
Discovering the Giant Nest Architecture of Grass-Cutting Ants, Atta capiguara (Hymenoptera, Formicidae)

Abstract
:1. Introduction
2. Material and Methods
2.1. Study Area
2.2. Nests Studied
2.3. Internal Architecture
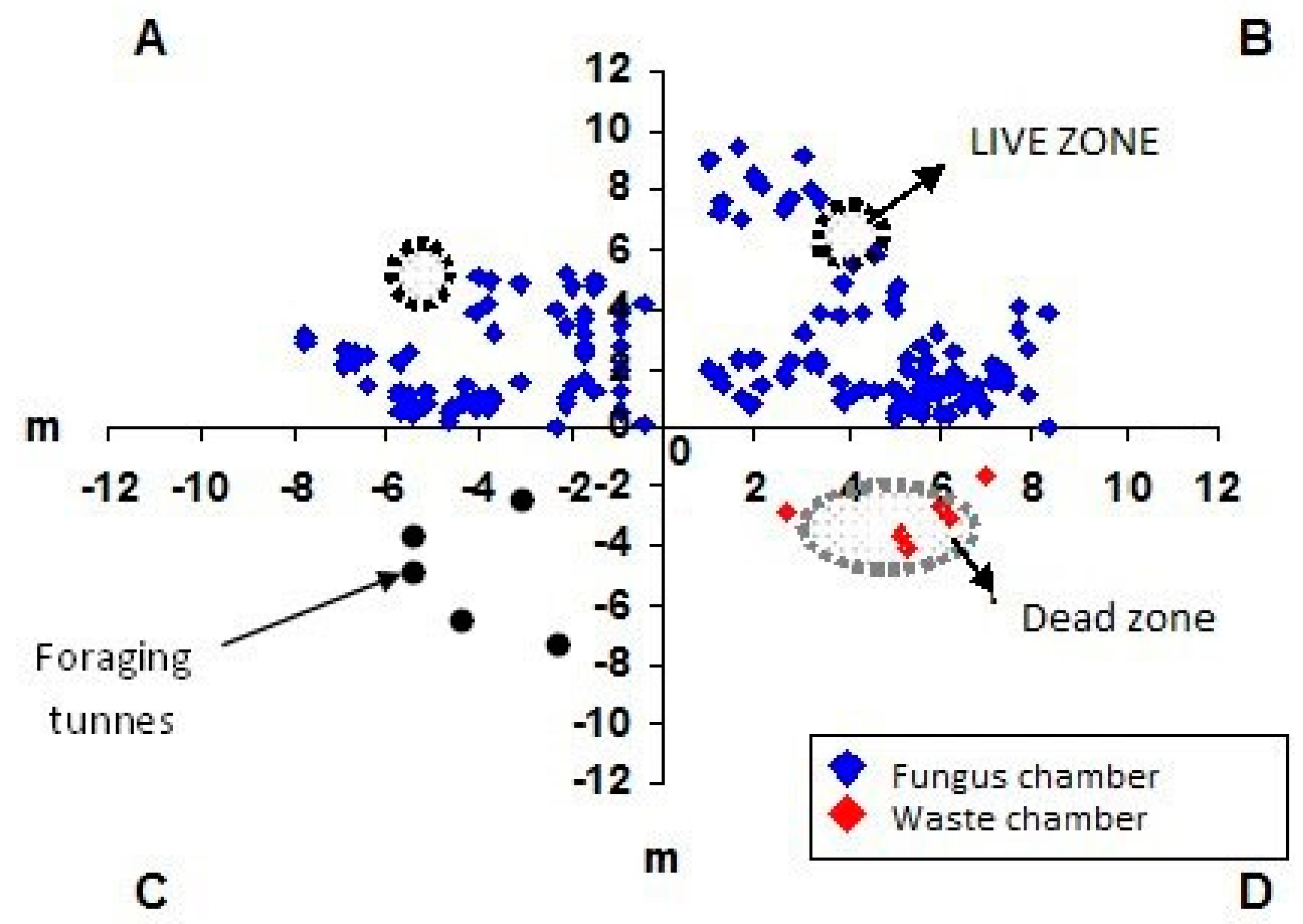
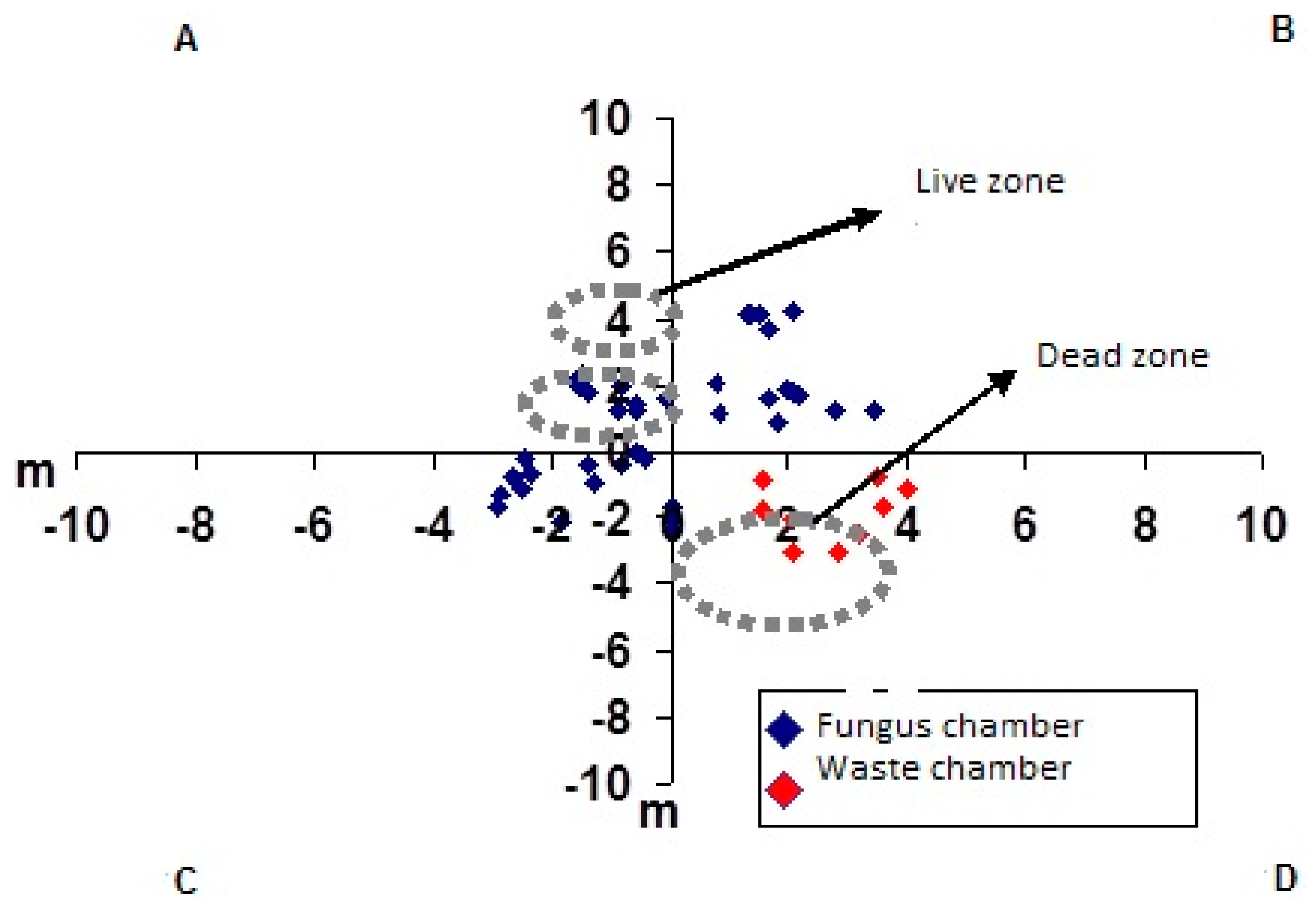
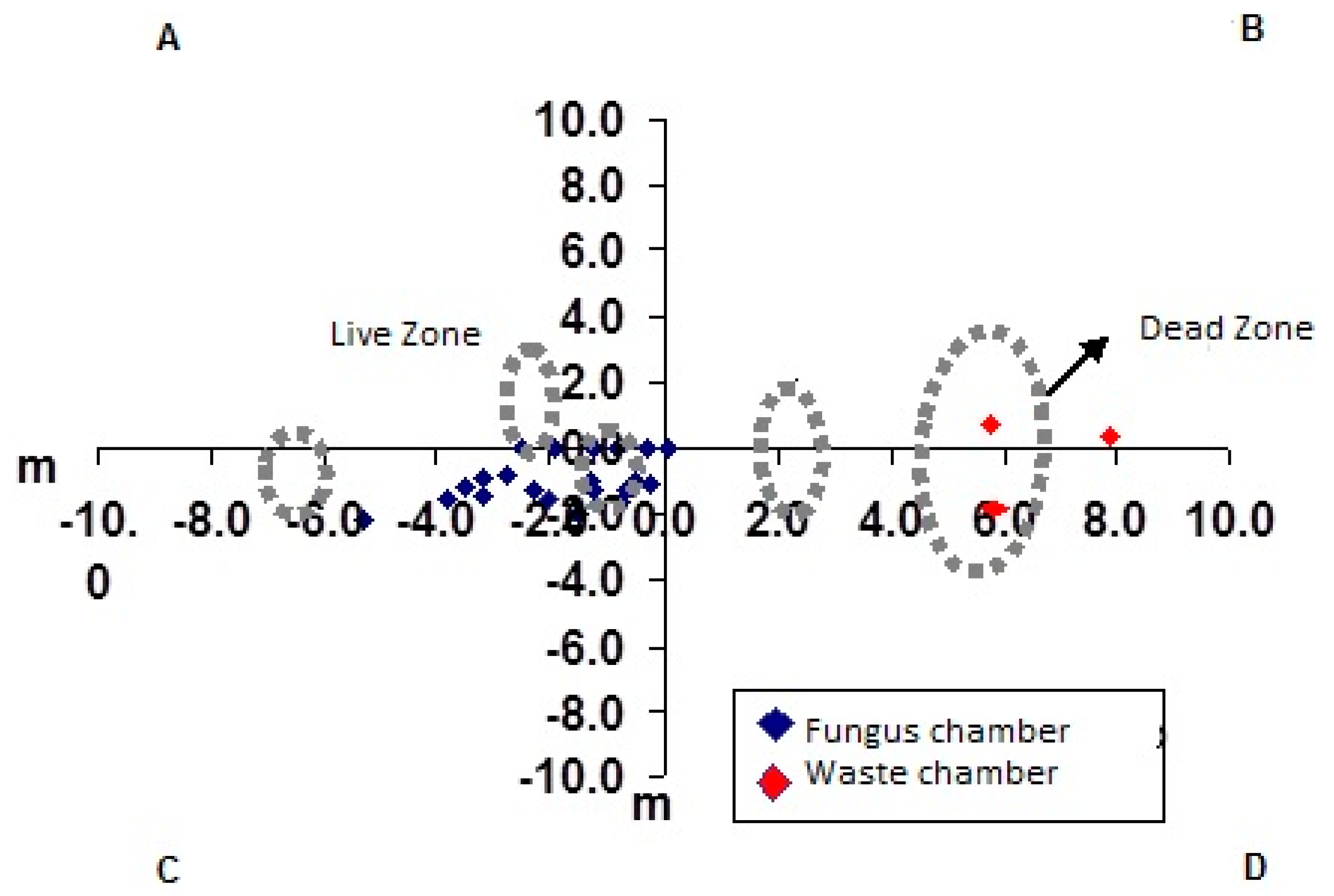
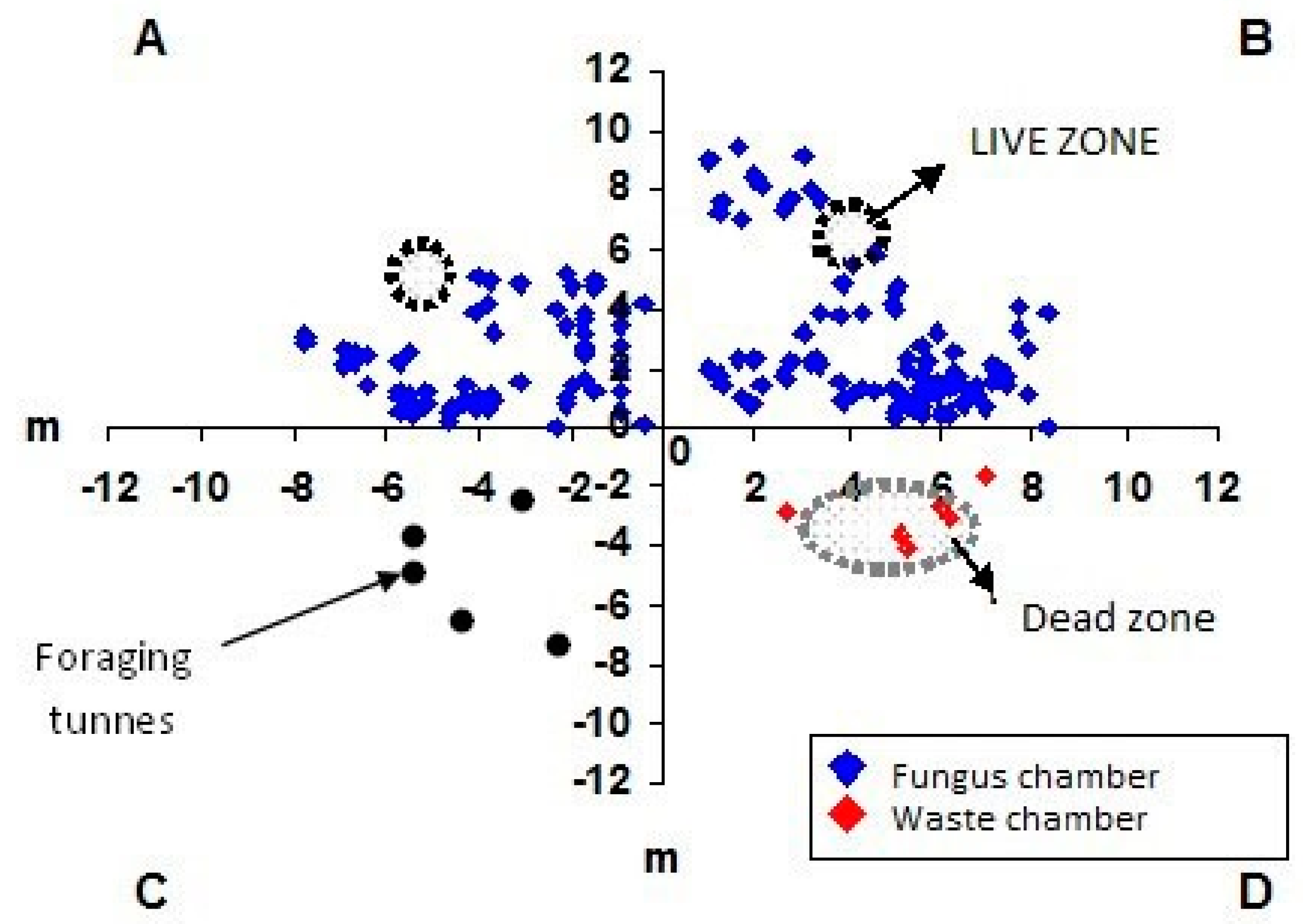
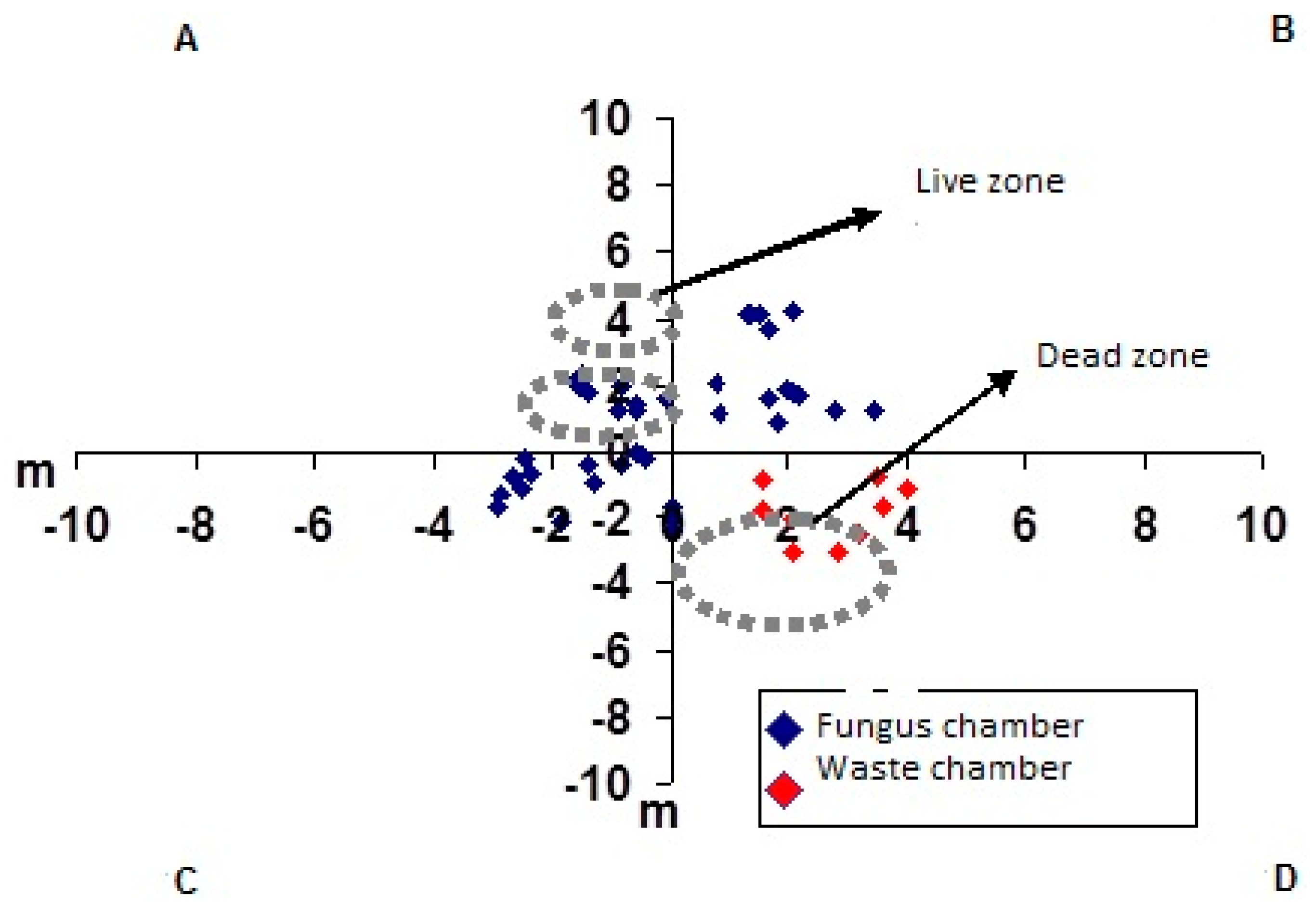
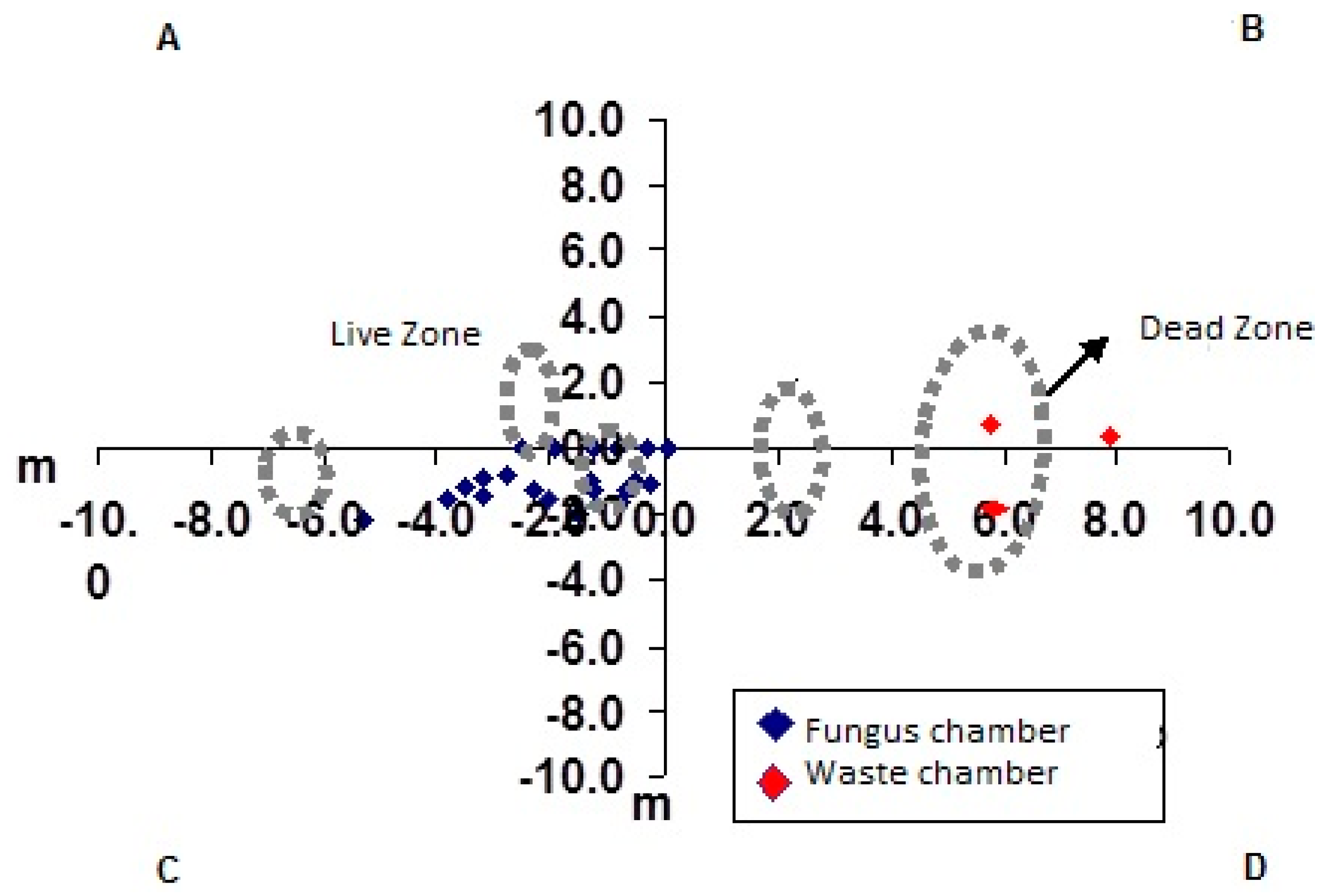
3. Results and Discussion
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Moreira, A.A.; Forti, L.C.; Andrade, A.P.P.; Boaretto, M.A.C.; Lopes, J.F.S. Nest Architecture of Atta laevigata (F. Smith, 1858) (Hymenoptera: Formicidae). Stud. Neotrop. Fauna Environ. 2004, 39, 109–116. [Google Scholar] [CrossRef]
- Pretto, D.R. Arquitetura dos Túneis de Forrageamento e do Ninho de Atta sexdens rubropilosa Forel, 1908 (Hym., Formicidae), Dispersão de Substrato e Dinâmica de Inseticida. [Dissertação (Mestrado em Agronomia/ Proteção de Plantas)]. Ph.D. Thesis, Faculdade de Ciências Agronômicas, Universidade Estadual Paulista, Botucatu, Brazil, 1996. (In Portuguese). [Google Scholar]
- Moreira, A.A.; Forti, L.C.; Boaretto, M.A.C.; Andrade, A.P.P.; Lopes, J.F.S.; Ramos, V.M. External and internal structure of Atta bisphaerica Forel (Hymenoptera, Formicidae) nests. J. Appl. Entomol. 2004, 128, 200–2003. [Google Scholar] [CrossRef]
- Cassill, D.; Tschinkel, W.R.; Vinson, S.B. Nest complexity, group size and brood rearing in the fire ant Solenopsis invicta. Insects Soc. 2002, 49, 158–163. [Google Scholar] [CrossRef]
- Tschinkel, W.R. Subterranean ant nests: trace fossils past and future? Palaeo 2003, 192, 321–333. [Google Scholar] [CrossRef]
- Pereira-da-Silva, V.; Forti, L.C.; Cardoso, L.G. Dinâmica populacional e caracterização dos ninhos de Acromyrmex coronatus (Fabricius, 1804) (Hymenoptera: Formicidae). Rev. Bras. Entomol. 1981, 25, 87–93. (In Portuguese) [Google Scholar]
- Fowler, H.G.; Forti, L.C.; Pereira-da-Silva, V.; Saes, N.B. Economics of Grass-Cutting Ants. In Fire Ants and Leaf-Cutting Ants; Lofgren, C.S., Vander Meer, R.K., Eds.; Westview Press: Boulder, CO, USA, 1986; pp. 18–35. [Google Scholar]
- Andrade, A.P.P. Biologia e Taxonomia Comparadas das Subespécies de Acromyrmex subterraneus Forel, 1893 (Hymenoptera, Formicidae) e a Contaminação das Operárias por Iscas Tóxicas. Ph.D. Thesis, Instituto de Biociências, Universidade Estadual Paulista, Botucatu, Brazil, 2002. (In Portuguese). [Google Scholar]
- Talbot, M. A comparison of two ants of the genus Fomica. Ecology 1948, 29, 316–325. [Google Scholar] [CrossRef]
- Autuori, M. Contribuição para o conhecimento da saúva (Atta spp.) (Hymenoptera: Formicidae): III—Escavação de um sauveiro (Atta sexdens rubropilosa Forel, 1908). Arq. Inst. Biol. 1942, 3, 136–148. (In Portuguese) [Google Scholar]
- Talbot, M.; Kennedy, C.H. The slave-making ant Formica sanguinea subintegra Emery, its raids, nuptial flights and nest structure. Ann. Entomol. Soc. Am. 1940, 33, 560–577. [Google Scholar] [CrossRef]
- Lavigne, R.J. Bionomics and nest structure of Pogonomyrmex occidentalis (Hymenoptera, Formicidae). Ann. Entomol. Soc. Am. 1969, 62, 1166–1175. [Google Scholar] [CrossRef]
- Wheeler, J.; Rissing, S.W. Natural history of Veromessor pergandei, I: The nest (Hymenoptera: Formicidae). Pan. Pac. Entomol. 1975, 51, 205–216. [Google Scholar]
- Mackay, W.P. A comparison of the nest phenologies of three species of Pogonomyrmex harvester ants (Hymenoptera: Formicidae). Psyche 1981, 88, 25–74. [Google Scholar] [CrossRef]
- Kugler, C.; Hincapie, C.M. Ecology of Pogonomyrmex mayri: Distribution, abundance, nest structure and diet. Biotropica 1983, 15, 190–198. [Google Scholar] [CrossRef]
- Tschinkel, W.R. Seasonal life history and nest architecture of a winter-active ant, Prenolepsis imparis. Insects Soc. 1987, 34, 143–164. [Google Scholar] [CrossRef]
- Bristow, C.M.; Cappaert, D.; Campbell, N.J.; Heise, A. Nest structure and colony cycle of the Allegheny mound ant, Formica exsectoides Forel (Hymenoptera: Formicidae). Insects Soc. 1992, 39, 385–402. [Google Scholar] [CrossRef]
- Crosland, M.W.J. Nest and colony structure in the primitive ant, Harpegnathos venator (Smith) (Hymenoptera, Formicidae). Pan. Pac. Entomol. 1995, 71, 18–23. [Google Scholar]
- Wilson, E.O. Sociobiology: The New Synthesis; Harvard University Press: Cambridge, MA, USA, 1975; p. 697. [Google Scholar]
- Jonkman, J.C.M. The external and internal structure and growth of nests of the leaf-cutting ant Atta vollenweideri Forel, 1893 (Hym.: Formicidae) Part II. J. Appl. Entomol. 1980, 89, 217–246. [Google Scholar]
- Jonkman, J.C.M. The external and internal structure and growth of nests of the leaf-cutting ant Atta vollenweideri Forel, 1893 (Hym.: Formicidae) Part I. J. Appl. Entomol. 1980, 89, 158–173. [Google Scholar] [CrossRef]
- Porter, S.D.; Tschinkel, W.R. Fire ant thermal preferences: Behavioral control of growth and metabolism. Behav. Ecol. Sociobiol. 1993, 32, 321–329. [Google Scholar] [CrossRef]
- Porter, S.D. Impact of temperature on colony growth and development rates of the ant, Solenopsis invicta. J. Insect Physiol. 1988, 34, 1127–1133. [Google Scholar] [CrossRef]
- Bollazzi, M.; Roces, F. Thermal preferences for fungus culturing and brood location by workers of the thatching grass-cutting ant Acromyrmex heyeri. Insects Soc. 2002, 49, 153–157. [Google Scholar] [CrossRef]
- Fowler, H.G.; Schlindwein, M.N.; Schlitter, F.M.; Forti, L.C. A simple method for determining location of foraging ant nests using leaf cutting ants as a model. J. Appl. Entomol. 1993, 116, 420–422. [Google Scholar] [CrossRef]
- Boaretto, M.A.C. Seleção de Substratos com Potencial Para uso em iscas Granuladas Para as Saúvas Atta capiguara Gonçalves, 1944 e Atta bisphaerica Forel, 1908 (Hymenoptera, Formicidae) e Isolamento do Fungo Simbionte. [Tese (Doutorado)]. Ph.D. Thesis, Faculdade de Ciências Agronômicas, Universidade Estadual Paulista, Botucatu, Brazil, 2000. (In Portuguese). [Google Scholar]
- Forti, L.C. Ecologia da saúva Atta capiguara Gonçalves, 1944 (Hymenoptera, Formicidae) em Pastagem. [Tese (Doutorado em Ciências)]. Ph.D. Thesis, Escola Superior de Agricultura “Luiz de Queiroz”, Universidade de São Paulo, Piracicaba, Brazil, 1985. (In Portuguese). [Google Scholar]
- Amante, E. Formiga saúva Atta capiguara, praga das pastagens. Biológico 1967, 33, 113–120. (In Portuguese) [Google Scholar]
- Amante, E. Saúva tira boi da pastagem. Coopercotia 1967, 23, 38–40. (In Portuguese) [Google Scholar]
- Carvalho, S. Atta (Neoatta) vollenweideri Forel, 1893, no Brasil: Ocorrência, Aspectos Externos e Internos do Sauveiro. Ph.D. Thesis, Centro de Ciências Rurais, Universidade Federal de Santa Maria, Santa Maria, Brazil, 1976. (In Portuguese). [Google Scholar]
- Bonetto, A. Las hormigas “cortadoras” de la provincia de Santa Fé (Generos: Atta y Acromyrmex). Direccion Gen. Rec. Nat. 1959, 2, 17–26. (In Portuguese) [Google Scholar]
- Jacoby, M. A arquitetura do ninho. In A Saúva: Uma Inteligência Nociva; Jacoby, M., Ed.; Serviço de Informação Agrícola: Rio de Janeiro, Brazil, 1950; pp. 21–31. (In Portuguese) [Google Scholar]
- Bollazzi, M. Ventilation of the giant nests of Atta leaf-cutting ants: Does underground circulating air enter the fungus chamber? Insects Soc. 2012, 59, 487–498. [Google Scholar] [CrossRef]
- Kleineidam, K.; Roces, F. Carbon dioxide concentrations and nest ventilations in nests of the leaf-cutting ant Atta vollenweideri. Insects Soc. 2000, 47, 241–248. [Google Scholar] [CrossRef]
- Kleineidam, K. Wind-induced ventilation of the giant nests of the leaf-cutting ant Atta vollenweideri. Naturwissenschaften 2001, 88, 301–305. [Google Scholar] [CrossRef] [PubMed]
- Waddington, S.J.; Hughes, W.H.O. Waste management in the leaf-cutting ant Acromyrmex echinatior: The role of worker size, age and plasticity. Behav. Ecol. Sociobiol. 2010, 64, 1219–1228. [Google Scholar] [CrossRef]
- Currie, C.R.; Mueller, U.G.; Malloch, D. The agricultural pathology of ant fungus gardens. Proc. Natl. Acad. Sci. USA 1999, 96, 7998–8002. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type | Width (cm) | Height (cm) | Length (m) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Mean | SD * | Max. | Min. | Mean | SD | Max. | Min. | Mean | SD | Max. | Min. | |
| Foraging Tunnels | 8 | 3.5 | 12 | 0.6 | 1.3 | 2.3 | 2 | 1 | 5.3 | 1.5 | 7.0 | 4.4 |
| Tunnels connecting to the chambers | 11 | 4.1 | 21 | 1.3 | 1.6 | 0.4 | 3.6 | 0.5 | 4.4 | 2.3 | 7.4 | 0.72 |
| Chambers | Height (m) | Width (m) | Volume (L) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Mean | SD ** | Max. | Min. | Mean | SD | Max. | Min. | Mean | SD | Max. | Min. | |
| Fungus (n * = 72) | 0.14 | 0.003 | 0.23 | 0.08 | 0.21 | 0.06 | 0.37 | 0.0006 | 5.7 | 4.0 | 17.5 | 0.9 |
| Waste (n = 10) | 1.4 | 0.57 | 2.7 | 0.6 | 0.32 | 0.11 | 0.53 | 0.16 | 172.9 | 146.6 | 535.5 | 20.7 |
| Chambers | Height (m) | Width (m) | Volume (L) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Mean | SD ** | Max. | Min. | Mean | SD | Max. | Min. | Mean | SD | Max. | Min. | |
| Fungus (n * = 40) | 0.17 | 0.5 | 0.38 | 0.8 | 0.21 | 0.5 | 0.32 | 0.08 | 7.5 | 5.5 | 22.3 | 0.8 |
| Waste (n = 8) | 0.8 | 0.3 | 1.1 | 0.7 | 0.14 | 0.04 | 0.4 | 0.16 | 32.07 | 39.8 | 77.9 | 5.24 |
| Chambers | Height (m) | Width (m) | Volume (L) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Mean | SD ** | Max. | Min. | Mean | SD | Max. | Min. | Mean | SD | Max. | Min. | |
| Fungus (n * = 28) | 0.13 | 0.4 | 0.2 | 0.6 | 0.18 | 0.7 | 0.4 | 0.1 | 4.5 | 4.9 | 23.1 | 0.5 |
| Waste (n = 8) | 0.68 | 0.21 | 0.8 | 0.4 | 0.47 | 0.08 | 0.60 | 0.15 | 43.55 | 33.05 | 75.4 | 9.42 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Forti, L.C.; Protti de Andrade, A.P.; Camargo, R.D.S.; Caldato, N.; Moreira, A.A. Discovering the Giant Nest Architecture of Grass-Cutting Ants, Atta capiguara (Hymenoptera, Formicidae). Insects 2017, 8, 39. https://doi.org/10.3390/insects8020039
Forti LC, Protti de Andrade AP, Camargo RDS, Caldato N, Moreira AA. Discovering the Giant Nest Architecture of Grass-Cutting Ants, Atta capiguara (Hymenoptera, Formicidae). Insects. 2017; 8(2):39. https://doi.org/10.3390/insects8020039
Chicago/Turabian StyleForti, Luiz Carlos, Ana Paula Protti de Andrade, Roberto Da Silva Camargo, Nadia Caldato, and Aldenise Alves Moreira. 2017. "Discovering the Giant Nest Architecture of Grass-Cutting Ants, Atta capiguara (Hymenoptera, Formicidae)" Insects 8, no. 2: 39. https://doi.org/10.3390/insects8020039
APA StyleForti, L. C., Protti de Andrade, A. P., Camargo, R. D. S., Caldato, N., & Moreira, A. A. (2017). Discovering the Giant Nest Architecture of Grass-Cutting Ants, Atta capiguara (Hymenoptera, Formicidae). Insects, 8(2), 39. https://doi.org/10.3390/insects8020039