

The Synergistic Effect of Lemongrass Essential Oil and Flometoquin, Flonicamid, and Sulfoxaflor on Bemisia tabaci (Genn.) (Hemiptera: Aleyrodidae): Insights into Toxicity, Biochemical Impact, and Molecular Docking
 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insect Population
2.2. Insecticides and Chemicals
2.3. Essential Oil Extraction and GC–MS Identification
2.4. Bioassays
2.5. Acute Toxicity
2.6. Biochemical Assays
2.6.1. Preparation of B. tabaci Homogenate
2.6.2. Acetylcholine Esterase (AChE) Assay
2.6.3. Cytochrome P450 Assay
2.6.4. Esterase Assay
2.6.5. Glutathione S-transferase (GST) Assay
2.7. Molecular Docking Study
2.8. Data Analysis
3. Results
3.1. Toxicity of the Tested Compounds to Bemisia tabaci
3.2. Interaction of Lemongrass EO with the Tested Insecticides on B. tabaci Adults
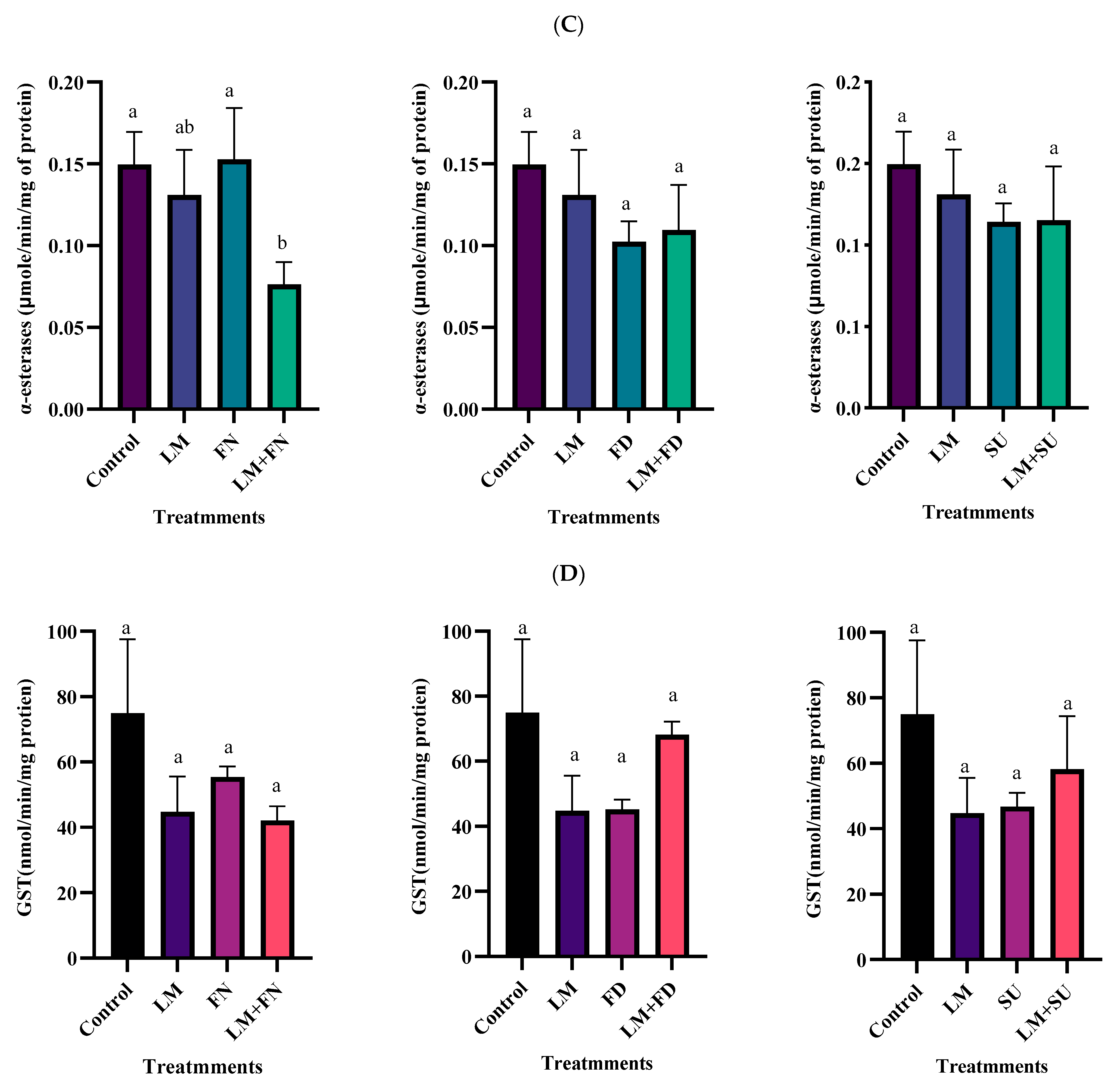
3.3. Biochemical Impact of the Tested Compounds
3.4. Biochemical Effect of Binary Combinations of Lemongrass EO with the Tested Insecticides
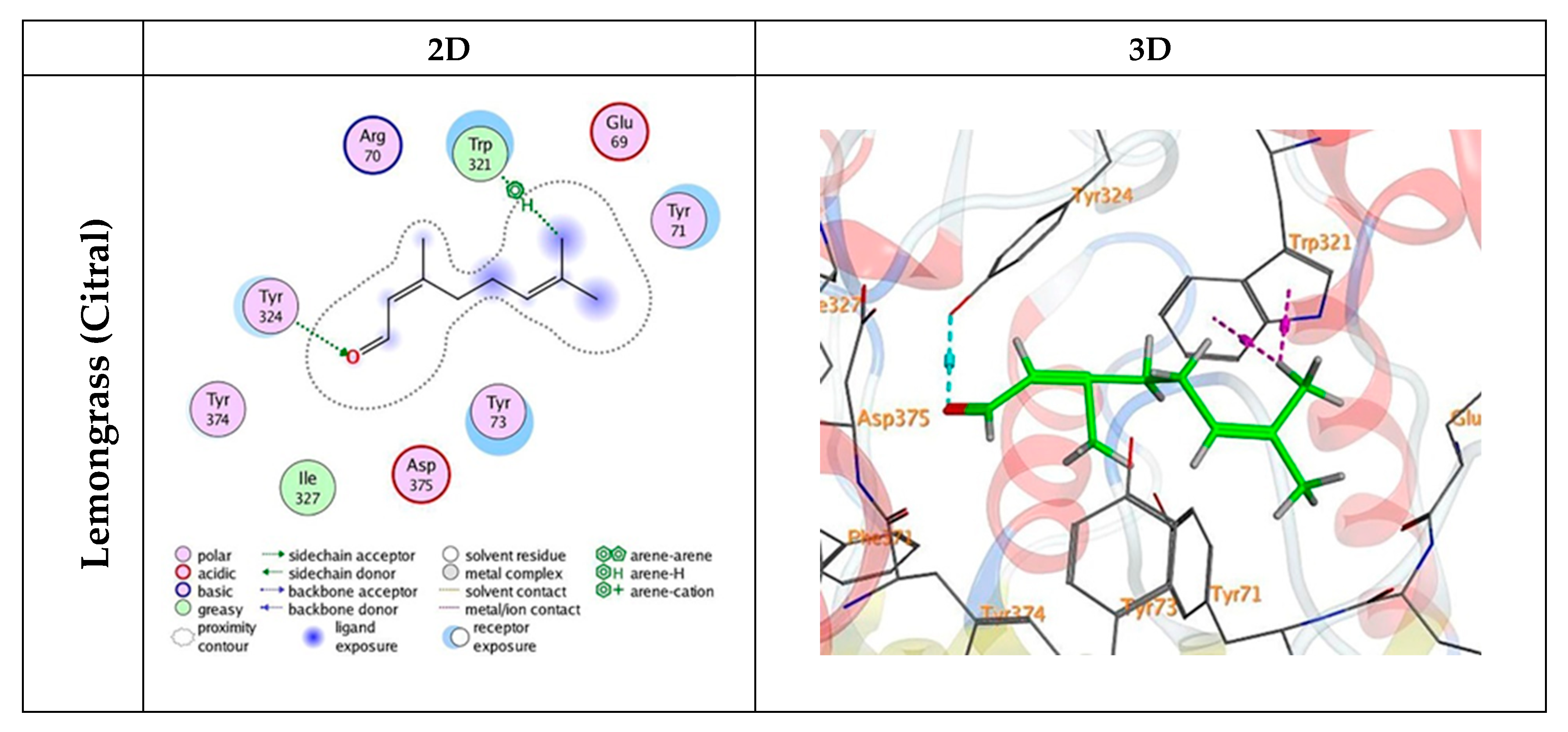
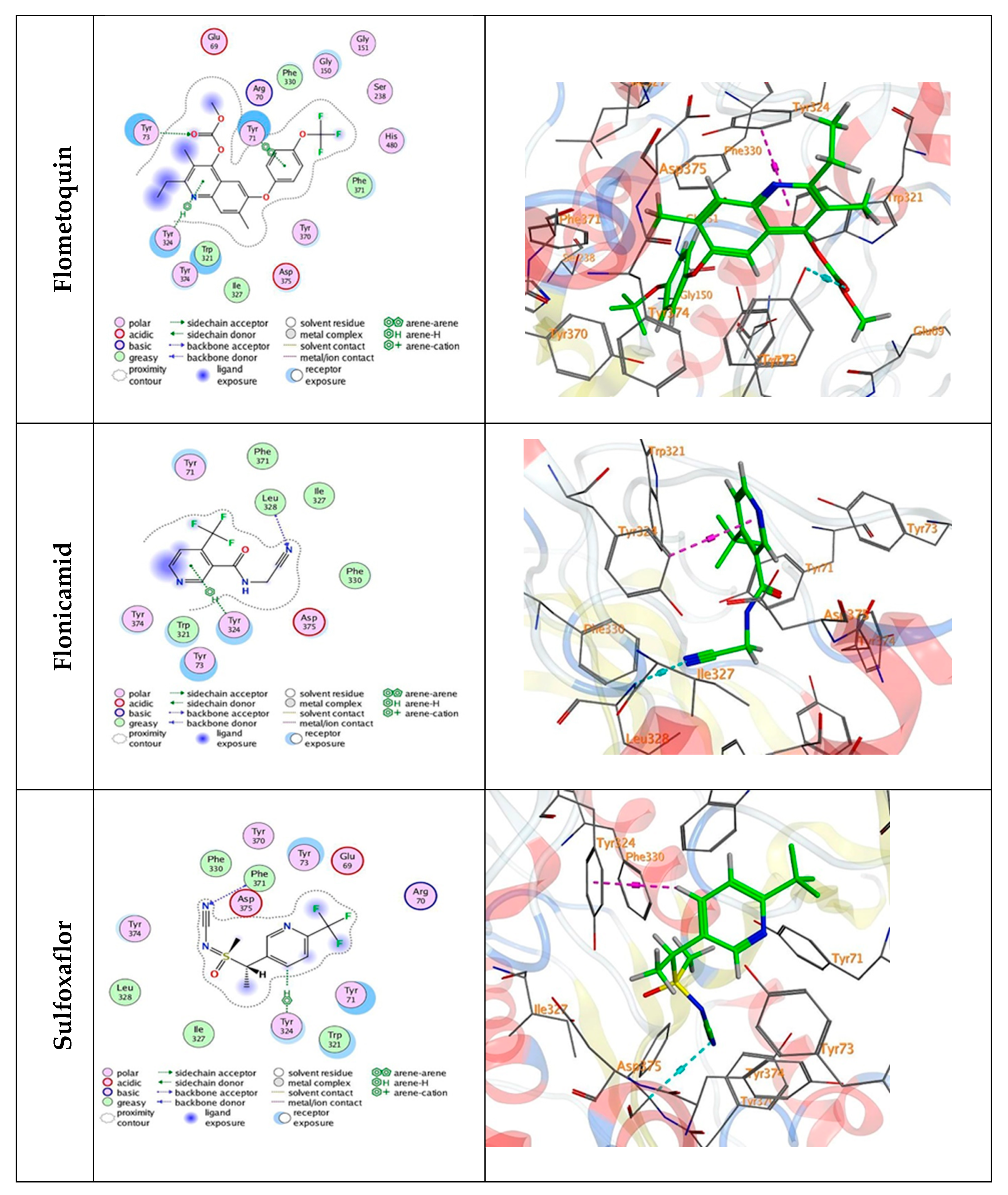
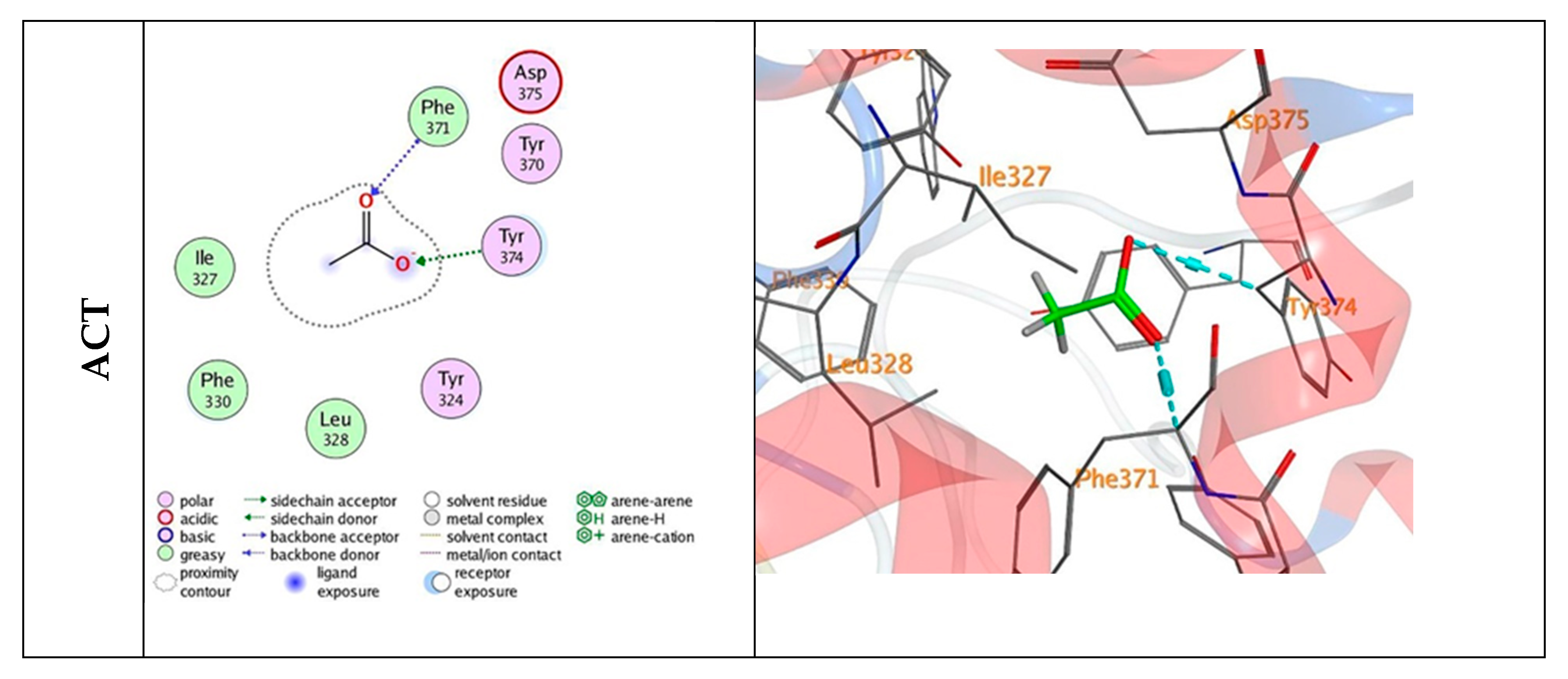
3.5. Docking Mechanisms of the Tested Compounds with AchE Enzyme
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Romba, R.; Gnankine, O.; Drabo, S.F.; Tiendrebeogo, F.; Henri, H.; Mouton, L.; Vavre, F. Abundance of Bemisia tabaci Gennadius (Hemiptera: Aleyrodidae) and its parasitoids on vegetables and cassava plants in Burkina Faso (West Africa). Ecol. Evol. 2018, 8, 6091–6103. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, S.; Ghanim, M. Factors Determining Transmission of Persistent Viruses by Bemisia Tabaci and Emergence of New Virus-Vector Relationships. Viruses 2021, 13, 1808. [Google Scholar] [CrossRef] [PubMed]
- Li, N.N.; Jiang, S.; Lu, K.Y.; Hong, J.S.; Wang, Y.B.; Yan, J.Y.; Luan, J.B. Bacteriocyte Development Is Sexually Differentiated in Bemisia Tabaci. Cell Rep. 2022, 38, 110613. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Jiang, Z.; Zhou, J.; Liu, X.; Zhang, Y.; Chu, D. Ecological Factors Associated with the Distribution of Bemisia tabaci Cryptic Species and Their Facultative Endosymbionts. Insects 2023, 14, 252. [Google Scholar] [CrossRef] [PubMed]
- EPPO Datasheets on Pests Recommended for Regulation: Bemisia Tabaci; European and Mediterranean Plant Protection Organization (EPPO): Paris, France, 2024; 12p.
- Moustafa, M.; Hefny, D.; Alfuhaid, N.; Helmy, R.; El-Said, N.; Ibrahim, E.-D. Effectiveness and Biochemical Impact of Flubendiamide and Flonicamid Insecticides against Bemisia Tabaci (Hemiptera: Aleyrodidae) and Residue Dissipation in Cherry Tomato Plants and Soil under Greenhouse Conditions 1. J. Entomol. Sci. 2024, 59, 1–22. [Google Scholar] [CrossRef]
- El Kady, H.; Devine, G.J. Insecticide Resistance in Egyptian Populations of the Cotton Whitefly, Bemisia Tabaci (Hemiptera: Aleyrodidae). Pest Manag. Sci. 2003, 59, 865–871. [Google Scholar] [CrossRef] [PubMed]
- Dawood, A.I. Monitoring Resistance in the Whitefly Bemisia Tabaci (Homoptera: Aleyrodidae) to the Efficiency of Three Insecticides in Relation to Some Detoxification Enzymes. Egypt. Acad. J. Biol. Sci. F Toxicol. Pest Control 2016, 8, 21–28. [Google Scholar] [CrossRef]
- Shaurub, E.-S.; Farghaley, S.; Dawood, A.; Mohamed, A. Current Insecticide-Resistance Status and Activity of Detoxifying Enzymes in Field Populations of Bemisia Tabaci from Egypt. Egypt. Sci. J. Pestic. 2016, 2, 1–14. [Google Scholar]
- Shaurub, E.-S.H.; Paredes-Montero, J.R.; Brown, J.K.; Zein, H.S.; Mohamed, A.A. Metabolic Resistance to Organophosphate Insecticides in Natural Populations of the Whitefly Bemisia Tabaci (Hemiptera: Aleyrodidae) in Egypt and Molecular Identification of Mitotypes. Phytoparasitica 2021, 49, 443–457. [Google Scholar] [CrossRef]
- Hassan, E.; Obaidoon, Y.; Mostafiz, M.M.; Senior, L. Effect of Plant Essential Oil Formulations on Bemisia Tabaci MEAM1 (Gennadius) and Its Parasitoid Eretmocerus Hayati (Zolnerowich and Rose). Plants 2023, 12, 4137. [Google Scholar] [CrossRef] [PubMed]
- Riyaz, S.U.M.; Kathiravan, K. Integrated Pest Management Approaches. In Geminiviruses: Impact, Challenges and Approaches; Kumar, R.V., Ed.; Springer International Publishing: Cham, Switzerland, 2019; pp. 219–235. ISBN 978-3-030-18248-9. [Google Scholar]
- Moustafa, A.M.M.; Elmenofy, H.W.; Osman, A.E.; El-Said, A.N.; Awad, M. Biological Impact, Oxidative Stress and Adipokinetic Hormone Activities of Agrotis Ipsilon in Response to Bioinsecticides. Plant Prot. Sci. 2022, 58, 326–337. [Google Scholar] [CrossRef]
- El-Shourbagy, N.M.; Farag, S.M.; Moustafa, M.A.M.; Al-Shuraym, L.; Sayed, S.; Zyaan, O. Biochemical and Insecticidal Efficacy of Clove and Basil Essential Oils and Two Photosensitizers and Their Combinations on Aphis Gossypii Glover (Hemiptera: Aphididae). Biosci. J. 2023, 39, e39100. [Google Scholar] [CrossRef]
- Onstad, D.W.; Knolhoff, L.M. Chapter One—Major Issues in Insect Resistance Management. In Insect Resistance Management, 3rd ed.; Onstad, D.W., Knolhoff, L.M., Eds.; Academic Press: Cambridge, MA, USA, 2023; pp. 1–29. ISBN 978-0-12-823787-8. [Google Scholar]
- Mansour, R.; Belzunces, L.P.; Suma, P.; Zappalà, L.; Mazzeo, G.; Grissa-Lebdi, K.; Russo, A.; Biondi, A. Vine and Citrus Mealybug Pest Control Based on Synthetic Chemicals. A Review. Agron. Sustain. Dev. 2018, 38, 37. [Google Scholar] [CrossRef]
- Kannan, M.; Bojan, N.; Swaminathan, J.; Zicarelli, G.; Hemalatha, D.; Zhang, Y.; Ramesh, M.; Faggio, C. Nanopesticides in Agricultural Pest Management and Their Environmental Risks: A Review. Int. J. Environ. Sci. Technol. 2023, 20, 10507–10532. [Google Scholar] [CrossRef]
- Suwannayod, S.; Sukontason, K.L.; Pitasawat, B.; Junkum, A.; Limsopatham, K.; Jones, M.K.; Somboon, P.; Leksomboon, R.; Chareonviriyaphap, T.; Tawatsin, A.; et al. Synergistic Toxicity of Plant Essential Oils Combined with Pyrethroid Insecticides against Blow Flies and the House Fly. Insects 2019, 10, 178. [Google Scholar] [CrossRef]
- Arokiyaraj, C.; Bhattacharyya, K.; Reddy, S.G.E. Toxicity and Synergistic Activity of Compounds from Essential Oils and Their Effect on Detoxification Enzymes against Planococcus Lilacinus. Front. Plant Sci. 2022, 13, 1016737. [Google Scholar] [CrossRef] [PubMed]
- Luijten, M.; Vlaanderen, J.; Kortenkamp, A.; Antignac, J.-P.; Barouki, R.; Bil, W.; van den Brand, A.; den Braver-Sewradj, S.; van Klaveren, J.; Mengelers, M.; et al. Mixture Risk Assessment and Human Biomonitoring: Lessons Learnt from HBM4EU. Int. J. Hyg. Environ. Health 2023, 249, 114135. [Google Scholar] [CrossRef] [PubMed]
- Rizzati, V.; Briand, O.; Guillou, H.; Gamet-Payrastre, L. Effects of Pesticide Mixtures in Human and Animal Models: An Update of the Recent Literature. Chem. Biol. Interact. 2016, 254, 231–246. [Google Scholar] [CrossRef] [PubMed]
- Alves, M.d.S.; Medeiros, E.A.D.P.d.; Pereira, C.d.S.B.; Moreira, Y.N.; Cappato, J.d.S.; Osorio, R.d.P.; Riger, C.J.; Santos, L.V.d.; Mesquita, R.D.; Pontes, E.G.; et al. Lemongrass Essential Oil: Scientific Bases for an Agroecological Approach to Seed Protection. Ind. Crops Prod. 2023, 199, 116760. [Google Scholar] [CrossRef]
- Pavela, R.; Benelli, G. Essential Oils as Ecofriendly Biopesticides? Challenges and Constraints. Trends Plant Sci. 2016, 21, 1000–1007. [Google Scholar] [CrossRef] [PubMed]
- Jeschke, P. Recent Developments in Fluorine-Containing Pesticides. Pest Manag. Sci. 2023. [Google Scholar] [CrossRef] [PubMed]
- Madesh, K.; Komala, G.; Chandralekha, R.; Tripathi, P. Mode of Action of Novel Insecticides. In Advanced Trends in Plant Protection; P.K. Publishers & Distributors: Delhi, India, 2024; pp. 255–291. ISBN 978-81-19428-xx-x. [Google Scholar]
- Kobayashi, T.; Hotta, H.; Miyake, T.; Nomura, M.; Horikoshi, R.; Yamamoto, K. Discovery of Flometoquin, a Novel Quinoline Insecticide. J. Pestic. Sci. 2023, 48, 168–174. [Google Scholar] [CrossRef] [PubMed]
- Ren, M.; Niu, J.; Hu, B.; Wei, Q.; Zheng, C.; Tian, X.; Gao, C.; He, B.; Dong, K.; Su, J. Block of Kir Channels by Flonicamid Disrupts Salivary and Renal Excretion of Insect Pests. Insect Biochem. Mol. Biol. 2018, 99, 17–26. [Google Scholar] [CrossRef] [PubMed]
- El-Hefny, D.E.; Ibrahim, E.-D.S.; Alfuhaid, N.A.; Fónagy, A.; Moustafa, M.A.M. Residual Effect of the Insecticides Flonicamid and Spiromesifen against Aphis Craccivora (Hemiptera: Aphididae) and Persistence Dynamics in Faba Bean1. J. Entomol. Sci. 2024, 59, 165–181. [Google Scholar] [CrossRef]
- Morita, M.; Yoneda, T.; Akiyoshi, N. Research and Development of a Novel Insecticide, Flonicamid. J. Pestic. Sci. 2014, 39, 179–180. [Google Scholar] [CrossRef]
- Shi, D.; Luo, C.; Lv, H.; Zhang, L.; Desneux, N.; You, H.; Li, J.; Ullah, F.; Ma, K. Impact of Sublethal and Low Lethal Concentrations of Flonicamid on Key Biological Traits and Population Growth Associated Genes in Melon Aphid, Aphis Gossypii Glover. Crop Prot. 2022, 152, 105863. [Google Scholar] [CrossRef]
- Arthur, A.L.; Kirkland, L.; Chirgwin, E.; van Rooyen, A.; Umina, P.A. Baseline Susceptibility of Australian Myzus Persicae (Hemiptera: Aphididae) to Novel Insecticides Flonicamid and Afidopyropen. Crop Prot. 2022, 158, 105992. [Google Scholar] [CrossRef]
- Moustafa, M.; Amer, A.; Al-Shuraym, L.; Ibrahim, E.-D.; Elhefny, D.; Salem, M.; Sayed, S. Efficacy of Chemical and Bio-Pesticides on Cowpea Aphid, Aphis Craccivora, and Their Residues on the Productivity of Fennel Plants (Foeniculum Vulgare). J. King Saud. Univ.-Sci. 2022, 34, 101900. [Google Scholar] [CrossRef]
- Moustafa, M.; Awad, M.; Amer, A.; Hassan, N.; Ibrahim, E.-D.; Ali, H.; Akrami, M.; Salem, M. Insecticidal Activity of Lemongrass Essential Oil as an Eco-Friendly Agent against the Black Cutworm Agrotis Ipsilon (Lepidoptera: Noctuidae). Insects 2021, 12, 737. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Ellman, G.L.; Courtney, K.D.; Andres, V.J.; Feather-Stone, R.M. A New and Rapid Colorimetric Determination of Acetylcholinesterase Activity. Biochem. Pharmacol. 1961, 7, 88–95. [Google Scholar] [CrossRef] [PubMed]
- Hansen, L.G.; Hodgson, E. Biochemical Characteristics of Insect Microsomes. N- and O-Demethylation. Biochem. Pharmacol. 1971, 20, 1569–1578. [Google Scholar] [CrossRef] [PubMed]
- Asperen, K. van A Study of Housefly Esterases by Means of a Sensitive Colorimetric Method. J. Insect Physiol. 1962, 8, 401–416. [Google Scholar] [CrossRef]
- Habig, W.H.; Pabst, M.J.; Jakoby, W.B. Glutathione S-Transferases. The First Enzymatic Step in Mercapturic Acid Formation. J. Biol. Chem. 1974, 249, 7130–7139. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.; Liu, Y.-X.; Zhang, S.-P.; Wang, Y.-Q.; Gao, P.; Li, Y.-T.; Yang, X.-Q. Transcription Factor AhR Regulates Glutathione S-Transferases Conferring Resistance to Lambda-Cyhalothrin in Cydia Pomonella. J. Agric. Food Chem. 2023, 71, 5230–5239. [Google Scholar] [CrossRef] [PubMed]
- Finney, D.J. Probit Analysis: A Statistical Treatment of the Sigmoid Response Curve; Cambridge University Press: Cambridge, UK, 1964; p. 25. [Google Scholar]
- Anaduaka, E.G.; Uchendu, N.O.; Asomadu, R.O.; Ezugwu, A.L.; Okeke, E.S.; Chidike Ezeorba, T.P. Widespread Use of Toxic Agrochemicals and Pesticides for Agricultural Products Storage in Africa and Developing Countries: Possible Panacea for Ecotoxicology and Health Implications. Heliyon 2023, 9, e15173. [Google Scholar] [CrossRef] [PubMed]
- Sonhafouo-Chiana, N.; Nkahe, L.D.; Kopya, E.; Awono-Ambene, P.H.; Wanji, S.; Wondji, C.S.; Antonio-Nkondjio, C. Rapid Evolution of Insecticide Resistance and Patterns of Pesticides Usage in Agriculture in the City of Yaoundé, Cameroon. Parasit. Vectors 2022, 15, 186. [Google Scholar] [CrossRef] [PubMed]
- Moustafa, M.A.M.; Moteleb, R.I.A.; Ghoneim, Y.F.; Hafez, S.S.; Ali, R.E.; Eweis, E.E.A.; Hassan, N.N. Monitoring Resistance and Biochemical Studies of Three Egyptian Field Strains of Spodoptera Littoralis (Lepidoptera: Noctuidae) to Six Insecticides. Toxics 2023, 11, 211. [Google Scholar] [CrossRef] [PubMed]
- Martín-López, B.; Varela, I.; Marnotes, S.; Cabaleiro, C. Use of Oils Combined with Low Doses of Insecticide for the Control of Myzus Persicae and PVY Epidemics. Pest Manag. Sci. 2006, 62, 372–378. [Google Scholar] [CrossRef] [PubMed]
- Faraone, N.; Hillier, N.K.; Cutler, G.C. Plant Essential Oils Synergize and Antagonize Toxicity of Different Conventional Insecticides against Myzus Persicae (Hemiptera: Aphididae). PLoS ONE 2015, 10, e0127774. [Google Scholar] [CrossRef] [PubMed]
- Silva, S.; Cunha, J.; Carvalho, S.; Zandonadi, C.; Martins, R.; Chang, R. Ocimum Basilicum Essential Oil Combined with Deltamethrin to Improve the Management of Spodoptera Frugiperda. Ciênc. E Agrotecnol. 2017, 41, 665–675. [Google Scholar] [CrossRef]
- Ismail, S. Synergistic Efficacy of Plant Essential Oils with Cypermethrin and Chlorpyrifos Against Spodoptera Littoralis, Field Populations in Egypt. Int. J. Adv. Biol. Biomed. Res. 2020, 9, 128–137. [Google Scholar] [CrossRef]
- Mantzoukas, S.; Kalyvas, N.; Ntoukas, A.; Lagogiannis, I.; Farsalinos, K.; Eliopoulos, P.A.; Poulas, K. Combined Toxicity of Cannabidiol Oil with Three Bio-Pesticides against Adults of Sitophilus Zeamais, Rhyzopertha Dominica, Prostephanus Truncatus and Trogoderma Granarium. Int. J. Environ. Res. Public. Health 2020, 17, 6664. [Google Scholar] [CrossRef] [PubMed]
- Silva, S.M.; Cunha, J.P.A.R.d.; Zandonadi, C.H.S.; Assunção, H.H.T.d.; Marques, M.G. Synergistic Effects of Binary Mixtures of Linalool with Pyrethroids against Fall Armyworm. Biosci. J. 2020, 36, 228–237. [Google Scholar] [CrossRef]
- Basit, M.; Saeed, S.; Saleem, M.A.; Denholm, I.; Shah, M. Detection of Resistance, Cross-Resistance, and Stability of Resistance to New Chemistry Insecticides in Bemisia Tabaci (Homoptera: Aleyrodidae). J. Econ. Entomol. 2013, 106, 1414–1422. [Google Scholar] [CrossRef] [PubMed]
- da Silva Santana, A.; Baldin, E.L.L.; Lima, A.P.S.; dos Santos, T.L.B.; Santos, M.C.; Vieira, T.M.; Crotti, A.E.M.; Takeara, R. New Challenges Demand New Solutions: Selected Essential Oils as an Alternative to Control Bemisia Tabaci MED in Brazil. Crop Prot. 2022, 155, 105909. [Google Scholar] [CrossRef]
- Costa, E.C.C.; Christofoli, M.; Costa, G.C.d.S.; Peixoto, M.F.; Fernandes, J.B.; Forim, M.R.; Pereira, K.d.C.; Silva, F.G.; Cazal, C.d.M. Essential Oil Repellent Action of Plants of the Genus Zanthoxylum against Bemisia Tabaci Biotype B (Homoptera: Aleyrodidae). Sci. Hortic. 2017, 226, 327–332. [Google Scholar] [CrossRef]
- El-Zahi, E.-Z.; El-Sarand, E.-S.; El Masry, G. Activity of Flonicamid and Two Neonicotinoid Insecticides against Bemisia Tabaci (Gennadius) and Its Associated Predators on Cotton Plants. Egypt. Acad. J. Biol. Sci. Entomol. 2017, 10, 25–34. [Google Scholar] [CrossRef]
- Watson, G.B.; Siebert, M.W.; Wang, N.X.; Loso, M.R.; Sparks, T.C. Sulfoxaflor—A Sulfoximine Insecticide: Review and Analysis of Mode of Action, Resistance and Cross-Resistance. Pestic. Biochem. Physiol. 2021, 178, 104924. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, F.S.; Helmy, W.S.; El-Din, H.S.; Al Naggar, Y. Chronic In-Hive Exposure to a Field-Relevant Concentration of CloserTM SC (24% Sulfoxaflor) Insecticide Altered Immunological and Physiological Markers of Honey Bee Foragers (Apis Mellifera). Apidologie 2023, 54, 4. [Google Scholar] [CrossRef]
- Ibrahim, E.-D.S.; Abd Alla, A.E.; El-Masarawy, M.S.; Salem, R.A.; Hassan, N.N.; Moustafa, M.A.M. Sulfoxaflor Influences the Biochemical and Histological Changes on Honeybees (Apis mellifera L.). Ecotoxicology 2023, 32, 674–681. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.J. Detoxification Mechanisms in Insects. In Encyclopedia of Entomology; Capinera, J.L., Ed.; Springer: Dordrecht, The Netherlands, 2008; pp. 1187–1201. ISBN 978-1-4020-6359-6. [Google Scholar]
- Damasceno, J.M.; Rato, L.D.; Simões, T.; Morão, I.F.C.; Meireles, G.; Novais, S.C.; Lemos, M.F.L. Exposure to the Insecticide Sulfoxaflor Affects Behaviour and Biomarkers Responses of Carcinus Maenas (Crustacea: Decapoda). Biology 2021, 10, 1234. [Google Scholar] [CrossRef] [PubMed]
- Schmidel, A.J.; Assmann, K.L.; Werlang, C.C.; Bertoncello, K.T.; Francescon, F.; Rambo, C.L.; Beltrame, G.M.; Calegari, D.; Batista, C.B.; Blaser, R.E.; et al. Subchronic Atrazine Exposure Changes Defensive Behaviour Profile and Disrupts Brain Acetylcholinesterase Activity of Zebrafish. Neurotoxicol. Teratol. 2014, 44, 62–69. [Google Scholar] [CrossRef]
- Morakchi, S.; Maïza, A.; Farine, P.; Aribi, N.; Soltani, N. Effects of a Neonicotinoid Insecticide (Acetamiprid) on Acetylcholinesterase Activity and Cuticular Hydrocarbons Profil in German Cockroaches. Commun. Agric. Appl. Biol. Sci. 2005, 70, 843–848. [Google Scholar] [PubMed]
- Topal, A.; Alak, G.; Ozkaraca, M.; Yeltekin, A.C.; Comaklı, S.; Acıl, G.; Kokturk, M.; Atamanalp, M. Neurotoxic Responses in Brain Tissues of Rainbow Trout Exposed to Imidacloprid Pesticide: Assessment of 8-Hydroxy-2-Deoxyguanosine Activity, Oxidative Stress and Acetylcholinesterase Activity. Chemosphere 2017, 175, 186–191. [Google Scholar] [CrossRef] [PubMed]
- Özdemir, H.H.; Kara, M.; Yumrutas, O.; Uckardes, F.; Eraslan, E.; Demir, C.F.; Bal, R. Determination of the Effects on Learning and Memory Performance and Related Gene Expressions of Clothianidin in Rat Models. Cogn. Neurodyn. 2014, 8, 411–416. [Google Scholar] [CrossRef] [PubMed]
- Piner Benli, P.; Çelik, M. Glutathione and Its Dependent Enzymes’ Modulatory Responses to Neonicotinoid Insecticide Sulfoxaflor Induced Oxidative Damage in Zebrafish in Vivo. Sci. Prog. 2021, 104, 00368504211028361. [Google Scholar] [CrossRef] [PubMed]
- Boily, M.; Sarrasin, B.; Deblois, C.; Aras, P.; Chagnon, M. Acetylcholinesterase in Honey Bees (Apis Mellifera) Exposed to Neonicotinoids, Atrazine and Glyphosate: Laboratory and Field Experiments. Environ. Sci. Pollut. Res. Int. 2013, 20, 5603–5614. [Google Scholar] [CrossRef] [PubMed]
- Sparks, T.C.; Nauen, R. IRAC: Mode of Action Classification and Insecticide Resistance Management. Pestic. Biochem. Physiol. 2015, 121, 122–128. [Google Scholar] [CrossRef] [PubMed]
- Nesterov, A.; Spalthoff, C.; Kandasamy, R.; Katana, R.; Rankl, N.B.; Andrés, M.; Jähde, P.; Dorsch, J.A.; Stam, L.F.; Braun, F.-J.; et al. TRP Channels in Insect Stretch Receptors as Insecticide Targets. Neuron 2015, 86, 665–671. [Google Scholar] [CrossRef] [PubMed]
- Kandasamy, R.; Costea, P.I.; Stam, L.; Nesterov, A. TRPV Channel Nanchung and TRPA Channel Water Witch Form Insecticide-Activated Complexes. Insect Biochem. Mol. Biol. 2022, 149, 103835. [Google Scholar] [CrossRef] [PubMed]
- Moustafa, M.A.M.; Hassan, N.N.; Alfuhaid, N.A.; Amer, A.; Awad, M. Insights into the Toxicity, Biochemical Activity, and Molecular Docking of Cymbopogon Citratus Essential Oils and Citral on Spodoptera Littoralis (Lepidoptera: Noctuidae). J. Econ. Entomol. 2023, 116, 1185–1195. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | LC25 (mg/L) (95% Confidence Limits) | LC50 (mg/L) (95% Confidence Limits) | Slope ± SE | χ2 |
|---|---|---|---|---|
| Lemongrass (C. citratus) | 68.85 (56.40–81.08) | 147.71 (128.19–171.02) | 2.03 ± 0.16 | 0.73 |
| Flometoquin | 11.79 (9.81–13.60) | 22.57 (19.88–25.55) | 2.37 ± 0.17 | 1.07 |
| Flonicamid | 3.98 (3.38–4.58) | 7.52 (6.64–8.54) | 2.44 ± 0.18 | 1.76 |
| Sulfoxaflor | 1.89 (1.60–2.19) | 3.69 (3.25–4.22) | 2.33 ± 0.18 | 1.94 |
| Compounds | Conc. | No. | % M a | ME | χ2 b | D b |
|---|---|---|---|---|---|---|
| LM | LC25 | 62 | 25.2 ± 5.5 b | |||
| FN | LC25 | 85 | 29.8 ± 9.2 b | |||
| FD | LC25 | 69 | 28.9 ± 10.7 b | |||
| SU | LC25 | 64 | 35.4 ± 5.0 b | |||
| LM + FN | LC25 + LC25 | 61 | 62.6 ± 3.5 a | 47.50 | 4.97 | 15.09 |
| LM + FD | LC25 + LC25 | 71 | 66.7 ± 3.8 a | 46.86 | 8.64 | 19.88 |
| LM + SU | LC25 + LC25 | 74 | 71.5 ± 4.3 a | 48.52 | 11.1 | 22.94 |
| Compound | Energy Score (S) (Kcal/mol) | Affinity Bond Strength (Kcal/mol) | Affinity Bond Length (in A° from Main Residue) | Amino Acids | Ligand | Interaction |
|---|---|---|---|---|---|---|
| Citral | −4.69 | −0.8 | 2.82 | TYR 324 | O 19 | H-acceptor |
| −0.6 | 3.79 | TRP 321 | C 20 | H-pi | ||
| −0.6 | 4.44 | TRP 321 | C 20 | H-pi | ||
| Flometoquin | −7.06 | −0.8 | 2.85 | TYR 73 | O 3 | H-acceptor |
| −0.6 | 4.00 | TYR 324 | 6-ring | pi-H | ||
| −0.0 | 3.97 | TYR 71 | 6-ring | pi-pi | ||
| Flonicamid | −5.26 | −0.8 | 3.44 | LEU 328 | N 22 | H-acceptor |
| −0.7 | 3.64 | TYR 324 | 6-ring | pi-H | ||
| Sulfoxaflor | −5.60 | −0.7 | 3.70 | PHE 371 | N 3 | H-acceptor |
| −0.6 | 4.53 | TYR 324 | C 14 | H-pi | ||
| ACT (acetate ion) | −3.41 | −1.8 | 3.00 | PHE 371 | O 6 | H-acceptor |
| −0.7 | 2.97 | TYR 374 | O 7 | H-acceptor |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moustafa, M.A.M.; Ahmed, F.S.; Alfuhaid, N.A.; El-Said, N.A.; Ibrahim, E.-D.S.; Awad, M. The Synergistic Effect of Lemongrass Essential Oil and Flometoquin, Flonicamid, and Sulfoxaflor on Bemisia tabaci (Genn.) (Hemiptera: Aleyrodidae): Insights into Toxicity, Biochemical Impact, and Molecular Docking. Insects 2024, 15, 302. https://doi.org/10.3390/insects15050302
Moustafa MAM, Ahmed FS, Alfuhaid NA, El-Said NA, Ibrahim E-DS, Awad M. The Synergistic Effect of Lemongrass Essential Oil and Flometoquin, Flonicamid, and Sulfoxaflor on Bemisia tabaci (Genn.) (Hemiptera: Aleyrodidae): Insights into Toxicity, Biochemical Impact, and Molecular Docking. Insects. 2024; 15(5):302. https://doi.org/10.3390/insects15050302
Chicago/Turabian StyleMoustafa, Moataz A. M., Fatma S. Ahmed, Nawal Abdulaziz Alfuhaid, Nourhan A. El-Said, El-Desoky S. Ibrahim, and Mona Awad. 2024. "The Synergistic Effect of Lemongrass Essential Oil and Flometoquin, Flonicamid, and Sulfoxaflor on Bemisia tabaci (Genn.) (Hemiptera: Aleyrodidae): Insights into Toxicity, Biochemical Impact, and Molecular Docking" Insects 15, no. 5: 302. https://doi.org/10.3390/insects15050302