Persistence of the Gypsy Moth Pheromone, Disparlure, in the Environment in Various Climates

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
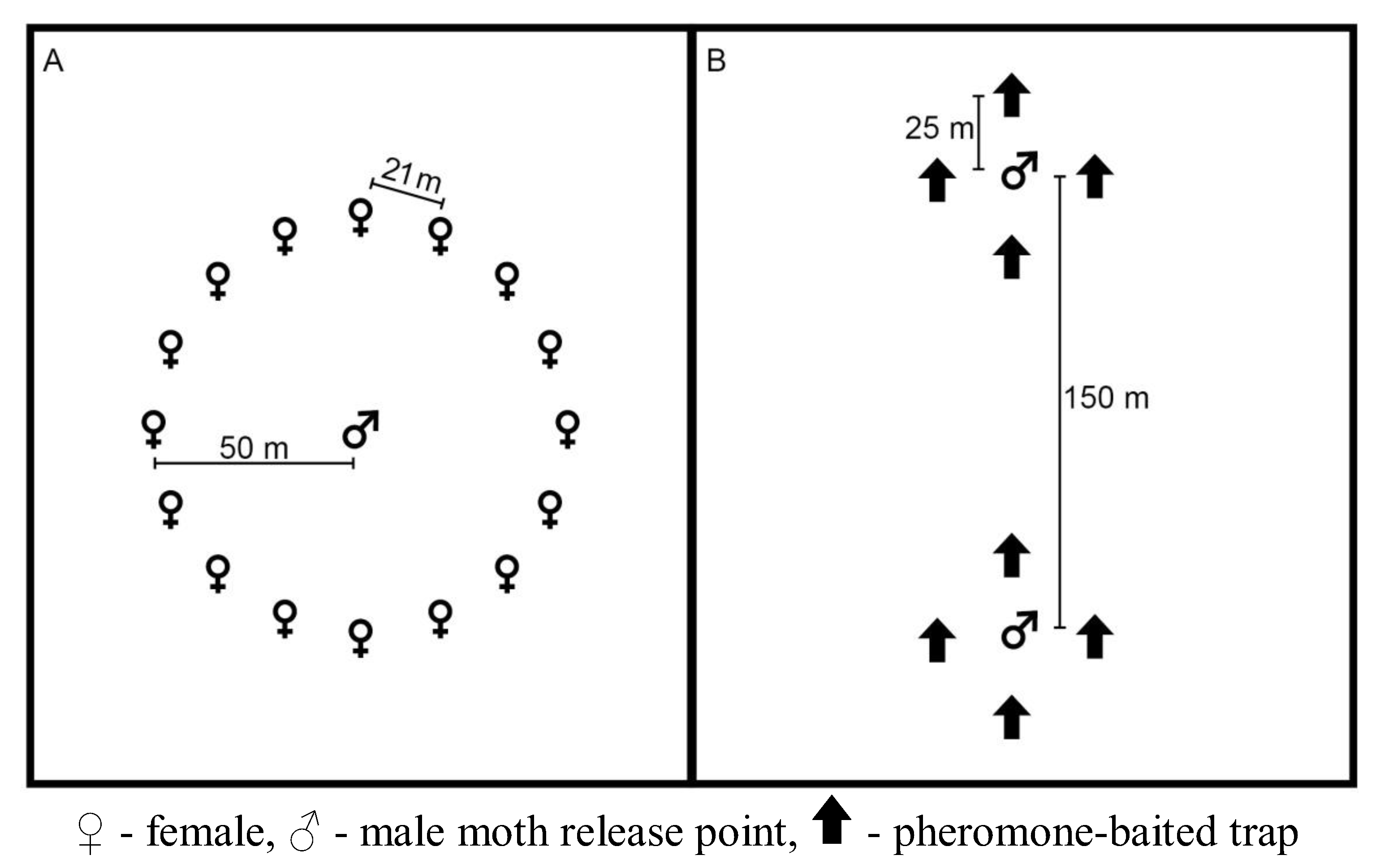
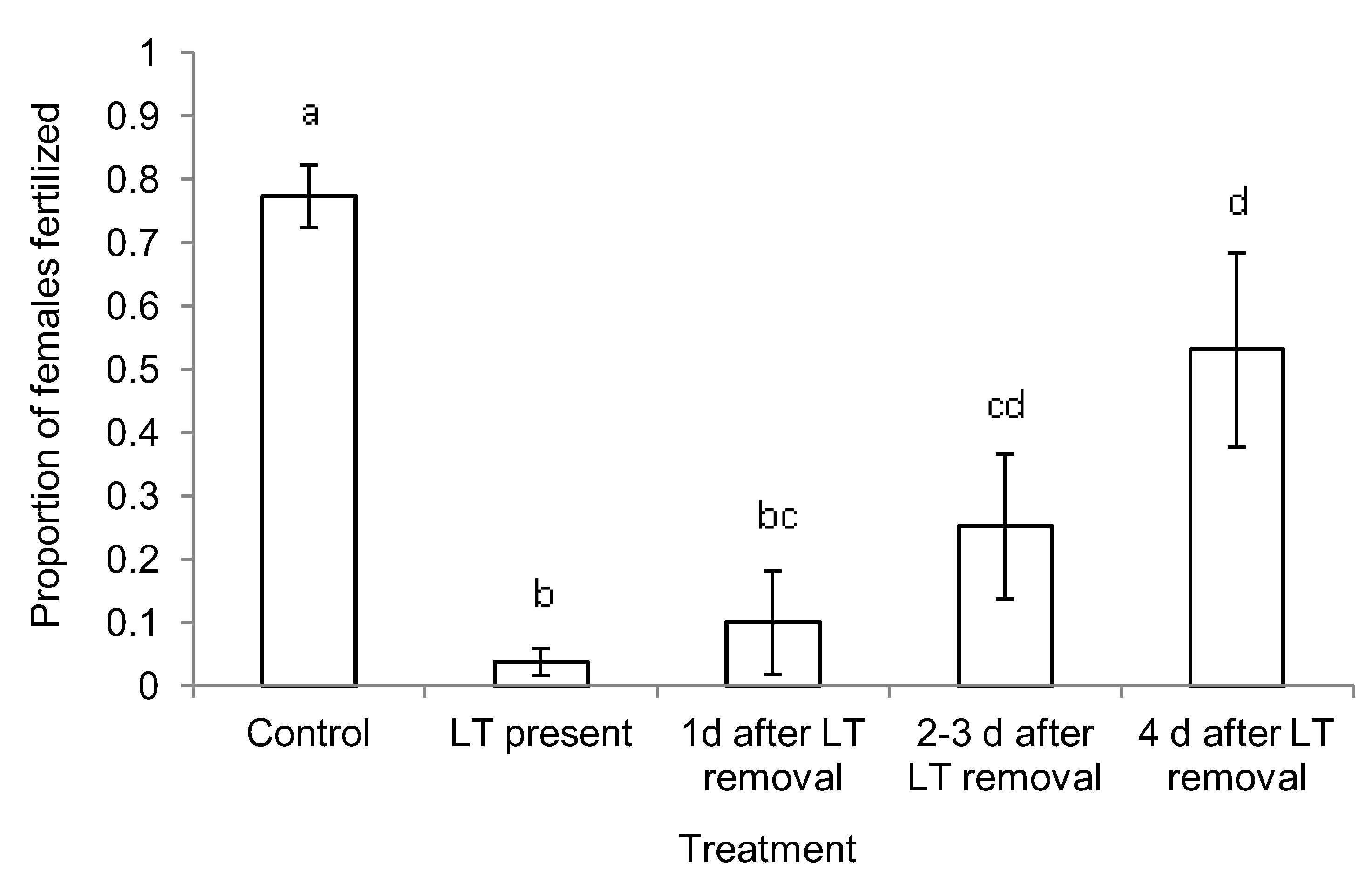
2.1. Mechanism of Short- and Long-term Persistence
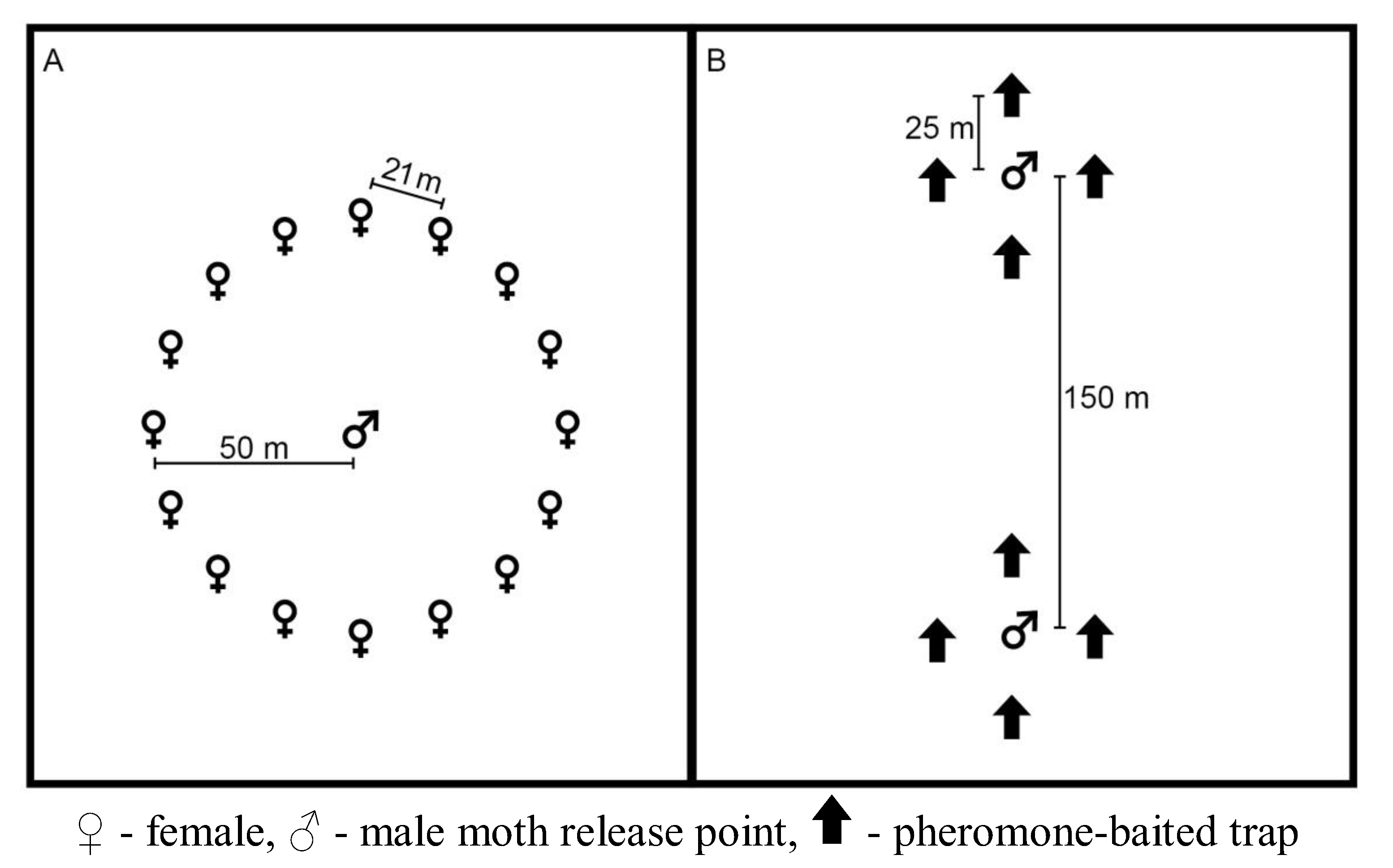
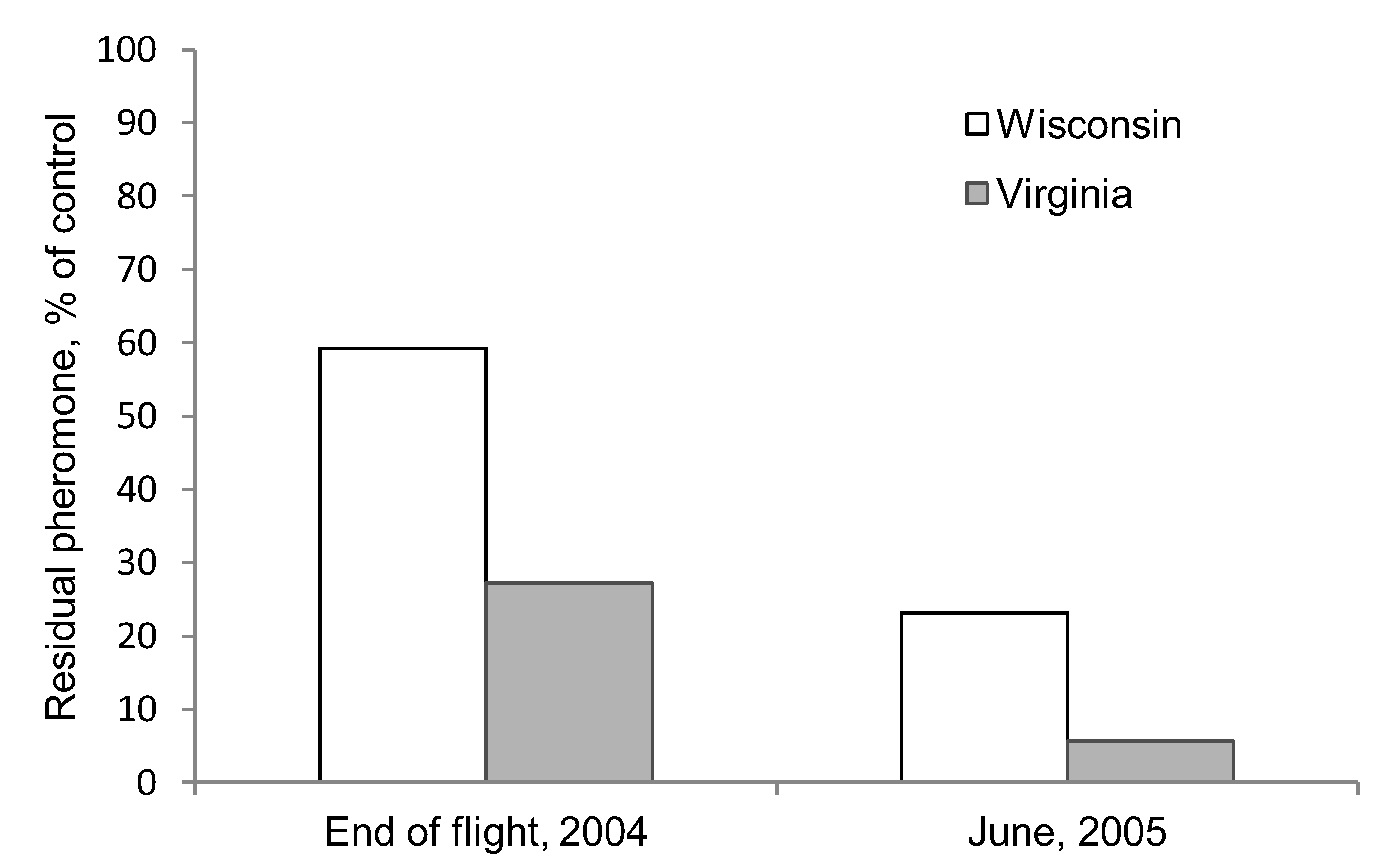
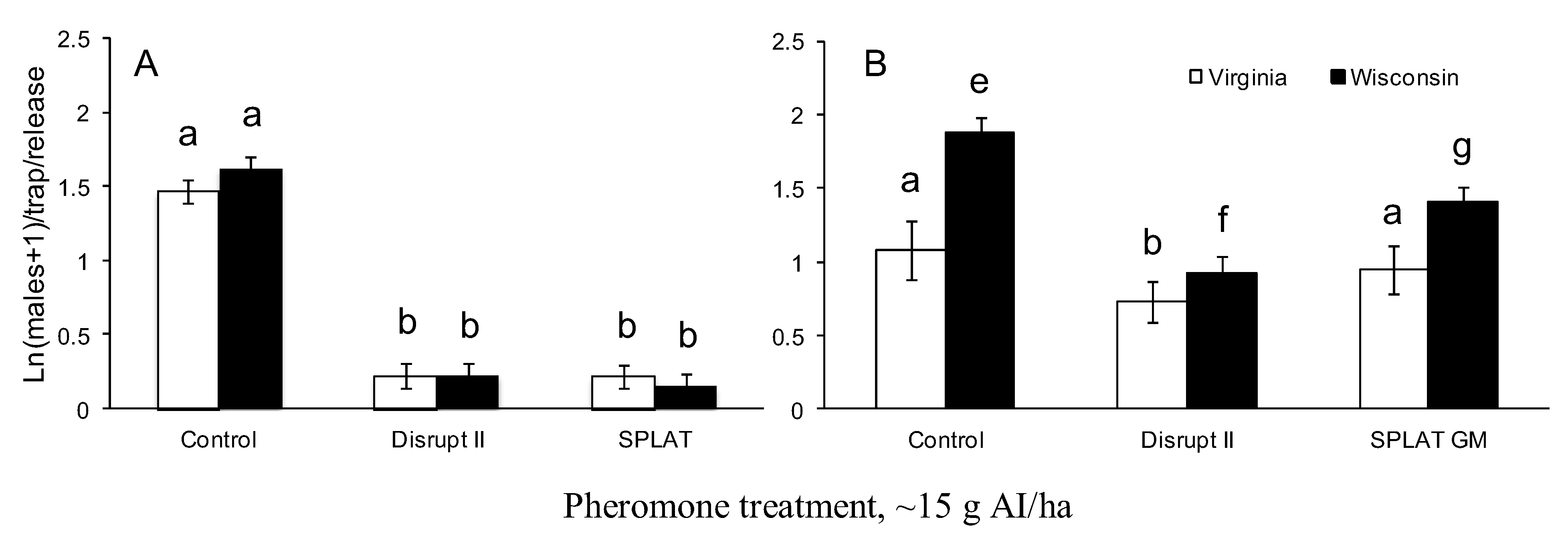
2.2. Pheromone Persistence Between Two Climate Regimes
3. Results
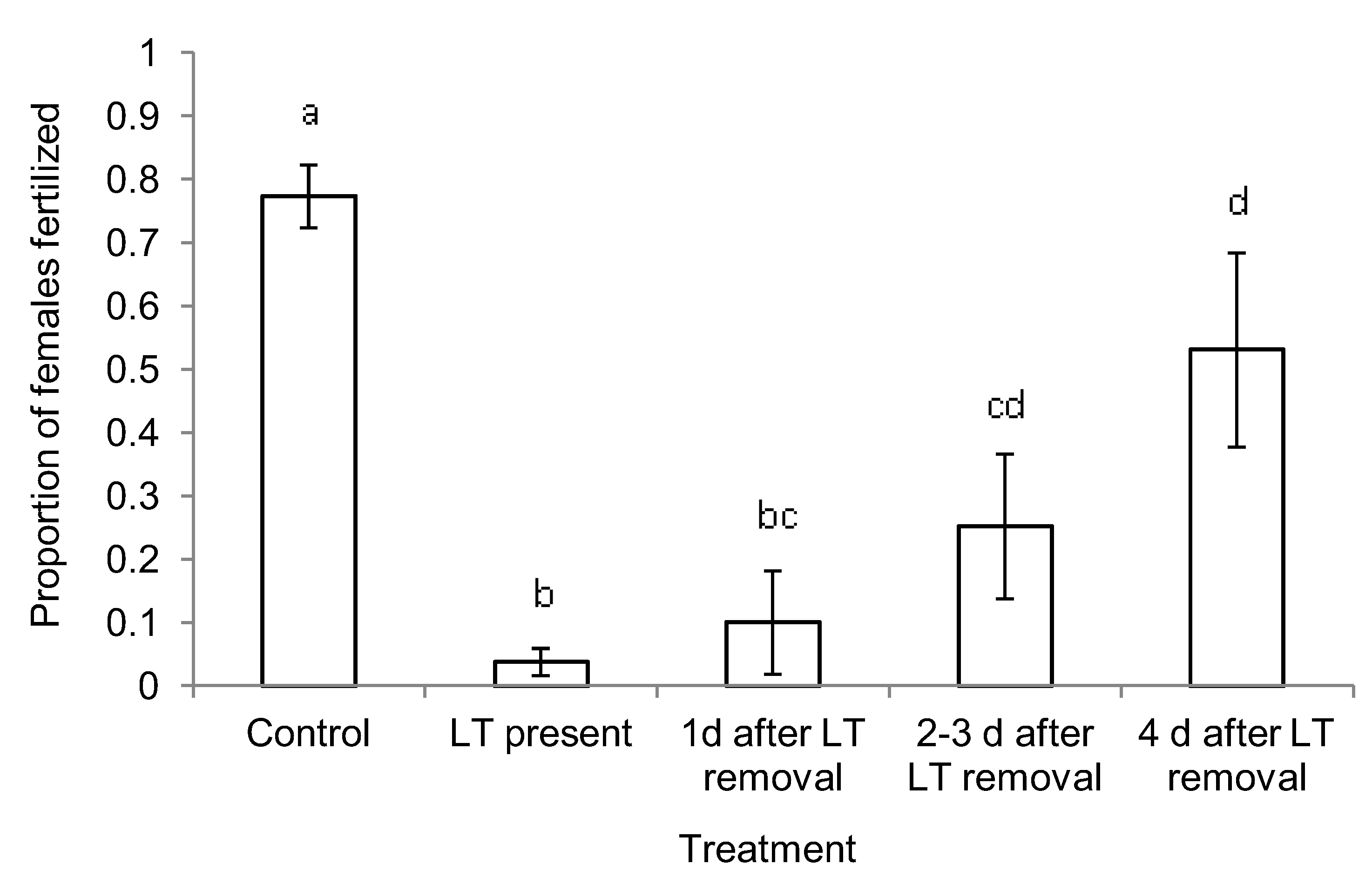
3.1. Mechanism of Short- and Long-term Persistence
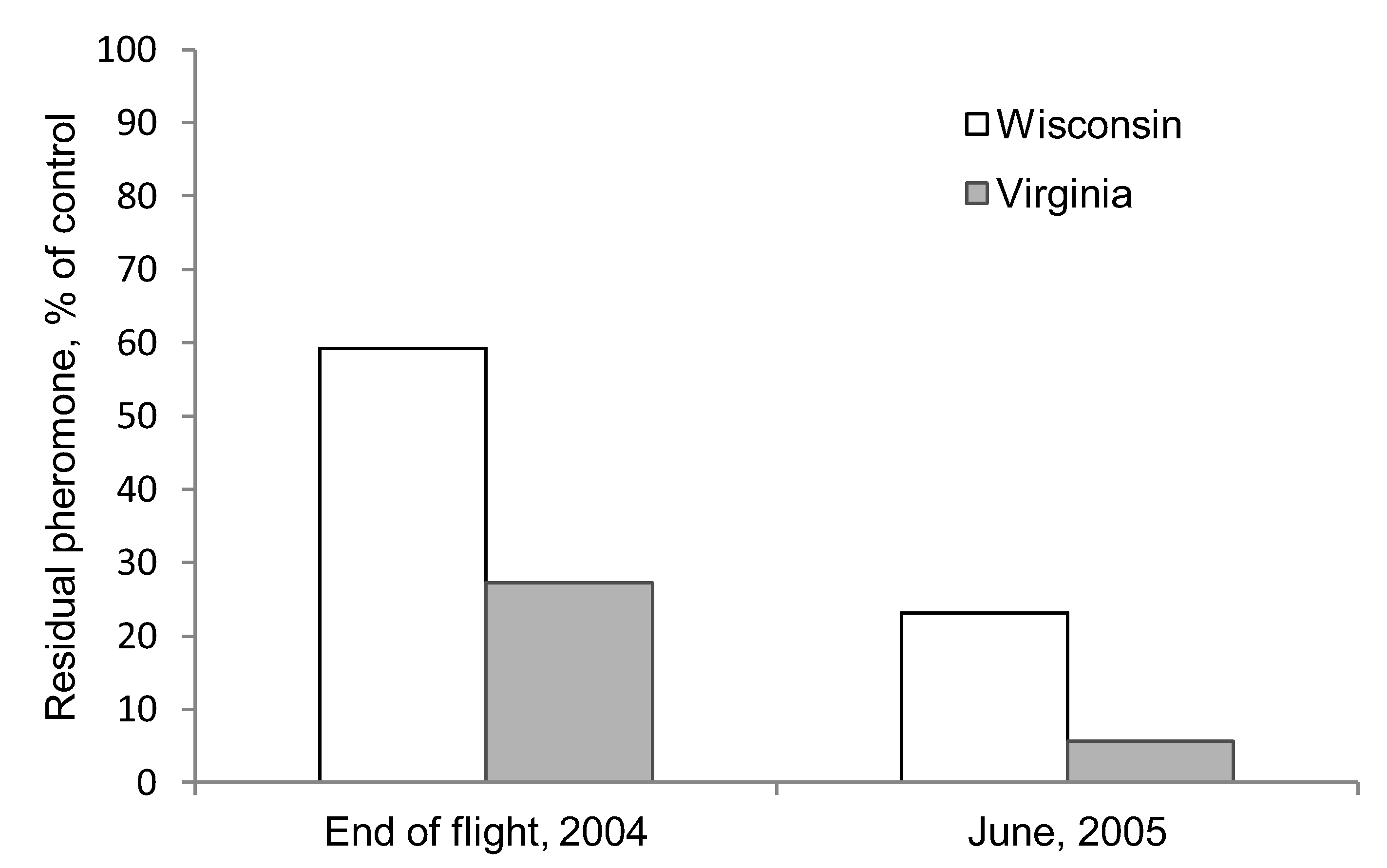
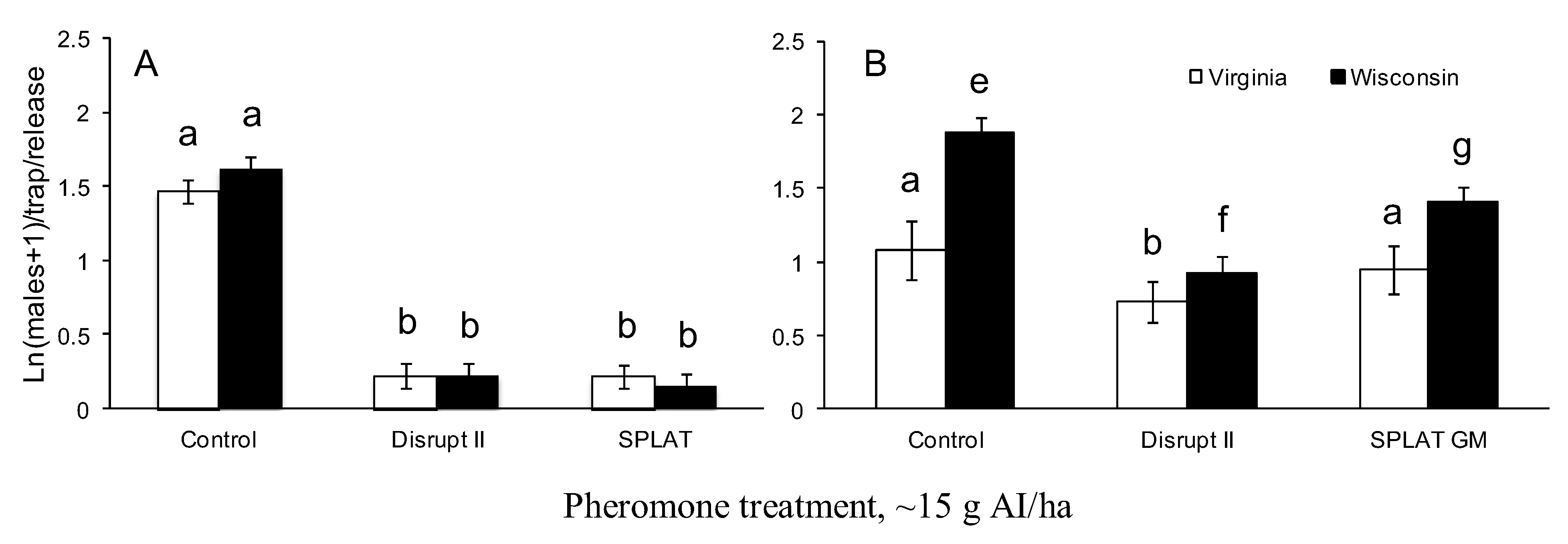
3.2. Pheromone Persistence between Two Climate Regimes
4. Discussion
Acknowledgements
References
- Carde, R.T.; Minks, A.K. Control of moth pests by mating disruption: successes and constraints. Annu. Rev. Entomol. 1995, 40, 559–585. [Google Scholar]
- El-Sayed, A.M.; Suckling, D.M.; Wearing, C.H.; Byers, J.A. Potential of mass trapping for long-term pest management and eradication of invasive species. J. Econ. Entomol. 2006, 99, 1550–1564. [Google Scholar]
- Suckling, D.M.; Tobin, P.C.; McCullough, D.G.; Herms, D.A. Combining tactics to exploit Allee effects for eradication of alien insect populations. J. Econ. Entomol. 2012, 105, 1–13. [Google Scholar]
- El-Sayed, A.M.; Mitchell, V.J.; Manning, L.-A.M.; Suckling, D.M. New sex pheromone blend for the lightbrown apple moth, Epiphyas postvittana. J. Chem. Ecol. 2011, 37, 640–646. [Google Scholar] [CrossRef]
- Miller, J.R.; Gut, L.J.; de Lame, F.M.; Stelinski, L.L. Differentiation of competitive vs. non-competitive mechanisms mediating disruption of moth sexual communication by point sources of sex pheromone (Part 2): Case studies. J. Chem. Ecol. 2006, 32, 2115–2143. [Google Scholar] [CrossRef]
- Yamanaka, T. Mating disruption or mass trapping? Numerical simulation analysis of a control strategy for lepidopteran pests. Popul. Ecol. 2007, 49, 75–86. [Google Scholar]
- Onufrieva, K.S.; Brewster, C.C.; Thorpe, K.W.; Sharov, A.A.; Leonard, D.S.; Reardon, R.C.; Mastro, V.C.; Sellers, P.; Roberts, E.A. Effects of the 3M (TM) MEC sprayable pheromone (R) formulation on gypsy moth mating success. J. Appl. Entomol. 2008, 132, 461–468. [Google Scholar]
- Wins-Purdy, A.H.; Judd, G.J.R.; Evenden, M.L. Mechanisms of pheromone communication disruption in choristoneura rosaceana exposed to microencapsulated (Z)-11-tetradecenyl acetate formulated with and without horticultural oil. J. Chem. Ecol. 2008, 34, 1096–1106. [Google Scholar]
- Thorpe, K.; Reardon, R.; Tcheslavskaia, K.; Leonard, D.; Mastro, V. A review of the use of mating disruption to manage gypsy moth, Lymantria dispar (L.); FHTET-2006–13. U.S. Department of Agriculture, Forest Service, Forest Health Technology Enterprise Team: Washington, DC, USA, 2006; Technical Report. [Google Scholar]
- Karg, G.; Suckling, D.M.; Bradley, S.J. Absorption and release of pheromone of Epiphyas postvittana (Lepidoptera: Tortricidae) by apple leaves. J. Chem. Ecol. 1994, 20, 1825–1841. [Google Scholar] [CrossRef]
- Suckling, D.M.; Karg, G.; Bradley, S.J. Apple foliage enhances mating disruption of light-brown apple moth. J. Chem. Ecol. 1996, 22, 325–341. [Google Scholar]
- Thorpe, K.W.; Tcheslavskaia, K.S.; Tobin, P.C.; Blackbum, L.M.; Leonard, D.S.; Roberts, E.A. Persistent effects of aerial applications of disparlure on gypsy moth: Trap catch and mating success. Entomol. Exp. Appl. 2007, 125, 223–229. [Google Scholar]
- Gut, L.J.; Stelinski, L.L.; Thomson, D.R.; Miller, J.R. Behaviour-modifying Chemicals: Prospects and Constraints in IPM; CABI Publishing: Wallingford, UK, 2004; pp. 73–121. [Google Scholar]
- Wall, C.; Sturgeon, D.M.; Greenway, A.R.; Perry, J.N. Contamination of vegetation with synthetic sex-attractant released from traps for the pea moth, Cydia nigricana. Entomol. Exp. Appl. 1981, 30, 111–115. [Google Scholar]
- Leonhardt, B.A.; Mastro, V.C.; Leonard, D.S.; McLane, W.; Reardon, R.C.; Thorpe, K.W. Control of low-density gypsy moth (Lepidoptera:Lymantriidae) populations by mating disruption with pheromone. J. Cheml. Ecol. 1996, 22, 1255–1272. [Google Scholar]
- Tobin, P.C.; Blackburn, L.M. Slow the Spread: A National Program to Manage the Gypsy Moth; USDA Forest Service: Newtown Square, PA, USA, 2007; Technical Report . [Google Scholar]
- Sharov, A.A.; Leonard, D.; Liebhold, A.M.; Roberts, E.A.; Dickerson, W. Slow the Spread": A national program to contain the gypsy moth. J. Forest. 2002, 100, 30–35. [Google Scholar]
- Tobin, P.C.; Bai, B.B.; Eggen, D.A.; Leonard, D.S. The ecology, geopolitics, and economics of managing the gypsy moth, Lymantria dispar (L.), in the United States. Int. J. Pest Manage. 2012, 58, 195–210. [Google Scholar] [CrossRef]
- Johnson, D.M.; Liebhold, A.M.; Bjornstad, O.N.; McManus, M.L. Circumpolar variation in periodicity and synchrony among gypsy moth populations. J. Anim. Ecol. 2005, 74, 882–892. [Google Scholar]
- Haynes, K.J.; Liebhold, A.M.; Johnson, D.M. Spatial analysis of harmonic oscillation of gypsy moth outbreak intensity. Oecologia 2009, 159, 249–256. [Google Scholar]
- Elkinton, J.S.; Liebhold, A.M. Population-Dynamics of Gypsy-Moth in North-America. Ann. Rev. Entomol. 1990, 35, 571–596. [Google Scholar]
- Epanchin-Niell, R.S.; Haight, R.G.; Berec, L.; Kean, J.M.; Liebhold, A.M. Optimal surveillance and eradication of invasive species in heterogeneous landscapes. Ecol. Lett. 2012, 15, 803–812. [Google Scholar]
- Liebhold, A.M.; Tobin, P.C. Growth of newly established alien populations: comparison of North American gypsy moth colonies with invasion theory. Popul. Ecol. 2006, 48, 253–262. [Google Scholar]
- Nation, J.L.; Foltz, J.L.; Dixon, W.N.; McAuslane, H.J. Evaluation of loss of (+)-disparlure from gypsy moth (Lepidoptera: Lymantriidae) pheromone dispenser tapes under field conditions in Florida. Fla. Entomol. 1993, 76, 584–589. [Google Scholar]
- Leonhardt, B.A.; Mastro, V.C.; Devilbiss, E.D. Evaluation of pheromone dispensers for use in gypsy moth detection (Lepidoptera: Lymantriidae). J. Entomol. Sci. 1992, 27, 280–284. [Google Scholar]
- Leonhardt, B.A.; Mastro, V.C.; Paszek, E.C.; Schwalbe, C.P.; Devilbiss, A.D. Dependence of gypsy moth (Lepidoptera: Lymantriidae) capture on pheromone release rate from laminate and other dispensers. J. Econ. Entomol. 1990, 83, 1977–1981. [Google Scholar]
- Bierl-Leonhardt, B.A.; DeVilbiss, E.D.; Plimmer, J.R. Rate of release of disparlure from laminated plastic dispensers. J. Econ. Entomol. 1979, 72, 319–321. [Google Scholar]
- Leonhardt, B.A.; Moreno, D.S. Evaluation of controlled release laminate dispensers for pheromones of several insect species. In Insect Pheromone Technology: Chemistry and Applications; Leonhardt, B.A., Beroza, M., Eds.; American Chemical Society: Washington, DC, USA, 1982; pp. 159–173, American Chemical Society Symposium Series 190. [Google Scholar]
- Tobin, P.C.; Zhang, A.; Onufrieva, K.; Leonard, D.S. Field evaluation of effect of temperature on release of disparlure from a pheromone-baited trapping system used to monitor gypsy moth (Lepidoptera: Lymantriidae). J. Econ. Entomol. 2011, 104, 1265–1271. [Google Scholar]
- Kolodny-Hirsch, D.M.; Webb, R.E.; Olsen, R.; Venables, L. Mating disruption of gypsy moth (Lepidoptera: Lymantriidae) following repeated ground application of racemic disparlure. J. Econ. Entomol. 1990, 83, 1972–1976. [Google Scholar]
- Thorpe, K.W.; Hickman, A.D.; Tcheslavskaia, K.S.; Leonard, D.S.; Roberts, E.A. Comparison of methods for deploying female gypsy moths to evaluate mating disruption treatments. Agr. Forest. Entomol. 2007, 9, 31–37. [Google Scholar]
- Tcheslavskaia, K.; Brewster, C.C.; Sharov, A.A. Mating success of gypsy moth (Lepidoptera : Lymantriidae) females in Southern Wisconsin. Great Lakes Entomol. 2002, 35, 1–7. [Google Scholar]
- Elkinton, J.S.; Carde, R.T. Effects of Intertrap Distance and Wind Direction on the Interaction of Gypsy-Moth (Lepidoptera, Lymantriidae) Pheromone-Baited Traps. Environ. Entomol. 1988, 17, 764–769. [Google Scholar]
- Thorpe, K.W.; Mastro, V.C.; Leonard, D.S.; Leonhardt, B.A.; McLane, W.; Reardon, R.C.; Talley, S.E. Comparative efficacy of two controlled-release gypsy moth mating disruption formulations. Entomol. Exp. App. 1999, 90, 267–277. [Google Scholar]
- Onufrieva, K.S.; Thorpe, K.W.; Hickman, A.D.; Tobin, P.C.; Leonard, D.S.; Roberts, E.A. Effects of SPLAT® GM sprayable pheromone formulation on gypsy moth mating success. Entomol. Exp. App. 2010, 136, 109–115. [Google Scholar]
- Karg, G.; Sauer, A.E.; Koch, U.T. The Influence of Plants on the Development of Pheromone Atmospheres Measured by EAG Method. In Proceedings of 18th Gottingen Neurobiology Conference, Thieme Verlag, Stuttgart, Germany, 4–6 June 1990.
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Onufrieva, K.S.; Thorpe, K.W.; Hickman, A.D.; Leonard, D.S.; Roberts, E.A.; Tobin, P.C. Persistence of the Gypsy Moth Pheromone, Disparlure, in the Environment in Various Climates. Insects 2013, 4, 104-116. https://doi.org/10.3390/insects4010104
Onufrieva KS, Thorpe KW, Hickman AD, Leonard DS, Roberts EA, Tobin PC. Persistence of the Gypsy Moth Pheromone, Disparlure, in the Environment in Various Climates. Insects. 2013; 4(1):104-116. https://doi.org/10.3390/insects4010104
Chicago/Turabian StyleOnufrieva, Ksenia S., Kevin W. Thorpe, Andrea D. Hickman, Donna S. Leonard, E. Anderson Roberts, and Patrick C. Tobin. 2013. "Persistence of the Gypsy Moth Pheromone, Disparlure, in the Environment in Various Climates" Insects 4, no. 1: 104-116. https://doi.org/10.3390/insects4010104