
Gut Microbiota Affects Host Fitness of Fall Armyworm Feeding on Different Food Types


State Key Laboratory for Biology of Plant Diseases and Insect Pests, Institute of Plant Protection, Chinese Academy of Agricultural Science, Beijing 100193, China
*
Author to whom correspondence should be addressed.
Insects 2024, 15(5), 304; https://doi.org/10.3390/insects15050304
Submission received: 27 March 2024
/
Revised: 19 April 2024
/
Accepted: 23 April 2024
/
Published: 24 April 2024
(This article belongs to the Section Insect Pest and Vector Management)
Abstract
:Simple Summary
The fall armyworm, a migratory invasive pest, poses a serious threat to the food security of major crops like maize, wheat, and rice. Understanding its adaptation mechanism to different hosts is crucial for developing effective control technology. This study examined the host fitness and gut microbial diversity of fall armyworms fed four different types of food. Based on life history parameters, pupa weight, and nutrient utilization indexes, the host fitness ranking from high to low was artificial diet, maize, wheat, and rice. Gut microbial composition and diversity varied significantly among fall armyworms fed different foods due to changes in low-abundant bacteria. Fall armyworms fed maize had the highest gut microbial diversity. The functions of gut microbes with significant abundance differences were enriched in nutrient and vitamin metabolism as well as other pathways closely related to host adaptation. Additionally, we identified five genera (Acinetobacter, Variovorax, Pseudomonas, Bacillus, and Serratia) that positively correlated with host fitness, while one genus (Rahnella) negatively correlated with it. This study reveals the potential role of gut microbes in the host adaptation of fall armyworms.
Abstract
The fall armyworm (FAW), Spodoptera frugiperda, seriously threatens food and cash crops. Maize, wheat, and even rice damage by FAWs have been reported in many areas of China. It is urgent to clarify the mechanism which FAWs adapt to different feeding hosts and develop effective control technologies. Two-sex life tables and 16s rDNA sequencing were used to determine the host fitness and gut microbial diversity of FAWs when fed four different food types. Considering the life history parameters, pupa weight, and nutrient utilization indexes, the host fitness of FAWs when fed different food types changed in descending order as follows: artificial diet, maize, wheat, and rice. The gut microbial composition and the diversity of FAWs when fed different food types were significantly different, and those changes were driven by low-abundant bacteria. The gut microbes of FAWs that were fed with maize had the highest diversity. The functions of the gut microbes with significant abundance differences were enriched in nutrient and vitamin metabolism and other pathways that were closely related to host adaptation. Furthermore, we identified five genera (Acinetobacter, Variovorax, Pseudomonas, Bacillus, and Serratia) and one genus (Rahnella) that were positively and negatively correlated with the host fitness, respectively. This study revealed the possible role of gut microbes in the host adaptation of FAWs.
1. Introduction
Due to its wide host range, strong migratory ability, and strong species competitiveness, the fall armyworm (FAW), Spodoptera frugiperda (Smith), seriously threatens food and cash crops, including maize, wheat, and cotton [1,2,3]. Meanwhile, being a major migratory pest, the speed at which this pest invades and spreads is concerning. After infesting Myanmar in late 2018, FAW spread to 27 Chinese provinces within a year and became a dominant pest [4,5].
Researchers have carried out extensive research on the host adaptation of/fitness differences in FAWs in different host plants [6,7]. In general, the FAWs invading China exhibit the highest host adaptability to maize [8,9]. In addition to maize, wheat has become an important wintering host plant of FAWs [10,11]. Furthermore, there have been reports of FAW infestations in rice fields across many areas of China [12,13,14]. The COI-R Tpi-C configuration, detected using the COI and Tpi (MspI) gene combination identification method, represents the primary host biotype that initially arrived in China [15,16]. Due to these reasons, the potential for FAWs to harm food crops other than maize must be considered. Therefore, it is crucial to investigate the adaptation mechanisms of FAWs to different feeding hosts such as maize, wheat, and rice.
In addition to studying the effects of different feeding hosts on insect life history parameters, the consumption and utilization of insects on different feeding hosts should not be ignored in the process of studying the formation mechanism of insect host adaptation [17,18]. When examining the food consumption, utilization, and detoxication enzyme (MFO: mixed-function oxidase) activities of two FAW host strains that fed on maize and rice, the results indicated that both behavioral and physiological factors were associated with host use [19]. The metabolization of foreign chemicals is among the key functions involved in the phenotypic variation in FAW strains [20]. Hafeez et al. [21] indicates the possible roles of S. frugiperda gut digestive protease enzymes and related genes (SfTry-3, SfTry-7, and Sfchym-9) in host plant adaptation. The consumption and use of food by insects not only represent their acceptance but also provide basic conditions for their growth, development, and reproduction, which may be the key factor affecting the formation of insect host adaptation characteristics [22,23].
Gut microbiotas play an important role in plant–insect interactions [24], including insect feeding and host digestion, insect–plant defense and immune resistance, and insect growth and development [25,26,27]. With the introduction of the holo-genome evolutionary theory and the development of high-throughput sequencing [28], more attention has been paid to the role of gut microbes in host adaptability [29,30,31]. There is growing evidence that variation in gut microbes can influence insect host phenotypes [32,33,34]. It has been proved that the Pseudomonas strain in the gut enables Hypothenemus hampei to acquire a caffeine degradation ability, allowing H. hampei to better utilize coffee beans [35]. Although some studies on the gut bacteria of FAWs have been conducted [36], it remains unclear whether the gut microbiotas of FAWs play a role in their adaptation to various host plants.
This research was devoted to investigating the potential role of the gut bacteria of FAWs in adapting to different host plants. In this study, four different food types were used, including an artificial diet and three host plants. The host fitness (life table parameters, pupal weight, and nutrient utilization indexes) of FAWs when feeding on different food types were measured. The gut bacterial compositions of FAWs feeding on different feed types were determined by 16s rDNA sequencing to explore the taxonomic diversity in different groups. By conducting correlation analysis between host fitness and gut bacterial taxa, we aimed to gain a better understanding of the interaction between microbes and insect host adaptations. Our research aimed to uncover the possible role of FAWs’ gut microbiotas in strategies that enable their adaptation to various host plants.
2. Materials and Methods
2.1. Insects and Food Types
FAWs were collected from maize fields in Nanning city, Guangxi Zhuang Autonomous Region (108.37° E, 22.82° N), in April 2019, and subsequently fed an artificial diet (for details, see Table S1) to establish a population for multiple generations under laboratory conditions. To ensure a consistent genetic background, we selected eggs laid after the mating of one adult male and one adult female from the laboratory population for subsequent experiments. After enough eggs were produced, we used the COI and Tpi [MspI] gene combination identification method [37] to determine the biotype of the parents, which both had the COI-R Tpi-C configuration. The environmental conditions in the incubator for raising FAWs were set at 27 ± 1 °C, with a light–dark cycle of 14:10 h and a relative humidity of 60–80%. Adults were fed with a 10% (w/v) honey–water mixture.
The artificial diet and three host plants were used as four different food types. The host plants, maize (Zea mays L.; Zhengdan 958, provided by the Institute of Crop Sciences, the Chinese Academy of Agricultural Sciences), wheat (Triticum aestivum L.; Luyuan 502, provided by the Institute of Crop Sciences, the Chinese Academy of Agricultural Sciences), and rice (Oryza sativa L.; Zhenghan 10, provided by the Henan Academy of Agricultural Sciences), were cultured in a greenhouse at 25 ± 1 °C at the Institute of Plant Protection, the Chinese Academy of Agricultural Sciences, Beijing, China. The host plants were fed to FAWs at the seedling stage (2–3 weeks after emergence).
2.2. Performance of FAWs Feeding on Different Food Types
Differences in the performance of FAWs when feeding on different food types were assessed by measuring the life table parameters, pupal weight, and nutrient utilization indexes.
According to Chi’s study [38], two-sex life tables were established for FAWs fed on four food types, respectively. For each food type, one hundred newly laid eggs from mated females were individually placed into 6-well plates and covered with toilet paper and a lid to prevent escape. Each FAW from the 1st to the 6th instar larva and the pupal stage was kept in the 6-well plate with food. The survival and development times of FAWs in each stage were recorded daily. The newly emerged females were individually paired with young males from the colony in a glass tube (5 × 12 cm diameter × height) and covered with cotton gauze as the oviposition substrate. The pairs were fed 10% (w/v) honey in sterile water. The survival and the number of eggs laid were recorded daily until all FAW adults died. The raw life history data were analyzed based on an age-stage, two-sex life table using TWO-SEX-MSChart software (v2023) [39]. The performances (developmental time of each stage; R0, net reproduction; r, intrinsic rate of increase; λ, finite rate; T, mean generation time; APOP, adult preoviposition period; oviposition days; and mean fecundity) of the FAW groups were calculated using TWO-SEX-MSChart (v2023).
Following the life table establishment, the 4th-day pupal weight of thirty FAWs was measured.
Twenty 6th-1st day FAW larvae that fed on each food type were used to determine the nutrient utilization indexes, which were measured as described in [22]. After a 6 h starvation treatment, each larva continued to feed on the corresponding food type for 24 h. The fresh weight and dry/fresh weight ratio of the larvae before feeding, the fresh weight and dry/fresh weight ratio of the food before feeding, the fresh weight and the dry weight of larvae after feeding for 24 h, including the dry weight of uneaten food and the dry weight of feces, were determined. The calculated indexes included the relative growth rate (RGR), approximate digestibility (AD), relative consumption rate (RCR), the efficiency of conversion for digested food (ECD), and ingested food (ECI). The calculation formula for each index was as follows:
WFI: dry weight of food introduced; WUF: dry weight of uneaten food; WIL: the initial dry weight of the larvae; WFL: the final dry weight of the larvae; WF: dry weight of feces.
2.3. Determining the Diversity of FAW Gut Microbial Communities after Feeding on Different Food Types
To clarify the differences in the gut microbiota diversities of FAWs induced by feeding on different food types, 16S rDNA sequencing was performed on the guts of 6th–1st day FAW larvae with the same genetic background that were continuously fed on different food types for more than five generations.
2.3.1. Sample Collection and DNA Extraction
Four groups of FAW samples were collected and named according to the food types; the groups were as follows: artificial diet, maize, wheat, and rice. For each group, the guts of four 6th-1st day larvae were dissected and placed in a 2 mL sterile tube, with five replicates per group. DNA was extracted using the E.Z.N.A. ®Stool DNA Kit (D4015, Omega, Inc., Norwalk, CT, USA), according to the manufacturer’s recommendations. Nuclease-free water was used as a blank. The total DNA was eluted into 50 μL of elution buffer and stored at −80 °C until PCR amplification by LC-BioTechnology Co., Ltd. (Hangzhou, China).
2.3.2. PCR Amplification and 16S rDNA Sequencing
The V3-V4 region of the 16S rRNA gene was amplified using the universal primers 341F (5′-CCTACGGGNGGCWGCAG-3′) and 806R (5′-GACTACHVGGGTATCTAATCC-3′). PCR amplification was performed in a reaction volume of 25 μL containing 25 ng template DNA, 12.5 μL PCR Premix, 2.5 μL of each primer, and PCR-grade water to adjust the volume. The PCR conditions included an initial denaturation at 98 °C for the 30 s, 32 denaturation cycles at 98 °C for the 10 s, annealing at 54 °C for the 30 s, an extension at 72 °C for 45 s, and the final extension at 72 °C for 10 min. The PCR products were confirmed with 2% agarose gel electrophoresis. Throughout DNA extraction, ultrapure water was used as a negative control to rule out false positives. The PCR products were purified using AMPure XT beads (Beckman Coulter Genomics, Danvers, MA, USA) and quantified by Qubit (Invitrogen, Carlsbad, CA, USA). The amplicon pools were prepared for sequencing, and the amplicon library size and quantity were assessed using the Agilent 2100 Bioanalyzer (Agilent, Santa Clara, CA, USA) and the Illumina Library Quantification Kit (Kapa Biosciences, Wilmington, MA, USA), respectively. The libraries were sequenced on the NovaSeq PE250 platform. The raw sequence reads are in the NCBI SRA database under the Project Accession ID PRJNA939966.
2.4. Data Analysis
The standard errors for each performance and significant differences among FAWs that fed on different food types were estimated using the paired bootstrap test in TWO-SEX-MSChart (v2023). Pupal weight and nutrient utilization indexes were compared using ANOVA, followed by Tukey’s HSD post hoc comparisons in SPSS 25.
For the 16S rDNA sequencing of the four groups of FAW gut microbial communities, paired-end reads were assigned to sequencing data samples based on their unique barcode and were truncated by removing the barcode and primer sequence. Paired-end reads were merged using FLASH. Raw and quality filtering was performed to obtain high-quality clean tags, according to fqtrim (v0.94). Chimeric sequences were filtered using Vsearch (v2.3.4). After dereplication using DADA2 [40], we obtained feature tables and sequences. Alpha and beta diversities were calculated by randomly normalizing them to the same sequences. Then, according to the SILVA (release 132, https://www.arb-silva.de/documentation/release-132/ (accessed on 24 October 2022) classifier, feature abundance was normalized using the relative abundance of each sample. The species diversity of each sample was analyzed using alpha diversity, Chao1, Observed_species, Goods_coverage, Shannon, and Simpson; these indices and the beta diversity were calculated using QIIME2 [41], and all graphs and diagrams were drawn in R (v3.5.2). Blast was used for sequence alignment, and feature sequences were annotated using the SILVA database (release 132). The PICRUSt2 (v2.2.0-b) [42] software (https://github.com/picrust/picrust2 (accessed on 24 October 2022) was used to predict the function of FAWs’ gut microbiota.
Spearman’s correlation was used to identify correlations between the biomarkers, which were selected from different groups of FAW gut microbial taxa, and FAW performance in R (v3.5.2), including the pupal weight (mg) and five nutrient utilization indexes. The relative abundance of gut bacterial taxa and FAW performance were transformed.
3. Results
3.1. Performances of FAWs Feeding on Different Food Types
The development and reproduction of the FAWs feeding on different food types showed significant differences. Compared with other food types, FAWs were fed an artificial diet, which showed the highest R0 (p < 0.05) (Table 1). FAWs feeding on rice had the lowest r and λ (p < 0.05) (Table 1). In comparison to those feeding on maize and wheat, FAWs that were fed the artificial diet and rice had a significantly longer T by approximately 2 days (p < 0.05) (Table 1). Different food types had little effect on the oviposition days of FAWs (p > 0.05), but the mean fecundity of reproductive females of FAWs when feeding on the artificial diet and maize was significantly higher than that of wheat and rice (p < 0.05) (Table 1).
The pupal weight of FAWs feeding on different food types was significantly different as follows: artificial diet > maize > wheat > rice (F(3,119) = 372.45, p < 0.01) (Figure 1).
The water content of different food types varied significantly. Based on the dry/fresh weight ratio, maize had the highest water content, followed by rice and wheat, while the artificial diet had the lowest (F(3,79) = 2001.40, p < 0.01) (Table 2). Therefore, dry weight was used to calculate nutrient use indicators. For each food type, when the 6th–1st day FAWs fed for 24 h, both the fresh weight and dry weight of worm bodies were heaviest with the artificial diet, followed by maize and wheat, and rice had the lightest weight (fresh weight: F(3,79) = 36.17, p < 0.01; dry weight: F(3,79) = 105.39, p < 0.01) (Table 2). FAWs exhibited significant differences in their ability to utilize and digest different food types. The RCR of FAWs when feeding on various food types showed the following pattern: maize > wheat ≈ rice > artificial diet (F(3,79) = 101.27, p < 0.01) (Table 2). In other words, FAWs had the largest relative feed intake and the highest palatability of maize. The RGR of FAWs when feeding on various food types showed the following pattern: maize ≈ artificial diet > wheat > rice (F(3,79) = 118.95, p < 0.01) (Table 2). In other words, the FAWs had the best absorption of maize during the artificial diet, followed by wheat, and the worst absorption was rice. The AD of FAWs when feeding on various food types showed the following pattern: artificial diet > maize > wheat > rice (F(3,79) = 275.06, p < 0.01) (Table 2). In other words, FAWs digested the artificial diet better, followed by maize and wheat, with the least digestible being rice. The ECI of FAWs when feeding on various food types showed the following pattern: artificial diet > maize ≈ wheat > rice (F(3,79) = 244.65, p < 0.01) (Table 2).
3.2. Diversity of FAW Gut Microbial Communities after Feeding on Different Food Types
After performing paired-end assembly, quality control, and chimera filtering on the raw reads, high-quality clean tags were obtained. The detailed statistical information of the raw and valid data for each sample can be found in Table S2. Based on the abundant table of features obtained after removing background noise using QIIME2, the number of common and unique features in each group was calculated. In total, 14, 152, 74, and 58 features were found in the four groups: artificial diet, maize, wheat, and rice, respectively. Only three features were shared among all groups, and 6, 128, 44, and 30 unique features were found in the four groups: artificial diet, maize, wheat, and rice, respectively (Figure S1).
The α-diversity of the four group samples at the feature level was estimated using the Chao1, good-coverage, Shannon, and Simpson indices. There was no difference in the good-coverage index among all groups, which were close to one. This indicates that the probability of new species not being detected in the samples was low and suggests that the sequencing results likely represent the real situation (Wilcoxon test, p < 0.01; Figure S2B). The Chao1, Shannon, and Simpson indices showed significant differences among the four groups (Wilcoxon test for Chao1, p < 0.01; Wilcoxon test for Shannon, p < 0.01; Wilcoxon test, p < 0.01; Figure S2A,C,D). The α-diversity suggests that species richness and community microorganism diversity in gut of FAW larvae were highest in the maize group and lowest in the artificial diet group.
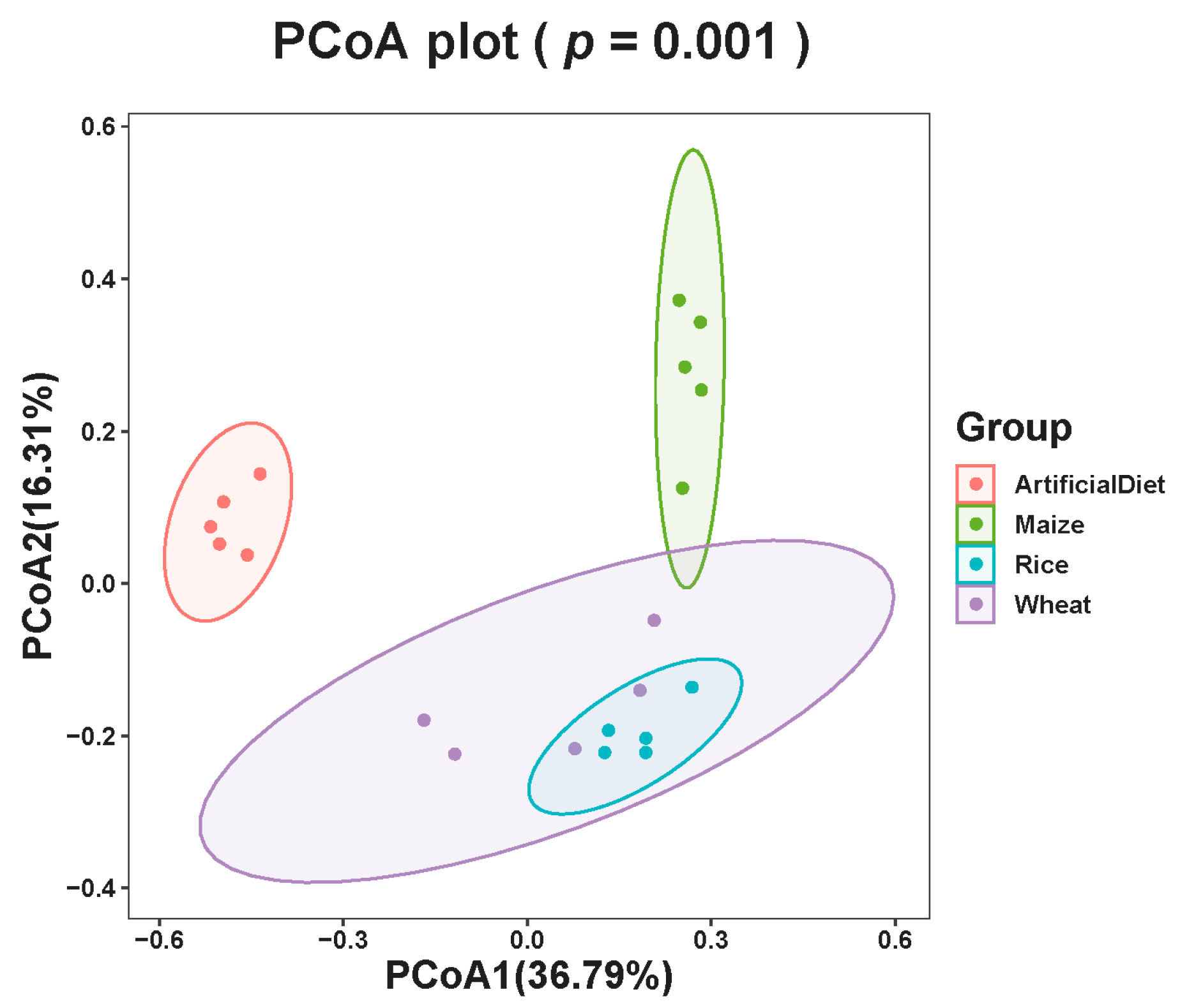
The β-diversity of four group samples at the feature level illustrated the similarities and differences in species composition and community structure. According to the principal coordinate analysis (PCoA), microbial communities were clearly divided into the following groups based on food types: artificial diet, maize, wheat, and rice (adonis, F(3,19) = 7.06, R2 = 0.57, p < 0.01; Figure 2). ANOSIM analysis also shows that feeding different food types significant impact FAWs’ gut bacterial composition (R2 = 0.70, p < 0.01).
In total, 13 phyla, 20 classes, 43 orders, 79 families, and 128 genera were detected in the 244 features of all groups. The community composition was analyzed at the phylum and genus levels (Figure 3). At the phylum level, Firmicutes occupied a dominant position in all groups, especially in the group’s artificial diet and wheat, with an average relative abundance of 99.99 and 99.36%, and 81.74 and 90.53% for groups maize and rice, respectively. Proteobacteria and Cyanobacteria were also the most abundant phyla in the maize group, with an average relative abundance of 10.78 and 7.43%, respectively. Moreover, Proteobacteria was also the most abundant phyla in the rice group, with an average relative abundance of 9.36% (Figure 3A). At the genus level, FAW gut bacterial genera demonstrated different distributions depending on their food types. Enterococcus was a dominant genus in all groups, with an average abundance of more than 81.66%, especially in the artificial diet group, where the average abundance reached 99.98%. In addition to Enterococcus, the top genera with an average abundance of >1% in the maize group were Oxyphotobacteria_unclassified, Enterobacter, and Acinetobacter, while those in group wheat were Turicibacter, and those in the rice group were Enterobacter and Rahnella (Figure 3B). Samples from the maize group showed a more abundant gut microbiota diversity.
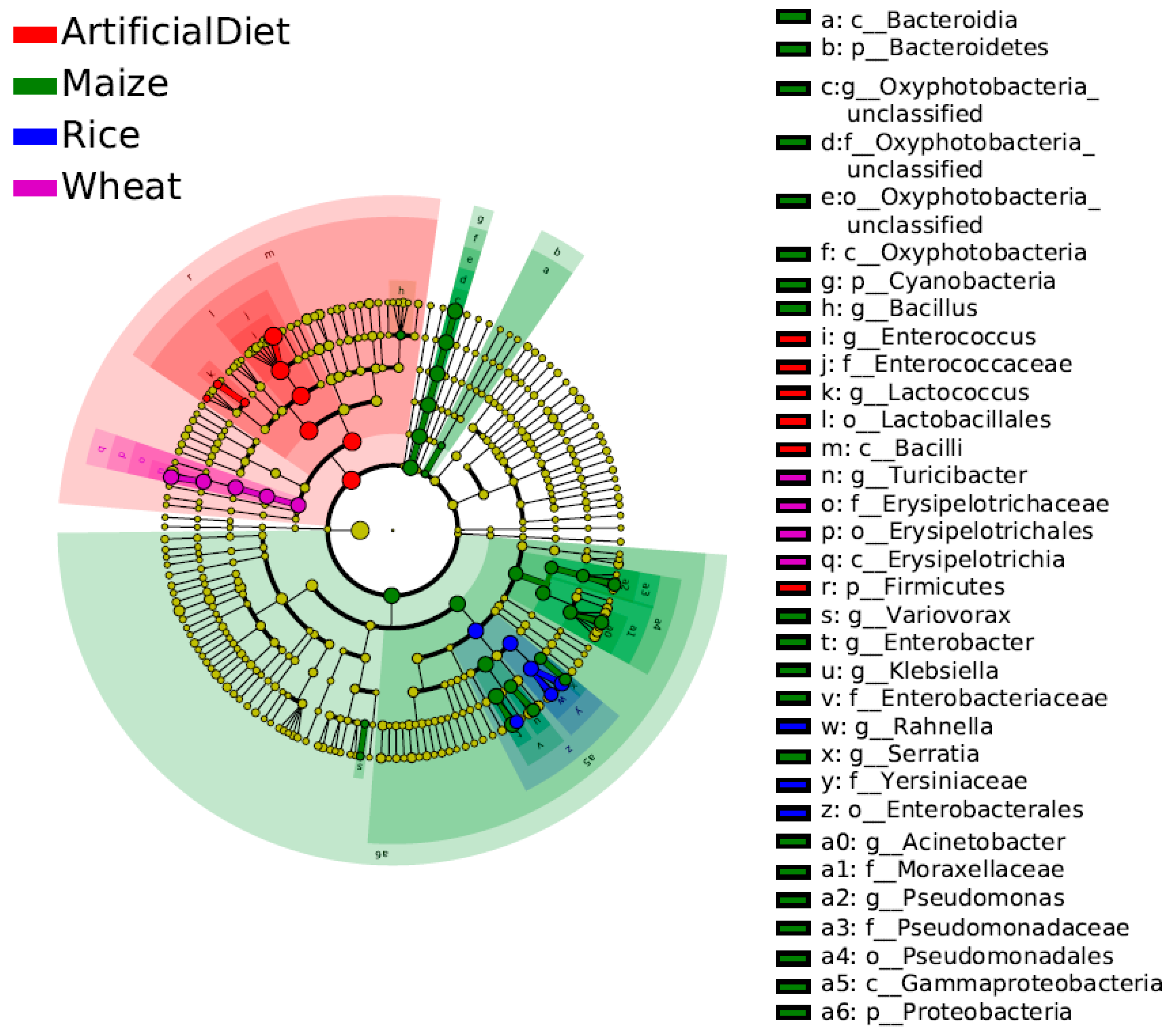
LEfSe (LDA Effect Size) analysis was performed to further explore biomarkers in each group with significant differences in the relative abundance across other groups (Figure 4; LDA scores > 3). At the genus level, Enterococcus and Lactococcus were identified as biomarkers with a higher abundance in the artificial diet group. Oxyphotobacteria_unclassified, Enterobacter, Acinetobacter, Variovorax, Klebsiella, Pseudomonas, Bacillus, and Serratia were found as biomarkers with a higher abundance in the Maize group. Only one biomarker was found with a higher abundance in the Wheat and Rice groups respectively, which was Turicibacter and Rahnella.
Based on PICRUSt2 functional prediction, at KEGG Level 2, the results showed that the functions of gut microbiota with significant differences in abundance across the groups were mainly involved in glycan biosynthesis and metabolism, carbohydrate metabolism, lipid metabolism, amino acid metabolism, cofactors, vitamin metabolism, etc., as well as digestive, endocrine, excretory, and immune systems closely related to insect adaptability (Figure S3).
3.3. Correlations between Gut Microbes and FAW Performance
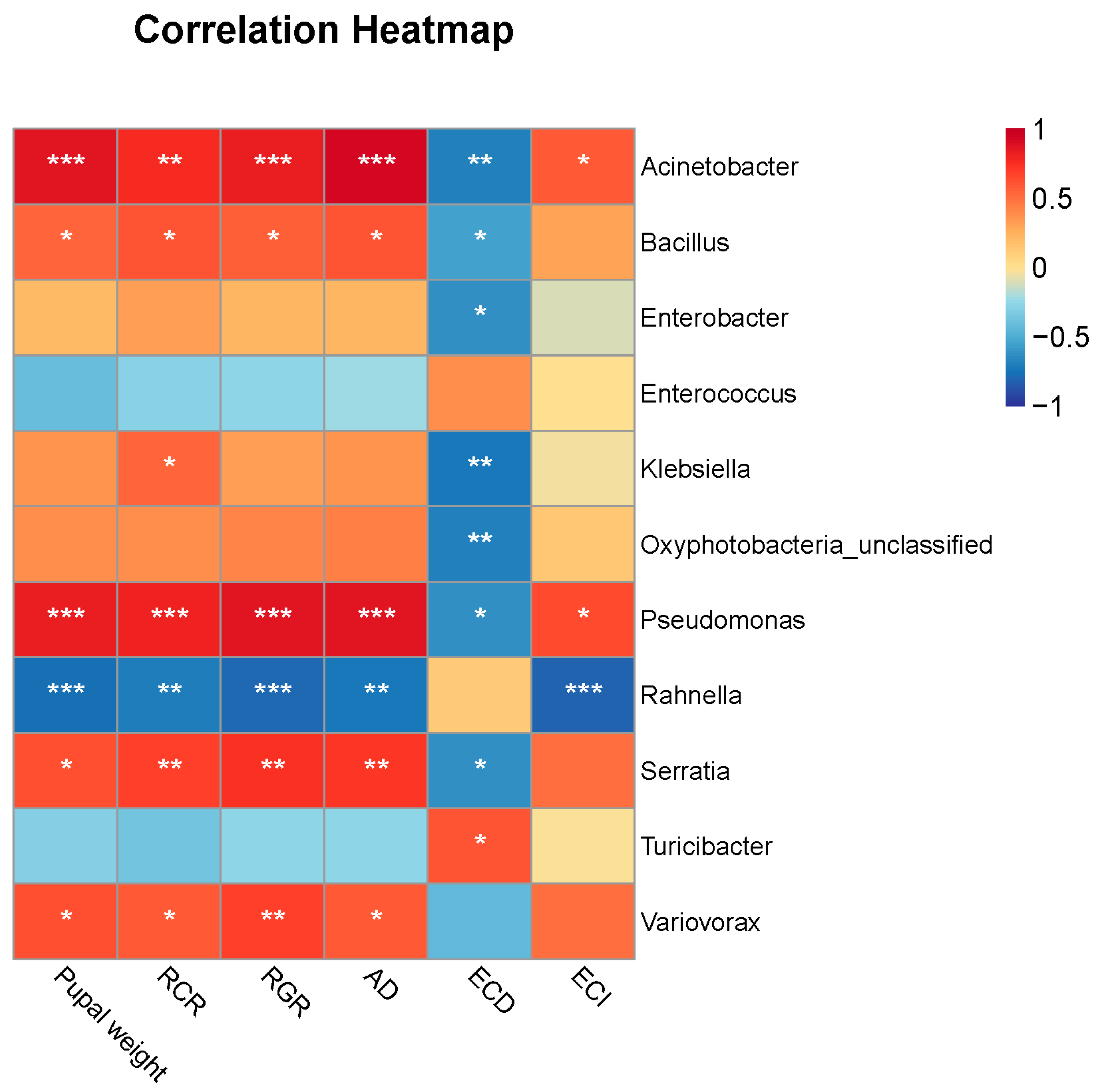
Feeding on different food types changed FAW host adaptation and gut microbiota communities. Therefore, the correlations between gut microbiota and FAW performance were calculated. Among all the biomarkers, the relative abundance of Enterococcus in each group was very high. Therefore, we used biomarkers other than Enterococcus to analyze the correlation with pupal weight and five nutrient utilization indexes of FAW when feeding on different food types via Spearman’s correlation. The result showed that relative abundances of the genera Acinetobacter, Variovorax, Pseudomonas, Bacillus, and Serratia were significantly and positively correlated with FAW pupal weight, RCR, RGR, and AD and also demonstrated a significantly negative correlation with ECD (Figure 5). Conversely, the relative abundance of Rahnella was significantly and negatively correlated with FAW pupal weight, RCR, RGR, AD, and ECI (Figure 5).
4. Discussion
The FAWs’ host range is wide, but their adaptation differs significantly from that of other hosts [6,7]. Herbivore preference and performance can be affected by the varying nutrition profiles and secondary metabolites in plants, which can even induce different levels of insect resistance [43]. In this study, the host fitness of FAWs varied significantly when feeding on four food types. The mean fertility of FAWs was highest when feeding on the artificial diet and maize. When FAWs fed on rice, its reproductive capacity was greatly reduced, resulting in the lowest r, λ, R0, and mean fecundity. Compared to maize and wheat, the mean generation time of FAWs fed the artificial diet or rice was significantly extended by about two days. Generally, shorter developmental times and higher reproduction rates on a particular plant species indicate a higher suitability for that plant [44,45,46]. In other words, FAWs exhibited greater adaptability to maize as a host compared to rice which showed the least adaptability. The prolonged generation time of FAWs when feeding on the artificial diet was more likely caused by the poor palatability of the young larvae whose period of development was prolonged on the artificial diet. Compared with the feeding on host plants, FAWs had the lowest RCR when feeding on the artificial diet, confirming that artificial diets are not highly palatable to FAWs. However, this did not affect the high host adaptability of FAWs to the artificial diet, which showed that they had a similarly high fecundity and even higher pupal weight when feeding on the artificial diet than on maize. The results regarding pupal weight and nutrient utilization indexes (RGR, AD, ECI) also indicated that the host adaptation of FAWs followed this descending order: artificial diet > maize > wheat > rice.
Changes in host plants dramatically alter the gut microbiota of herbivores [47]. Proteobacteria and Firmicutes were identified as the most dominant bacterial phyla in S. frugiperda collected from four maize-growing regions in Kenya [36]. In this study, the composition and diversity of the gut microbiota of FAWs varied significantly when feeding on different food types, with the highest diversity observed when they were fed maize among the four food types. Besides Firmicutes and Proteobacteria, Cyanobacteria was also identified as the dominant bacterial phylum of FAWs fed on maize. Interestingly, Firmicutes was absolutely dominant in all groups. Especially when FAWs were fed the artificial diet, their gut microorganisms were almost all Firmicutes. This suggests that dietary changes did not affect Firmicutes as the core microbiota, and compositional changes in FAWs’ gut microbiota after dietary alterations were driven by low-abundant bacteria [48,49].
The composition of the gut microbiota significantly influenced insect growth and development [50], with the most dominant contributions of insect gut microbiotas including nutrients and vitamins provision, enhanced digestion efficiency, and the detoxification metabolism of allelochemicals [51,52]. Using PICRUSt2 function prediction, our results also showed that the functions of FAWs’ gut microbiota with significant differences in abundance across the groups are mainly enriched in nutrient (glycan, carbohydrate, lipid, and amino acid) and vitamin metabolisms, as well as the detoxification metabolisms of allelochemicals, etc. Through correlation analysis, five genera (Acinetobacter, Variovorax, Pseudomonas, Bacillus, and Serratia) and one genus (Rahnella) showed significant positive and negative correlations with FAW host fitness, respectively. Beneficial gut microbiotas have been shown to be critical for insect host health [27,53,54]. Acinetobacter exhibited enhanced esterase activity and facilitated insecticide metabolism, including cypermethrin and Bt toxins, which could contribute to insect resistance [55,56]. Pseudomonas may benefit bark beetles by providing nutrients, protecting them from chemical defenses, and antagonizing entomopathogenic fungi [57]. Serratia promotes pea aphid development and growth by enhancing fatty acid biosynthesis and metabolism [58]. Although harmful gut bacteria could also adversely affect insects [59,60], the effect of Rahnella on insect fitness has not yet been reported.
Based on the differences in host fitness and the gut microbial composition and diversity of FAWs when feeding on different food types, we can make possible inferences about the host adaptation mechanism of the fall armyworm. When the food source is extremely nutritious, such as the artificial diet, the fall armyworm obtains all kinds of nutrients required for growth, development, and reproduction solely through feeding. However, when feeding on host plants, the fall armyworm cannot obtain all the necessary nutrients through feeding alone. Therefore, it is necessary to adjust the community structure of intestinal microorganisms to assist in synthesizing, utilizing, and metabolizing nutrients (such as glycan, carbohydrate, lipid, and amino acids) to promote normal growth and development [61]. When the host plants are nutrient-deficient, and the diversity of the intestinal microbes is insufficient to compensate for the nutrient deficiency caused by food sources, the fall armyworm shows poor performance. The key gut microbiota which affects insect host adaptation and the specific functions of different microbiota strains require further clarification through bacterial clearance and re-inoculation experiments.
5. Conclusions
Two-sex life tables and 16s rDNA sequencing analysis were used to determine the host fitness and gut microbial composition and diversity of FAWs when feeding on different food types. Our results showed that the food types significantly affect the host fitness of FAWs. Considering the life history parameters, pupa weight and the nutrient utilization indexes, the fitness of fall armyworms feeding on different food types changed from high to low with the following: artificial diet, maize, wheat, and rice. Similarly, the food types also significantly influenced the gut microbial composition and diversity of FAWs, and those changes were primarily caused by low-abundant bacteria. The functional prediction results for gut microbes with significant abundance differences indicated their main functions involvement in nutrient and vitamin metabolism, as well as other pathways closely associated with host adaptation. Interestingly, we identified five gut genera (Acinetobacter, Variovorax, Pseudomonas, Bacillus, and Serratia) and one genus (Rahnella) that were positively and negatively correlated with the host fitness, respectively. This study revealed the possible role of gut microbes in the host adaptation of FAWs, providing a reference for further exploration into the adaptive mechanisms of FAWs to their hosts and offering a new perspective on pest control.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/insects15050304/s1. Table S1: Artificial diet formulation for FAWs; Table S2: Statistical table of raw and valid data for 16s rDNA sequencing of gut samples from FAWs larvae feeding on different food types; Figure S1: Venn diagrams of features shared among the guts of FAW larvae feeding on different food types; Figure S2: α-diversity of gut bacteria in FAWs feeding on different food types; Figure S3: PICRUSt2 functional prediction of FAW gut bacteria at KEGG level 2 (t-test, p < 0.05).
Author Contributions
Conceiving and designing the experiments, L.M., X.J., Y.C. and L.Z.; performing the experiments, L.M., D.W., Q.R. and J.S.; analyzing the data, L.M., D.W., Q.R. and J.S.; writing the paper, L.M. and X.J.; project administration, L.M., X.J., Y.C. and L.Z.; funding acquisition, X.J., Y.C. and L.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the National Key Research and Development Program of China [2021YFD1400701, 2022YFD1400601], National Natural Science Foundation of China (32072420, 31871951), China Agriculture Research System of MOF and MARA [CARS-22], Government Procurement of Public Services of MARA (15226006), and Basic Research Funds of Chinese Academy of Agricultural Sciences (S2022XM05).
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Acknowledgments
We are grateful for the assistance of all staff members and students in the State Key Laboratory for Biology of Plant Diseases and Insect Pests, Institute of Plant Protection, the Chinese Academy of Agricultural Science in Beijing, China.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Sparks, A.N. A review of the biology of the fall armyworm. Fla. Entomol. 1979, 62, 82–87. [Google Scholar] [CrossRef]
- Westbrook, J.K.; Nagoshi, R.N.; Meagher, R.L.; Fleischer, S.J.; Jairam, S. Modeling seasonal migration of fall armyworm moths. Int. J. Biometeorol. 2016, 60, 255–267. [Google Scholar] [CrossRef]
- Montezano, D.G.; Specht, A.; Sosa-Gómez, D.R.; Roque-Specht, V.F.; Sousa-Silva, J.C.; Paula-Moraes, S.V.; Peterson, J.A.; Hunt, T.E. Host plants of Spodoptera frugiperda (Lepidoptera: Noctuidae) in the Americas. Afr. Entomol. 2018, 26, 286–300. [Google Scholar] [CrossRef]
- Jiang, Y.Y.; Liu, J.; Xie, M.C.; Li, Y.H.; Zhang, M.L.; Qiu, K. Observation on law of diffusion damage of Spodoptera frugiperda in China in 2019. Plant Prot. 2019, 45, 10–19. [Google Scholar] [CrossRef]
- Sun, X.X.; Hu, C.X.; Jia, H.R.; Wu, Q.L.; Shen, X.J.; Zhao, S.Y.; Jiang, Y.Y.; Wu, K.M. Case study on the first immigration of fall armyworm, Spodoptera frugiperda invading into China. J. Integr. Agric. 2021, 20, 664–672. [Google Scholar] [CrossRef]
- Guo, J.F.; Zhang, M.D.; Gao, Z.P.; Wang, D.J.; He, K.L.; Wang, Z.Y. Comparison of larval performance and oviposition preference of Spodoptera frugiperda among three host plants: Potential risks to potato and tobacco crops. Insect Sci. 2021, 28, 602–610. [Google Scholar] [CrossRef]
- He, L.M.; Wu, Q.L.; Gao, X.W.; Wu, K.M. Population life tables for the invasive fall armyworm, Spodoptera frugiperda fed on major oil crops planted in China. J. Integr. Agric. 2021, 20, 745–754. [Google Scholar] [CrossRef]
- Wu, Z.W.; Shi, P.Q.; Zeng, Y.H.; Huang, W.F.; Huang, Q.Z.; Ma, X.H.; Guo, L.Z. Population life tables of Spodoptera frugiperda (Lepidoptera: Noctuidae) fed on three host plants. Plant Prot. 2019, 45, 59–64. [Google Scholar] [CrossRef]
- Xu, P.J.; Zhang, D.D.; Wang, J.; Wu, K.M.; Wang, X.W.; Wang, X.F.; Ren, G.W. The host perference of Spodoptera frugiperda on maize and tobacco. Plant Prot. 2019, 45, 61–64. [Google Scholar] [CrossRef]
- Yang, X.M.; Sun, X.X.; Zhao, S.Y.; Li, J.Y.; Chi, X.C.; Jiang, Y.Y.; Wu, K.M. Population occurrence, spatial distribution and sampling technique of fall armyworm Spodoptera frugiperda in wheat fields. Plant Prot. 2020, 46, 10–16. [Google Scholar] [CrossRef]
- Li, Y.P.; Li, M.; Liu, H.H.; Xiao, Q.; Li, X.Y. Occurrence and control of Spodoptera frugiperda in early sowing wheat field in northern Jiangsu province. Plant Prot. 2020, 46, 212–215. [Google Scholar] [CrossRef]
- Xu, L.N.; Chen, Y.T.; Xu, T.T.; Bi, S.J.; Tong, Q.; Hu, B.J.; Hu, F.; Wang, Z.Y. Fall armyworm damaging rice seedlings in Anhui province. Plant Prot. 2022, 48, 310–313. [Google Scholar] [CrossRef]
- Yang, J.J.; Tao, Y.Q.; LIu, Q.; Zheng, Z.W.; Zhou, H.Z. Rice seedlings were damaged by fall armyworm in Wuxue of Hubei province. China Plant Prot. 2020, 40, 44–45. (In Chinese) [Google Scholar]
- Zhang, H. Preliminary report of rice seedlings affected by fall armyworm and biotype identification in Yunxiao of Fujian province. China Plant Prot. 2020, 40, 41–43+53. (In Chinese) [Google Scholar]
- Zhang, L.; Jin, M.H.; Zhang, D.D.; Jiang, Y.Y.; Liu, J.; Wu, K.M.; Xiao, Y.T. Molecular identification of invasive fall armyworm Spodoptera frugiperda in Yunnan Province. Plant Prot. 2019, 45, 19–24. (In Chinese) [Google Scholar] [CrossRef]
- Xu, L.N.; Hu, B.J.; Su, J.Y.; Qi, R.D.; Su, W.H.; Qiu, K.; Zhou, Z.Y.; Zheng, Z.Y.; Zhang, Q.Y.; Hu, F.; et al. Genetic analysis of the fall armyworm Spodoptera frugiperda invaded in Anhui province. Plant Prot. 2019, 45, 47–53. [Google Scholar] [CrossRef]
- Ansari, M.S.; Hasan, F.; Ahmad, N. Influence of various host plants on the consumption and utilization of food by Pieris brassicae (Linn.). Bull. Entomol. Res. 2012, 102, 231–237. [Google Scholar] [CrossRef]
- Truzi, C.C.; Holzhausen, H.G.; Alvaro, J.C.; De Laurentis, V.L.; Vieira, N.F.; Vacari, A.M.; De Bortoli, S.A. Food consumption utilization, and life history parameters of Helicoverpa armigera (Lepidoptera: Noctuidae) reared on diets of varying protein level. J. Insect Sci. 2019, 19, 12. [Google Scholar] [CrossRef]
- Veenstra, K.H.; Pashley, D.P.; Ottea, J.A. Host-plant adaptation in fall armyworm host strains: Comparison of food consumption, utilization, and detoxication enzyme activities. Ann. Entomol. Soc. Am. 1995, 88, 80–91. [Google Scholar] [CrossRef]
- Silva-Brandão, K.L.; Horikoshi, R.J.; Bernardi, D.; Omoto, C.; Figueira, A.; Brandao, M.M. Transcript expression plasticity as a response to alternative larval host plants in the speciation process of corn and rice strains of Spodoptera frugiperda. BMC Genom. 2017, 18, 792. [Google Scholar] [CrossRef]
- Hafeez, M.; Li, X.W.; Zhang, J.M.; Zhang, Z.J.; Huang, J.; Wang, L.K.; Khan, M.M.; Shah, S.; Fernandez-Grandon, G.M.; Lu, Y.B. Role of digestive protease enzymes and related genes in host plant adaptation of a polyphagous pest, Spodoptera frugiperda. Insect Sci. 2021, 28, 611–626. [Google Scholar] [CrossRef] [PubMed]
- Waldbauer, G.P. The consumption and utilization of food by insects. In Advances in Insect Physiology; Beament, J.W.L., Treherne, J.E., Wigglesworth, V.B., Eds.; Academic Press: Cambridge, MA, USA, 1968; Volume 5, pp. 229–288. [Google Scholar] [CrossRef]
- Parra, J.R.P.; Panizzi, A.R.; Haddad, M.L. Nutritional indices for measuring insect food intake and utilization. In Insect Bioecology and Nutrition for Integrated Pest Management; Panizzi, A.R., Parra, J.R., Eds.; CRC Press: Boca Raton, FL, USA, 2012; pp. 13–49. [Google Scholar] [CrossRef]
- Biere, A.; Bennett, A.E.; Fox, C. Three-way interactions between plants, microbes and insects. Funct. Ecol. 2013, 27, 567–573. [Google Scholar] [CrossRef]
- Engel, P.; Moran, N.A. The gut microbiota of insects—Diversity in structure and function. FEMS Microbiol. Rev. 2013, 37, 699–735. [Google Scholar] [CrossRef] [PubMed]
- Itoh, H.; Tago, K.; Hayatsu, M.; Kikuchi, Y. Detoxifying symbiosis: Microbe-mediated detoxification of phytotoxins and pesticides in insects. Nat. Prod. Rep. 2018, 35, 434–454. [Google Scholar] [CrossRef] [PubMed]
- Acevedo, F.E.; Peiffer, M.; Tan, C.W.; Stanley, B.A.; Stanley, A.; Wang, J.; Jones, A.G.; Hoover, K.; Rosa, C.; Luthe, D.; et al. Fall armyworm-associated gut bacteria modulate plant defense responses. Mol. Plant-Microbe Interact. 2017, 30, 127–137. [Google Scholar] [CrossRef] [PubMed]
- Zilber-Rosenberg, I.; Rosenberg, E. Role of microorganisms in the evolution of animals and plants: The hologenome theory of evolution. FEMS Microbiol. Rev. 2008, 32, 723–735. [Google Scholar] [CrossRef] [PubMed]
- Anand, A.A.P.; Vennison, S.J.; Sankar, S.G.; Prabhu, D.I.G.; Vasan, P.T.; Raghuraman, T.; Geoffrey, C.J.; Vendan, S.E. Isolation and characterization of bacteria from the gut of Bombyx mori that degrade cellulose, xylan, pectin and starch and their impact on digestion. J. Insect Sci. 2010, 10, 20. [Google Scholar] [CrossRef]
- Biere, A.; Tack, A.J.M.; Bennett, A. Evolutionary adaptation in three-way interactions between plants, microbes and arthropods. Funct. Ecol. 2013, 27, 646–660. [Google Scholar] [CrossRef]
- Ge, S.X.; Shi, F.M.; Pei, J.H.; Hou, Z.H.; Zong, S.X.; Ren, L.L. Gut bacteria associated with Monochamus saltuarius (Coleoptera: Cerambycidae) and their possible roles in host plant adaptations. Front. Microbiol. 2021, 12, 687211. [Google Scholar] [CrossRef]
- Bost, A.; Martinson, V.G.; Franzenburg, S.; Adair, K.L.; Albasi, A.; Wells, M.T.; Douglas, A.E. Functional variation in the gut microbiome of wild Drosophila populations. Mol. Ecol. 2018, 27, 2834–2845. [Google Scholar] [CrossRef]
- Schretter, C.E.; Vielmetter, J.; Bartos, I.; Marka, Z.; Marka, S.; Argade, S.; Mazmanian, S.K. A gut microbial factor modulates locomotor behaviour in Drosophila. Nature 2018, 563, 402–406. [Google Scholar] [CrossRef] [PubMed]
- Douglas, A.E. Gut microbes alter the walking activity of fruit flies. Nature 2018, 563, 2. [Google Scholar] [CrossRef] [PubMed]
- Ceja-Navarro, J.A.; Vega, F.E.; Karaoz, U.; Hao, Z.; Jenkins, S.; Lim, H.C.; Kosina, P.; Infante, F.; Northen, T.R.; Brodie, E.L. Gut microbiota mediate caffeine detoxification in the primary insect pest of coffee. Nat. Commun. 2015, 6, 7618. [Google Scholar] [CrossRef] [PubMed]
- Gichuhi, J.; Sevgan, S.; Khamis, F.; Van den Berg, J.; du Plessis, H.; Ekesi, S.; Herren, J.K. Diversity of fall armyworm, Spodoptera frugiperda and their gut bacterial community in Kenya. PeerJ 2020, 8, e8701. [Google Scholar] [CrossRef] [PubMed]
- Nagoshi, R.N. Improvements in the identification of strains facilitate population studies of fall armyworm subgroups. Ann. Entomol. Soc. Am. 2012, 105, 351–358. [Google Scholar] [CrossRef]
- Chi, H. Life-table analysis incorporating both sexes and variable development rates among individuals. Environ. Entomol. 1988, 17, 26–34. [Google Scholar] [CrossRef]
- Chi, H. TWOSEX-MSChart: A Computer Program for the Age-Stage, Two-Sex Life Table Analysis; National Chung Hsing University: Taichung, Taiwan, 2023. [Google Scholar]
- Callahan, B.; McMurdie, P.; Rosen, M.; Han, A.; Johnson, A.; Holmes, S. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [PubMed]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef] [PubMed]
- Langille, M.; Zaneveld, J.; Caporaso, J.; McDonald, D.; Knights, D.; Reyes, J.A.; Clemente, J.C.; Burkepile, D.E.; Thurber, R.L.V.; Knight, R.; et al. Predictive functional profiling of microbial communities using 16S rRNA marker gene sequences. Nat. Biotechnol. 2013, 31, 814–821. [Google Scholar] [CrossRef]
- Couture, J.J.; Mason, C.J.; Habeck, C.W.; Lindroth, R.L. Behavioral and morphological responses of an insect herbivore to low nutrient quality are inhibited by plant chemical defenses. Arthropod-Plant Interact. 2016, 10, 341–349. [Google Scholar] [CrossRef]
- Awmack, C.S.; Leather, S.R. Host plant quality and fecundity in herbivorous insects. Annu. Rev. Entomol. 2002, 47, 817–844. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.W.; He, P.Y.; Zhang, Y.Y.; Liu, T.X.; Jing, X.F.; Zhang, S.Z. The population growth of Spodoptera frugiperda on six cash crop species and implications for its occurrence and damage potential in China. Insects 2020, 11, 639. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.H.; Zhou, C.; Long, G.Y.; Yang, X.B.; Wei, Z.Y.; Liao, Y.J.; Yang, H.; Hu, C.X. Fitness of fall armyworm, Spodoptera frugiperda to three solanaceous vegetables. J. Integr. Agric. 2021, 20, 755–763. [Google Scholar] [CrossRef]
- Moran, N.A.; Ochman, H.; Hammer, T.J. Evolutionary and ecological consequences of gut microbial communities. Annu. Rev. Ecol. Evol. Syst. 2019, 50, 451–475. [Google Scholar] [CrossRef] [PubMed]
- Mikaelyan, A.; Dietrich, C.; Kohler, T.; Poulsen, M.; Sillam-Dusses, D.; Brune, A. Diet is the primary determinant of bacterial community structure in the guts of higher termites. Mol. Ecol. 2015, 24, 5284–5295. [Google Scholar] [CrossRef]
- Benjamino, J.; Lincoln, S.; Srivastava, R.; Graf, J. Low-abundant bacteria drive compositional changes in the gut microbiota after dietary alteration. Microbiome 2018, 6, 86. [Google Scholar] [CrossRef] [PubMed]
- Ruokolainen, L.; Ikonen, S.; Makkonen, H.; Hanski, I. Larval growth rate is associated with the composition of the gut microbiota in the Glanville fritillary butterfly. Oecologia 2016, 181, 895–903. [Google Scholar] [CrossRef] [PubMed]
- Graf, J. Lessons from digestive-tract symbioses between bacteria and invertebrates. Annu. Rev. Microbiol. 2016, 70, 375–393. [Google Scholar] [CrossRef]
- Jing, T.Z.; Qi, F.H.; Wang, Z.Y. Most dominant roles of insect gut bacteria: Digestion, detoxification, or essential nutrient provision? Microbiome 2020, 8, 38. [Google Scholar] [CrossRef]
- Dillon, R.J.; Dillon, V.M. The gut bacteria of insects: Nonpathogenic interactions. Annu. Rev. Entomol. 2004, 49, 71–92. [Google Scholar] [CrossRef]
- Berasategui, A.; Salem, H.; Paetz, C.; Santoro, M.; Gershenzon, J.; Kaltenpoth, M.; Schmidt, A. Gut microbiota of the pine weevil degrades conifer diterpenes and increases insect fitness. Mol. Ecol. 2017, 26, 4099–4110. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Xu, X.; De Mandal, S.; Shakeel, M.; Hua, Y.; Shoukat, R.F.; Fu, D.; Jin, F. Gut microbiota mediate Plutella xylostella susceptibility to Bt Cry1Ac protoxin is associated with host immune response. Environ. Pollut. 2021, 271, 116271. [Google Scholar] [CrossRef] [PubMed]
- Zhan, H.; Wang, H.; Liao, L.; Feng, Y.; Fan, X.; Zhang, L.; Chen, S. Kinetics and novel degradation pathway of permethrin in Acinetobacter baumannii ZH-14. Front. Microbiol. 2018, 9, 98. [Google Scholar] [CrossRef] [PubMed]
- Saati-Santamaria, Z.; Rivas, R.; Kolarik, M.; Garcia-Fraile, P. A new perspective of Pseudomonas-Host Interactions: Distribution and potential ecological functions of the genus Pseudomonas within the bark beetle holobiont. Biology 2021, 10, 164. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Ling, X.; Guo, H.; Zhu-Salzman, K.; Ge, F.; Sun, Y. Serratia symbiotica enhances fatty acid metabolism of pea aphid to promote host development. Int. J. Mol. Sci. 2021, 22, 5951. [Google Scholar] [CrossRef] [PubMed]
- Gould, A.L.; Zhang, V.; Lamberti, L.; Jones, E.W.; Obadia, B.; Korasidis, N.; Gavryushkin, A.; Carlson, J.M.; Beerenwinkel, N.; Ludington, W.B. Microbiome interactions shape host fitness. Proc. Natl. Acad. Sci. USA 2018, 115, E11951–E11960. [Google Scholar] [CrossRef] [PubMed]
- Fast, D.; Kostiuk, B.; Foley, E.; Pukatzki, S. Commensal pathogen competition impacts host viability. Proc. Natl. Acad. Sci. USA 2018, 115, 7099–7104. [Google Scholar] [CrossRef]
- Luo, J.; Cheng, Y.; Guo, L.; Wang, A.; Lu, M.; Xu, L. Variation of gut microbiota caused by an imbalance diet is detrimental to bugs’ survival. Sci. Total Environ. 2021, 771, 144880. [Google Scholar] [CrossRef]
Figure 1.
Pupal weight of FAWs feeding on different food types. Lowercase letters indicate the significance of the difference (compared using ANOVA, followed by Tukey’s HSD post hoc comparisons); the error bars indicate the standard errors.
Figure 1.
Pupal weight of FAWs feeding on different food types. Lowercase letters indicate the significance of the difference (compared using ANOVA, followed by Tukey’s HSD post hoc comparisons); the error bars indicate the standard errors.

Figure 2.
PCoA of gut bacteria in FAW fed different food types based on Bray–Curtis distances generated from feature tables. Different colors represent different groupings (adonis).
Figure 2.
PCoA of gut bacteria in FAW fed different food types based on Bray–Curtis distances generated from feature tables. Different colors represent different groupings (adonis).

Figure 3.
Relative abundance of gut microbiota phyla (A) and genera (B) of FAW larvae when fed different food types. Different colors represent the relative abundance of gut microbiota. ‘AD’ stands for artificial diet.
Figure 3.
Relative abundance of gut microbiota phyla (A) and genera (B) of FAW larvae when fed different food types. Different colors represent the relative abundance of gut microbiota. ‘AD’ stands for artificial diet.

Figure 4.
LEfSe analysis of gut microbiota in FAW larvae when fed different food types (LDA scores > 3). Different colors represent different groups; the phyla with significant differences are directly marked in the figure, and the significant differences in other levels are identified by letters.
Figure 4.
LEfSe analysis of gut microbiota in FAW larvae when fed different food types (LDA scores > 3). Different colors represent different groups; the phyla with significant differences are directly marked in the figure, and the significant differences in other levels are identified by letters.

Figure 5.
Correlations between biomarkers and FAW performance. Colors showed correlation coefficients. * p < 0.05, ** p < 0.01, *** p < 0.001.
Figure 5.
Correlations between biomarkers and FAW performance. Colors showed correlation coefficients. * p < 0.05, ** p < 0.01, *** p < 0.001.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Life table parameters of FAWs feeding on different food types.
| Parameter | Artificial Diet | Maize | Wheat | Rice | ||||
|---|---|---|---|---|---|---|---|---|
| Intrinsic rate of increase, r (d−1) | 0.19 ± 0.00 a | 0.18 ± 0.00 a | 0.19 ± 0.00 a | 0.15 ± 0.01 b | ||||
| Finite rate of increase, λ (d−1) | 1.21 ± 0.00 a | 1.20 ± 0.00 a | 1.21 ± 0.00 a | 1.17 ± 0.01 b | ||||
| Net reproductive rate, R0 (offspring) | 573.01 ± 95.19 a | 306.99 ± 65.71 bc | 321.83 ± 61.13 b | 162.15 ± 39.31 c | ||||
| Mean generation time, T (d) | 32.79 ± 0.28 a | 30.79 ± 0.31 b | 30.08 ± 0.38 b | 32.98 ± 0.69 a | ||||
| APOP (for female) | n | days | n | days | n | days | n | days |
| 28 | 4.03 ± 0.17 ab | 20 | 3.80 ± 0.22 ab | 25 | 3.60 ± 0.21 b | 16 | 4.50 ± 0.35 a | |
| Oviposition days (for female) | n | days | n | days | n | days | n | days |
| 37 | 6.82 ± 0.22 a | 21 | 6.70 ± 0.46 a | 35 | 7.04 ± 0.67 a | 21 | 7.81 ± 0.59 a | |
| Mean fecundity (for reproductive female) | n | offspring/ individual | n | Offspring/ individual | n | offspring/ individual | n | offspring/ individual |
| 37 | 1548.68 ± 161.33 a | 21 | 1461.86 ± 134.09 a | 35 | 919.51 ± 122.08 b | 21 | 772.14 ± 114.69 b | |
Lowercase letters indicate the significance of the difference (compared using the paired bootstrap test, p-value < 0.05); n represents the sample size.
Table 2.
The nutrient utilization indexes of FAWs when feeding on different food types.
| Indexes | Food Types | |||
|---|---|---|---|---|
| Artificial Diet | Maize | Wheat | Rice | |
| Dry/fresh weight ratio of food (%) | 21.32 ± 0.10 a | 8.24 ± 0.08 d | 14.19 ± 0.08 c | 14.97 ± 0.18 b |
| Fresh weight of FAW larvae after feeding for 24 h (mg) | 322.48 ± 8.64 a | 258.55 ± 4.79 b | 262.77 ± 7.74 b | 221.86 ± 5.86 c |
| Dry weight of FAW larvae after feeding for 24 h (mg) | 58.14 ± 1.97 a | 43.80 ± 1.27 b | 40.12 ± 1.14 b | 24.75 ± 0.60 c |
| RCR (%) | 194.83 ± 5.35 c | 381.10 ± 6.58 a | 294.33 ± 11.34 b | 271.94 ± 5.61 b |
| RGR (%) | 74.13 ± 1.24 a | 76.94 ± 1.07 a | 56.87 ± 2.64 b | 38.25 ± 1.06 c |
| AD (%) | 68.39 ± 1.28 a | 45.04 ± 1.16 b | 31.87 ± 0.73 c | 26.51 ± 1.24 d |
| ECD (%) | 56.87 ± 2.29 a | 45.95 ± 2.06 b | 61.16 ± 1.99 a | 55.66 ± 3.15 a |
| ECI (%) | 38.51 ± 1.08 a | 20.29 ± 0.44 b | 19.36 ± 0.55 b | 14.14 ± 0.43 c |
Lowercase letters indicate the significance of difference (compared using ANOVA, followed by Tukey’s HSD post hoc comparisons).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Ma, L.; Wang, D.; Ren, Q.; Sun, J.; Zhang, L.; Cheng, Y.; Jiang, X. Gut Microbiota Affects Host Fitness of Fall Armyworm Feeding on Different Food Types. Insects 2024, 15, 304. https://doi.org/10.3390/insects15050304
AMA Style
Ma L, Wang D, Ren Q, Sun J, Zhang L, Cheng Y, Jiang X. Gut Microbiota Affects Host Fitness of Fall Armyworm Feeding on Different Food Types. Insects. 2024; 15(5):304. https://doi.org/10.3390/insects15050304
Chicago/Turabian StyleMa, Lin, Daotong Wang, Qilin Ren, Jiaqi Sun, Lei Zhang, Yunxia Cheng, and Xingfu Jiang. 2024. "Gut Microbiota Affects Host Fitness of Fall Armyworm Feeding on Different Food Types" Insects 15, no. 5: 304. https://doi.org/10.3390/insects15050304
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.