1. Introduction
Parasitic Hymenoptera are extremely species-rich and of wide significance in terrestrial ecosystems, because they are a major group of insects with a parasitoid lifestyle [
1]. The fact that they are particularly sensitive to environmental disturbance makes them good indicators of diversity and environmental stability [
1,
2] and allows the establishment of a pattern of ecosystem conservation [
3,
4,
5].
Within the parasitic Hymenoptera, the family of Braconidae is the second largest family, comprising about 40,000 species, which are distributed throughout the world in several different habitats [
6]. They are considered essential for the maintenance of the balance of the communities that include them [
7], due to their role in controlling the caterpillar populations [
8,
9,
10,
11]. These wasps are primary parasitoids of the immature stages of Lepidoptera, Diptera and Coleoptera [
6,
12] and the economic importance of braconid species resides in their potential for the biological control of insect pests [
8,
13,
14].
Within the Braconidae, the subfamily Microgastrinae with over 2,000 species described and an estimated global diversity of 5,000–10,000 [
15], is one of the most important groups of parasitoids in terms of both species richness and economic importance [
16]. This fact has caused the Microgastrinae to be one of the most studied parasitic wasps by DNA-barcode in recent years [
16,
17,
18,
19].
These wasps are all koinobiont endoparasitoids of Lepidopteran larvae and the vast majority of known hosts belong to the
Ditrysia-group [
2]. More than 100 species of Microgastrinae have been used in the biological control of Lepidoptera pests and this total is likely to rise [
20].
Despite considerable knowledge concerning the ecology of some species, the same is not true at the family level with the exception of some recent studies [
21,
22]. In the Iberian Peninsula (Andorra, Spain and Portugal), Braconidae communities have been insufficiently analyzed [
4,
23,
24,
25,
26,
27,
28], with the exception of those in the Pyrenees [
29] or in Navarra [
30,
31].
Within this context, this study analyzed alpha, beta and gamma diversity of Microgastrinae in the Artikutza Forest (Navarra) located in the western Pyrenees and having an enormous ecological value. This area includes two adjacent types of forest delimited by a stream: mixed pine forest and beech forest. Data on the phenology of the subfamily and its relationship with environmental and climatic conditions were also studied.
3. Results and Discussion
During the sampling period, 3,534 specimens of the Braconidae family were captured. Of these, 524 belonged to the Microgastrinae subfamily (14.82%), 364 of which were males and 160 were females. Because only females could be identified at the species level, males were not included in the analyses. From the total of 160 females, 154 could be identified because the remaining specimens lacked some structure that was essential for their identification.
A total of 27 species was captured, belonging to nine genera: Choeras dorsalis (Spinola 1808), Choeras hedymeles (Nixon 1973), Choeras parasitellae (Bounché 1834), Cotesia ancilla (Nixon 1974), Cotesia chares (Nixon 1974), Cotesia jucundus (Marshall 1885), Cotesia sp1, Dolichogenidea celsus (Papp 1975), Dolichogenidea laevigatoides (Nixon 1972), Dolichogenidea laevigatus (Ratzeburg 1848), Dolichogenidea sp1, Dolichogenidea sp2, Dolichogenidea varifemur (Abdinbekova 1969), Glyptapanteles aliphera (Nixon 1973), Glyptapanteles fulvipes (Haliday 1834), Glyptapanteles mygdonia (Nixon 1973), Glyptapanteles porthetriae (Muesebeck 1928), Glyptapanteles vitripennis (Curtis 1830), Microgaster sp1, Microplitis marshalii Kokijev 1898, Paroplitis sp1, Paroplitis wesmaeli (Ruthe 1860), Pholetesor circumscriptus (Nees 1834), Pholetesor sp1, Pholetesor sp2, Protapanteles anchisiades (Nixon 1973) and Protapanteles sp1.
The number of genera and species differed between each habitat and trap. In the mixed forest, 23 species were collected (15 in M-1, 8 in M-2 and 10 in M-3) whereas 11 species were captured in the beech forest (7 in H-1, 7 in H-2 and 7 in H-3) (
Table 2). These differences are because the mixed forest represents an area with a richer diversity of plants and consequently also of host species. Individually, M-1 was the trap with the largest number of species as it was located on the edge of a clearing which was windier than other locations.
The genus analysis showed that the genus
Choeras is the most abundant in this study, with 64 specimens, followed by the genera
Paroplitis (45) and
Glyptapanteles (35) which contrasts with the results from other studies of Braconidae in the Iberian Peninsula [
55]. When these results are checked with the results obtained by Falcó-Garí
et al. in Andorra [
29], it is possible to see that
Glyptapanteles remains very abundant. However, only one specimen has been captured of
Microplitis.
The species analysis showed that
Choeras hedymeles was the most common species with 59 specimens (38.31%) followed by
Glyptapanteles vitripennis with 21 and
Dolichogenidea sp1 with 15 (13.63% and 9.74%, respectively). However, analysis of the number of captures showed that 102 specimens were collected in the mixed forest habitat (37 in M-1, 28 in M-2 and 37 in M-3) and 52 in the beech forest (18 in H-1, 18 in H-2 and 16 in H-3) (
Table 2). Note the absence of the genus
Apanteles, genus characterized by its ubiquity and dominance [
56]. However, checking the data capture of Falcó-Garí
et al. [
29], in Andorra (Pyrenees) also note that only 15 specimens were captured of a total of 494 (3.03%). Both facts show that
Apanteles is not abundant in this area.
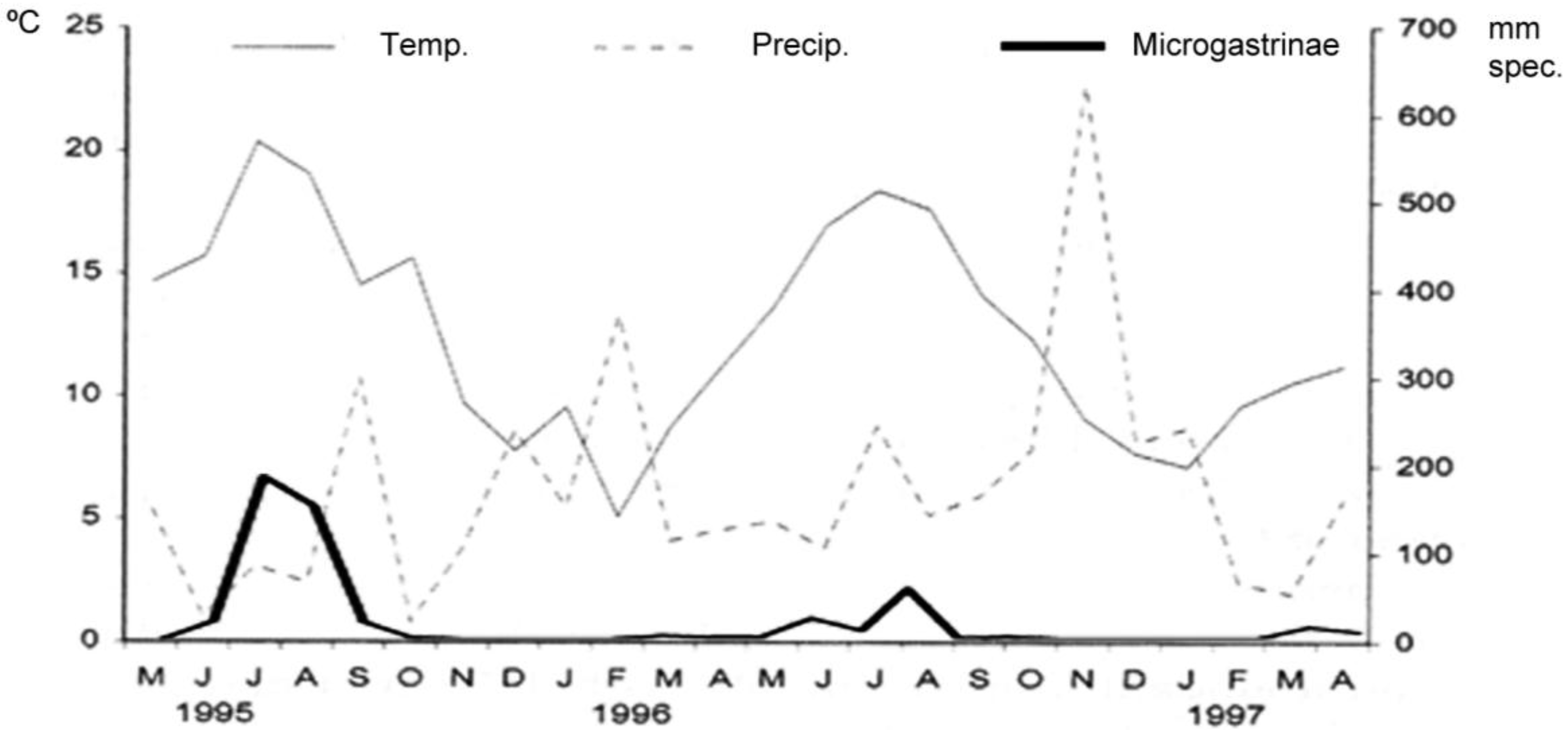
The phenology of the Microgastrinae (
Figure 1) established relationships between the abundance of Microgastrinae and climatic conditions. The capture of Microgastrinae was higher during the summer months when temperatures are moderate with average of 15–20 °C. In contrast, increased rainfall during the winter months caused a decrease in temperature and in Microgastrinae abundance.
Table 2.
Distribution of Microgastrinae specimens per habitat.
Table 2.
Distribution of Microgastrinae specimens per habitat.
| Species | Trap number | Total Specimens |
|---|
| M-1 | M-2 | M-3 | H-1 | H-2 | H-3 | ΣM | ΣH | Total |
|---|
| Choeras dorsalis | 1 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 1 |
| Choeras hedymeles | 5 | 10 | 18 | 10 | 10 | 6 | 33 | 26 | 59 |
| Choeras parasitellae | 3 | 0 | 0 | 0 | 0 | 1 | 3 | 1 | 4 |
| Cotesia ancilla | 1 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 1 |
| Cotesia chares | 0 | 1 | 0 | 0 | 0 | 0 | 1 | 0 | 1 |
| Cotesia jucundus | 0 | 1 | 0 | 0 | 0 | 0 | 1 | 0 | 1 |
| Cotesia sp1 | 4 | 0 | 0 | 0 | 0 | 0 | 4 | 0 | 4 |
| Dolichogenidea celsus | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 1 |
| Dolichogenidea laevigatoides | 0 | 0 | 0 | 2 | 0 | 0 | 0 | 2 | 2 |
| Dolichogenidea laevigatus | 0 | 0 | 0 | 1 | 1 | 0 | 0 | 2 | 2 |
| Dolichogenidea sp1 | 3 | 3 | 5 | 2 | 2 | 0 | 11 | 4 | 15 |
| Dolichogenidea sp2 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 1 |
| Dolichogenidea varifemur | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 2 | 2 |
| Glyptapanteles aliphera | 1 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 1 |
| Glyptapanteles fulvipes | 2 | 0 | 0 | 0 | 0 | 0 | 2 | 0 | 2 |
| Glyptapanteles mygdonia | 0 | 1 | 3 | 1 | 0 | 4 | 4 | 5 | 9 |
| Glyptapanteles porthetriae | 2 | 0 | 0 | 0 | 0 | 0 | 2 | 0 | 2 |
| Glyptapanteles vitripennis | 4 | 9 | 4 | 1 | 2 | 1 | 17 | 4 | 21 |
| Microgaster sp1 | 1 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 1 |
| Microplitis marshalii | 1 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 1 |
| Paroplitis macrocephalus | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 1 |
| Paroplitis wesmaeli | 1 | 0 | 0 | 0 | 1 | 2 | 1 | 3 | 4 |
| Pholetesor circumscriptus | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 1 |
| Pholetesor sp1 | 3 | 0 | 1 | 0 | 1 | 1 | 4 | 2 | 6 |
| Pholetesor sp2 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 1 |
| Protapanteles anchisiades | 5 | 2 | 2 | 0 | 0 | 0 | 9 | 0 | 9 |
| Protapanteles sp1 | 0 | 1 | 0 | 0 | 0 | 0 | 1 | 0 | 1 |
| Total specimens | 37 | 28 | 37 | 18 | 18 | 16 | 102 | 52 | 154 |
Figure 1.
Relationship between climatic conditions and abundance of Microgastrinae.
Figure 1.
Relationship between climatic conditions and abundance of Microgastrinae.
3.1. Alpha Diversity
The mixed forest had a higher species richness with a D
Mg = 4.76 than the beech forest with a value of 2.53. At the trap level, M-1 (D
Mg = 3.877) had the highest richness, followed by H-3 (2.164) and H‑1 and H-2 (1.750 and 1.650, respectively) (
Table 3). In addition to alpha diversity, the Shannon–Wiener and the Simpson indexes are shown in
Table 3. The Shannon index for mixed forest (2.346) and beech forest (2.017) suggested a similar trend in the distribution of dominant genera and discrepancies were due to different numbers of rare genera (those represented by few specimens). This trend was also supported by the Simpson index (0.159 in mixed forest and 0.281 in beech forest). At the trap level, the Shannon-Wiener index values were higher in M-1, M-3 and H-3 (2.537, 1.711 and 1.667, respectively) whereas for the Simpson index, the ranking was H-1 and H-2 (both 0.345) M-2 (0.525) and M-3 (0.280).
Table 3.
Diversity and abundance values of the collected Microgastrinae species.
Table 3.
Diversity and abundance values of the collected Microgastrinae species.
| Data | Trap number | Total |
|---|
| M-1 | M-2 | M-3 | H-1 | H-2 | H-3 | ΣM | ΣH |
|---|
| Species | 15 | 8 | 10 | 7 | 7 | 7 | 23 | 11 |
| Specimens | 37 | 28 | 37 | 18 | 18 | 16 | 102 | 52 |
| Shannon I. | 2.537 | 1.636 | 1.711 | 1.453 | 1.453 | 1.667 | 2.346 | 2.174 |
| Simpson I. | 0.090 | 0.252 | 0.280 | 0.345 | 0.345 | 0.234 | 0.159 | 0.281 |
| Margalef I. | 3.877 | 2.101 | 2.492 | 2.076 | 2.076 | 2.164 | 4.760 | 2.530 |
3.2. Beta Diversity
Beta diversity was calculated using values between the different areas with the Jaccard and complementarity indices (
Table 4). Firstly, the Jaccard index showed a degree of dissimilarity at the species level between the two habitats (0.259). At the trap level, H-2 and H-3 both showed the highest similarity with each other (0.556) and M-1 and H-1 were the least similar traps (0.158). Secondly, the complementarity index between mixed and beech forest (0.741) indicated that the species composition similarity between both communities was low. At the trap level, M-1 and H-1 shows the highest complementarity (0.842) whereas the comparison between H-2 and H-3 showed the least complementarity, with a value of 0.444.
Table 4.
Jaccard index and Complementarity Values.
Table 4.
Jaccard index and Complementarity Values.
| | M-1 | M-2 | M-3 | H-1 | H-2 | H-3 | |
| M-1 | | 0.211 | 0.250 | 0.158 | 0.294 | 0.294 |
| M-2 | 0.789 | | 0.385 | 0.364 | 0.250 | 0.250 |
| M-3 | 0.750 | 0.615 | | 0.308 | 0.308 | 0.308 | Jaccard I. |
| H-1 | 0.842 | 0.636 | 0.818 | | 0.400 | 0.273 | |
| H-2 | 0.706 | 0.750 | 0.818 | 0.600 | | 0.556 |
| H-3 | 0.706 | 0.750 | 0.818 | 0.545 | 0.444 | |
| | Complementarity | |
3.3. Gamma Diversity
Gamma diversity using the Lande index reached a value of 37 and can be decomposed into alpha and beta diversity. Alpha diversity contributed 45.95% towards the gamma index and beta diversity, the remaining 54.05%. These results suggest that a greater degree of change in species composition than differences in species richness exists between mixed and beech communities. This fact can be explained by the specificity of habitat in both potential hosts and their parasitoids.
4. Conclusions
To sum up, on checking with studies carried out in Artikutza about Braconidae subfamilies, it has been shown that Alysiinae was the most abundant subfamily captured with approximately 64.23%, followed by Microgastrinae with 14.82%, while Opiinae has only 2.97%. The information about the Braconidae abundance and alpha diversity is very interesting due to the specific relationships that these parasitic wasps have with their host and with the host plants. This information about trophic relationships could be used indirectly to further knowledge concerning the biodiversity appearing in similar areas.
On the other hand, it is possible to consider that Microgastrinae is one of the most abundant subfamilies in both the mixed forest and in the beech forest with 524 collected specimens, mainly collected during the months of moderate temperatures (15–20 °C), July and August. In turn, as can be expected, the mixed forest shows a higher Microgastrinae abundance than in the beech forest due to the greater variety of potential host plants of Lepidoptera. However, in contrast to the results from other studies of Braconidae in the Iberian Peninsula, Choeras is the most abundant genus with the unexpected absence of Apanteles.
In addition, on analyzing the community structure, it is possible to see that Microgastrinae community of mixed and beech forest are composed of a few abundant species and a large number of rare species. The observed pattern in beta diversity shows that the two sampled sites have a higher complementarity with different species composition, which belongs to areas with an important ecological value.
Finally, we conclude that, although this study was conducted to determine the diversity and community structure of Microgastrinae, further studies of Braconidae are recommended in different areas together with DNA-barcode studies to increase the knowledge of this large subfamily that remains mostly unknown.


{kind=link}