Rapid Cold Hardening Confers a Transient Increase in Low Temperature Survival in Diapausing Chilo suppressalis Larvae


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Insects and Diapause Induction
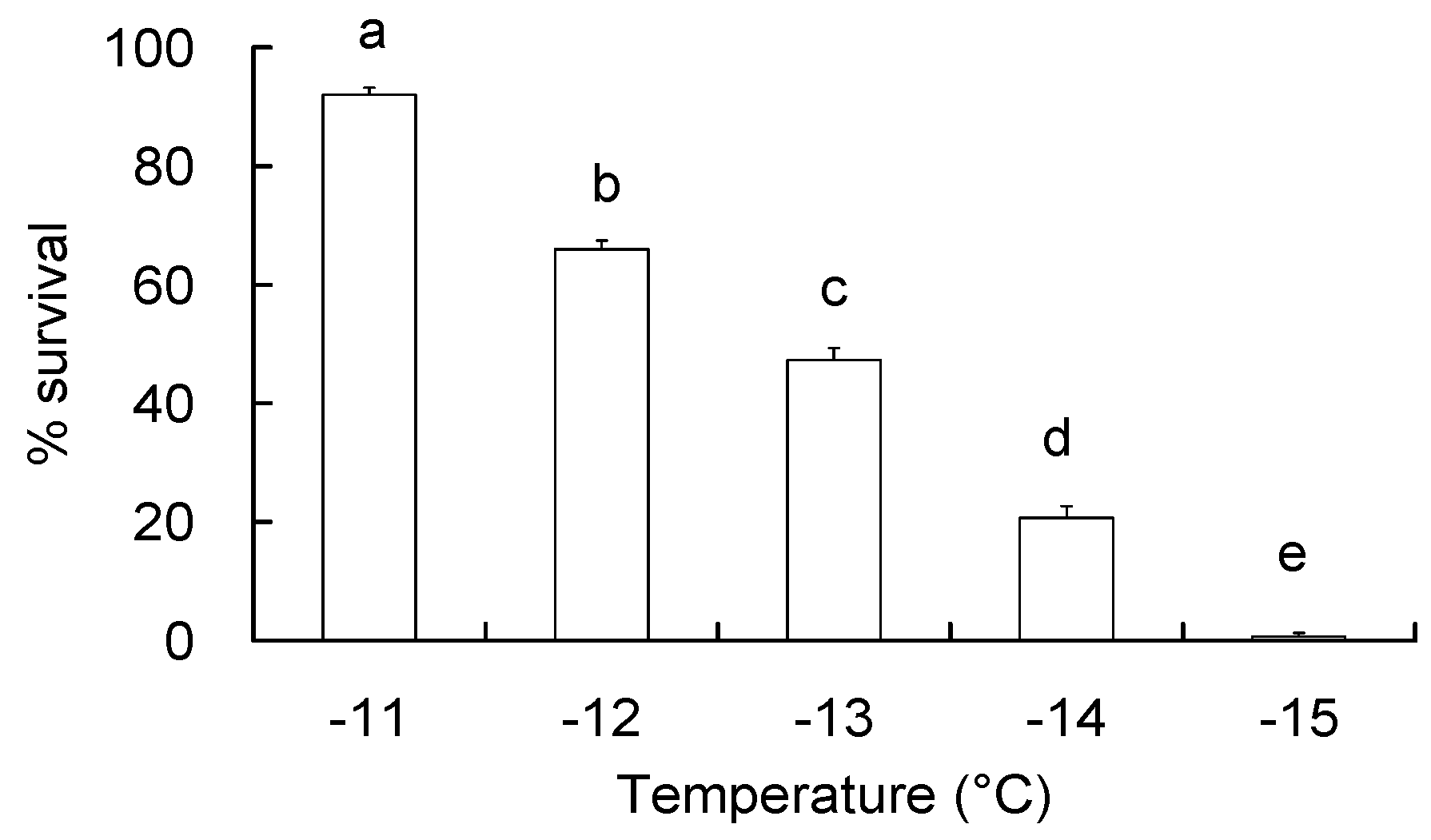
2.2. Determination of Discriminating Temperature
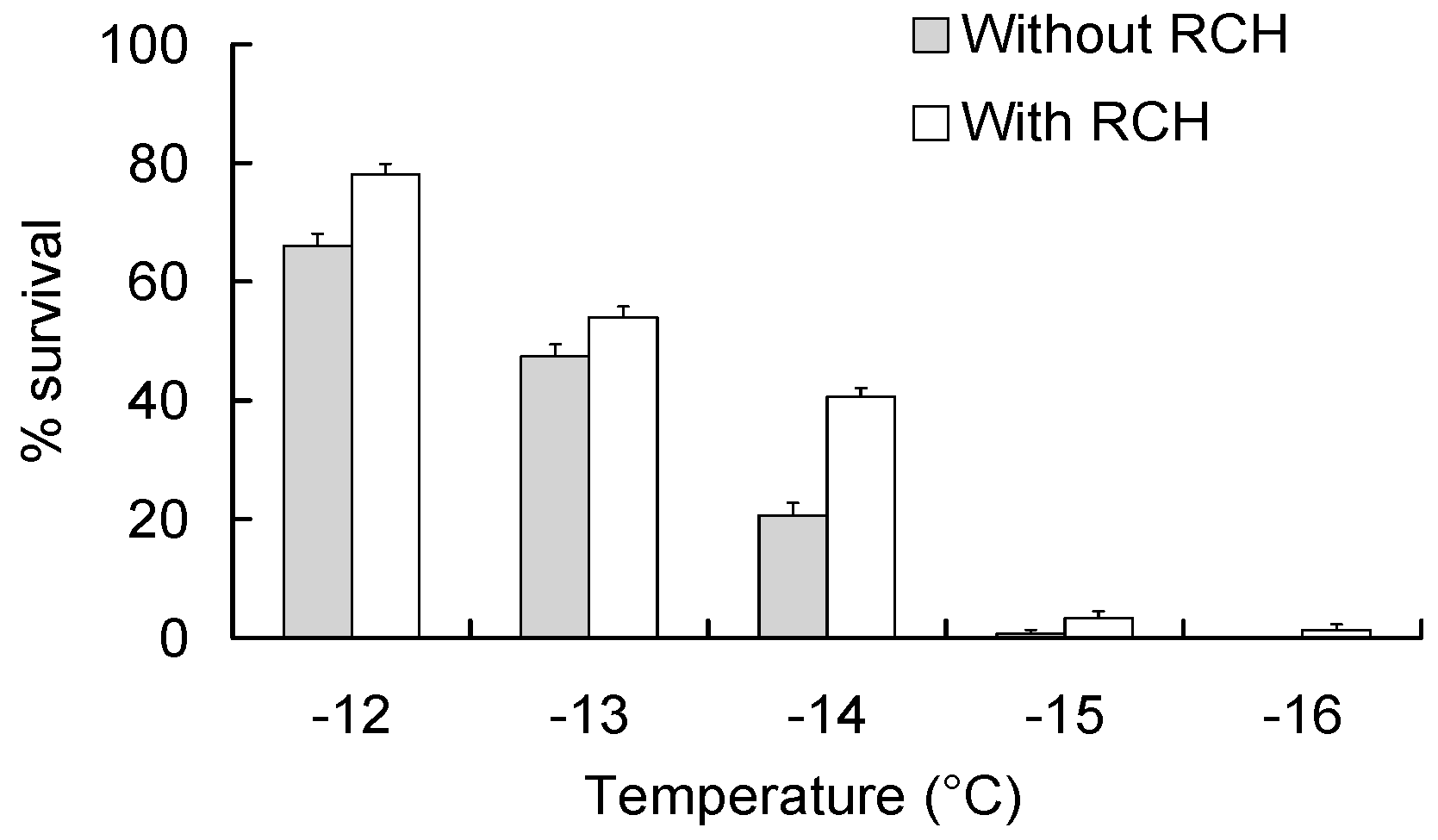
2.3. Induction and Detection of RCH Response
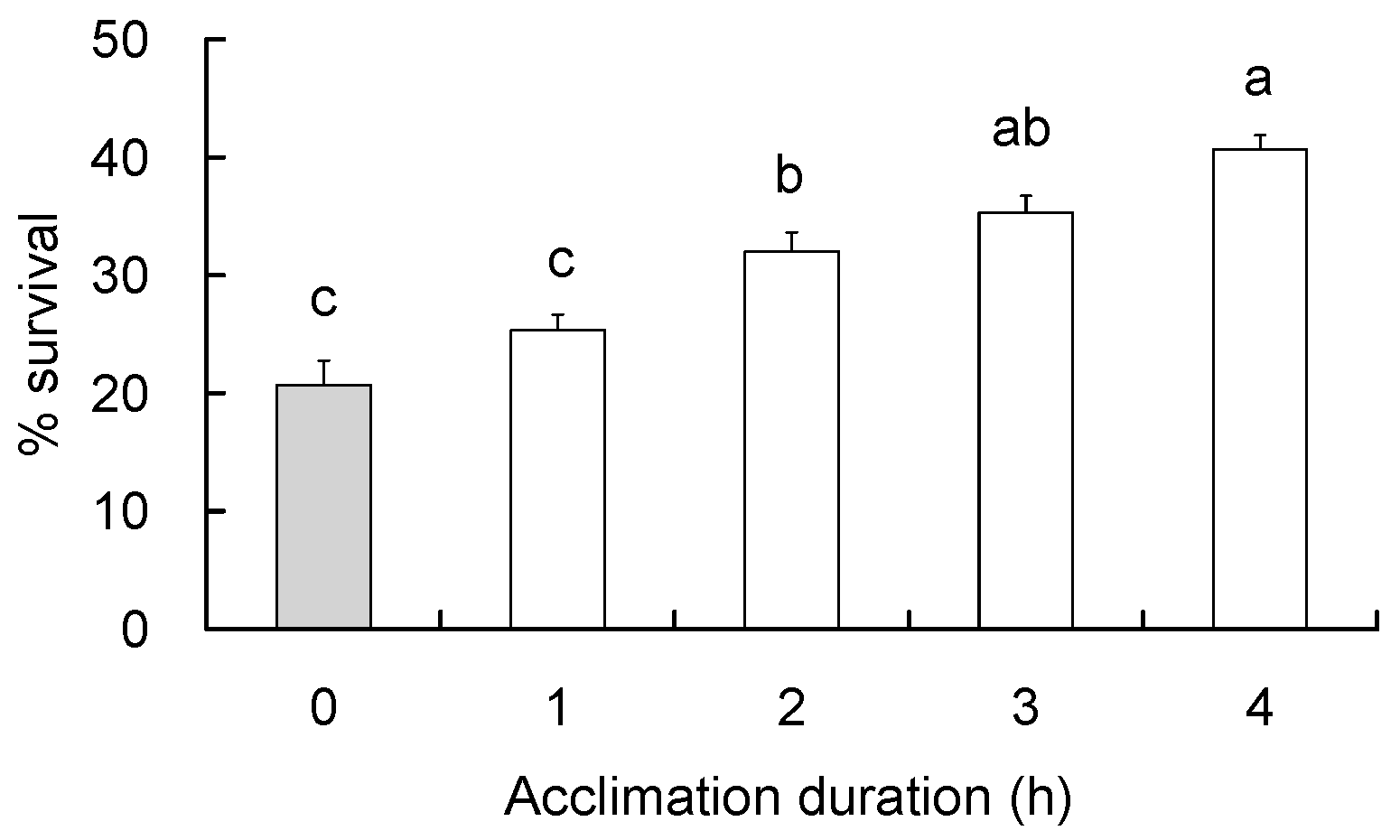
2.4. Extent of RCH Response
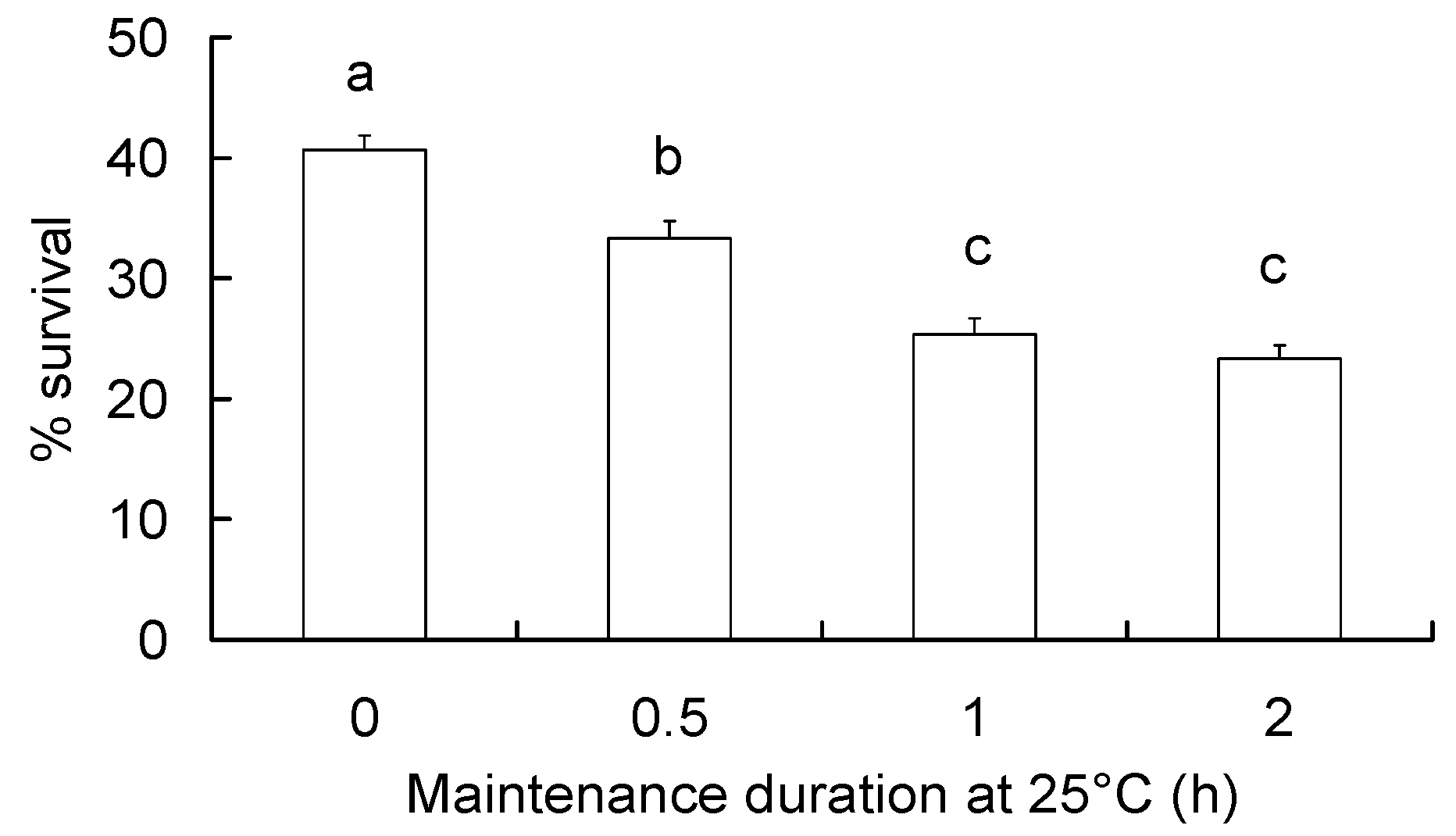
2.5. Durability of RCH Response
2.6. Statistical Analysis
3. Results
3.1. Discriminating Temperature
3.2. RCH Response
3.3. Extent of RCH Response
3.4. Durability of RCH Response
4. Discussion
5. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Overgaard, J.; MacMillan, H.A. The integrative physiology of insect chill tolerance. Annu. Rev. Physiol. 2017, 79, 187–208. [Google Scholar] [CrossRef] [PubMed]
- Lee, R.E., Jr.; Denlinger, D.L. Rapid cold-hardening: Ecological significance and underpinning mechanisms. In Low Temperature Biology of Insects; Denlinger, D.L., Lee, R.E., Jr., Eds.; Cambridge University Press: Cambridge, UK, 2010; pp. 35–58. ISBN 9780521886352. [Google Scholar]
- Coulson, S.J.; Bale, J.S. Characterisation and limitations of the rapid cold-hardening response in the housefly Musca domestica (Diptera: Muscidae). J. Insect Physiol. 1990, 36, 207–211. [Google Scholar] [CrossRef]
- Powell, S.J.; Bale, J.S. Cold shock injury and ecological costs of rapid cold hardening in the grain aphid Sitobion avenae (Hemiptera: Aphididae). J. Insect Physiol. 2004, 50, 277–284. [Google Scholar] [CrossRef] [PubMed]
- Lee, R.E., Jr.; Chen, C.P.; Denlinger, D.L. A rapid cold-hardening process in insects. Science 1987, 238, 1415–1417. [Google Scholar] [CrossRef] [PubMed]
- MacMillan, H.A.; Guglielmo, C.G.; Sinclair, B.J. Membrane remodeling and glucose in Drosophila melanogaster: A test of rapid cold-hardening and chilling tolerance hypotheses. J. Insect Physiol. 2009, 55, 243–249. [Google Scholar] [CrossRef] [PubMed]
- Czajka, M.C.; Lee, R.E., Jr. A rapid cold-hardening response protecting against cold shock injury in Drosophila melanogaster. J. Exp. Biol. 1990, 148, 245–254. [Google Scholar] [PubMed]
- McDonald, J.R.; Bale, J.S.; Walters, K.F.A. Rapid cold hardening in the western flower thrips Frankliniella occidentalis. J. Insect Physiol. 1997, 43, 759–766. [Google Scholar] [CrossRef]
- Shintani, Y.; Ishikawa, Y. Relationship between rapid cold-hardening and cold acclimation in the eggs of the yellow-spotted longicorn beetle, Psacothea hilaris. J. Insect Physiol. 2007, 53, 1055–1062. [Google Scholar] [CrossRef] [PubMed]
- Qiang, C.-K.; Du, Y.-Z.; Yu, L.-Y.; Cui, Y.-D.; Zheng, F.-S.; Lu, M.-X. Effect of rapid cold hardening on the cold tolerance of the larvae of the rice stem borer, Chilo suppressalis (Walker). Agric. Sci. China 2008, 7, 321–328. [Google Scholar] [CrossRef]
- Overgaard, J.; Malmendal, A.; Sorensen, J.G.; Bundy, J.G.; Loeschcke, V.; Nielsen, N.C.; Holmstrup, M. Metabolomic profiling of rapid cold hardening and cold shock in Drosophila melanogaster. J. Insect Physiol. 2008, 53, 1218–1232. [Google Scholar] [CrossRef] [PubMed]
- Teets, N.M.; Denlinger, D.L. Physiological mechanisms of seasonal and rapid cold-hardening in insects. Physiol. Entomol. 2013, 38, 105–116. [Google Scholar] [CrossRef]
- Overgaard, J.; Sorensen, J.G.; Comc, E.; Colinet, H. The rapid cold hardening response of Drosophila melanogaster: Complex regulation across different levels of biological organization. J. Insect Physiol. 2014, 62, 46–53. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.; Kim, K.; Kim, Y. Rapid cold hardening of Thrips palmi (Thysanoptera: Thripidae). Environ. Entomol. 2014, 43, 1076–1083. [Google Scholar] [CrossRef] [PubMed]
- Hou, M.; Lin, W.; Han, Y. Seasonal changes in supercooling points and glycerol content in overwintering larvae of the Asiatic rice borer from rice and water-oat plants. Environ. Entomol. 2009, 38, 1182–1188. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Han, Y.; Hu, Q.; Hou, M. Diapause induction and photoperiodic clock in Chilo suppressalis Walker (Lepidoptera: Crambidae). Entomol. Sci. 2011, 14, 283–290. [Google Scholar] [CrossRef]
- Su, J.-W.; Xuan, W.-J.; Sheng, C.-F.; Ge, F. Biology of overwintering larvae of the Asiatic rice borer, Chilo suppressalis, in paddy fields of Northeast China. Entomol. Knowl. 2003, 40, 323–325. [Google Scholar]
- Izumi, Y.; Sonoda, S.; Tsumuki, H. Effects of diapause and cold-acclimation on the avoidance of freezing injury in fat body tissue of the rice stem borer, Chilo suppressalis Walker. J. Insect Physiol. 2007, 53, 685–690. [Google Scholar] [CrossRef] [PubMed]
- Hou, M.; Han, Y.; Lin, W. Influence of soil moisture on supercooling capacity and associated physiological parameters of overwintering larvae of the rice stem borer. Entomol. Sci. 2009, 12, 155–161. [Google Scholar] [CrossRef]
- Han, L.; Li, S.; Liu, P.; Peng, Y.; Hou, M. New artificial diet for continuous rearing of Chilo suppressalis (Lepidoptera: Crambidae). Ann. Entomol. Soc. Am. 2012, 105, 253–258. [Google Scholar] [CrossRef]
- Broufas, G.D.; Koveos, D.S. Rapid cold hardening in the predatory mite Euseius (Amblyseius) finlandicus (Acari: Phytoseiidae). J. Insect Physiol. 2001, 47, 699–708. [Google Scholar] [CrossRef]
- Koveos, D.S. Rapid cold hardening in the olive fruit fly Bactrocera oleae under laboratory and field conditions. Ent. Exp. Appl. 2001, 101, 257–263. [Google Scholar] [CrossRef]
- Ju, R.-T.; Xiao, Y.-Y.; Li, B. Rapid cold hardening increases cold and chilling tolerances more than acclimation in the adults of the sycamore lace bug, Corythucha ciliata (Say) (Hemiptera: Tingidae). J. Insect Physiol. 2011, 57, 1577–1582. [Google Scholar] [CrossRef] [PubMed]
- Cui, F.; Wang, H.; Zhang, H.; Kang, L. Anoxic stress and rapid cold hardening enhance cold tolerance of the migratory locust. Cryobiology 2014, 69, 243–248. [Google Scholar] [CrossRef] [PubMed]
- Cha, W.H.; Lee, D. Identification of rapid cold hardening-related genes in the tobacco budworm, Helicoverpa assulta. J. Asia-Pac. Entomol. 2016, 19, 1061–1066. [Google Scholar] [CrossRef]
- Sinclair, B.J.; Chown, S.L. Rapid cold-hardening in a Karoo beetle, Afrinus sp. Physiol. Entomol. 2006, 31, 98–101. [Google Scholar] [CrossRef]
- Wang, X.H.; Kang, L. Rapid cold-hardening in young hoppers of the migratory locust Locusta migratoria L. (Orthoptera: Acridiidae). CryoLetters 2003, 24, 331–340. [Google Scholar] [PubMed]




© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, G.; Wen, J.; Han, Y.; Hou, M. Rapid Cold Hardening Confers a Transient Increase in Low Temperature Survival in Diapausing Chilo suppressalis Larvae. Insects 2018, 9, 53. https://doi.org/10.3390/insects9020053
Yang G, Wen J, Han Y, Hou M. Rapid Cold Hardening Confers a Transient Increase in Low Temperature Survival in Diapausing Chilo suppressalis Larvae. Insects. 2018; 9(2):53. https://doi.org/10.3390/insects9020053
Chicago/Turabian StyleYang, Guangping, Jihui Wen, Yongqiang Han, and Maolin Hou. 2018. "Rapid Cold Hardening Confers a Transient Increase in Low Temperature Survival in Diapausing Chilo suppressalis Larvae" Insects 9, no. 2: 53. https://doi.org/10.3390/insects9020053
APA StyleYang, G., Wen, J., Han, Y., & Hou, M. (2018). Rapid Cold Hardening Confers a Transient Increase in Low Temperature Survival in Diapausing Chilo suppressalis Larvae. Insects, 9(2), 53. https://doi.org/10.3390/insects9020053