.292 | 0.447 | 0.462 | 0.347 | 0.368 | 0.032 | 7.577 |
| MS12A | 43.857 | 3 | 1.894 | 0.787 | 0.589 | 0.47 | 0.477 | −0.253 | −0.233 | 0.016 | 15.846 |
| Bd85b | 45 | 3.714 | 2.613 | 1.055 | 0.438 | 0.616 | 0.644 | 0.288 | 0.32 | 0.044 | 5.368 |
| MS4 | 44.286 | 4.429 | 3.013 | 1.235 | 0.791 | 0.664 | 0.68 | −0.191 | −0.164 | 0.023 | 10.732 |
| MS3 | 44 | 3.143 | 1.623 | 0.688 | 0.162 | 0.378 | 0.384 | 0.571 | 0.577 | 0.015 | 16.091 |
| Mean | 44.314 | 3.114 | 1.924 | 0.722 | 0.356 | 0.402 | 0.413 | 0.208 | 0.228 | 0.027 | 11.36 |
| SE | 0.111 | 0.130 | 0.089 | 0.047 | 0.031 | 0.026 | 0.048 | 0.103 | 0.101 | 0.004 | 1.623 |
| Pop | N | Na | Ne | I | Ho | He | uHe | F | PPL |
|---|---|---|---|---|---|---|---|---|---|
| HOUET | 44.6 | 3.3 | 1.895 | 0.718 | 0.347 | 0.390 | 0.394 | 0.128 | 80% |
| KNDG | 44.8 | 3.5 | 1.963 | 0.741 | 0.328 | 0.397 | 0.402 | 0.202 | 80% |
| CMOE | 43.2 | 3.2 | 2.032 | 0.771 | 0.348 | 0.431 | 0.436 | 0.269 | 100% |
| BGRB | 44.6 | 3.1 | 1.916 | 0.740 | 0.384 | 0.423 | 0.428 | 0.167 | 90% |
| PONI | 44.5 | 2.9 | 1.926 | 0.716 | 0.365 | 0.398 | 0.402 | 0.159 | 90% |
| SSLI | 44.6 | 2.8 | 1.819 | 0.657 | 0.354 | 0.370 | 0.374 | 0.164 | 90% |
| BK-SG | 43.9 | 3.0 | 1.919 | 0.712 | 0.369 | 0.403 | 0.407 | 0.062 | 90% |
| Mean | 44.3 | 3.1 | 1.924 | 0.722 | 0.356 | 0.402 | 0.406 | 0.166 | 88.6% |
| Pop | BGRB | BK-SG | CMOE | HOUET | KNDG | PONI | SSLI |
|---|---|---|---|---|---|---|---|
| BGRB | 16.416 | 20.583 | 11.114 | 12.250 | 22.477 | 12.908 | |
| BK-SG | 0.015 | 17.607 | 11.655 | 11.114 | 13.639 | 12.250 | |
| CMOE | 0.012 | 0.014 | 14.456 | 22.478 | 27.528 | 13.639 | |
| HOUET | 0.022 | 0.021 | 0.017 | 14.456 | 13.639 | 14.456 | |
| KNDG | 0.020 | 0.022 | 0.011 | 0.017 | 17.607 | 10.620 | |
| PONI | 0.011 | 0.018 | 0.009 | 0.018 | 0.014 | 24.75 | |
| SSLI | 0.019 | 0.020 | 0.018 | 0.017 | 0.023 | 0.010 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Traoré, N.; Kientega, M.; Maïga, H.; Nebié, K.; Zida, I.; Galizi, R.; Kiendrebeogo, E.; Sow, B.B.D.; Belem, A.M.G.; Dabiré, R.A.; et al. Genetic Diversity and Population Structure of the Invasive Oriental Fruit Fly, Bactrocera dorsalis (Diptera: Tephritidae) in Burkina Faso. Insects 2024, 15, 298. https://doi.org/10.3390/insects15050298
Traoré N, Kientega M, Maïga H, Nebié K, Zida I, Galizi R, Kiendrebeogo E, Sow BBD, Belem AMG, Dabiré RA, et al. Genetic Diversity and Population Structure of the Invasive Oriental Fruit Fly, Bactrocera dorsalis (Diptera: Tephritidae) in Burkina Faso. Insects. 2024; 15(5):298. https://doi.org/10.3390/insects15050298
Chicago/Turabian StyleTraoré, Nouhoun, Mahamadi Kientega, Hamidou Maïga, Karim Nebié, Issaka Zida, Roberto Galizi, Emmanuel Kiendrebeogo, Bazoumana B. D. Sow, Adrien M. G. Belem, Remy A. Dabiré, and et al. 2024. "Genetic Diversity and Population Structure of the Invasive Oriental Fruit Fly, Bactrocera dorsalis (Diptera: Tephritidae) in Burkina Faso" Insects 15, no. 5: 298. https://doi.org/10.3390/insects15050298