Nutritional Quality during Development Alters Insulin-Like Peptides’ Expression and Physiology of the Adult Yellow Fever Mosquito, Aedes aegypti


Abstract
1. Introduction
2. Materials and Methods
2.1. Mosquitoes
2.2. Quantitative Expression Analysis of ILPs by Real Time RT-PCR
2.3. Mosquito Physiology Bioassays
2.3.1. Developmental Time
2.3.2. Fecundity
2.3.3. Metabolic Assays
2.4. Data Analysis
3. Results
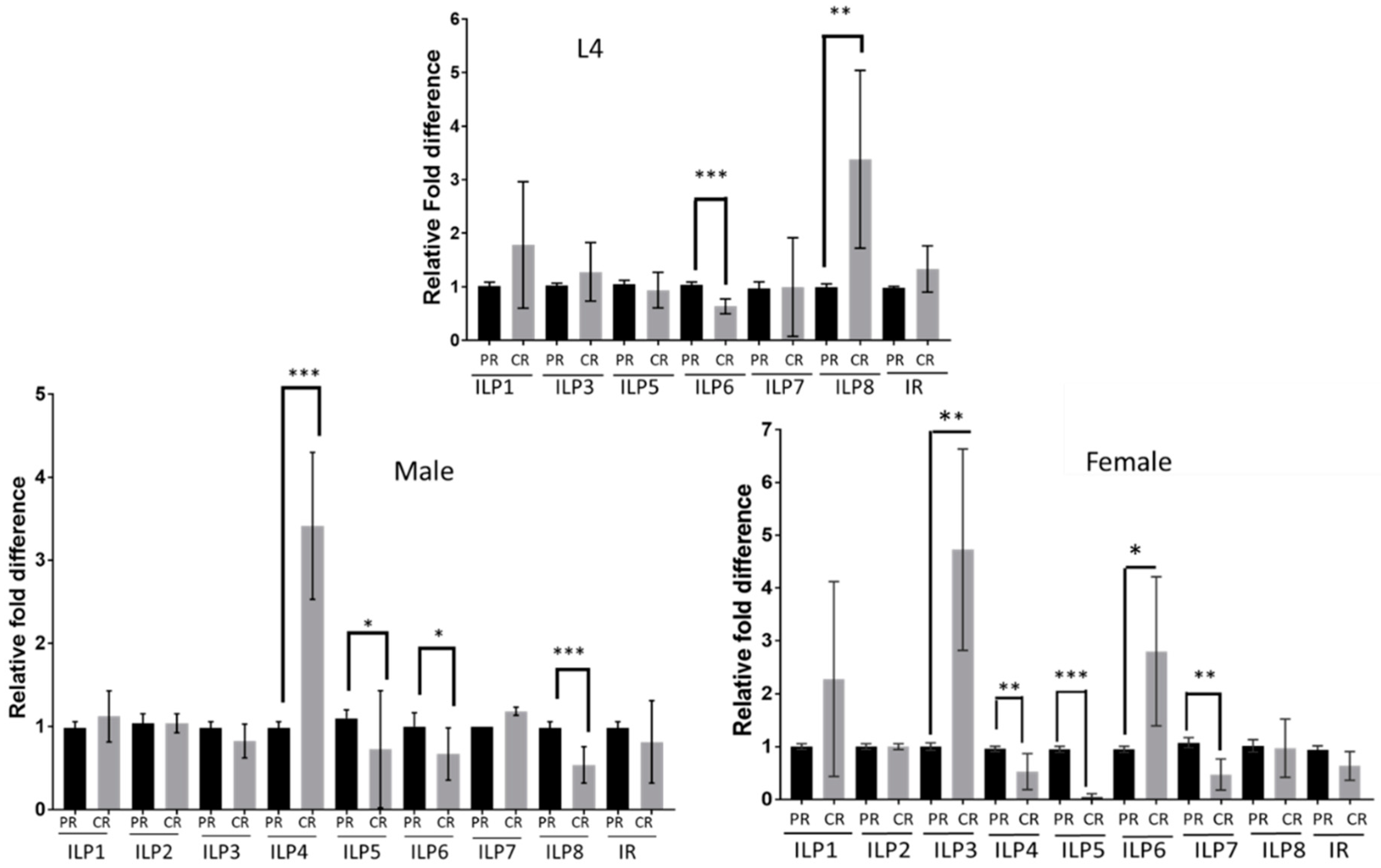
3.1. ILP Expression Changes in Mosquitoes Reared on Different Diets
3.2. Protein Rich Diet Resulted in Shorter Developmental Time
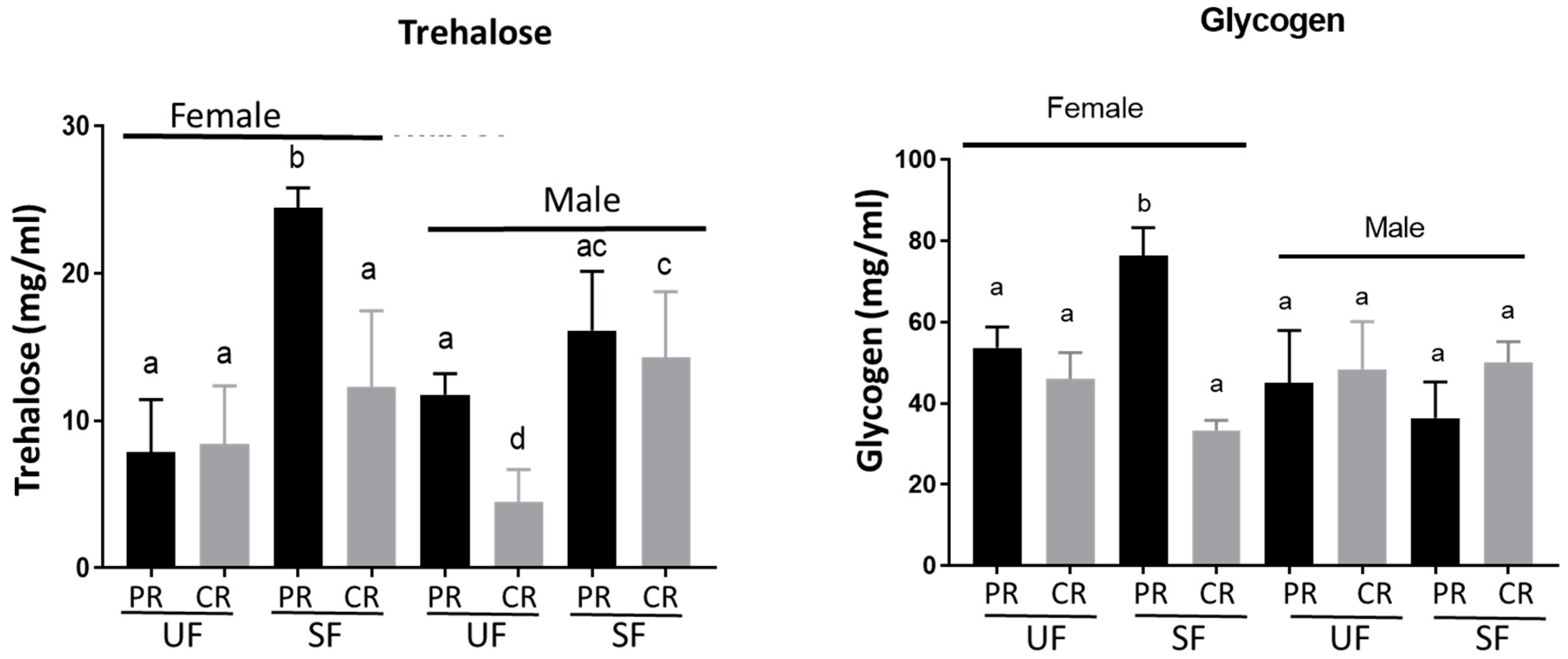
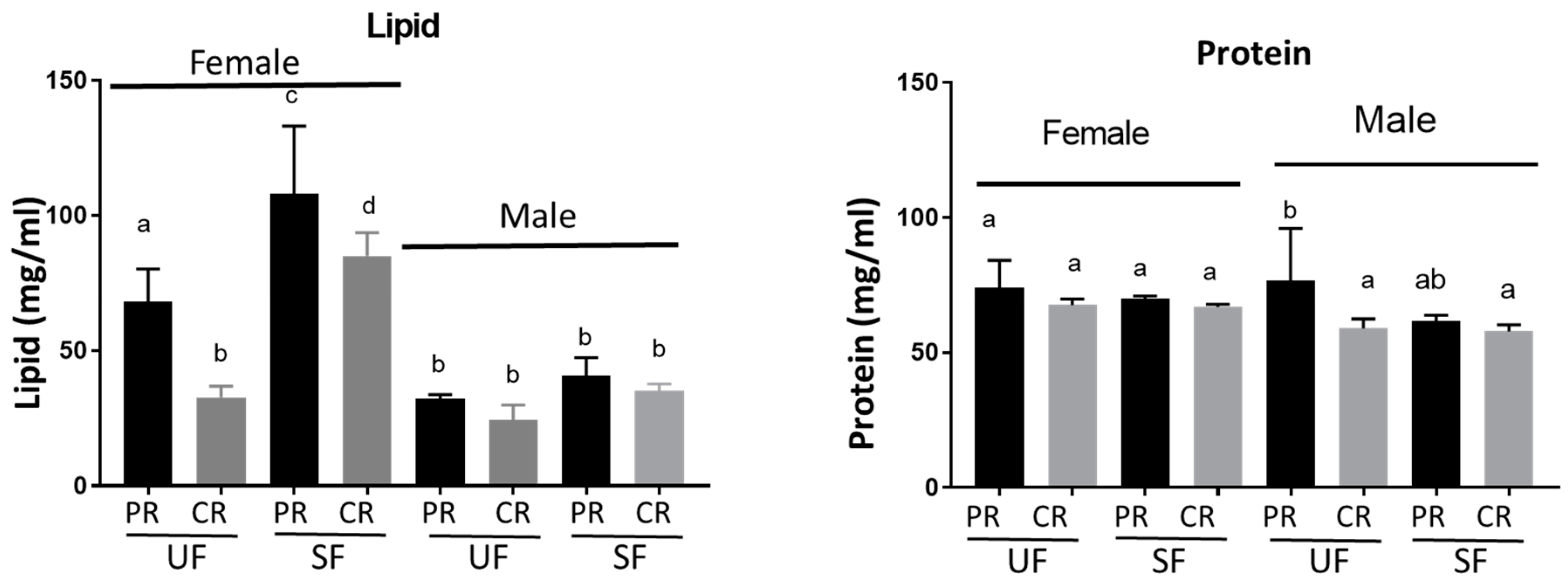
3.3. Levels of Circulating Carbohydrates, Glycogen, and Lipids Varied in Adults from Different Diet Fed Larvae, whereas Protein Levels Remained Constant
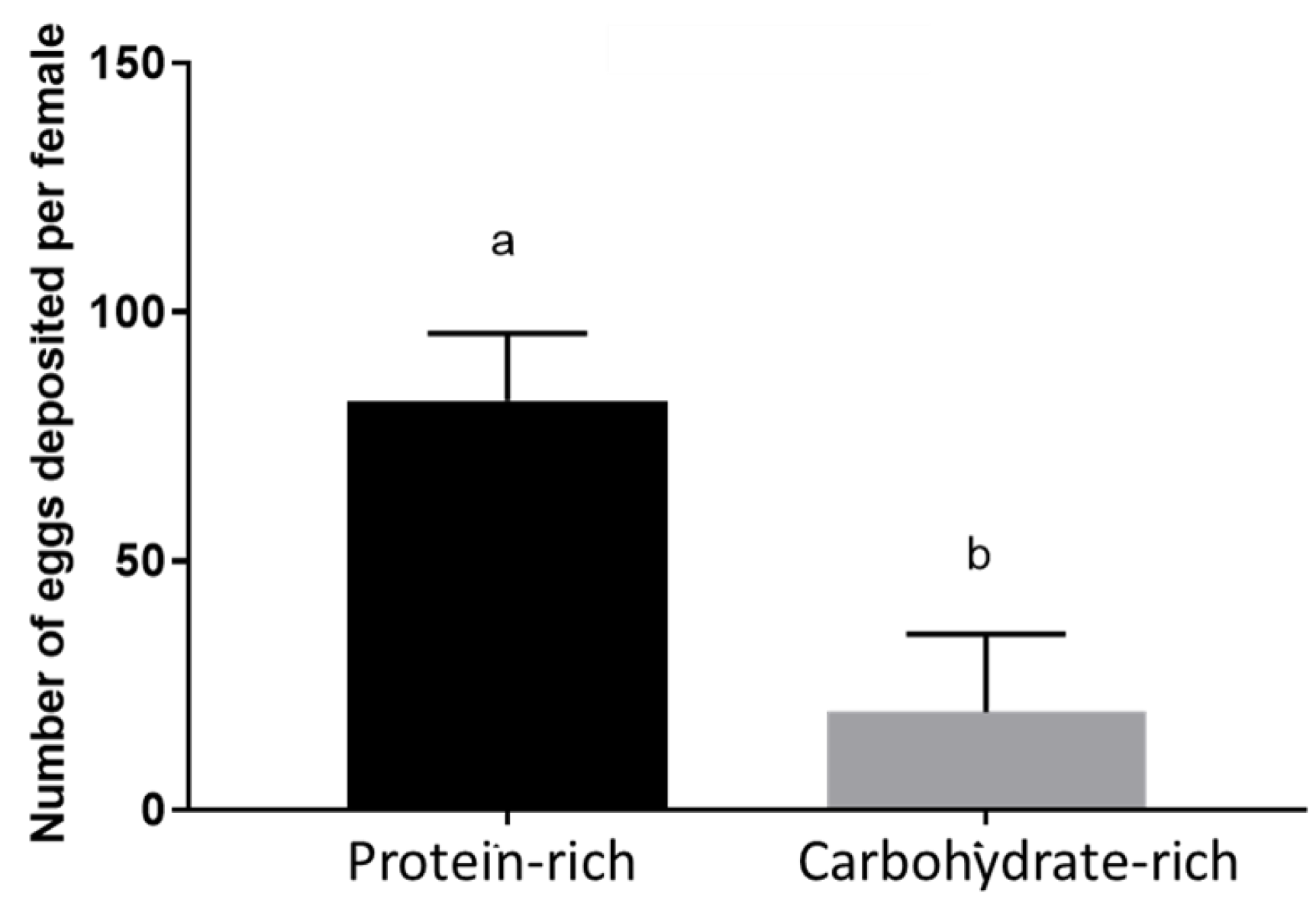
3.4. Protein-Rich Diet Resulted in Higher Fecundity
3.5. Protein-Rich Diet Resulted in Longer Life Span and Difference in Female Body Size
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ILP | insulin-like peptides |
| Dilp | Drosophila insulin-like peptides |
| IR | insulin receptor |
| L1-4 | Larval instars 1-4 |
| RT-PCR | Reverse transcription Polymerase Chain Reaction |
| qRT-PCR | Reverse transcription Polymerase Chain Reaction |
References
- Matsuda, H.; Yamada, T.; Yoshida, M.; Nishimura, T. Flies without Trehalose. J. Biol. Chem. 2015, 290, 1244–1255. [Google Scholar] [CrossRef] [PubMed]
- Saltiel, A.R.; Kahn, C.R. Insulin signaling and the regulation of glucose and lipid metabolism. Nature 2001, 414, 799–806. [Google Scholar] [CrossRef] [PubMed]
- Broughton, S.J.; Piper, M.D.; Ikeya, T.; Bass, T.M.; Jacobson, J.; Driege, Y.; Martinez, P.; Hafen, E.; Withers, D.J.; Leevers, S.J.; et al. Longer lifespan, altered metabolism, and stress resistance in Drosophila from ablation of cells making insulin-like ligands. Proc. Natl. Acad. Sci. USA 2005, 102, 3105–3110. [Google Scholar] [CrossRef] [PubMed]
- Grönke, S.; Clarke, D.F.; Broughton, S.; Andrews, T.D.; Partridge, L. Molecular Evolution and Functional Characterization of Drosophila Insulin-Like Peptides. PLoS Genet. 2010, 6, e1000857. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.K.; Rulifson, E.J. Conserved mechanisms of glucose sensing and regulation by Drosophila corpora cardiaca cells. Nature 2004, 431, 316–320. [Google Scholar] [CrossRef] [PubMed]
- Riehle, M.A.; Fan, Y.; Cao, C.; Brown, M.R. Molecular characterization of insulin-like peptides in the yellow fever mosquito, Aedes aegypti: Expression, cellular localization, and phylogeny. Peptides 2006, 27, 2547–2560. [Google Scholar] [CrossRef] [PubMed]
- Brown, M.R.; Clark, K.D.; Gulia, M.; Zhao, Z.; Garczynski, S.F.; Crim, J.W.; Suderman, R.J.; Strand, M.R. An insulin-like peptide regulates egg maturation and metabolism in the mosquito Aedes aegypti. Proc. Natl. Acad. Sci. USA 2008, 105, 5716–5721. [Google Scholar] [CrossRef] [PubMed]
- Gulia-Nuss, M.; Eum, J.H.; Strand, M.R.; Brown, M.R. Ovary ecdysteroidogenic hormone activates egg maturation in the mosquito Georgecraigius atropalpus after adult eclosion or a blood meal. J. Exp. Biol. 2012, 215, 3758–3767. [Google Scholar] [CrossRef] [PubMed]
- Gulia-Nuss, M.; Robertson, A.E.; Brown, M.R.; Strand, M.R. Insulin-Like Peptides and the Target of Rapamycin Pathway Coordinately Regulate Blood Digestion and Egg Maturation in the Mosquito Aedes aegypti. PLoS ONE 2011, 6, e20401. [Google Scholar] [CrossRef] [PubMed]
- Wen, Z.; Gulia, M.; Clark, K.D.; Dhara, A.; Crim, J.W.; Strand, M.R.; Brown, M.R. Two insulin-like peptide family members from the mosquito Aedes aegypti exhibit differential biological and receptor binding activities. Mol. Cell. Endocrinol. 2010, 328, 47–55. [Google Scholar] [CrossRef] [PubMed]
- Gulia-Nuss, M.; Elliot, A.; Brown, M.R.; Strand, M.R. Multiple factors contribute to anautogenous reproduction by the mosquito Aedes aegypti. J. Insect Physiol. 2015, 82, 8–16. [Google Scholar] [CrossRef] [PubMed]
- Ling, L.; Kokoza, V.A.; Zhang, C.; Aksoy, E.; Raikhel, A.S. MicroRNA-277 targets insulin-like peptides 7 and 8 to control lipid metabolism and reproduction in Aedes aegypti mosquitoes. Proc. Natl. Acad. Sci. USA 2017, 114, E8017–E8024. [Google Scholar] [CrossRef] [PubMed]
- Ihle, K.E.; Baker, N.A.; Amdam, G.V. Insulin-like peptide response to nutritional input in honey bee workers. J. Insect Physiol. 2014, 69, 49–55. [Google Scholar] [CrossRef] [PubMed]
- Da-Silva AraújoEmail, M.; Gil, L.H.S.; de-Almeida e-Silva, A. Larval food quantity affects development time, survival and adult biological traits that influence the vectorial capacity of Anopheles darlingi under laboratory conditions. Malar. J. 2012, 11, 261. [Google Scholar] [CrossRef]
- Lopes, T.F.; Holcman, M.M.; Barbosa, G.L.; de Fatima Domingos, M.; Barreiros, R.M.O.V. Laboratory evaluation of the development of Aedes aegypti in two seasons: Influence of different places and different densities. Revista Do Instituto de Medicina Tropical de São Paulo 2014, 56, 369–374. [Google Scholar] [CrossRef] [PubMed]
- Good, T.P.; Tatar, M. Age-specific mortality and reproduction respond to adult dietary restriction in Drosophila melanogaster. J. Insect Physiol. 2001, 47, 1467–1473. [Google Scholar] [CrossRef]
- Walker, G.; Houthoofd, K.; Vanfleteren, J.R.; Gems, D. Dietary restriction in C. elegans: From rate-of-living effects to nutrient sensing pathways. Mech. Ageing Dev. 2005, 126, 929–937. [Google Scholar] [CrossRef] [PubMed]
- Fontana, L.; Partridge, L.; Longo, V.D. Extending healthy life span-from yeast to humans. Science 2010, 328, 321–326. [Google Scholar] [CrossRef] [PubMed]
- Gluckman, P.D.; Hanson, M.A. The developmental origins of the metabolic syndrome. Trends Endocrinol. Metab. 2004, 15, 183–187. [Google Scholar] [CrossRef] [PubMed]
- Boggs, C.L.; Freeman, K.D. Larval food limitation in butterflies: Effects on adult resource allocation and fitness. Oecologia 2005, 144, 353–361. [Google Scholar] [CrossRef] [PubMed]
- Barrett, E.L.B.; Hunt, J.; Moore, A.J.; Moore, P.J. Separate and combined effects of nutrition during juvenile and sexual development on female life-history trajectories: The thrifty phenotype in a cockroach. Proc. R. Soc. B Biol. Sci. 2009, 276, 3257–3264. [Google Scholar] [CrossRef] [PubMed]
- Joy, T.K.; Arik, A.J.; Corby-Harris, V.; Johnson, A.A.; Riehle, M.A. The impact of larval and adult dietary restriction on lifespan, reproduction and growth in the mosquito Aedes aegypti. Exp. Gerontol. 2010, 45, 685–690. [Google Scholar] [CrossRef] [PubMed]
- Dmitriew, C.; Rowe, L. The effects of larval nutrition on reproductive performance in a food-limited adult environment. PLoS ONE 2011, 6, e17399. [Google Scholar] [CrossRef] [PubMed]
- Kaspi, R.; Mossinson, S.; Drezner, T.; Kamensky, B.; Yuval, B. Effects of larval diet on development rates and reproductive maturation of male and female Mediterranean fruit flies. Physiol. Entomol. 2002, 27, 29–38. [Google Scholar] [CrossRef]
- Colasurdo, N.; Gélinas, Y.; Despland, E. Larval nutrition affects life history traits in a capital breeding moth. J. Exp. Biol. 2009, 212, 1794–1800. [Google Scholar] [CrossRef] [PubMed]
- Kolss, M.; Vijendravarma, R.K.; Schwaller, G.; Kawecki, T.J. Life-history consequences of adaptation to larval nutritional stress in Drosophila. Evol. Int. J. Org. Evol. 2009, 63, 2389–2401. [Google Scholar] [CrossRef] [PubMed]
- Colombani, J.; Andersen, D.S.; Leopold, P. Secreted peptide Dilp8 coordinates Drosophila tissue growth with developmental timing. Science 2012, 336, 582–585. [Google Scholar] [CrossRef] [PubMed]
- Garelli, A.; Gontijo, A.M.; Miguela, V.; Caparros, E.; Dominguez, M. Imaginal discs secrete insulin-like peptide 8 to mediate plasticity of growth and maturation. Science 2012, 336, 579–582. [Google Scholar] [CrossRef] [PubMed]
- Ikeya, T.; Galic, M.; Belawat, P.; Nairz, K.; Hafen, E. Nutrient-dependent expression of insulin-like peptides from neuroendocrine cells in the CNS contributes to growth regulation in Drosophila. Curr. Biol. 2002, 12, 1293–1300. [Google Scholar] [CrossRef]
- Slaidina, M.; Delanoue, R.; Gronke, S.; Partridge, L.; Leopold, P. A Drosophila insulin-like peptide promotes growth during nonfeeding states. Dev. Cell 2009, 17, 874–884. [Google Scholar] [CrossRef] [PubMed]
- Bai, H.; Kang, P.; Tatar, M. Drosophila insulin-like peptide-6 (dilp6) expression from fat body extends lifespan and represses secretion of Drosophila insulin-like peptide-2 from the brain. Aging Cell 2012, 11, 978–985. [Google Scholar] [CrossRef] [PubMed]
- Min, K.; Yamamoto, R.; Buch, S.; Pankratz, M.; Tatar, M. Drosophila lifespan control by dietary restriction independent of insulin-like signaling. Aging Cell 2008, 7, 199–206. [Google Scholar] [CrossRef] [PubMed]
- Broughton, S.J.; Slack, C.; Alic, N.; Metaxakis, A.; Bass, T.M.; Driege, Y.; Partridge, L. DILP-producing median neurosecretory cells in the Drosophila brain mediate the response of lifespan to nutrition. Aging Cell 2010, 9, 336–346. [Google Scholar] [CrossRef] [PubMed]
- Okamoto, N.; Nishimura, T. Signaling from glia and cholinergic neurons controls nutrient-dependent production of an insulin-like peptide for Drosophila body growth. Dev. Cell 2015, 35, 295–310. [Google Scholar] [CrossRef] [PubMed]
- Post, S.; Tatar, M. Nutritional Geometric Profiles of Insulin/IGF Expression in Drosophila melanogaster. PLoS ONE 2016, 11, e0155628. [Google Scholar] [CrossRef] [PubMed]
- Bunning, H.; Bassett, L.; Clowser, C.; Rapkin, J.; Jensen, K.; House, C.M.; Hunt, J. Dietary choice for a balanced nutrient intake increases the mean and reduces the variance in the reproductive performance of male and female cockroaches. Ecol. Evol. 2016, 6, 4711–4730. [Google Scholar] [CrossRef] [PubMed]
- Briegel, H. Fecundity, metabolism, and body size in Anopheles (Diptera: Culicidae), vectors of malaria. J. Med. Entomol. 1990, 27, 839–850. [Google Scholar] [CrossRef] [PubMed]
- Briegel, H. Metabolic relationship between female body size, reserves, and fecundity of Aedes aegypti. J. Insect Physiol. 1990, 36, 165–172. [Google Scholar] [CrossRef]
- Telang, A.; Wells, M.A. The effect of larval and adult nutrition on successful autogenous egg production by a mosquito. J. Insect Physiol. 2004, 50, 677–685. [Google Scholar] [CrossRef] [PubMed]
- Ziegler, R.; Ibrahim, M.M. Formation of lipid reserves in fat body and eggs of the yellow fever mosquito, Aedes aegypti. J. Insect Physiol. 2001, 47, 623–627. [Google Scholar] [CrossRef]
- Shapiro, L.L.M.; Murdock, C.C.; Jacobs, G.R.; Thomas, R.J.; Thomas, M.B. Larval food quantity affects the capacity of adult mosquitoes to transmit human malaria. Proc. R. Soc. B Biol. Sci. 2016, 283. [Google Scholar] [CrossRef] [PubMed]
- Davies, L.R.; Schou, M.F.; Kristensen, T.N.; Loeschcke, V. Linking developmental diet to adult foraging choice in Drosophila melanogaster. J. Exp. Biol. 2018, 221. [Google Scholar] [CrossRef] [PubMed]
- Reis, T. Effects of Synthetic Diets Enriched in Specific Nutrients on Drosophila Development, Body Fat, and Lifespan. PLoS ONE 2016, 11, e0146758. [Google Scholar] [CrossRef] [PubMed]
- Van Handel, E.; Lum, P.T.M. Sex as regulator of triglyceride metabolism in the mosquito. Science 1961, 134, 1979–1980. [Google Scholar] [CrossRef] [PubMed]
- Van Handel, E. Metabolism of nutrients in the adult mosquito. Mosq. News 1984, 44, 573–579. [Google Scholar]
- Lamont, B.J.; Waters, M.F.; Andrikopoulos, S. A low-carbohydrate high-fat diet increases weight gain and does not improve glucose tolerance, insulin secretion or β-cell mass in NZO mice. Nutr. Diabetes 2016, 6, e194. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Primer Sequence |
|---|---|
| ILP1 Fwd | 5’-ACTGGTTTGCAACAGCTACC-3’ |
| ILP1 Rev | 5’-TCCAGGTCCTGTTTGATCTC-3’ |
| ILP2 Fwd | 5’-CATCACCGCTCAGAATACCT-3’ |
| ILP2 Rev | 5’-AGAACGGAAAACCGTGACTA-3’ |
| ILP3 Fwd | 5’-ACCAACTTGCGAGTATCGAG-3’ |
| ILP3 Rev | 5’-TGTACTACGGTTCCGACCAT-3’ |
| ILP4 Fwd | 5’-TACTCGAAGCACGACCCTAT-3’ |
| ILP4 Rev | 5’-GGCAACATTCCTCTACGATG-3’ |
| ILP5 Fwd | 5’-CTAATCCGGCACCTTTACTG-3’ |
| ILP5 Rev | 5’-AAGGGTAGCGCATTAGCAC-3’ |
| ILP6 Fwd | 5’-GAGCAAATCCACAACTCCAG-3’ |
| ILP6 Rev | 5’-GCACAGTTCCAAATTCCATC-3’ |
| ILP7 Fwd | 5’- GCGCCAACTATGACAAAACT-3’ |
| ILP7 Rev | 5’- AGGGTTTGTAGCAACAGTCG-3’ |
| ILP8 Fwd | 5’- AGGGCCATTCTACAAGCTCT-3’ |
| ILP8 Rev | 5’- AGGAATGTTTCTCCGTGTCC-3’ |
| Insulin receptor Fwd | 5’- AATGGTTACCGCCACTGAAG-3’ |
| Insulin receptor Rev | 5’- GCACTGATCCGCAGTACAGA-3’ |
| Ribosomal protein S7 Fwd | 5’- ACCGCCGTCTACGATGCCA-3’ |
| Ribosomal protein S7 Rev | 5’- ATGGTGGTCTGCTGGTTCTT-3’ |
| Diet | Developmental Time per Life Stage | Total | |||
|---|---|---|---|---|---|
| L1 to L2 | L2 to L3 | L3 to L4 | L4 to pupae | ||
| PR | 38 ± 5 | 44 ± 4 | 48 ± 5 | 48 ± 4 | 178 ± 18 |
| CR | 38 ± 4 | 50 ± 6 | 64 ± 6 | 84 ± 8 | 236 ± 24 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pooraiiouby, R.; Sharma, A.; Beard, J.; Reyes, J.; Nuss, A.; Gulia-Nuss, M. Nutritional Quality during Development Alters Insulin-Like Peptides’ Expression and Physiology of the Adult Yellow Fever Mosquito, Aedes aegypti. Insects 2018, 9, 110. https://doi.org/10.3390/insects9030110
Pooraiiouby R, Sharma A, Beard J, Reyes J, Nuss A, Gulia-Nuss M. Nutritional Quality during Development Alters Insulin-Like Peptides’ Expression and Physiology of the Adult Yellow Fever Mosquito, Aedes aegypti. Insects. 2018; 9(3):110. https://doi.org/10.3390/insects9030110
Chicago/Turabian StylePooraiiouby, Rana, Arvind Sharma, Joshua Beard, Jeremiah Reyes, Andrew Nuss, and Monika Gulia-Nuss. 2018. "Nutritional Quality during Development Alters Insulin-Like Peptides’ Expression and Physiology of the Adult Yellow Fever Mosquito, Aedes aegypti" Insects 9, no. 3: 110. https://doi.org/10.3390/insects9030110
APA StylePooraiiouby, R., Sharma, A., Beard, J., Reyes, J., Nuss, A., & Gulia-Nuss, M. (2018). Nutritional Quality during Development Alters Insulin-Like Peptides’ Expression and Physiology of the Adult Yellow Fever Mosquito, Aedes aegypti. Insects, 9(3), 110. https://doi.org/10.3390/insects9030110