1. Introduction
Deforestation in Venezuela has greatly increased in the last few decades, with about 326,000 hectares of native forests being destroyed per year for agricultural and livestock purposes with the aim of helping to guarantee the “food and agriculture security” of the country [
1]. However, the expectations for production have not been realized, and Venezuela is now suffering the consequences of deforestation, such as more frequent and more severe landslides and a substantial increase in emissions of greenhouse gases [
2]. Consequently, there is an urgent need to promote a sustainable farming model that ensures food production and forest conservation in an economically profitable framework for rural communities. Agroforestry, i.e., the combination of trees with crops or livestock [
3] may represent an opportunity to achieve these goals.
Cacao crops have been traditionally cultivated under shade conditions, but presently there is an active debate amongst both scientists and farmers as to whether the cacao crop should be placed under shade conditions or not [
4,
5,
6,
7,
8]. Physiological traits such as the photosynthetic rates of cacao, which are saturated at low photosynthetic photon flux densities (400–600 μmol photons m
−2 s
−1), and low leaf stomatal conductance (g
s) help explain why cacao is not only a shade-tolerant species [
9,
10,
11,
12], but also why it benefits by growing under the shade of other trees [
13]. However, monocultures of new sun-grown hybrids are spreading in many tropical countries, especially in those regions where radiation hours and intensity are low [
14], resulting in high short-term productivity but with possible long-term implications with regard to the depletion of soils and loss of biodiversity, ultimately affecting the plantation yield [
5,
6]. Shade varieties may therefore have more advantages than disadvantages [
4,
7], as long as shade trees contribute additional benefits [
15,
16], do not compete with cacao for light or nutrients, and are properly managed.
Insects are known to be important in almost all ecological processes within terrestrial ecosystems [
17], and their relationship with those ecological processes needs to be established with regard to cultivated lands. This includes both the ecosystem services they provide, such as biological control [
18] and pollination [
19], and the ecosystem disservice they cause, such as physical and physiological damage produced by herbivorous insects [
20,
21,
22]. However, the implications that different light regimes or the species identity may have on the insect assemblages have been poorly studied. Insects attacking herbivorous insects, such as parasitoid Hymenoptera and some predator beetles, while helping to balance the agroecosystem by natural control, are more sensitive to disturbances than their hosts [
23,
24]. However, it is unclear whether shade trees in cacao agroforestry systems induce pest outbreaks or provide a useful ecosystem service by increasing their natural enemies’ populations [
25,
26,
27,
28].
The Venezuelan Criollo cacao cultivars (
Theobroma cacao L.) have exquisite flavours and aromas [
29,
30], and are highly appreciated by the chocolate industry. Nevertheless, they have low fruit production and are highly sensitive to pests and diseases, which make them of little interest economically to the local farmers. Hence, these cultivars have been replaced by more resistant Trinitario- and Forastero-type cultivars and as a consequence, Criollo cacaos are endangered. However, in Venezuela, since the beginning of 2000, several national projects have been promoted in order to rescue the high-yielding Criollo cultivars by increasing their production and gradually introducing them on cacao plantations. Managing the cacao cultures in order to favour pest natural control by favouring the occurrence of their potential natural enemies may also help to recover these traditional cultivars [
27,
31].
In the present study we assessed the insect assemblages associated to four native timber trees in a cacao agroforestry system by using two indicator groups: Coleoptera and parasitoid Hymenoptera, since both groups have been demonstrated to be very sensitive to changes in the environment [
32,
33,
34,
35]. We used a higher-level taxa for the tests, i.e., family level. Family richness can be used as a surrogate for species richness, since it is closely correlated to species richness in some groups [
36,
37]. The aims of this study were: (1) to assess insect richness and abundance at different taxonomic levels in the agroforestry system of Criollo cacao cultivars with native timber trees; and (2) to evaluate the influence of the native timber trees and the cacao cultivars over the insect assemblages associated.
3. Results
We collected 49,538 individuals, belonging to seven orders: Orthoptera, Lepidoptera, Hymenoptera, Hemiptera, Diptera, Coleoptera, and Dictyoptera. The orders Hymenoptera, Diptera and Hemiptera were the most abundant orders, comprising 95% of the total insects with 19,118, 13,363 and 14,550 individuals collected, respectively.
We found 23 families of parasitoid Hymenoptera (
Table 2) and 24 of Coleoptera (
Table 3). In
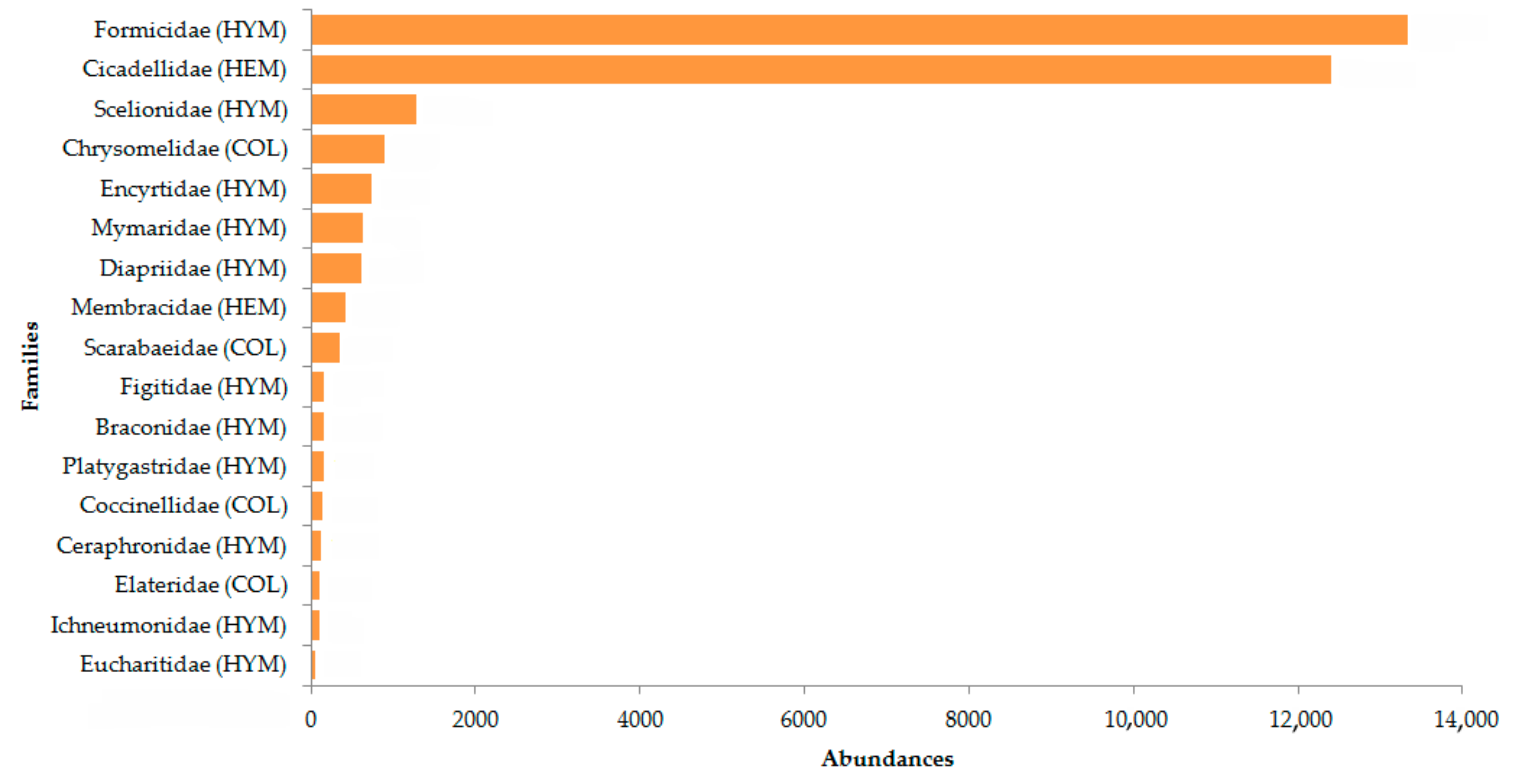
Figure 2, the most abundant families (
n > 50 individuals) are shown. Formicidae and Cicadellidae were included in the count because they were extremely abundant within the samples, representing more than 80%.
Influence of Native Timber Tree Species and Cacao Cultivars on Entomofauna
No significant differences were found between total abundance nor richness of orders occurring associated to every timber tree (F = 0.119,
p = 0.949 and F = 0.974,
p = 0.409, respectively), nor to every cacao cultivar (F = 0.483,
p = 0.695 and F = 0.487,
p = 0.692, respectively), nor the interaction of both (F = 0.909,
p = 0.522, and F = 0.812,
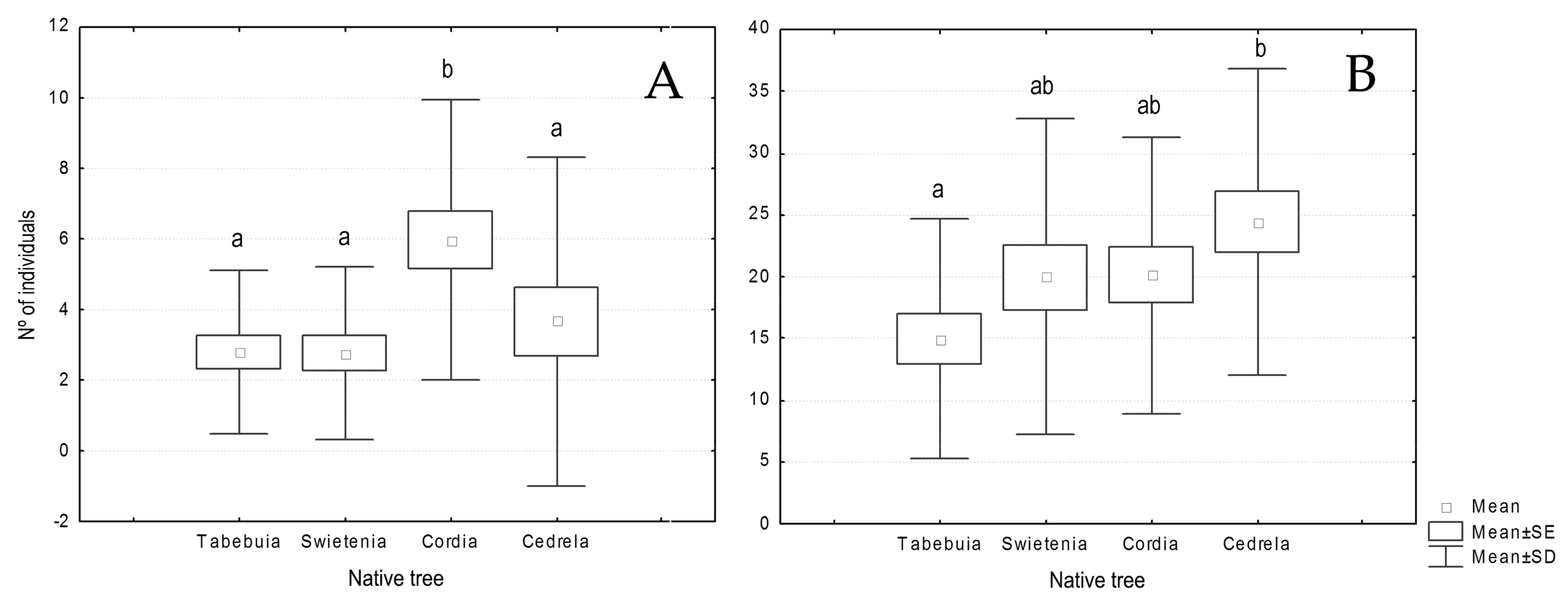
p = 0.607). However, when analysing every order separately, we found that Lepidoptera and Coleoptera abundances were influenced by the tree species, and Lepidoptera by the cacao cultivar also. While Lepidoptera was significantly (F = 4.77,
p = 0.004) more abundant in
Cordia plots than in
Tabebuia and
Swietenia (
Figure 3A), Coleoptera was significantly (F = 2.598,
p = 0.050) more present in
Cedrela than in
Tabebuia plots (
Figure 3B). Regarding the cacao cultivar, Lepidoptera was significantly more abundant in Porcelana plots (F = 4.066,
p = 0.009).
Regarding family richness (including all Coleoptera and parasitoid Hymenoptera families), no significant differences between the different native trees were found (F = 0.751, p = 0.525), nor with the interaction of timber tree and cacao cultivar (F = 0.868, p = 0.557), but the Merideño cultivar had a significantly higher family richness than Guasare (F = 3.694, p = 0.015).
The PERMANOVA analysis showed significant differences in both Hymenoptera and Coleoptera assemblages associated to the native timber trees (F = 1.792,
p = 0.001 and F = 1.728,
p = 0.024, respectively). When comparing cacao cultivar, there were significant effects only on the Coleoptera assemblages (F = 1.995,
p = 0.005), and none of the interactions between cacao cultivars and native trees were significant, indicating the independence of these factors. Considering parasitoid Hymenoptera family assemblages, those occurring in
Cedrela plots were significantly different to the
Swietenia and
Tabebuia ones, as well as those in
Cordia when compared to those in
Swietenia (
Table 4). The families Diapriidae, Scelionidae, Encyrtidae, Mymaridae and Platygastridae explained about 50% of these differences: they were relatively less abundant in
Cedrela than in the other trees, whilst Diapriidae and Encyrtidae were more abundant in
Cordia than in
Swietenia (see
Supplementary Materials Table S1); however, only the families Eulophidae and Chalcididae were considered significant as indicator species for
Cordia and
Swietenia trees species, respectively (
Table 5).
On the other hand, considering Coleoptera families’ assemblages, significant differences were found between
Tabebuia and
Cordia (
Table 4). Chrysomelidae, Scarabaeidae, Coccinellidae, Elateridae, and Curculionidae contributed the most, explaining 62% of the differences. Most families were more abundant in
Cordia plots than in those with
Tabebuia, while only Scarabaeidae and Curculionidae were more present in
Tabebuia than in
Cordia (see
Supplementary Materials Table S2). The family Lycidae was a significant indicator for
Cordia trees, as was Biphyllidae for
Cedrela (
Table 6). There were also significant differences in the Coleoptera assemblages amongst cacao cultivars: those occurring in Guasare plots were different to Lobatera and Merideño. Chrysomelidae, Scarabaeidae, Coccinellidae, Elateridae, Scydnaenidae, Curculionidae, and Staphylinidae contributed to 70–77% of the total dissimilarity, they were less abundant in Guasare than in the other cultivars (see
Supplementary Materials Table S3). Scydmaenidae and Scarabaeidae turned out to be significant indicator families for Lobatera and Merideño, respectively (
Table 7).
4. Discussion
The entomofauna associated with the agroforestry system studied included the major insect orders, with samples being dominated by Hymenoptera, Hemiptera, and Diptera. Ants (Formicidae) and leafhoppers (Cicadellidae) were by far the most abundant groups. The dominance of these groups was influenced by the type of trap used, since it was placed on the ground and by consequence it mainly collected ground insects or those living in the herbaceous and litter layers. Coloured pan traps are considered a standardized methodology for comparison of Hymenoptera diversity amongst sites [
45,
46] and for collecting some Coleoptera families [
47]. The high abundance of ants may be favoured by the presence of
Cordia trees, since a close symbiotic relationship has been described between ants and some
Cordia species [
48]. Jaimez et al. [
20] found that
Cordia trees were not attacked by insects in this plantation, while
Tabebuia and
Swietenia were highly susceptible to ant attacks during the first two years of establishment. In our study, Formicidae abundance was not significantly higher in
Cordia plots, although that may be explained by the young age of the trees, since one year after sampling this relationship was more noticeable [
20]. Besides Formicidae and Cicadellidae, yellow traps, as shown in this study, are effective for collecting Proctotrupoidea and Platygastroidea [
49], as well as some Chalcidoidea families [
50], while other insect traps commonly used, like malaise traps, usually collect greater amounts of Ichneumonoidea [
38,
45,
49,
50,
51].
Concerning Hymenopteran parasitoids, we found 23 of the 44 Neotropical families belonging to the superfamilies mentioned above [
52]. This is of low richness compared to the 33 families of parasitoid Hymenoptera found in different traditional cacao plantations of the same Venezuelan state [
53], but significantly higher than the 11 Hymenoptera families collected in a monospecific silvopastoral system of
Brachiaria decumbens in Minas Gerais [
50]. Furthermore, the richness of parasitoid families recorded in this study was similar to that found in some Amazonian forest reserves [
51], and their relative abundances are consistent with those found by [
45] by means of yellow pan traps at the edge of a rainforest in Sulawesi.
Many of the hymenopteran families we found in this survey, specifically Ichneumonidae, Braconidae, Eurytomidae, Chalcididae, Trichogrammatidae, Encyrtidae, Aphelinidae, Eulophidae, Eupelmidae, and Signiphoridae, have been recorded as parasitoids of phytophagous species that feed on cacao [
54]. Although we did not identify the individuals to species level, both abundance and family richness of parasitoid Hymenoptera are good predictors of the richness of other taxa [
55]. The occurrence of these families within the farm, while indicating relatively high overall biodiversity, may also be contributing to long-term sustainability due to the potential regulation of phytophagous insect assemblages, since parasitoids can keep pest populations at low densities [
56] while generating responses at a radius of hundreds of meters [
57]. However, the high incidence of
Hypsipyla grandella (Zeller) attacking mainly
Cedrela in the Judibana plantation during the early stages [
20], which is one of the main pests of
Cedrela and
Swietenia in Venezuela [
58], may indicate the absence of a suitable population of their native parasitoids. One Eulophidae species,
Palmistichus elaeisis (Delvare & LaSalle), has been reported as parasitizing
H. grandella pupae in Brazil [
59], and actually this family was more abundant in the
Cedrela plots than in others, but it may not be present enough. In
Tabebuia, the pyralid
Eulepte gastralis (Gn.) may be a severe defoliator in young plantations, especially when sowing density is high [
60], but in 2–3 years plantations, leaf-rolling insects tend to be the most harmful [
61]; none of them were reported in Judibana plantation, or at least not being economically important [
20].
We also found a high richness of Coleoptera families, which represents a high diversity of trophic guilds too, contributing to the maintenance of several ecosystem services (e.g., natural pest control, nutrients recycling). The most abundant families, i.e., Chrysomelidae and Scarabaeidae, coincide with those found on the lower canopy level of some cacao agroforestry systems in Indonesia [
62]. Some species of chrysomelids have been recorded from cocoa plants [
63], so it is important to make identifications at genus or species level in this family. Besides, the relatively low abundance of Scolytinae subfamily suggests a more or less healthy farm, compared for example to a sampling made in another Venezuelan Criollo cacao plantation, where about 20 Scolytinae were collected per trap [
22]. This group may constitute one of the most important pests in cacao by helping to spread many phytopathogenic fungi [
64,
65], which have been recorded as the main concern for cacao farmers in Mérida plantations [
22] and have also been associated with the shade trees of cacao agroforestry systems, such as
Swietenia and
Tabebuia [
66]. However, the lack of complementary collecting methods, such as alcohol traps, light traps, and direct capture, in addition to the seasonal sampling, may be affecting this result [
66]. The occurrence of families such as Coccinellidae, Staphylinidae, and Carabidae indicates a community of generalist predator beetles that could be effective at controlling populations of herbivores such as Chrysomelidae, Scarabaeidae, and Elateridae, whereas the low abundance of bark beetle predators such as Histeridae could be related to the low abundance of prey [
67].
The type of timber tree shade may determine the presence or absence of determined species due to effects on thermoregulation [
68]. A moderate shade seems to benefit some insects, like Scarabaeidae dung beetles or parasitic wasps [
26], but implications of shade management over Coleoptera diversity are unclear [
62]. In the study area, Coleoptera were significantly more abundant in
Cedrela than in
Tabebuia, both with similar shade cast. It seems that, independently of the shade cast, the identity of the trees do determine Coleoptera preferences [
69], and may influence the preference of a determined herbivorous pest to an herbaceous host plant [
70].
Cordia timber species have been recommended for coffee plantations because of their potentially sustainable output of timber [
16,
71].
Cedrela may constitute a suitable option as a shade tree in coffee plantations, provided that it does not compete with coffee plants [
72], although the survival rate may be low during the first years after plantation [
73].
Cordia and
Cedrela showed the best growing rates during the first three years in the studied plantation [
20,
74], and gathered more abundant and significantly different insect assemblages according to the present work. Besides, their shade cast is consistent with Beer’s recommendation [
4]: a “regular mottled shade pattern” in order to not reduce the light quality that would be available for the cacao trees.
The limited relationship between the native timber trees and the characteristics of the insect assemblages tested in this study may indicate that the local habitat variables do not have a great impact on these groups with relatively high mobility. Instead, landscape variables may have more weight to explain their diversity and abundance, as shown for different insect groups [
35,
75,
76,
77]. Mixed-species plantations of timber trees may reduce the herbivore damage [
78], although this will depend on the particular pest species, since mixed plantations may favour or be detrimental to them by the opposite resource concentration and dilution effects, as seen for
Tabebuia plantations [
78]. Furthermore, the presence of non-host herbivores may reduce the efficiency of several natural enemies by increasing the time they have to spend to find their specific hosts [
79].
Despite the limitations of the sampling design, this study gives a general picture of the overall insect diversity associated with some of the timber species that may be used as shade trees in cacao plantations. The trap type we used may show a bias in the sampled groups, so further studies should be done in order to know which species are directly related to the timber species and how they may effectively help to improve productivity of cacao plantations, and, as a consequence, ensure the long-term livelihoods of cacao farmers.





{kind=link}
{kind=link}
{kind=link}