
Getting More Power from Your Flowers: Multi-Functional Flower Strips Enhance Pollinators and Pest Control Agents in Apple Orchards

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design
2.2. Flower Strips
2.3. Flower-Visitor Surveys
2.4. Apple Tree Surveys
2.5. Sentinel Egg Cards
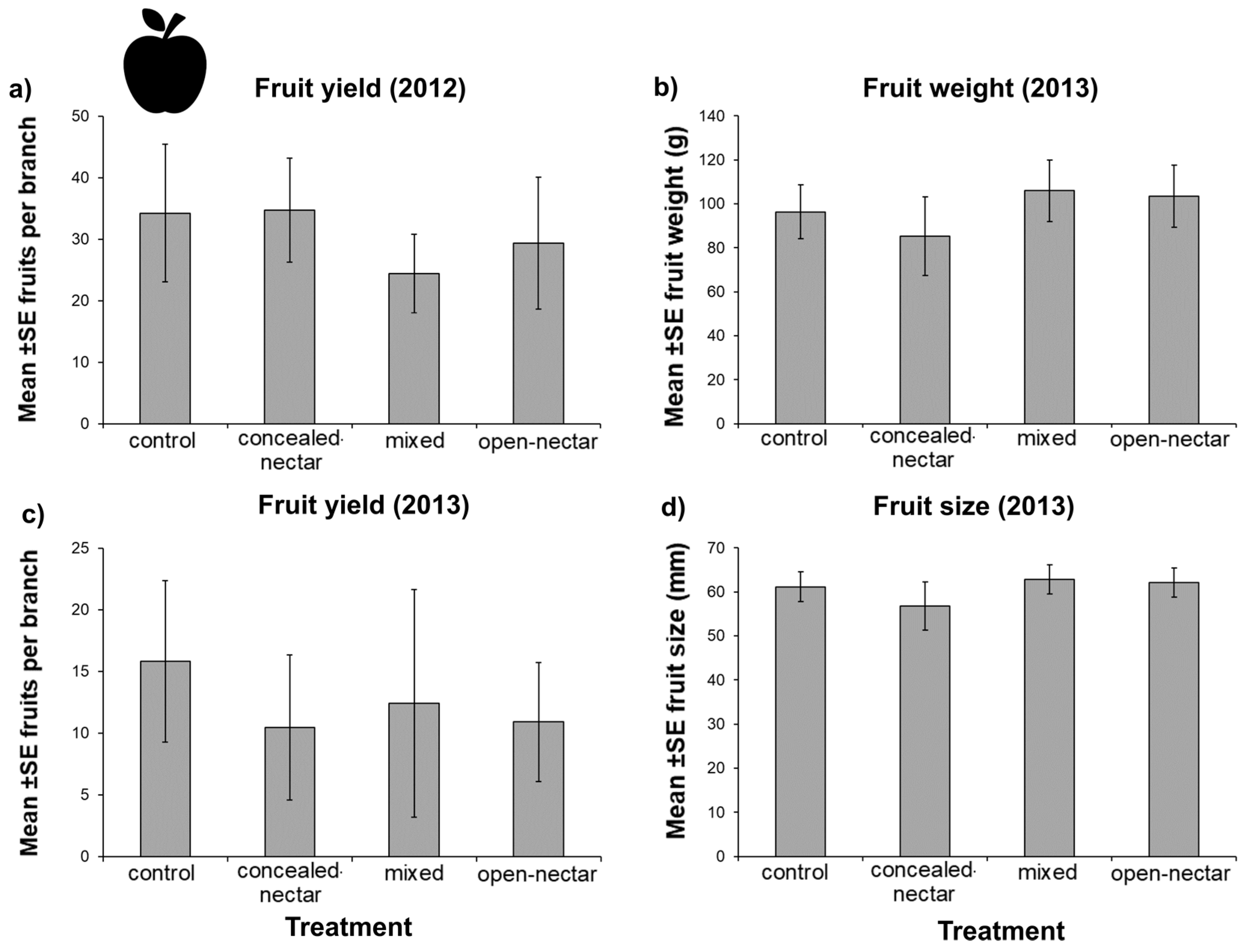
2.6. Fruit Yield
2.7. Statistical Analyses
3. Results
3.1. Flower Strips
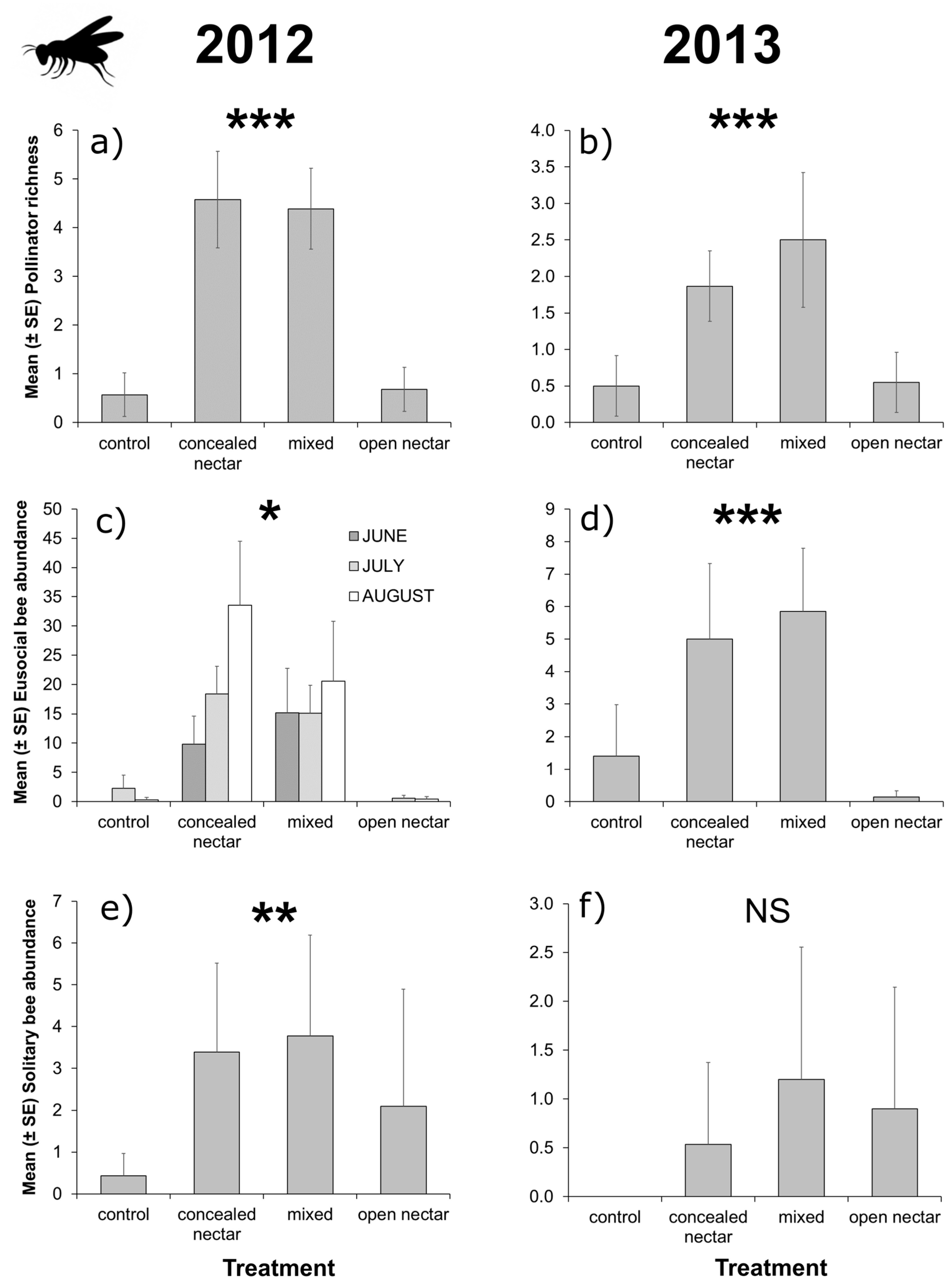
3.2. Flower-Visitor Surveys
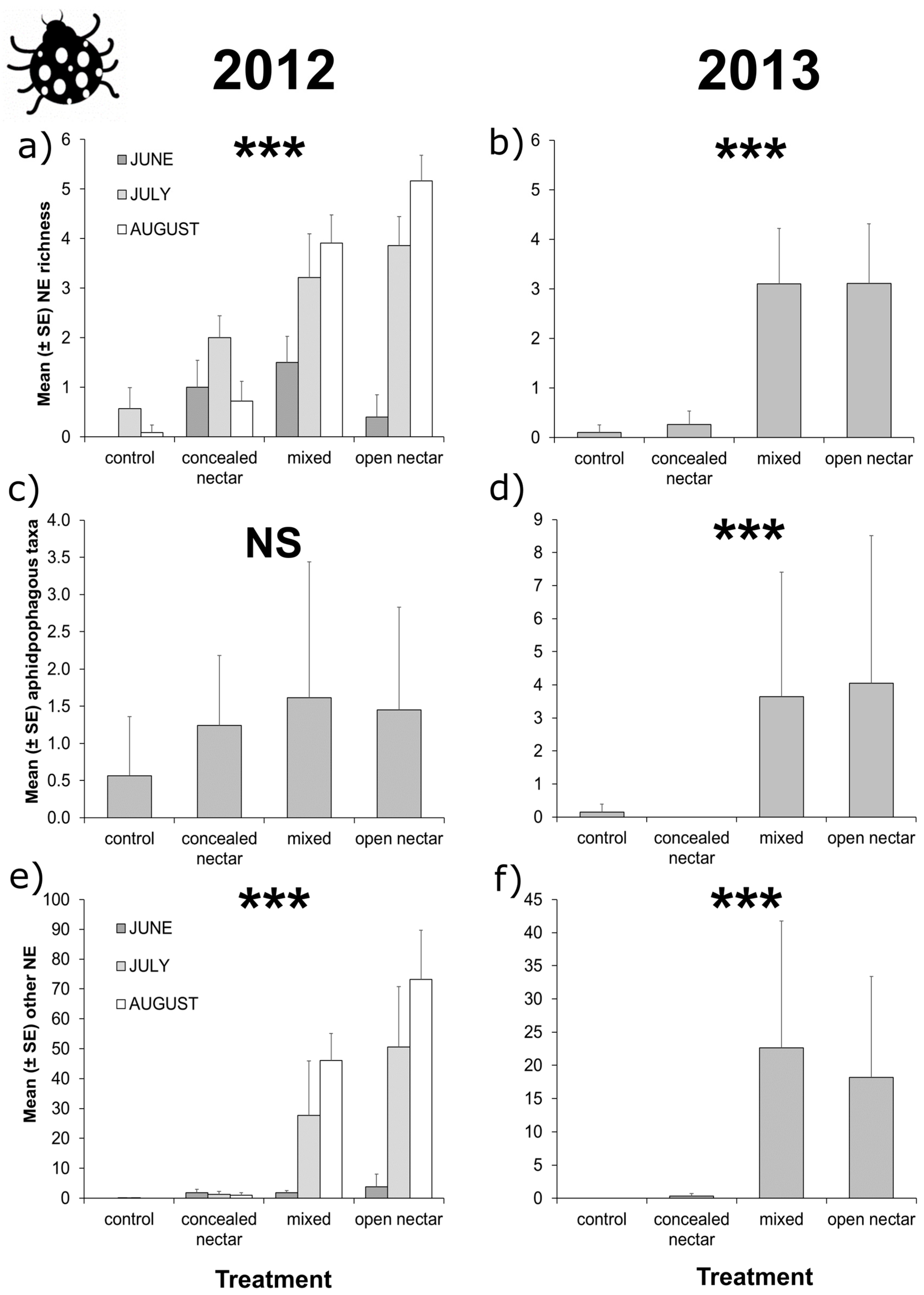
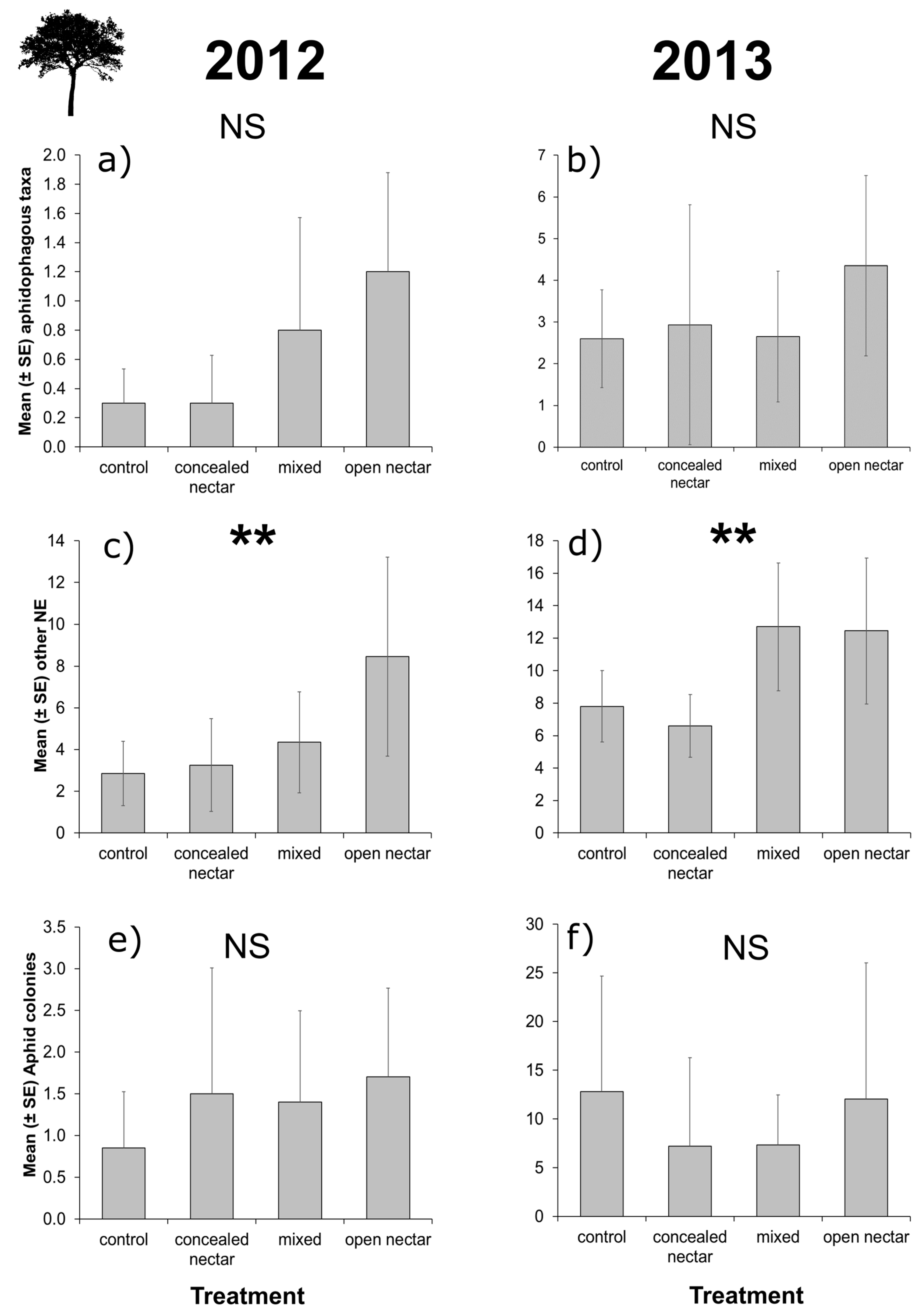
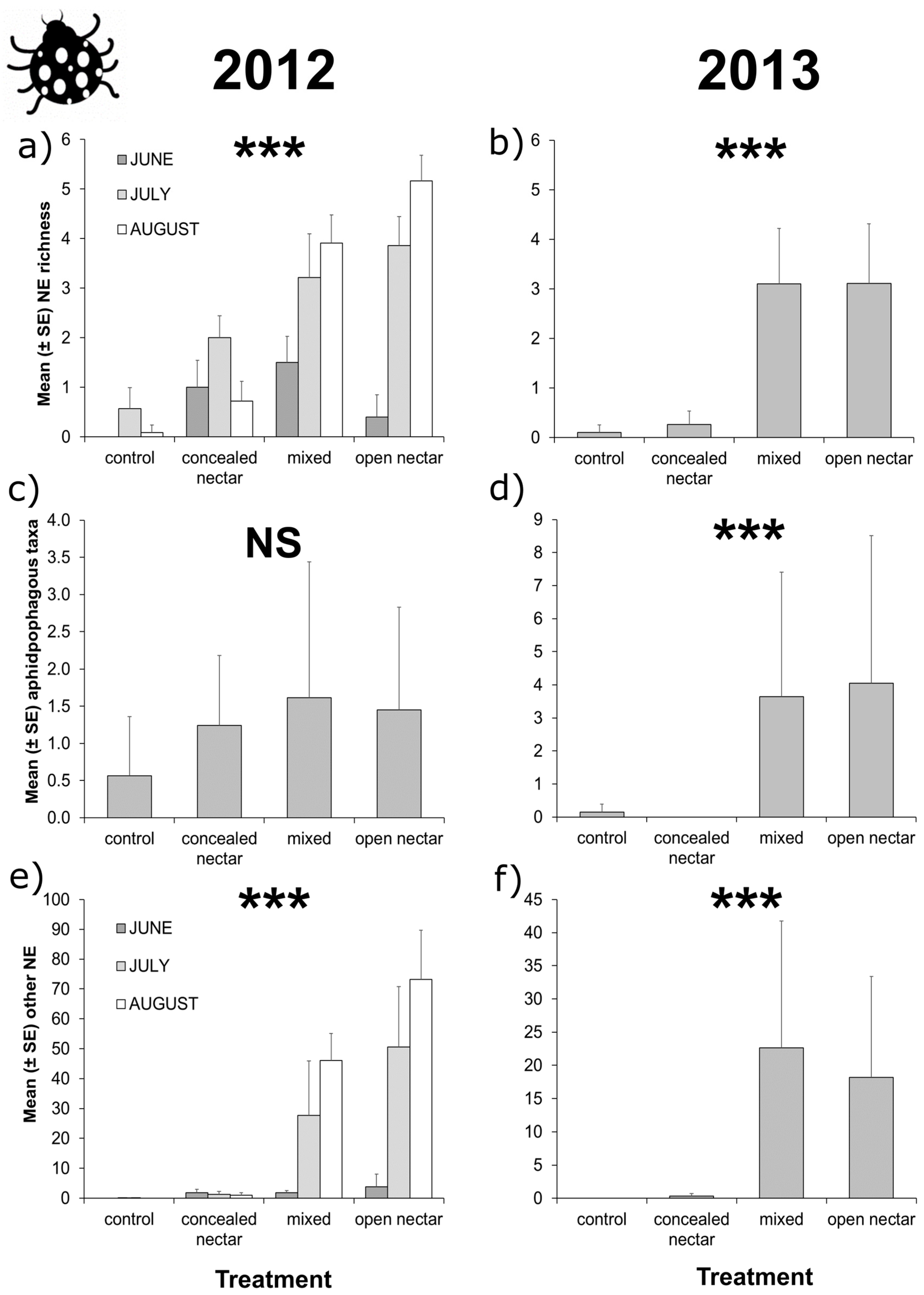
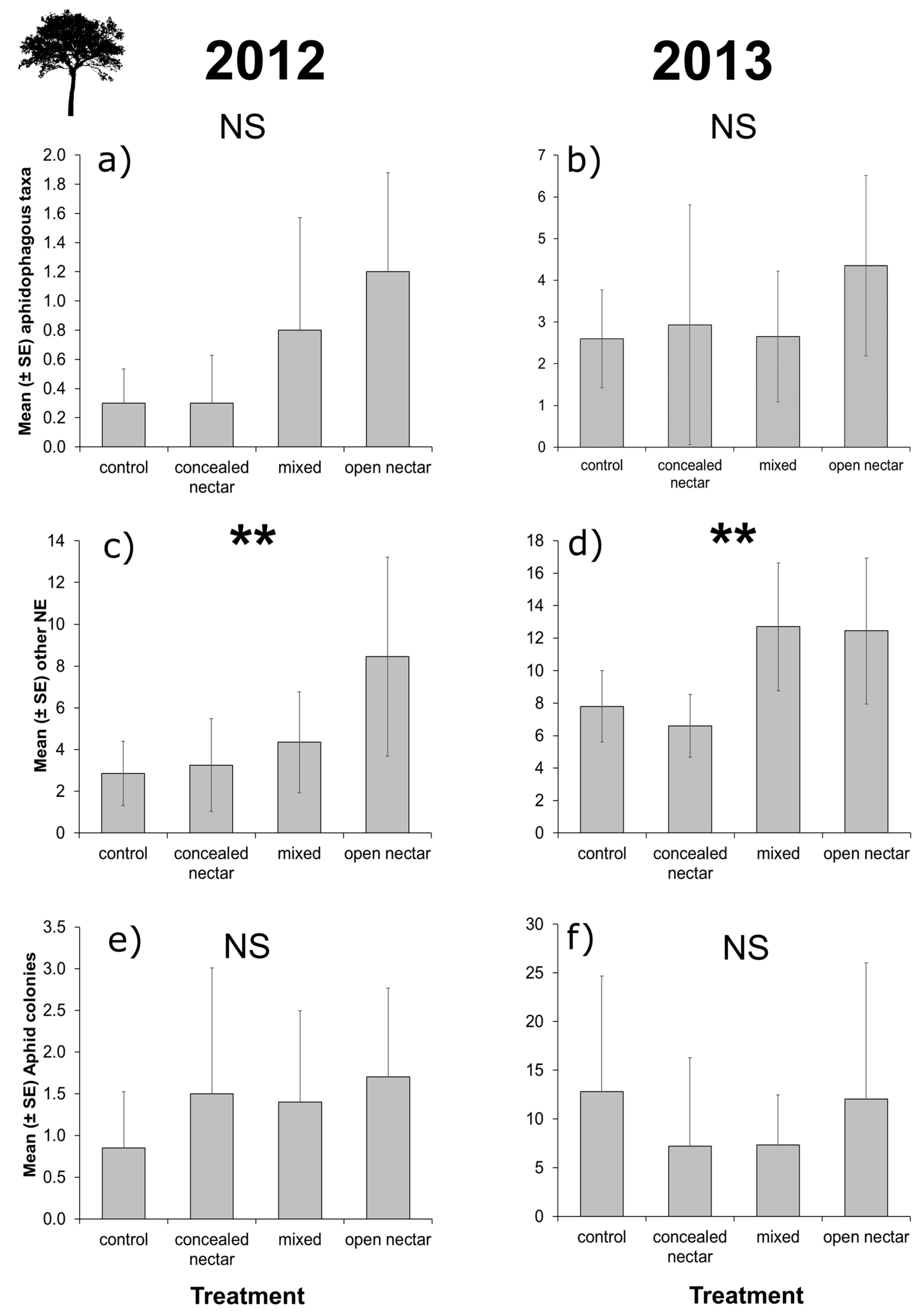
3.3. Apple Tree Surveys
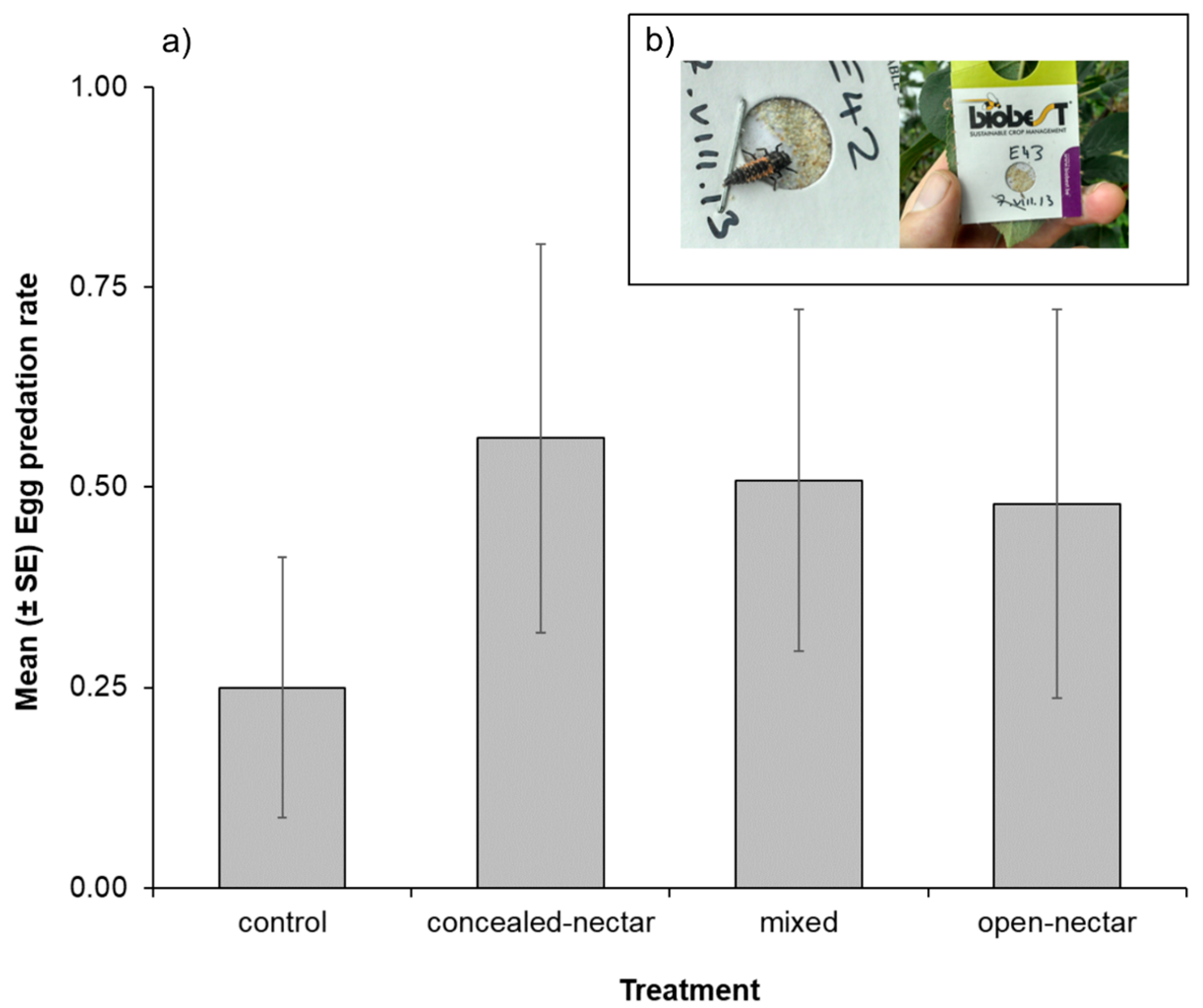
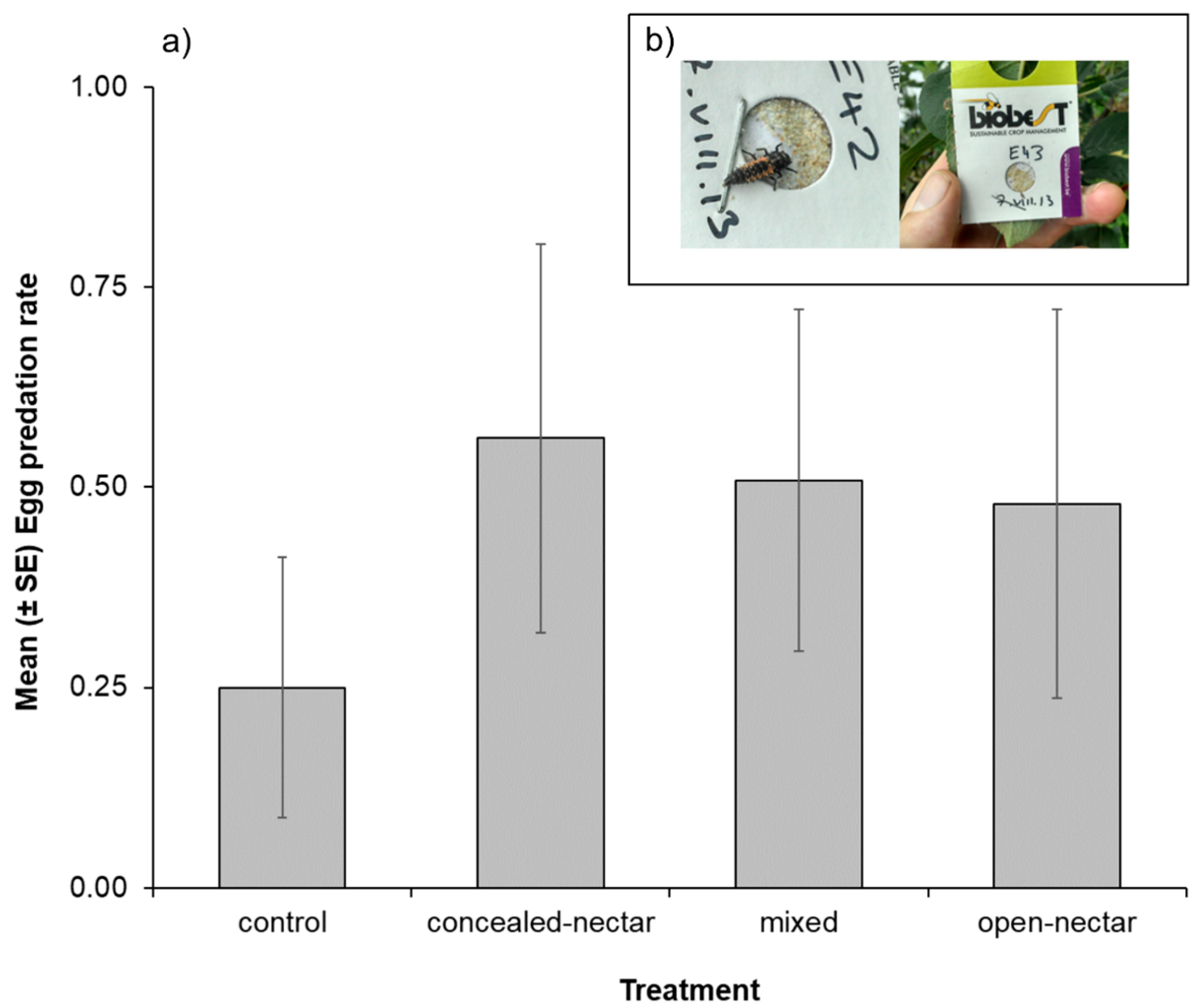
3.4. Sentinel Egg Cards
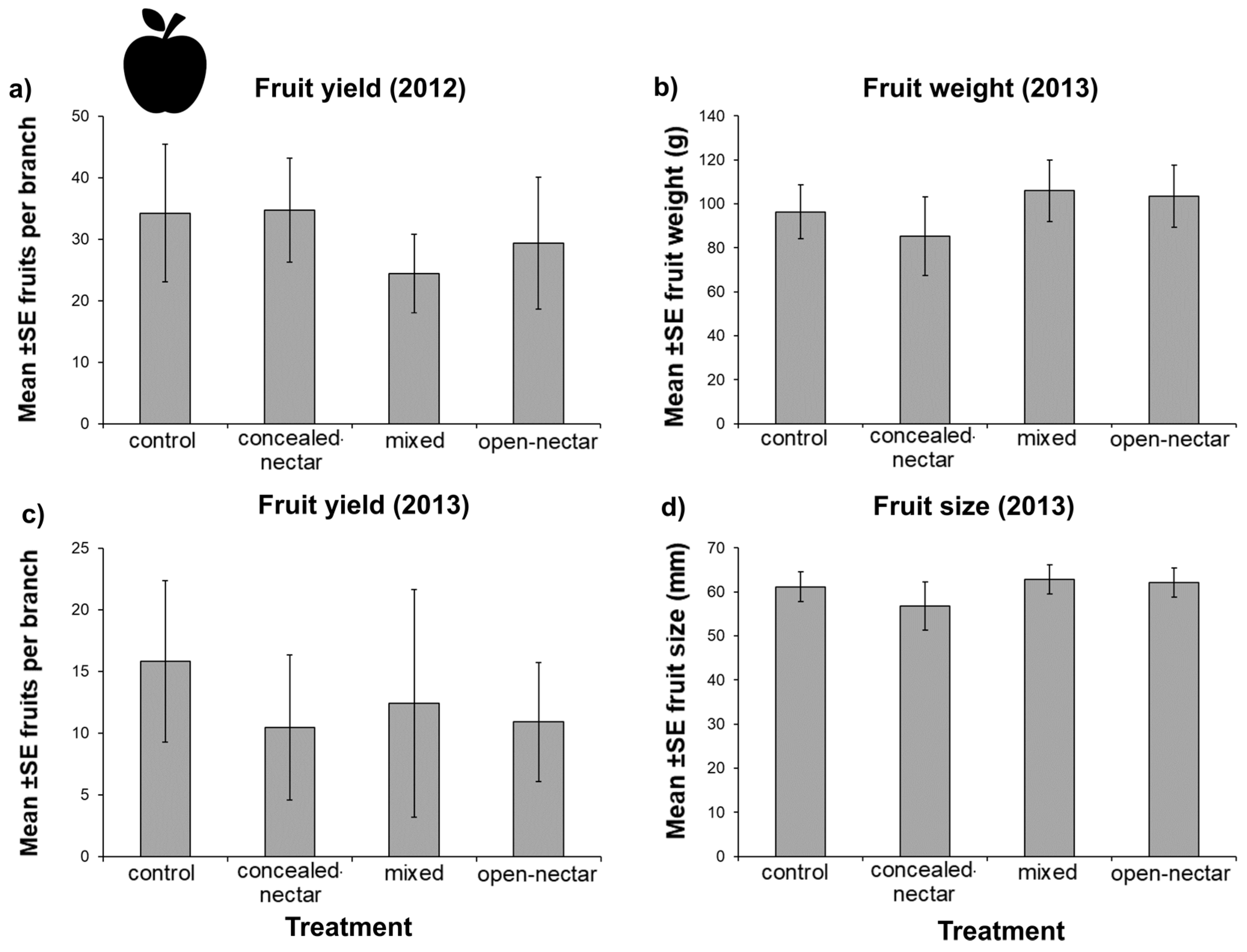
3.5. Fruit Yield
4. Discussion
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Godfray, H.C.J. Food and biodiversity. Science 2011, 333, 1231–1232. [Google Scholar] [CrossRef] [PubMed]
- Biesmeijer, J.C.; Roberts, S.P.M.; Reemer, M.; Ohlemüller, R.; Edwards, M.; Peeters, T.; Schaffers, A.P.; Potts, S.G.; Kleukers, R.; Thomas, C.D.; et al. Parallel Declines in pollinators and insect-pollinated plants in Britain and the Netherlands. Science 2006, 313, 351–354. [Google Scholar] [CrossRef] [PubMed]
- Evenson, R.E.; Gollin, D. Assessing the impact of the green revolution, 1960 to 2000. Science 2003, 300, 758–762. [Google Scholar] [CrossRef] [PubMed]
- Phalan, B.; Onial, M.; Balmford, A.; Green, R.E.; Godfray, H.C.J.; Tilman, D.; Green, R.E.; Cornell, S.J.; Scharlemann, J.P.W.; Balmford, A.; et al. Reconciling food production and biodiversity conservation: Land sharing and land sparing compared. Science 2011, 333, 1289–1291. [Google Scholar] [CrossRef] [PubMed]
- Kremen, C.; Williams, N.M.; Thorp, R.W. Crop pollination from native bees at risk from agricultural intensification. Proc. Natl. Acad. Sci. USA 2002, 99, 16812–16816. [Google Scholar] [CrossRef] [PubMed]
- Tscharntke, T.; Klein, A.M.; Kruess, A.; Steffan-Dewenter, I.; Thies, C. Landscape perspectives on agricultural intensification and biodiversity—Ecosystem service management. Ecol. Lett. 2005, 8, 857–874. [Google Scholar] [CrossRef]
- Garibaldi, L.A.; Gemmill-Herren, B.; D’Annolfo, R.; Graeub, B.E.; Cunningham, S.A.; Breeze, T.D. Farming Approaches for Greater Biodiversity, Livelihoods, and Food Security. Trends Ecol. Evol. 2016, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Bommarco, R.; Kleijn, D.; Potts, S.G. Ecological intensification: Harnessing ecosystem services for food security. Trends Ecol. Evol. 2013, 28, 230–238. [Google Scholar] [CrossRef] [PubMed]
- Losey, J.E.; Vaughan, M. The Economic Value of Ecological Services Provided by Insects. Bioscience 2006, 56, 311. [Google Scholar] [CrossRef]
- Klein, A.-M.; Vaissière, B.E.; Cane, J.H.; Steffan-Dewenter, I.; Cunningham, S.A.; Kremen, C.; Tscharntke, T. Importance of pollinators in changing landscapes for world crops. Proc. Biol. Sci. 2007, 274, 303–313. [Google Scholar] [CrossRef] [PubMed]
- Potts, S.G.; Imperatriz-Fonseca, V.; Ngo, H.T.; Aizen, M.A.; Biesmeijer, J.C.; Breeze, T.D.; Dicks, L.V.; Garibaldi, L.A.; Hill, R.; Settele, J.; et al. Safeguarding pollinators and their values to human well-being. Nature 2016, 540, 220–229. [Google Scholar] [CrossRef] [PubMed]
- Chaplin-Kramer, R.; O’Rourke, M.E.; Blitzer, E.J.; Kremen, C. A meta-analysis of crop pest and natural enemy response to landscape complexity. Ecol. Lett. 2011, 14. [Google Scholar] [CrossRef] [PubMed]
- Landis, D.A.; Wratten, S.D.; Gurr, G.M. Habitat Management to Conserve Natural Enemies of Arthropod Pests in Agriculture. Annu. Rev. Entomol. 2000, 45, 175–201. [Google Scholar] [CrossRef] [PubMed]
- Wratten, S.D.; Gillespie, M.; Decourtye, A.; Mader, E.; Desneux, N. Pollinator habitat enhancement: Benefits to other ecosystem services. Agric. Ecosyst. Environ. 2012, 159, 112–122. [Google Scholar] [CrossRef]
- Cross, J.; Fountain, M.; Markó, V.; Nagy, C. Arthropod ecosystem services in apple orchards and their economic benefits. Ecol. Entomol. 2015, 40, 82–96. [Google Scholar] [CrossRef]
- Geiger, F.; Wäckers, F.L.; Bianchi, F.J.J. A Hibernation of predatory arthropods in semi-natural habitats. BioControl 2009, 54, 529–535. [Google Scholar] [CrossRef]
- Wäckers, F.L.; Van Rijn, P.C.J. Pick and Mix: Selecting flowering plants to meet requirements of target biological control insects. In Biodiversity and Insect Pests; Gurr, G.M., Ed.; Wiley Blackwell: Hoboken, NJ, USA, 2012. [Google Scholar]
- Scheper, J.; Holzschuh, A.; Kuussaari, M.; Potts, S.G.; Rundlöf, M.; Smith, H.G.; Kleijn, D. Environmental factors driving the effectiveness of European agri-environmental measures in mitigating pollinator loss—A meta-analysis. Ecol. Lett. 2013, 16, 912–920. [Google Scholar] [CrossRef] [PubMed]
- Tschumi, M.; Albrecht, M.; Collatz, J.; Dubsky, V.; Entling, M.H.; Najar-Rodriguez, A.J.; Jacot, K. Tailored flower strips promote natural enemy biodiversity and pest control in potato crops. J. Appl. Ecol. 2016, 53, 1169–1176. [Google Scholar] [CrossRef]
- Wood, T.J.; Holland, J.M.; Hughes, W.O.H.; Goulson, D. Targeted agri-environment schemes significantly improve the population size of common farmland bumblebee species. Mol. Ecol. 2015, 24, 1668–1680. [Google Scholar] [CrossRef] [PubMed]
- Blaauw, B.R.; Isaacs, R. Flower plantings increase wild bee abundance and the pollination services provided to a pollination-dependent crop. J. Appl. Ecol. 2014, 51, 890–898. [Google Scholar] [CrossRef]
- Carvell, C.; Meek, W.R.; Pywell, R.F.; Goulson, D.; Nowakowski, M. Comparing the efficacy of agri-environment schemes to enhance bumble bee abundance and diversity on arable field margins. J. Appl. Ecol. 2007, 44, 29–40. [Google Scholar] [CrossRef]
- Steward, P.R.; Shackelford, G.; Carvalheiro, L.G.; Benton, T.G.; Garibaldi, L.A.; Sait, S.M. Pollination and biological control research: Are we neglecting two billion smallholders. Agric. Food Secur. 2014. [Google Scholar] [CrossRef]
- Shackelford, G.; Steward, P.R.; Benton, T.G.; Kunin, W.E.; Potts, S.G.; Biesmeijer, J.C.; Sait, S.M. Comparison of pollinators and natural enemies: A meta-analysis of landscape and local effects on abundance and richness in crops. Biol. Rev. 2013, 88, 1002–1021. [Google Scholar] [CrossRef] [PubMed]
- Campbell, A.J.; Biesmeijer, J.C.; Varma, V.; Wäckers, F.L. Realising multiple ecosystem services based on the response of three beneficial insect groups to floral traits and trait diversity. Basic Appl. Ecol. 2012, 13, 363–370. [Google Scholar] [CrossRef]
- Olson, D.M.; Wäckers, F.L. Management of field margins to maximize multiple ecological services. J. Appl. Ecol. 2007, 44, 13–21. [Google Scholar] [CrossRef]
- Isaacs, R.; Tuell, J.; Fiedler, A.; Gardiner, M.; Landis, D. Maximizing arthropod-mediated ecosystem services in agricultural landscapes: The role of native plants. Front. Ecol. Environ. 2009, 7, 196–203. [Google Scholar] [CrossRef]
- Gurr, G.; Wratten, S.D.; Landis, D.A.; You, M. Habitat management to suppress pest populations: Progress and prospects. Annu. Rev. Entomol. 2017, 62, 62–91. [Google Scholar] [CrossRef] [PubMed]
- Van Rijn, P.C.J.; Wäckers, F.L. Nectar accessibility determines fitness, flower choice and abundance of hoverflies that provide natural pest control. J. Appl. Ecol. 2016, 53, 925–933. [Google Scholar] [CrossRef]
- Wood, T.J.; Holland, J.M.; Goulson, D. Pollinator-friendly management does not increase the diversity of farmland bees and wasps. Biol. Conserv. 2015, 187, 120–126. [Google Scholar] [CrossRef]
- Wäckers, F.L. Assessing the suitability of flowering herbs as parasitoid food sources: Flower attractiveness and nectar accessibility. Biol. Control 2004, 29, 307–314. [Google Scholar] [CrossRef]
- Pontin, D.R.; Wade, M.R.; Kehrli, P.; Wratten, S.D. Attractiveness of single and multiple species flower patches to beneficial insects in agroecosystems. Ann. Appl. Biol. 2006, 148, 39–47. [Google Scholar] [CrossRef]
- Balzan, M.V.; Bocci, G.; Moonen, A.C. Utilisation of plant functional diversity in wildflower strips for the delivery of multiple agroecosystem services. Entomol. Exp. Appl. 2016, 158, 304–319. [Google Scholar] [CrossRef]
- Brown, M.W.; Mathews, C.R. Conservation biological control of rosy apple aphid, Dysaphis plantaginea (Passerini), in eastern north America. Environ. Entomol. 2007, 36, 1131–1139. [Google Scholar] [CrossRef]
- Wyss, E.; Villiger, M.; Müller-Schärer, H. The potential of three native insect predators to control the rosy apple aphid, Dysaphis plantaginea. BioControl 1999, 44, 171–182. [Google Scholar] [CrossRef]
- Dib, H.; Simon, S.; Sauphanor, B.; Capowiez, Y. The role of natural enemies on the population dynamics of the rosy apple aphid, Dysaphis plantaginea Passerini (Hemiptera: Aphididae) in organic apple orchards in south-eastern France. Biol. Control 2010, 55, 97–109. [Google Scholar] [CrossRef]
- Gontijo, L.M.; Beers, E.H.; Snyder, W.E. Flowers promote aphid suppression in apple orchards. Biol. Control 2013, 66, 8–15. [Google Scholar] [CrossRef]
- Wyss, E. The effects of artificial weed strips on diversity and abundance of the arthropod fauna in a Swiss experimental apple orchard. Agric. Ecosyst. Environ. 1996, 60, 47–59. [Google Scholar] [CrossRef]
- Rosa García, R.; Miñarro, M. Role of floral resources in the conservation of pollinator communities in cider-apple orchards. Agric. Ecosyst. Environ. 2014, 183, 118–126. [Google Scholar] [CrossRef]
- Campbell, A.J.; Wilby, A.; Sutton, P.; Wäckers, F.L. Do sown flower strips boost wild pollinator abundance and pollination services in a spring-flowering crop? A case study from UK cider apple orchards. Agric. Ecosyst. Environ. 2017, 239, 20–29. [Google Scholar] [CrossRef]
- Blitzer, E.J.; Gibbs, J.; Park, M.G.; Danforth, B.N. Pollination services for apple are dependent on diverse wild bee communities. Agric. Ecosyst. Environ. 2016, 221, 1–7. [Google Scholar] [CrossRef]
- Garratt, M.P.D.; Breeze, T.D.; Jenner, N.; Polce, C.; Biesmeijer, J.C.; Potts, S.G. Avoiding a bad apple: Insect pollination enhances fruit quality and economic value. Agric. Ecosyst. Environ. 2014, 184, 34–40. [Google Scholar] [CrossRef] [PubMed]
- Russo, L.; Debarros, N.; Yang, S.; Shea, K.; Mortensen, D. Supporting crop pollinators with floral resources: Network-based phenological matching. Ecol. Evol. 2013, 3, 3125–3140. [Google Scholar] [CrossRef] [PubMed]
- Fournier, D.A.; Skaug, H.J.; Ancheta, J.; Ianelli, J.; Magnusson, A.; Maunder, M.N.; Nielsen, A.; Sibert, J. AD Model Builder: Using automatic differentiation for statistical inference of highly parameterized complex nonlinear models. Optim. Methods Softw. 2012, 27, 233–249. [Google Scholar] [CrossRef] [Green Version]
- Pinheiro, J.; Bates, D.; DebRoy, S.; Sarkar, D. nlme: Linear and Nonlinear Mixed Effects Models, R Package version 3.1-131; 2016. Available online: https://CRAN.R-project.org/package=nlme (accessed on 12 September 2017).
- Zuur, A.F.; Ieno, E.N.; Walker, N.J.; Saveliev, A.A.; Smith, G. Mixed Effects Models and Extensions in Ecology with R; Springer-Verlag: New York, NY, USA, 2009. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; The R Foundation for Statistical Computing: Vienna, Austria, 2015; Volume 55, pp. 275–286. ISBN 3-900051-07-0. [Google Scholar]
- Junker, R.R.; Blüthgen, N.; Brehm, T.; Binkenstein, J.; Paulus, J.; Martin Schaefer, H.; Stang, M. Specialization on traits as basis for the niche-breadth of flower visitors and as structuring mechanism of ecological networks. Funct. Ecol. 2013, 27, 329–341. [Google Scholar] [CrossRef]
- Bartomeus, I.; Ascher, J.S.; Gibbs, J.; Danforth, B.N.; Wagner, D.L.; Hedtke, S.M. Historical changes in northeastern US bee pollinators related to shared ecological traits. Proc. Natl. Acad. Sci. USA 2013, 110, 4656–4660. [Google Scholar] [CrossRef] [PubMed]
- Winkler, K.; Wäckers, F.L.; Termorshuizen, A.J.; Lenteren, J.C. Assessing risks and benefits of floral supplements in conservation biological control. BioControl 2010, 55, 719–727. [Google Scholar] [CrossRef]
- Blaauw, B.R.; Isaacs, R. Larger wildflower plantings increase natural enemy density, diversity, and biological control of sentinel prey, without increasing herbivore density. Ecol. Entomol. 2012, 37, 386–394. [Google Scholar] [CrossRef]
- Power, A.G. Ecosystem services and agriculture: Tradeoffs and synergies. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2010, 365, 2959–2971. [Google Scholar] [CrossRef] [PubMed]
- Bugg, R.L.; Waddington, C. Using cover crops to manage arthropod pests of orchards: A review. Agric. Ecosyst. Environ. 1994, 50, 11–28. [Google Scholar] [CrossRef]
- Bianchi, F.J.J.A.; Wäckers, F.L. Effects of flower attractiveness and nectar availability in field margins on biological control by parasitoids. Biol. Control 2008, 46, 400–408. [Google Scholar] [CrossRef]
- Simon, S.; Bouvier, J.-C.; Debras, J.-F.; Sauphanor, B. Biodiversity and pest management in orchard systems. A review. Agron. Sustain. Dev. 2010, 30, 139–152. [Google Scholar] [CrossRef]
- Lundgren, J.G. Nutritional aspects of non-prey foods in the life histories of predaceous Coccinellidae. Biol. Control 2009, 51, 294–305. [Google Scholar] [CrossRef]
- Spellman, B.; Brown, M.W.; Mathews, C.R. Effect of floral and extrafloral resources on predation of Aphis spiraecola by Harmonia axyridis on apple. BioControl 2006, 51, 715–724. [Google Scholar] [CrossRef]
- Kuhlmann, U.; Carl, K.P.; Mills, N.J. Quantifying the impact of insect predators and parasitoids on populations of the apple ermine moth, Yponomeuta malinellus (Lepidoptera: Yponomeutidae), in Europe. Bull. Entomol. Res. 1998, 88, 165. [Google Scholar] [CrossRef]
- Bostanian, N.J.; Goulet, H.; O’Hara, J.; Masner, L.; Racette, G. Towards Insecticide Free Apple Orchards: Flowering Plants to Attract Beneficial Arthropods. Biocontrol Sci. Technol. 2004, 14, 25–37. [Google Scholar] [CrossRef]
- Bos, M.M.; Veddeler, D.; Bogdanski, A.K.; Klein, A.M.; Tscharntke, T.; Steffan-Dewenter, I.; Tylianakis, J.M. Caveats to quantifying ecosystem services: Fruit abortion blurs benefits from crop pollination. Ecol. Appl. 2007, 17, 1841–1849. [Google Scholar] [CrossRef] [PubMed]
- Saunders, M.E. Ecosystem services in agriculture: Understanding the multifunctional role of invertebrates. Agric. For. Entomol. 2017. [Google Scholar] [CrossRef]
- Klein, A.-M.; Hendrix, S.D.; Clough, Y.; Scofield, A.; Kremen, C. Interacting effects of pollination, water and nutrients on fruit tree performance. Plant Biol. (Stuttgart) 2014, 1–8. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 2012 | 2013 | ||||||
|---|---|---|---|---|---|---|---|
| Within Flower Strip | Within Flower Strip | ||||||
| Response | Predictor | d.f. | LRT | p | d.f. | LRT | p |
| Pollinators | |||||||
| Richness | Treatment | 3 | 34.23 | <0.001 | 3 | 17.44 | 0.001 |
| Month | 2 | 5.63 | 0.060 | 2 | 9.13 | 0.010 | |
| T × M | 6 | 9.67 | 0.139 | 6 | 7.72 | 0.259 | |
| Eusocial bees | Treatment | 3 | 22.51 | <0.001 | |||
| Month | 2 | 18.37 | <0.001 | ||||
| T × M | 6 | 15.34 | 0.018 | 6 | 6.79 | 0.341 | |
| Solitary bees | Treatment | 3 | 13.96 | 0.003 | 3 | 7.02 | 0.071 |
| Month | 2 | 8.98 | 0.011 | 2 | 10.79 | 0.005 | |
| T × M | 6 | 8.49 | 0.204 | 6 | 4.85 | 0.564 | |
| Natural enemies | |||||||
| Richness | Treatment | 3 | 35.22 | <0.001 | |||
| Month | 2 | 23.14 | <0.001 | ||||
| T × M | 6 | 14.51 | 0.024 | 6 | 3.91 | 0.689 | |
| Aphidophagous taxa | Treatment | 3 | 3.27 | 0.352 | 3 | 18.47 | <0.001 |
| Month | 2 | 21.46 | <0.001 | 2 | 24.82 | <0.001 | |
| T × M | 6 | 2.27 | 0.893 | 6 | 9.53 | 0.146 | |
| Other taxa | Treatment | 3 | 28.00 | <0.001 | |||
| Month | 2 | 8.42 | 0.015 | ||||
| T × M | 6 | 40.02 | <0.001 | 6 | 8.74 | 0.189 | |
| 2012 | 2013 | ||||||
|---|---|---|---|---|---|---|---|
| Within Crop | Within Crop | ||||||
| Response | Predictor | d.f. | LRT | p | d.f. | LRT | p |
| Natural enemies | |||||||
| Richness | Treatment | 3 | 2.40 | 0.493 | 3 | 2.65 | 0.449 |
| Month | 2 | 34.80 | <0.001 | 2 | 0.60 | 0.742 | |
| T × M | 6 | 2.32 | 0.888 | 6 | 1.09 | 0.982 | |
| Aphidophagous taxa | Treatment | 3 | 6.35 | 0.096 | 3 | 1.93 | 0.587 |
| Month | 2 | 18.29 | <0.001 | 2 | 5.61 | 0.060 | |
| T × M | 6 | 6.16 | 0.406 | 6 | 11.25 | 0.081 | |
| Other taxa | Treatment | 3 | 13.87 | 0.003 | 3 | 12.77 | 0.005 |
| Month | 2 | 83.47 | <0.001 | 2 | 3.33 | 0.189 | |
| T × M | 6 | 5.01 | 0.543 | 6 | 7.76 | 0.256 | |
| Pest control | |||||||
| Aphid colonies | Treatment | 3 | 1.54 | 0.672 | 3 | 3.88 | 0.275 |
| Month | 2 | 18.97 | <0.001 | 2 | 55.28 | <0.001 | |
| T × M | 6 | 9.37 | 0.154 | 6 | 5.39 | 0.495 | |
| Egg cards | Treatment | - | - | - | 3 | 9.54 | 0.023 |
| Month * | - | - | - | 3 | 77.71 | <0.001 | |
| T × M | - | - | - | 9 | 15.58 | 0.076 | |
| Treatment | |||
|---|---|---|---|
| Response Variable | LRT | d.f. | p |
| 2012 | |||
| Fruit number | 7.01 | 3 | 0.071 |
| 2013 | |||
| Fruit number | 4.65 | 3 | 0.200 |
| Fruit size (mm) | 5.09 | 3 | 0.165 |
| Fruit weight (g) | 5.94 | 3 | 0.114 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Campbell, A.J.; Wilby, A.; Sutton, P.; Wäckers, F. Getting More Power from Your Flowers: Multi-Functional Flower Strips Enhance Pollinators and Pest Control Agents in Apple Orchards. Insects 2017, 8, 101. https://doi.org/10.3390/insects8030101
Campbell AJ, Wilby A, Sutton P, Wäckers F. Getting More Power from Your Flowers: Multi-Functional Flower Strips Enhance Pollinators and Pest Control Agents in Apple Orchards. Insects. 2017; 8(3):101. https://doi.org/10.3390/insects8030101
Chicago/Turabian StyleCampbell, Alistair John, Andrew Wilby, Peter Sutton, and Felix Wäckers. 2017. "Getting More Power from Your Flowers: Multi-Functional Flower Strips Enhance Pollinators and Pest Control Agents in Apple Orchards" Insects 8, no. 3: 101. https://doi.org/10.3390/insects8030101