
Enhanced Trapping of Yellowjacket Wasps (Hymenoptera: Vespidae) via Spatial Partitioning of Attractants

{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Yellowjacket Traps
2.2. Attractants
2.3. Yellowjacket Field Studies
2.4. Data Analysis
3. Results
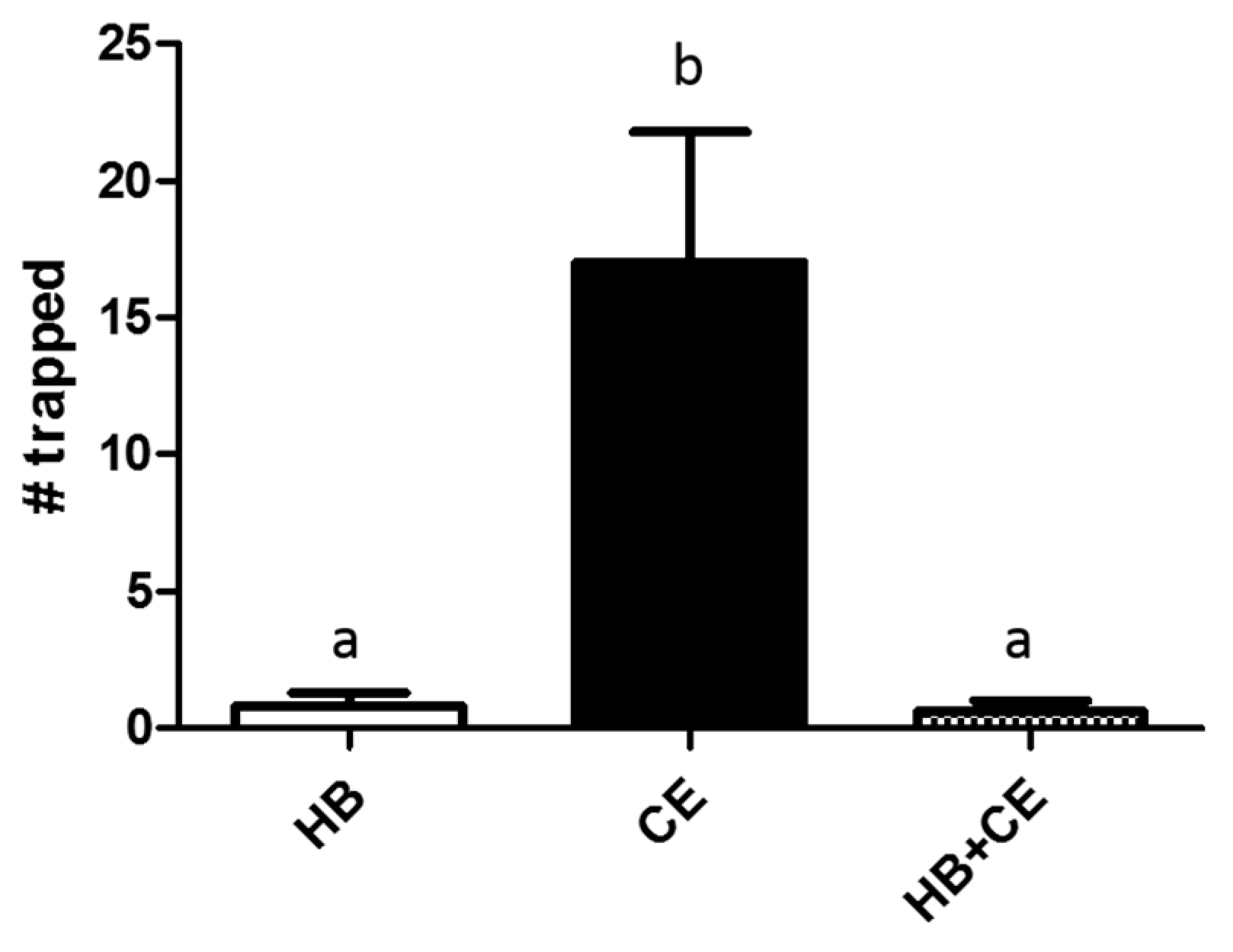
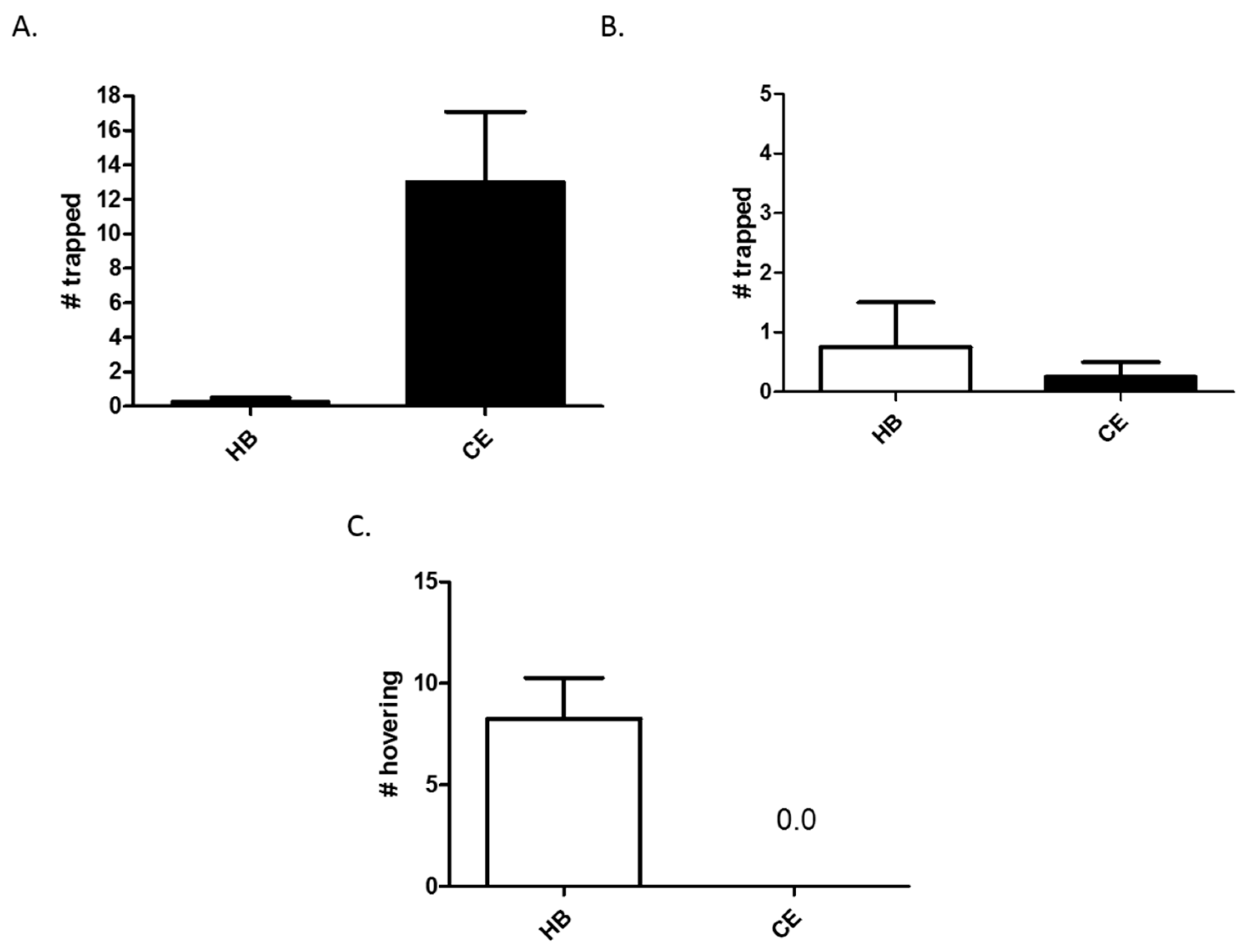
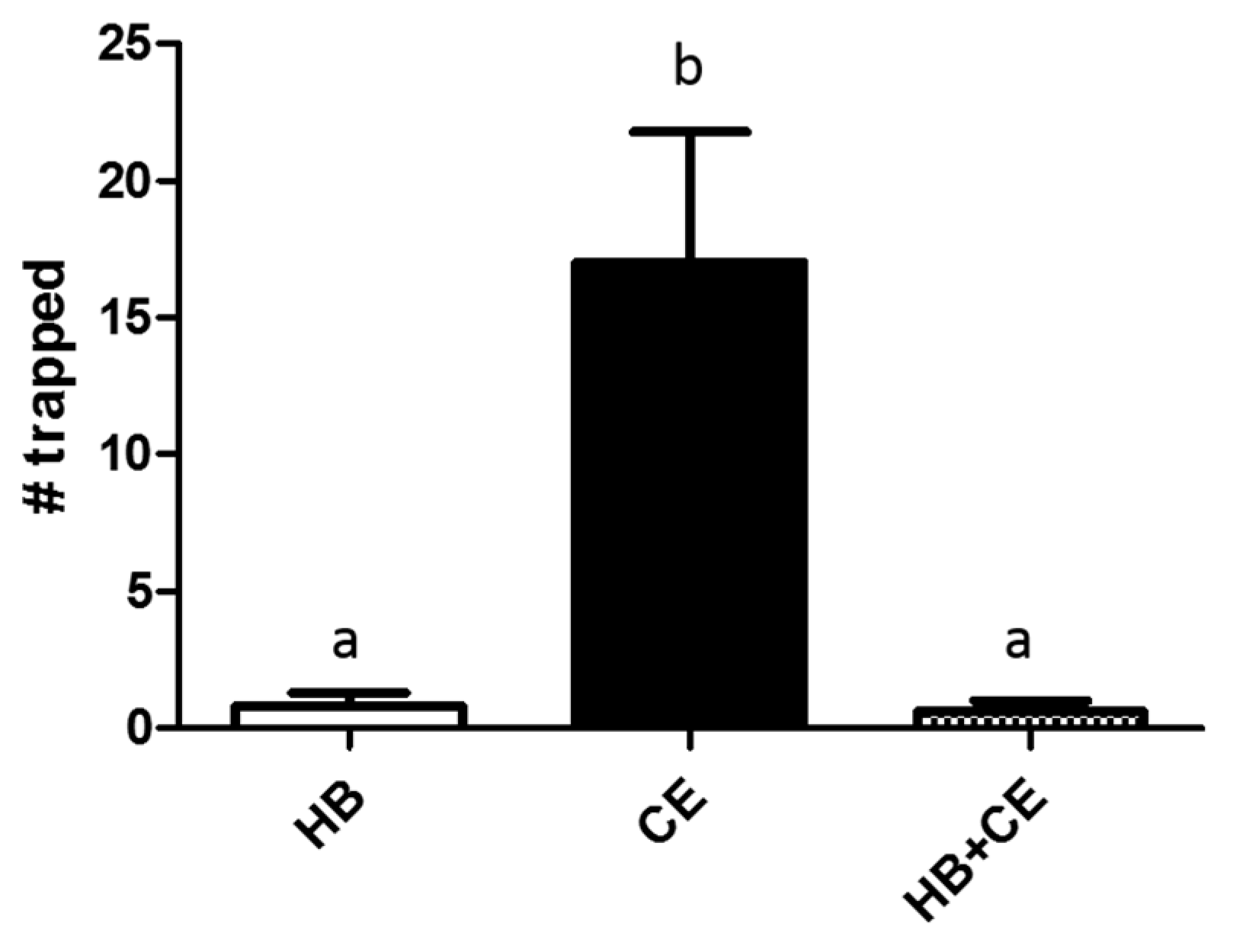
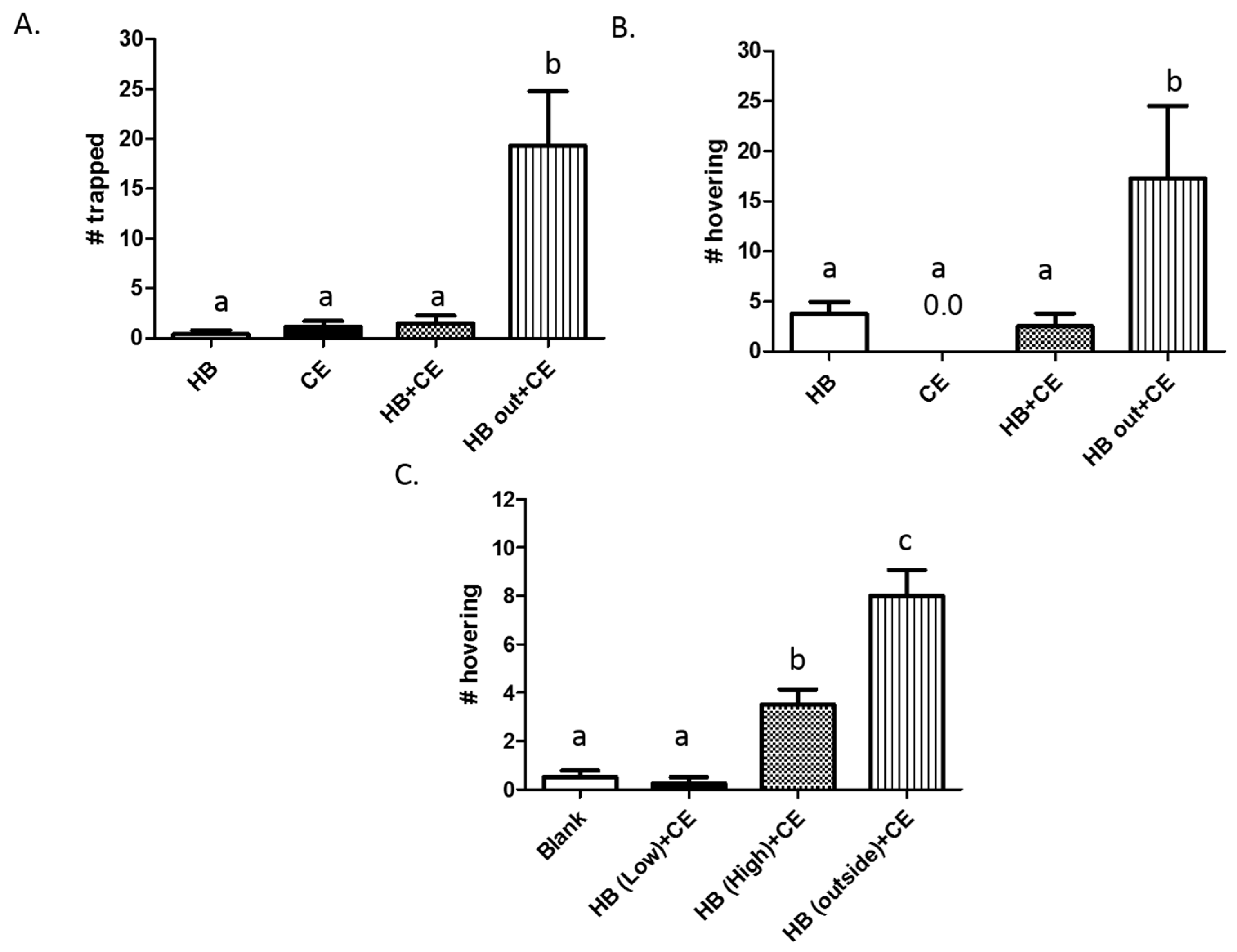
3.1. Comparison of Heptyl Butyrate and Chicken Extract as Attractants
3.2. Effects of Heptyl Butyrate on the Attractiveness of Chicken Extract
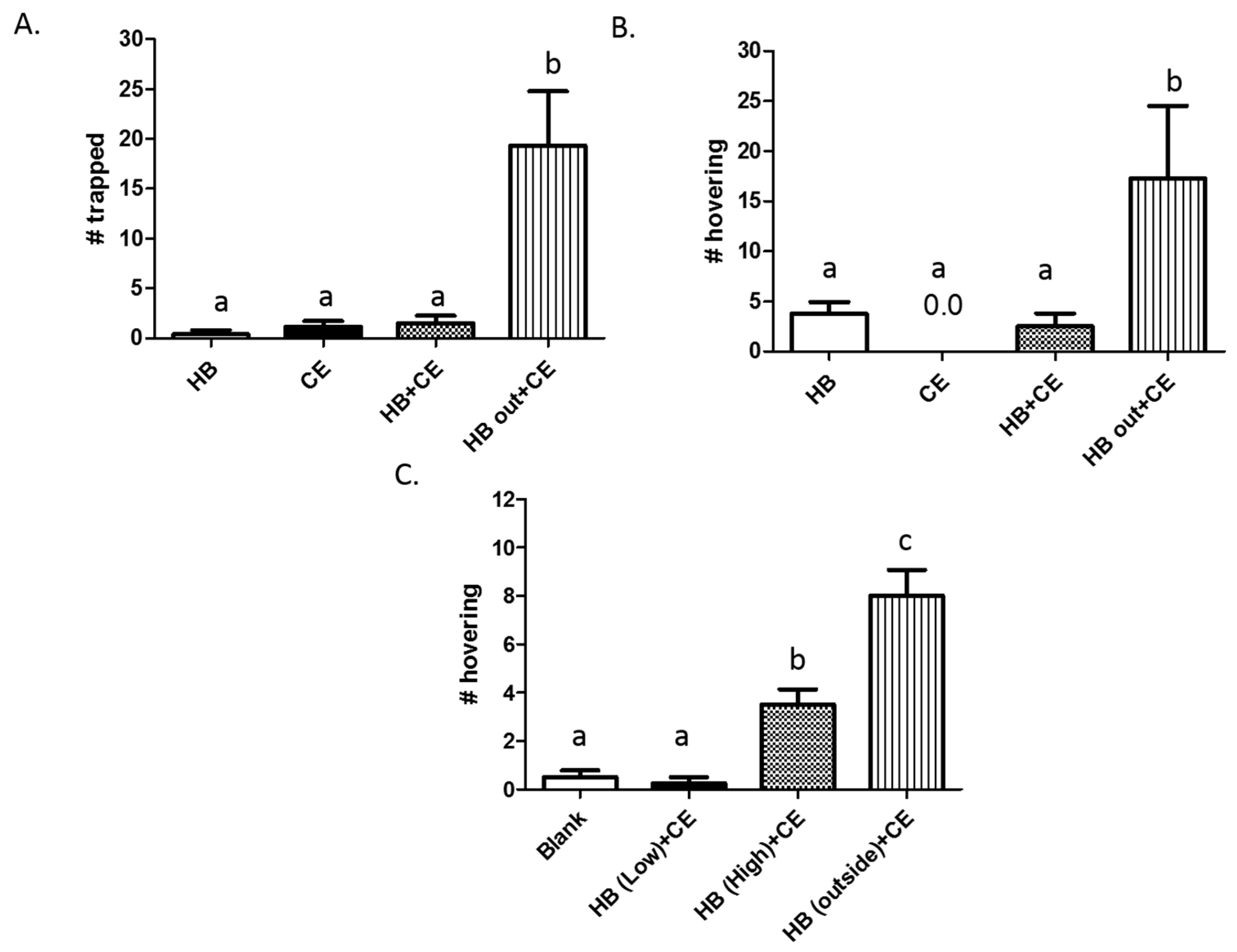
3.3. Partitioning of Heptyl Butyrate and Chicken Extract Results in Increased Trapping
4. Discussion
5. Conclusions
Author Contributions
Conflicts of Interest
References
- MacDonald, J.F.; Akre, R.D.; Keyel, R.E. The German Yellowjacket (Vespula germanica) problem in the United States (Hymenoptera: Vespidae). Bull. Entomol. Soc. Am. 1980, 26, 436–442. [Google Scholar] [CrossRef]
- MacDonald, J.F.; Matthews, R.W. Nesting biology of the southern yellow jacket, Vespula squamosal (Hymenoptera: Vespidae): Social parasitism and independent founding. J. Kansas Entomol. Soc. 1984, 57, 134–151. [Google Scholar]
- Langley, R.L.; Mack, K.; Haileyesus, T.; Proescholdbell, S.; Annest, J.L. National estimates of noncanine bite and sting injuries treated in US hospital emergency departments, 2001–2010. Wilderness Environ. Med. 2014, 25, 14–23. [Google Scholar] [CrossRef] [PubMed]
- Vetter, R.S.; Visscher, P.K.; Camazine, S. Mass envenomations by honey bees and wasps. West J. Med. 1999, 170, 223–227. [Google Scholar] [PubMed]
- MacIntyre, P.; Hellstrom, J. An Evaluation of the Cost of Pest Wasps (Vespula spp) in New Zealand; Department of Conservation and Ministry for Primary Industries: Wellington, DC, USA, 2015; p. 44.
- Waddell, L.R.; Drobatz, L.J. Massive envenomation by Vespula spp in two dogs. J. Vet. Emerg. Crit. Care 1999, 9, 67–71. [Google Scholar] [CrossRef]
- Landolt, P.; Zhang, Q. Discovery and development of chemical attractants used to trap pestiferous social wasps (Hymenoptera: Vespidae). J. Chem. Ecol. 2016, 42, 655–665. [Google Scholar] [CrossRef] [PubMed]
- Braband, B.; Frye, M.; Lampman, J.; Marvin, D. Yellowjacket Trapping Efficacy Trials: NYS IPM Program 2014 & 2015; New York State IPM Program: New York, NY, USA, 2016. [Google Scholar]
- Rust, M.K.; Reierson, D.A.; Vetter, R.S. Developing Baits for the Control of Yellowjackets in California; Final Report 2010 for Structural Pest Control Board. Structural Pest Control Board, Grant No. 041-04; University of California: Oakland, CA, USA, 2010; pp. 1–33. [Google Scholar]
- Mattheis, J.P.; Buchanan, D.A.; Feldmann, J.R. Change in apple fruit volatiles after storage in atmospheres inducing anaerobic metabolism. J. Agric. Food Res. 1991, 39, 1602–1605. [Google Scholar] [CrossRef]
- McGovern, T.P.; Davis, H.G.; Beroza, M.; Ingani, J.C.; Eddy, G.W. Esters highly attractive to Vespula spp. J. Econ. Entomol. 1970, 63, 1134–1136. [Google Scholar] [CrossRef]
- Landolt, P. Chemical attractants for trapping yellow jackets Vespula germanica and Vespula pensylvanica (Hymenoptera: Vespidae). Environ. Entomol. 1998, 27, 1229–1234. [Google Scholar] [CrossRef]
- Richter, M.R. Social wasp (Hymenoptera: Vespidae) foraging behavior. Annu. Rev. Entomol. 2000, 45, 121–150. [Google Scholar] [CrossRef] [PubMed]
- Spurr, E.B. Protein bait preferences of wasps (Vespula vulgaris and Vespula germanica) at Mt Thomas, Canterbury, New Zealand. N. Z. J. Zool. 1995, 22, 281–289. [Google Scholar] [CrossRef]
- Wood, G.M.; Hopkins, D.C.; Schellhorn, N.A. Preference by Vespula germanica (Hymenoptera: Vespidae) for processed meats: implications for toxic baiting. J. Econ. Entomol. 2006, 99, 263–267. [Google Scholar] [CrossRef] [PubMed]
- Rust, M.K.; Choe, W.H.; Erin, W.R.; Campbell, K.; Kabashima, J.; Dimson, M. Controlling yellow jackets with fipronil-based protein baits in urban recreational areas. Int. J. Pest Manag. 2016. [Google Scholar] [CrossRef]
- Reirson, D.A.; Rust, M.K.; Vetter, R.S. Traps and protein bait to suppress populations of yellowjackets (Hymenoptera: Vespidae). In Proceedings of the Sixth International Conference on Urban Pests, Budapest, Hungary, 13–16 July 2008; pp. 267–274.
- Unelius, C.; El-Sayed, A.M.; Twidle, A.M.; Stringer, L.D.; Manning, L.M.; Sullivan, T.E.E.; Brown, R.L.; Noble, A.D.L. Volatiles from green lipped mussel as a lead to vespid wasp attractants. J. Appl. Entomol. 2013, 138, 87–95. [Google Scholar] [CrossRef]
- Unelius, C.; Suckling, D.; Brown, R.L.; Josvai, J.K.; El-Sayed, A.M. Combining odours isolated from phylogenetically diverse sources yields a better lure for yellowjackets. Pest Manag. Sci. 2016, 72, 760–769. [Google Scholar] [CrossRef] [PubMed]
- Aldrich, J.R.; Zhang, Q.H.; Zhang, A. Synergistic chemical attraction of the eastern yellowjacket Vespula maculifrons. J. Entomol. Sci. 2004, 39, 643–653. [Google Scholar]
- Foster, S.P.; Harris, M.O. Behavioral manipulation methods for insect pest-management. Annu. Rev. Entomol. 1997, 42, 123–146. [Google Scholar] [CrossRef] [PubMed]
- El-Sayed, A.M.; Manning, L.A.; Unelius, C.R.; Park, K.C.; Stringer, L.D.; White, N.; Bunn, B.; Twidle, A.; Suckling, D.M. Attraction and antennal response of the common wasp, Vespula vulgaris, to selected synthetic chemicals in New Zealand beech forest. Pest Manag. Sci. 2009, 65, 975–981. [Google Scholar] [CrossRef] [PubMed]
- Ross, D.R.; Shukle, R.H.; MacDonald, J.F. Meat extracts attractive to scavenger Vespula in Eastern North America (Hymenoptera: Vespidae). J. Econ. Entomol. 1984, 77, 637–642. [Google Scholar] [CrossRef]
- Landolt, P.J.; Toth, M.; Josvai, J. First European report of social wasps trapped in response to acetic acid, isobutanol, 2-methyl-2-propanol, and heptyl butyrate in tests conducted in Hungary. Bull. Insectol. 2007, 60, 7–11. [Google Scholar]
- Buteler, M.; Lozada, M.; Adamo, P.D.; Melo, R.A.L.; Stadler, T. Behavioural responses of Vespula germanica (Hymenoptera: Vespidae) wasps exposed to essential oils. Austral Entomol. 2016, 55, 308–315. [Google Scholar] [CrossRef]
- Zhang, Q.H.; Schneidmiller, R.G.; Hoover, D.R. Essential oils and their compositions as spatial repellents for pestiferous social wasps. Pest. Manag. Sci. 2013, 69, 542–552. [Google Scholar] [CrossRef] [PubMed]
- Boeve, J.L.; Honraet, K.; Rossel, B. Screening of repellents against vespid wasps. Insects 2014, 5, 272–286. [Google Scholar] [CrossRef] [PubMed]
- Weston, R.J.; Woolhouse, A.D.; Spurr, E.B.; Harris, R.K.; Suckling, D.M. Spiroacetals and other venom constituents as potential was attractants. J. Chem. Ecol. 1997, 23, 553–568. [Google Scholar] [CrossRef]
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liang, D.; Pietri, J.E. Enhanced Trapping of Yellowjacket Wasps (Hymenoptera: Vespidae) via Spatial Partitioning of Attractants. Insects 2017, 8, 17. https://doi.org/10.3390/insects8010017
Liang D, Pietri JE. Enhanced Trapping of Yellowjacket Wasps (Hymenoptera: Vespidae) via Spatial Partitioning of Attractants. Insects. 2017; 8(1):17. https://doi.org/10.3390/insects8010017
Chicago/Turabian StyleLiang, Dangsheng, and Jose E. Pietri. 2017. "Enhanced Trapping of Yellowjacket Wasps (Hymenoptera: Vespidae) via Spatial Partitioning of Attractants" Insects 8, no. 1: 17. https://doi.org/10.3390/insects8010017