
Flood Stress as a Technique to Assess Preventive Insecticide and Fungicide Treatments for Protecting Trees against Ambrosia Beetles


Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Trees and Imposing Flood-Stress
2.2. Effects of Preventive Insecticide Treatment on Preference and Colonization Success
2.3. Effects of Preventive Fungicide Treatment on Preference and Colonization Success
2.4. Statistics
3. Results
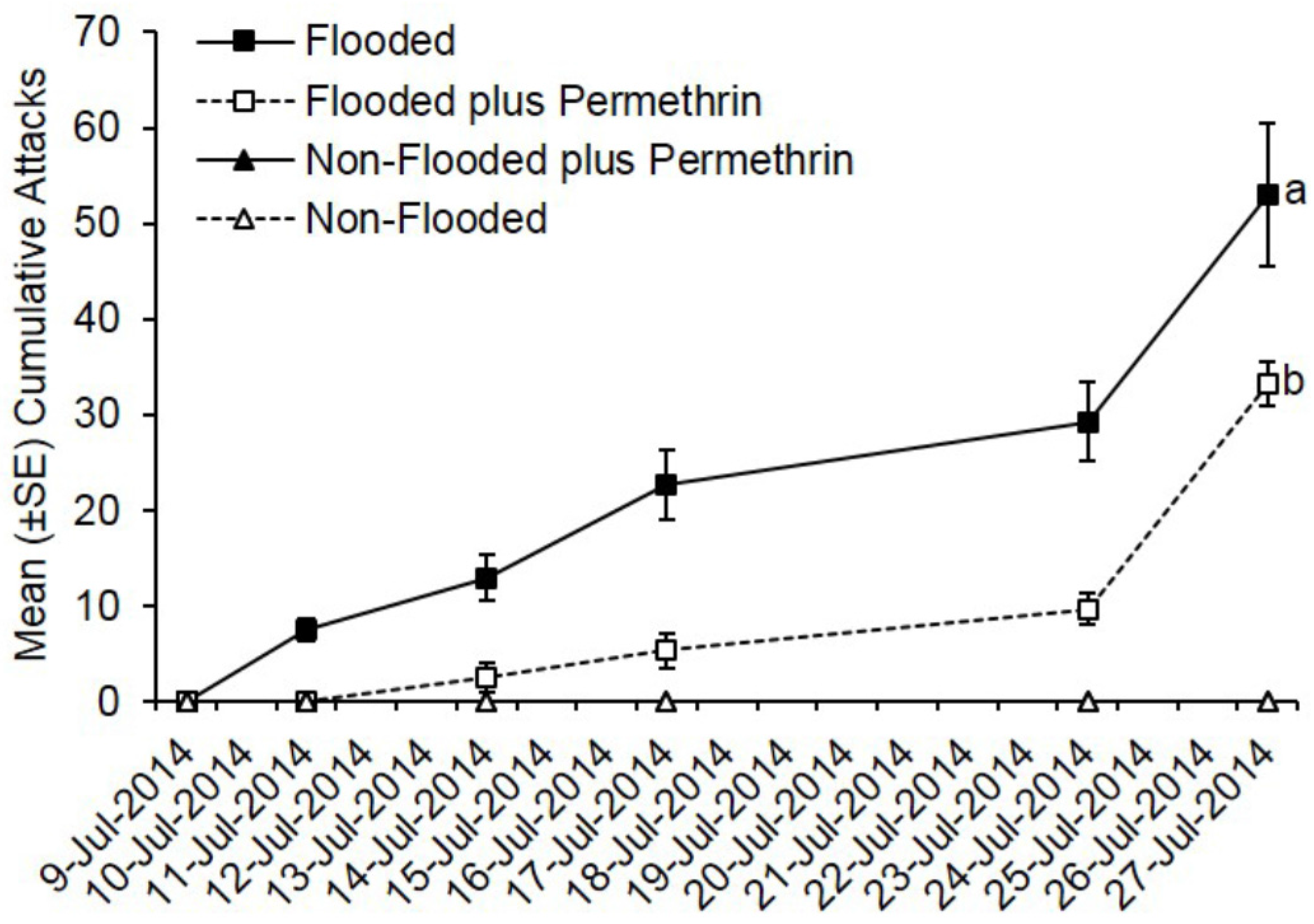
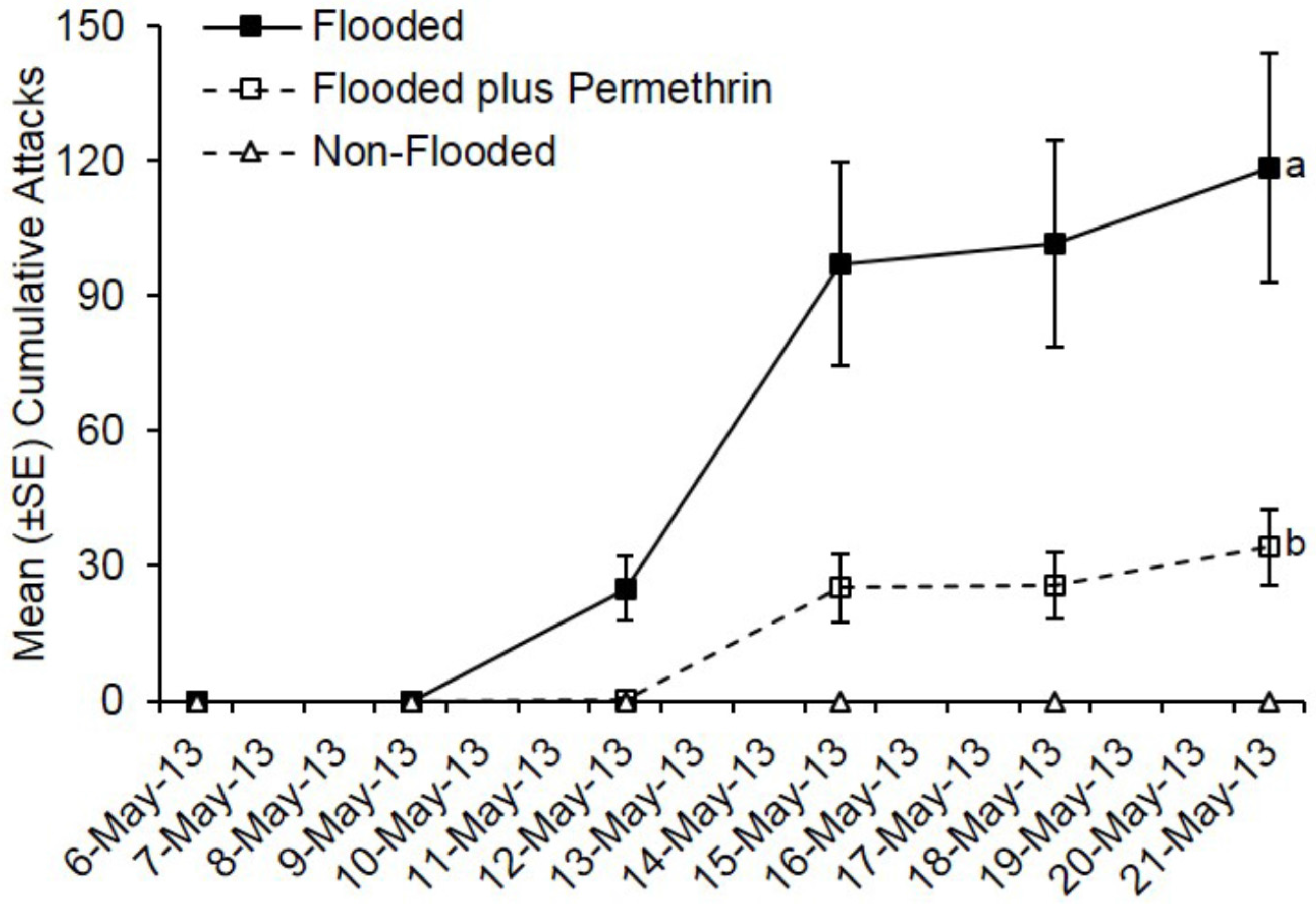
3.1. Preventive Insecticide Effects on Preference and Colonization Success
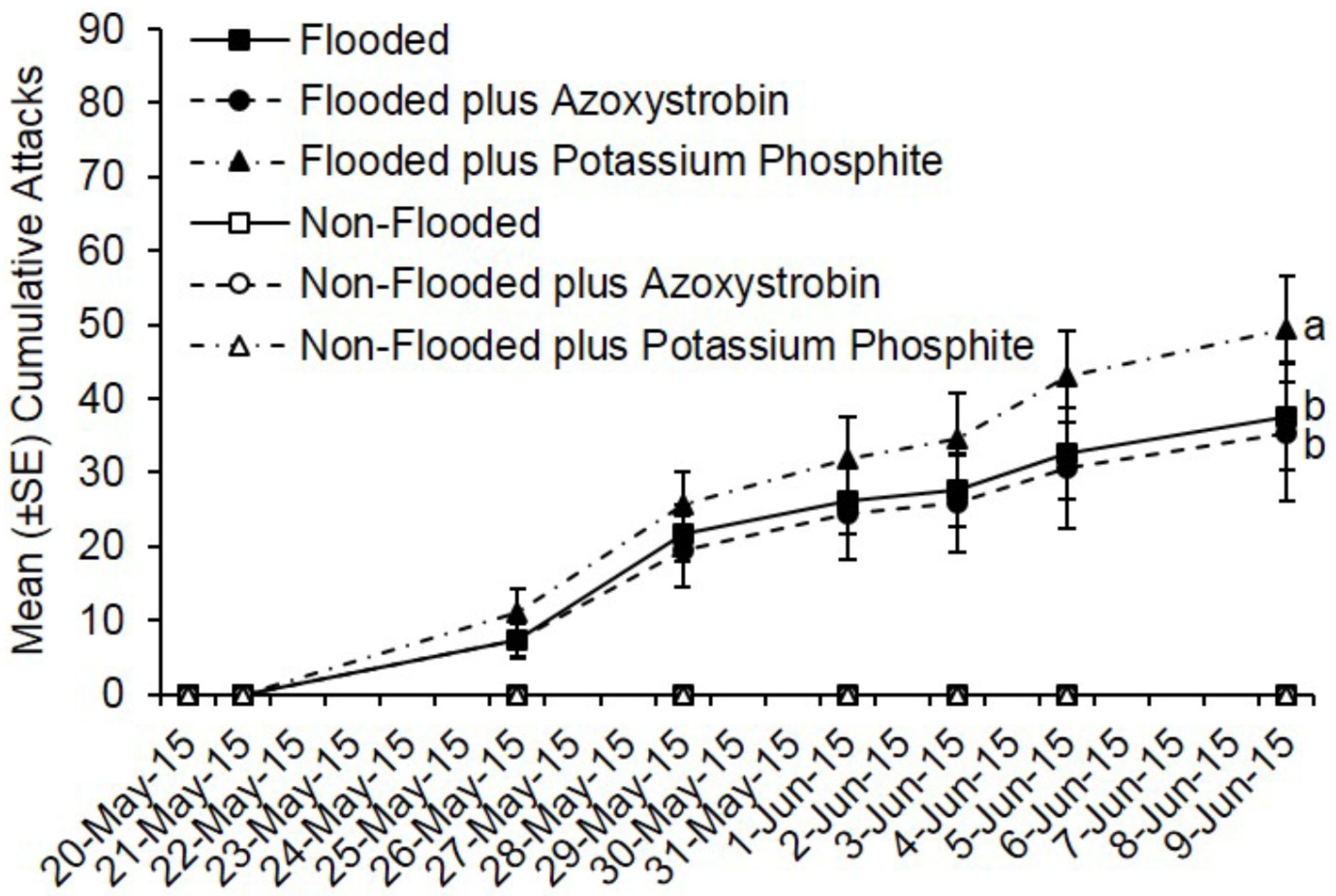
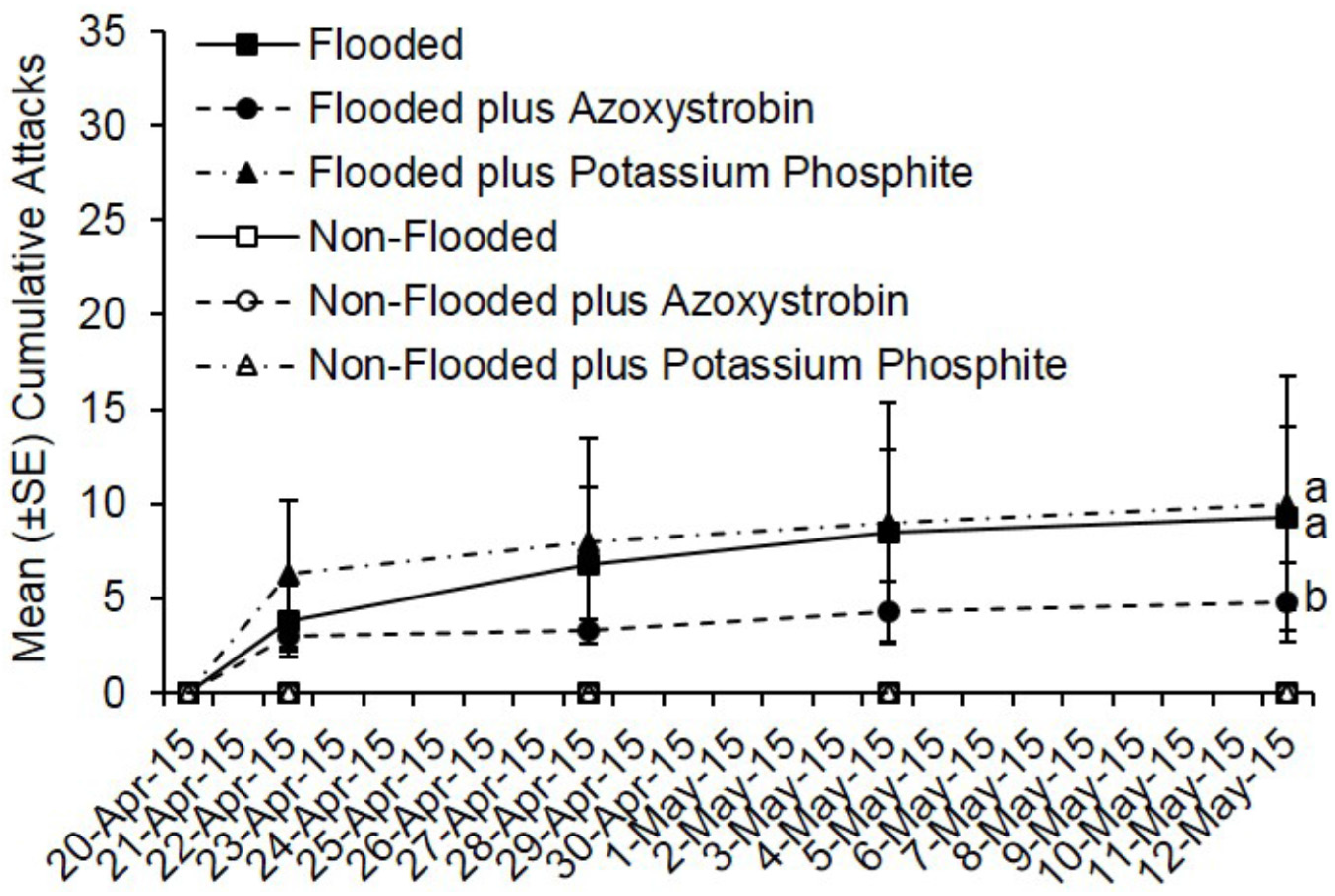
3.2. Preventive Fungicide Effects on Preference and Colonization Success
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Frank, S.D.; Klingeman, W.E.; White, S.A.; Fulcher, A. Biology, injury, and management of maple tree pests in nurseries and urban landscapes. J. Int. Pest Manag. 2013, 4, B1–B14. [Google Scholar] [CrossRef]
- Agnello, A.; Breth, D.; Tee, E.; Cox, K.; Warren, H.R. Ambrosia beetle—An emergent apple pest. N. Y. Fruit Q. 2015, 23, 25–28. [Google Scholar]
- Ranger, C.M.; Reding, M.E.; Schultz, P.B.; Oliver, J.B.; Frank, S.D.; Addesso, K.M.; Chong, J.H.; Sampson, B.; Werle, C.; Gill, S.; et al. Biology, ecology, and management of nonnative ambrosia beetles (Coleoptera: Curculionidae: Scolytinae) in ornamental plant nurseries. J. Integr. Pest Manag. 2016, 7, 1–23. [Google Scholar]
- (CABI) Commonwealth Agricultural Bureaux International Invasive Species Compendium. Xylosandrus compactus (Shot-Hole Borer); CABI Head Office: Wallingford, UK, 2015; Available online: http://www.cabi.org/isc/datasheet/57234 (accessed on 27 May 2016).
- Schedl, K.E. Scolytidae und Platypodidae Afrikans. Band 2. Familie Scolytidae (Fortsetzung). Revista de Entomologia de Moçambique 1963, 5, 1–593. [Google Scholar]
- Weber, B.C.; McPherson, J.E. World list of host plants of Xylosandrus germanus (Blandford) (Coleoptera: Scolytidae). Coleop. Bull. 1983, 37, 114–134. [Google Scholar]
- Ngoan, N.D.; Wilkinson, R.C.; Short, D.E.; Moses, C.S.; Mangold, J.R. Biology of an introduced ambrosia beetle, Xylosandrus compactus, in Florida. Ann. Entomol. Soc. Am. 1976, 69, 872–876. [Google Scholar] [CrossRef]
- Harrington, T.C.; McNew, D.; Mayers, C.; Fraedrich, S.W.; Reed, S.E. Ambrosiella roeperi sp. nov. is the mycangial symbiont of the granulate ambrosia beetle, Xylosandrus crassiusculus. Mycologia 2014, 106, 835–845. [Google Scholar] [CrossRef] [PubMed]
- Mayers, C.G.; McNew, D.L.; Harrington, T.C.; Roeper, R.A.; Fraedrich, S.W.; Biedermann, P.H.; Castrillo, L.A.; Reed, S.E. Three genera in the Ceratocystidaceae are the respective symbionts of three independent lineages of ambrosia beetles with large, complex mycangia. Fungal Biol. 2015, 119, 1075–1092. [Google Scholar] [CrossRef] [PubMed]
- French, J.R.; Roeper, R.A. Interactions of the ambrosia beetle, Xyleborus dispar (Coleoptera: Scolytidae), with its symbiotic fungus Ambrosiella hartigii (Fungi: Imperfecti). Can. Entomol. 1972, 104, 1635–1641. [Google Scholar] [CrossRef]
- Weber, B.C.; McPherson, J.E. Life history of the ambrosia beetle Xylosandrus germanus (Coleoptera: Scolytidae). Ann. Entomol. Soc. Am. 1983, 76, 455–462. [Google Scholar] [CrossRef]
- Weber, B.C.; McPherson, J.E. The ambrosia fungus of Xylosandrus germanus (Coleoptera: Scolytidae). Can. Entomol. 1984, 116, 281–283. [Google Scholar] [CrossRef]
- Kinuura, H. Symbiotic fungi associated with ambrosia beetles. Jpn. Agric. Res. Quart. 1995, 29, 57–63. [Google Scholar]
- Hulcr, J.; Rountree, N.R.; Diamond, S.E.; Stelinski, L.L.; Fierer, N.; Dunn, R.R. Mycangia of ambrosia beetles host communities of bacteria. Microb. Ecol. 2012, 64, 784–793. [Google Scholar] [CrossRef] [PubMed]
- Ploetz, R.C.; Hulcr, J.; Wingfield, M.J.; de Beer, Z.W. Destructive tree diseases associated with ambrosia and bark beetles: Black swan events in tree pathology? Plant Dis. 2013, 97, 856–872. [Google Scholar] [CrossRef]
- Carrillo, D.; Duncan, R.E.; Ploetz, J.N.; Campbell, A.F.; Ploetz, R.C.; Peña, J.E. Lateral transfer of a phytopathogenic symbiont among native and exotic ambrosia beetles. Plant Pathol. 2014, 63, 54–62. [Google Scholar] [CrossRef]
- Buchanan, W.D. Experiments with an ambrosia beetle, Xylosandrus germanus (Blfd.). J. Econ. Entomol. 1941, 34, 367–369. [Google Scholar] [CrossRef]
- Ranger, C.M.; Reding, M.E.; Persad, A.B.; Herms, D.A. Ability of stress-related volatiles to attract and induce attacks by Xylosandrus germanus and other ambrosia beetles (Coleoptera: Curculionidae, Scolytinae). Agric. For. Entomol. 2010, 12, 177–185. [Google Scholar] [CrossRef]
- Reding, M.E.; Oliver, J.; Schultz, P.; Ranger, C.M.; Youssef, N. Ethanol injection of ornamental trees facilitates testing insecticide efficacy against ambrosia beetles (Coleoptera: Curculionidae: Scolytinae). J. Econ. Entomol. 2013, 106, 289–298. [Google Scholar] [CrossRef] [PubMed]
- Frank, S.D.; Sadof, C.S. Reducing insecticide volume and nontarget effects of ambrosia beetle management in nurseries. J. Econ. Entomol. 2011, 104, 1960–1968. [Google Scholar] [CrossRef] [PubMed]
- Ranger, C.M.; Reding, M.E.; Oliver, J.B.; Schultz, P.B.; Moyseenko, J.J.; Youssef, N. Comparative efficacy of botanical formulations for managing ambrosia beetles and their corresponding mass spectral characterization. J. Econ. Entomol. 2011, 104, 1665–1674. [Google Scholar] [CrossRef] [PubMed]
- Ranger, C.M.; Tobin, P.C.; Reding, M.E.; Bray, A.M.; Oliver, J.B.; Schultz, P.B.; Frank, S.D.; Persad, A.B. Interruption of semiochemical-based attraction of ambrosia beetles to ethanol-baited traps and ethanol-injected trap trees by verbenone. Environ. Entomol. 2013, 42, 539–547. [Google Scholar] [CrossRef] [PubMed]
- Ranger, C.M.; Reding, M.E.; Schultz, P.; Oliver, J. Influence of flood-stress on ambrosia beetle (Coleoptera: Curculionidae, Scolytinae) host-selection and implications for their management in a changing climate. Agric. For. Entomol. 2013, 15, 56–64. [Google Scholar] [CrossRef]
- Ranger, C.M.; Schultz, P.B.; Frank, S.D.; Chong, J.H.; Reding, M.E. Non-native ambrosia beetles as opportunistic exploiters of living but weakened trees. PLoS ONE 2015, 10, e0131496. [Google Scholar] [CrossRef] [PubMed]
- Ranger, C.M.; Reding, M.E.; Schultz, P.B.; Oliver, J.B. Ambrosia beetle (Coleoptera: Curculionidae) responses to volatile emissions associated with ethanol-injected Magnolia virginiana. Environ. Entomol. 2012, 41, 636–647. [Google Scholar] [CrossRef] [PubMed]
- Strom, B.L.; Roton, L.M. A small-bolt method for screening tree protectants against bark beetles (Coleoptera: Curculionidae). J. Entomol. Sci. 2009, 44, 1–12. [Google Scholar]
- Dejonckheere, W.; Verstraeten, R.; Steurbaut, W.; Melkebeke, G.; Kips, R.H. Permethrin and deltamethrin residues on lettuce. Pestic. Sci. 1982, 13, 351–356. [Google Scholar] [CrossRef]
- MacKenzie, S.J.; Mertely, J.C.; Peres, N.A. Curative and protectant activity of fungicides for control of crown rot of strawberry caused by Colletotrichum gloeosporioides. Plant Dis. 2009, 93, 815–820. [Google Scholar] [CrossRef]
- Mizell, R.F.; Riddle, T.C. Evaluation of insecticides to control Asian ambrosia beetle, Xylosandrus crassiusculus. In Proceedings the 49th Annual Southern Nursey Association Research Conference; James, B.L., Ed.; Southern Nursery Association: Marietta, GA, USA, 2004; pp. 152–155. [Google Scholar]
- Frank, S.D.; Bambara, S. The Granulate [Asian] Ambrosia Beetle; North Carolina Extension Service, 2009. Available online: https://www.ces.ncsu.edu/depts/ent/notes/O&T/trees/note111/note111.html (accessed on 25 May 2016).
- Reddy, G.K.; Verghese, A. Management of shot hole borer, Xylosandrus crassiusculus (Motschulsky) (Coleoptera: Scolytidae) on grapes. Pest Manag. Hortic. Ecosyst. 2006, 12, 107–115. [Google Scholar]
- Joseph, M.; Prakasan, C.B.; Vijayalakshmi, C.K.; Sreedharan, K. Laboratory evaluation of the systemic fungicide propiconazole (Tilt 25EC) against the ‘ambrosia’ fungus, the food of shot hole borer Xylosandrus compactus (Eichhoff) (Coleoptera: Scolytidae) attacking coffee. In Proceedings of the 15th Plantation Crops Symposium Placrosym XV; Central Coffee Research Institute, Coffee Research Station: Mysore, India, 2002; pp. 613–616. [Google Scholar]
- Kagezi, G.; Kucel, P.; Olal, S.; Pinard, F.; Seruyange, J.; Musoli, P.; Kangire, A. In vitro inhibitory effect of selected fungicides on mycelial growth of ambrosia fungus associated with the black coffee twig borer, Xylosandrus compactus Eichhoff (Coleoptera: Curculionidae) in Uganda. Afr. J. Agric. Res. 2015, 10, 2322–2328. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Mean (±SE) Number of | |||
|---|---|---|---|---|
| Galleries with Fungal Growth per Tree | Galleries with Eggs per Tree | Foundress X. germanus Recovered per Tree | Living X. germanus Recovered per Tree | |
| Flooded | 22.2 ± 5.5 a | 4.3 ± 1.7 | 37.5 ± 2.6 a | 3.2 ± 1.2 a |
| Flooded plus Permethrin | 10.2 ± 2.3 b | 0.0 ± 0.0 | 18.7 ± 3.2 b | 0.3 ± 0.2 b |
| Non-Flooded | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 |
| Non-Flooded plus Permethrin | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 |
| t; P | −5.04; p < 0.0001 | – | −6.03; <0.0001 | −3.03; 0.003 |
| Treatment | Mean (±SE) Number of | |||
|---|---|---|---|---|
| Galleries with Fungal Growth per Tree | Galleries with Eggs per Tree | Foundress X. germanus Recovered per Tree | Living X. germanus Recovered per Tree | |
| Flooded | 10.4 ± 3.8 a | 2.6 ± 1.9 | 24.8 ± 6.5 b | 13.8 ± 7.2 a |
| Flooded plus Azoxystrobin | 2.0 ± 1.0 b | 0.0 ± 0.0 | 21.8 ± 6.7 b | 3.6 ± 1.9 b |
| Flooded plus Potassium Phosphite | 4.4 ± 1.3 b | 0.0 ± 0.0 | 31.6 ± 4.9 a | 7.2 ± 2.2 b |
| Non-Flooded | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 |
| Non-Flooded plus Azoxystrobin | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 |
| Non-Flooded plus Potassium Phosphite | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 |
| t; p: Flooded vs. Flooded plus Azoxystrobin | −4.77; <0.0001 | – | −0.98; 0.33 | −5.08; <0.0001 |
| t; p: Flooded vs. Flooded plus Potassium Phosphite | −3.38; 0.0007 | – | 2.02; 0.043 | −3.16; 0.002 |
| Treatment | Mean (± SE) Number of | |||
|---|---|---|---|---|
| Eggs Recovered per Tree | Larvae Recovered per Tree | Pupae Recovered per Tree | X. germanus Recovered per Tree | |
| Flooded | 0.0 ± 0.0 | 2.5 ± 1.5 a | 1.0 ± 1.0 | 0.5 ± 0.3 a |
| Flooded plus Azoxystrobin | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.3 ± 0.3 a |
| Flooded plus Potassium Phosphite | 0.0 ± 0.0 | 1.0 ± 0.6 a | 0.0 ± 0.0 | 1.3 ± 0.9 a |
| Non-Flooded | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 |
| Non-Flooded plus Azoxystrobin | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 |
| Non-Flooded plus Potassium Phosphite | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 |
| t; P: Flooded vs. Flooded plus Azoxystrobin | – | – | – | −0.57; 0.57 |
| t; P: Flooded vs. Flooded plus Potassium Phosphite | – | −1.55; 0.12 | – | 1.10; 0.27 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ranger, C.M.; Schultz, P.B.; Reding, M.E.; Frank, S.D.; Palmquist, D.E. Flood Stress as a Technique to Assess Preventive Insecticide and Fungicide Treatments for Protecting Trees against Ambrosia Beetles. Insects 2016, 7, 40. https://doi.org/10.3390/insects7030040
Ranger CM, Schultz PB, Reding ME, Frank SD, Palmquist DE. Flood Stress as a Technique to Assess Preventive Insecticide and Fungicide Treatments for Protecting Trees against Ambrosia Beetles. Insects. 2016; 7(3):40. https://doi.org/10.3390/insects7030040
Chicago/Turabian StyleRanger, Christopher M., Peter B. Schultz, Michael E. Reding, Steven D. Frank, and Debra E. Palmquist. 2016. "Flood Stress as a Technique to Assess Preventive Insecticide and Fungicide Treatments for Protecting Trees against Ambrosia Beetles" Insects 7, no. 3: 40. https://doi.org/10.3390/insects7030040
APA StyleRanger, C. M., Schultz, P. B., Reding, M. E., Frank, S. D., & Palmquist, D. E. (2016). Flood Stress as a Technique to Assess Preventive Insecticide and Fungicide Treatments for Protecting Trees against Ambrosia Beetles. Insects, 7(3), 40. https://doi.org/10.3390/insects7030040