An Advanced Numerical Trajectory Model Tracks a Corn Earworm Moth Migration Event in Texas, USA




Abstract
:1. Introduction
2. Materials and Methods
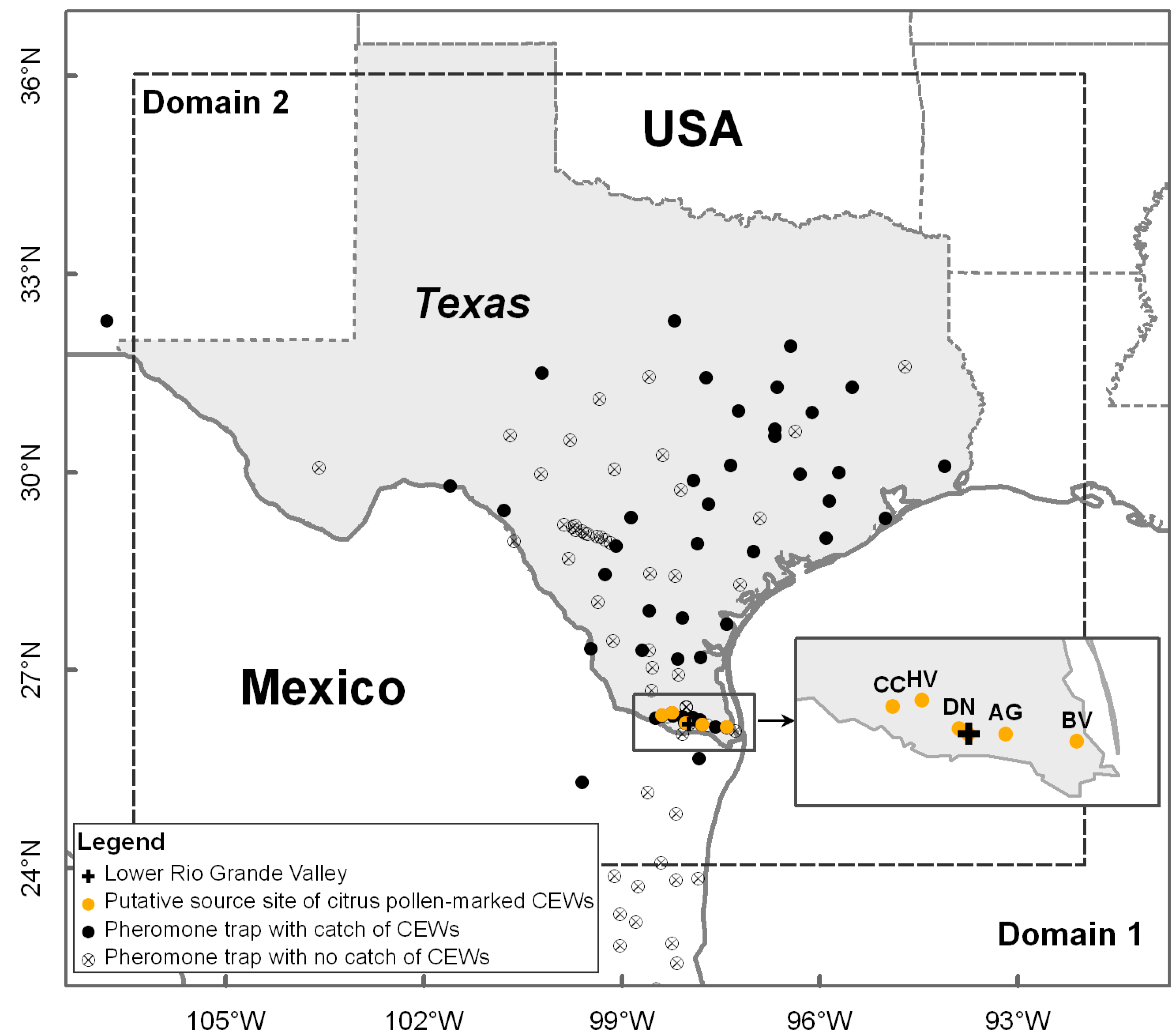
2.1. Study Area and Occurrence of H. zea During 20–22 March 1995 in South-Central Texas
2.2. Trajectory Modelling Methods
2.2.1. HYSPLIT Model Simulation of H. zea Migration
2.2.2. WRF Model Simulation of H. zea Migration
2.3. Evaluation of the Modelling Methods
3. Results
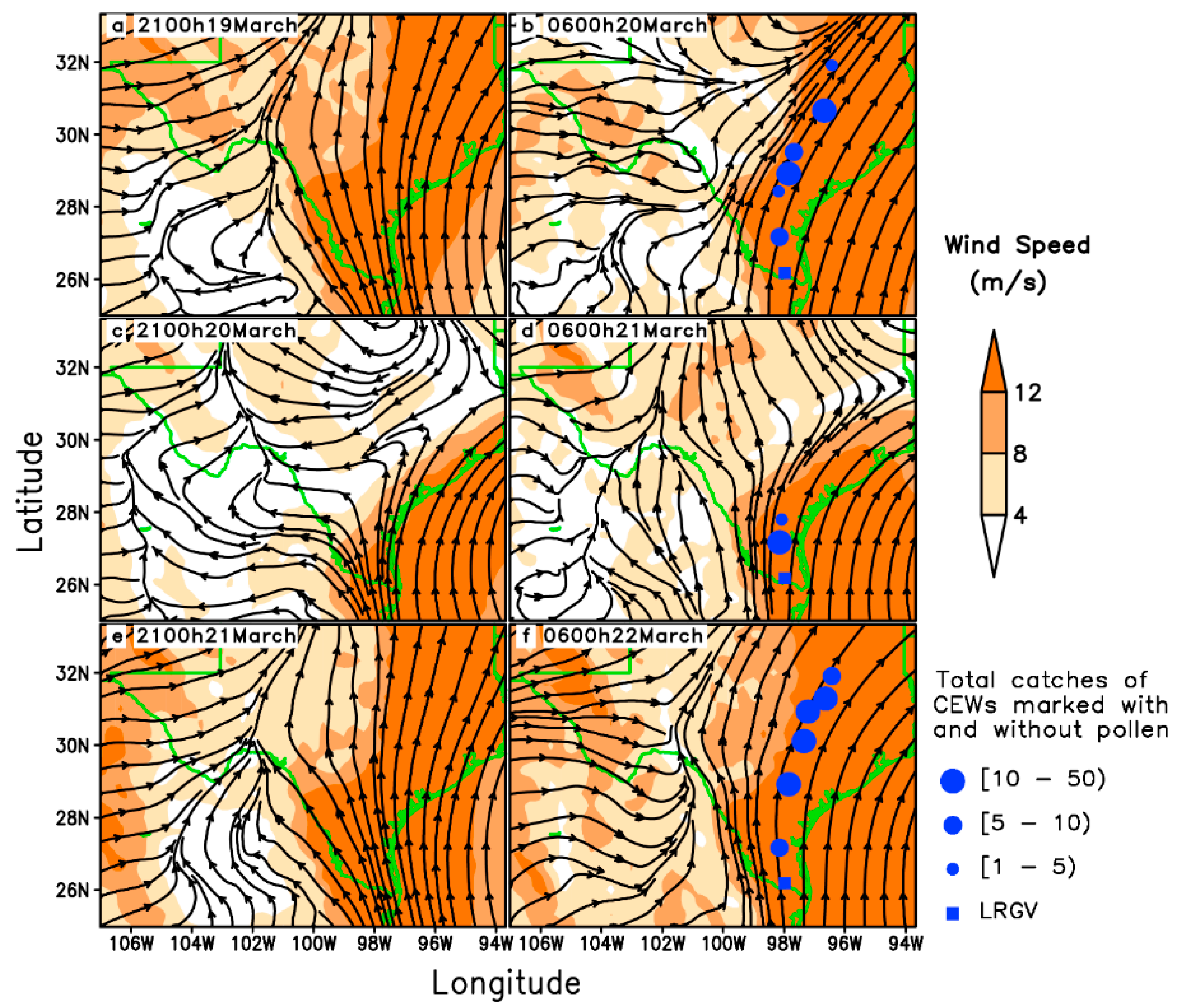
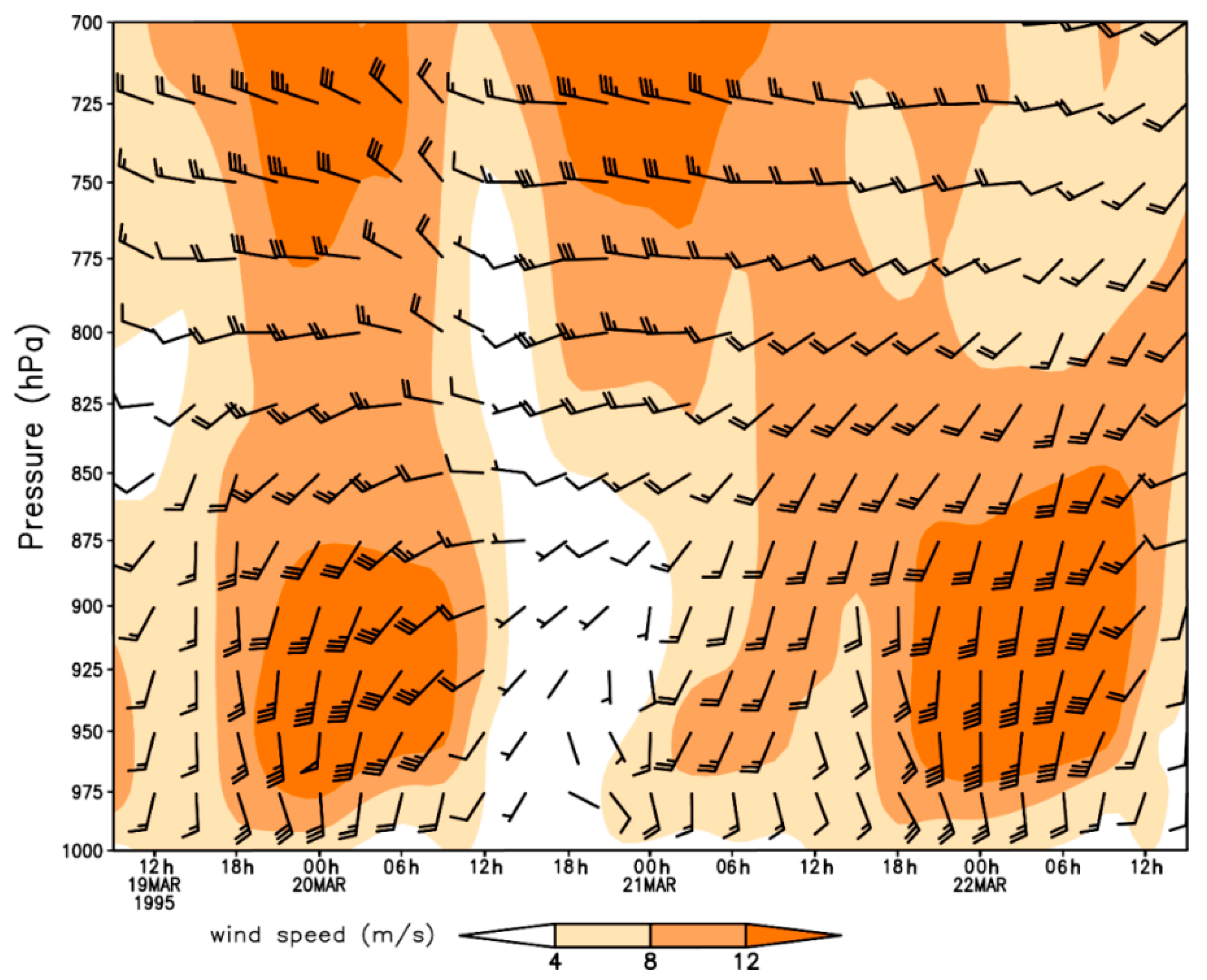
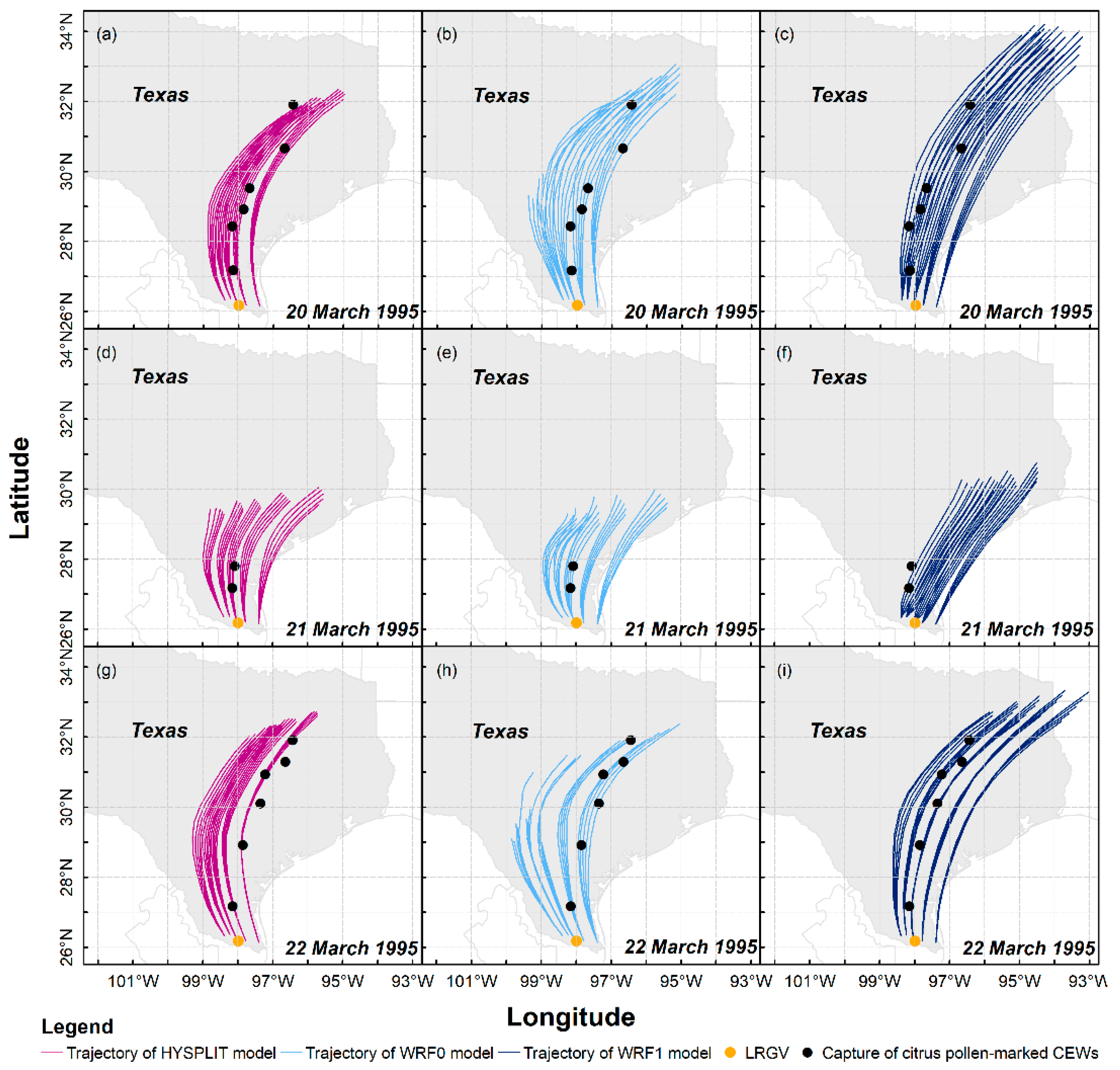
3.1. Long-Distance Migration Processes of Pollen-Marked H. zea Males Over Texas on 20–22 March 1995
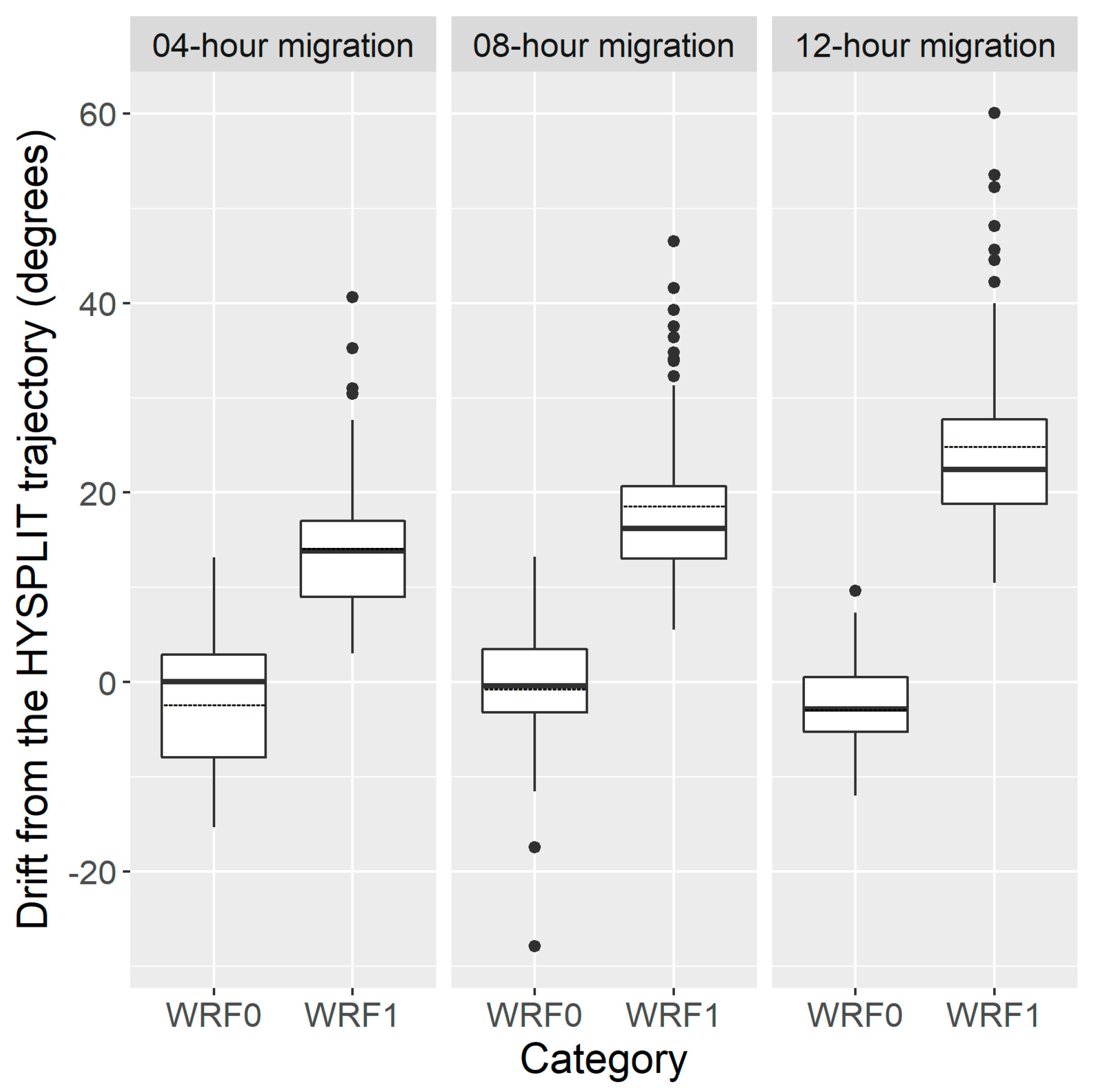
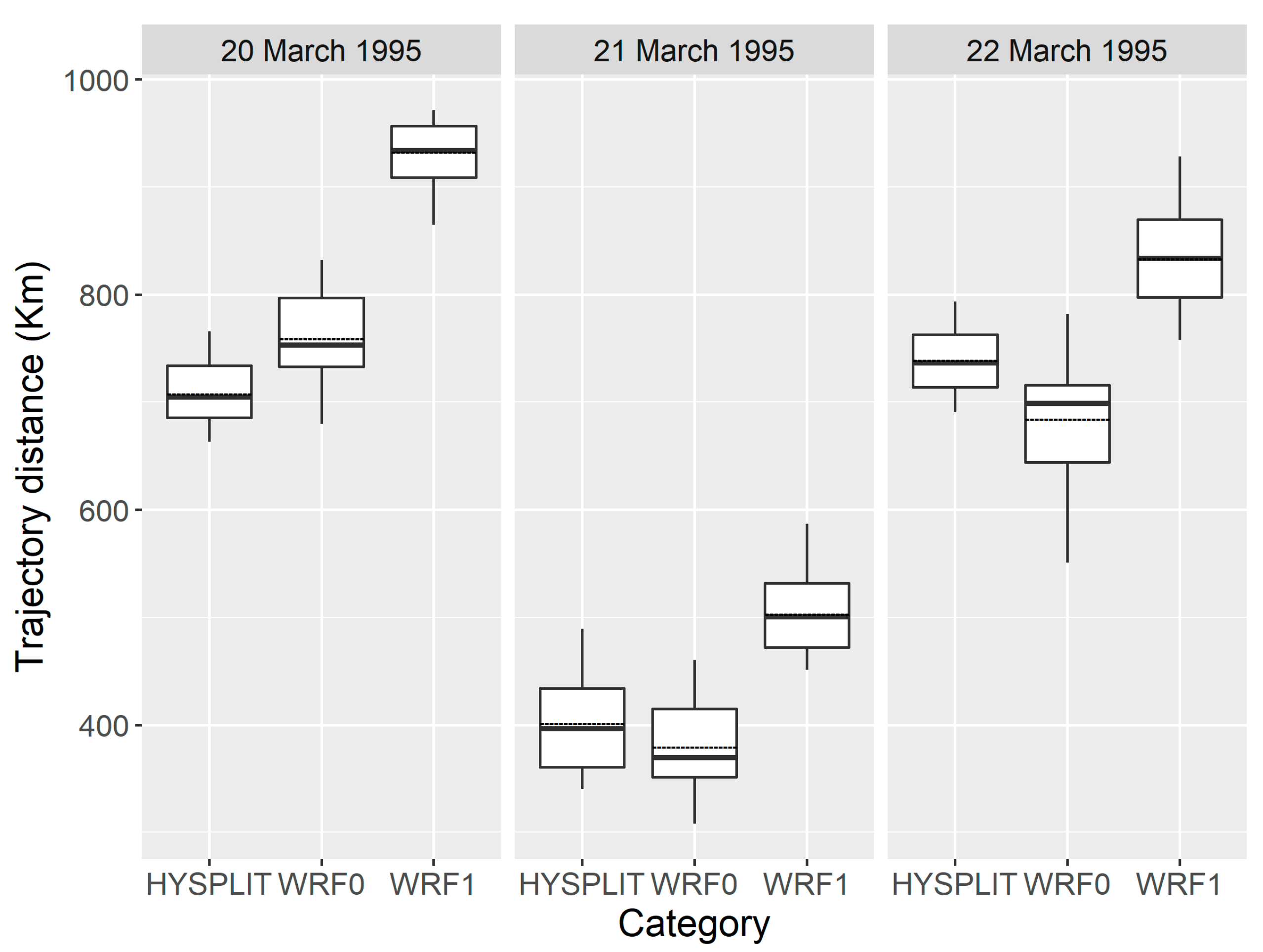
3.2. Statistical Comparisons of Simulations on H. zea Migration Trajectories over Texas
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Johnson, S.J. Insect migration in North America: Synoptic-scale transport in a highly seasonal environment. In Insect Migration: Tracking Resources through Space and Time; Drake, V.A., Gatehouse, A.G., Eds.; Cambridge University Press: Cambridge, UK, 1995; pp. 31–66. [Google Scholar]
- Pedgley, D.E.; Reynolds, D.R.; Tatchel, G.M. Long-range insect migration in relation to climate and weather: Africa and Europe. In Insect Migration: Tracking Resources through Space and Time; Drake, V.A., Gatehouse, A.G., Eds.; Cambridge University Press: Cambridge, UK, 1995; pp. 3–29. [Google Scholar]
- Chapman, J.W.; Nesbit, R.L.; Burgin, L.E.; Reynolds, D.R.; Smith, A.D.; Middleton, D.R.; Hill, J.K. Flight orientation behaviors promote optimal migration trajectories in high-flying insects. Science 2010, 327, 682–685. [Google Scholar] [CrossRef] [PubMed]
- Green, K. The transport of nutrients and energy into the Australian Snowy Mountains by migrating bogong moths Agrotis infusa. Austral Ecol. 2011, 36, 25–34. [Google Scholar] [CrossRef]
- Bauer, S.; Hoye, B.J. Migratory animals couple biodiversity and ecosystem functioning worldwide. Science 2014, 344, 1242552. [Google Scholar] [CrossRef] [PubMed]
- Hu, G.; Lim, K.S.; Horvitz, N.; Clark, S.J.; Reynolds, D.R.; Sapir, N.; Chapman, J.W. Mass seasonal bioflows of high-flying insect migrants. Science 2016, 354, 1584–1587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, F.; Zhai, B.P.; Hu, G. Trajectory analysis methods for insect migration research. Chin. J. Appl. Entomol. 2013, 50, 853–862. [Google Scholar]
- Rainey, R.C. Meteorology and the migration of desert locusts. Applications of Synoptic Meteorology in Locust Control. Anti-Locust Mem. 1963, 7, 1–115. [Google Scholar]
- Taylor, R.A.J. A simulation model of locust migratory behaviour. J. Anim. Ecol. 1979, 48, 577–602. [Google Scholar] [CrossRef]
- Rosenberg, L.J.; Magor, J.I. Predicting windborne displacements of the brown planthopper, Nilaparvata lugens from synoptic weather data. 1. Long-distance displacements in the north-east monsoon. J. Anim. Ecol. 1987, 56, 39–51. [Google Scholar] [CrossRef]
- Westbrook, J.K.; Wolf, W.W.; Lingren, P.D.; Raulston, J.R.; Lopez, J.D., Jr.; Matis, J.H.; Eyster, R.S.; Esquivel, J.F.; Schleider, P.G. Early-season migratory flights of corn earworm (Lepidoptera: Noctuidae). Environ. Entomol. 1997, 26, 12–20. [Google Scholar] [CrossRef]
- Westbrook, J.K.; Esquivel, J.F.; Lopez, J.D.; Jones, G.D.; Wolf, W.W.; Raulston, J.R. Validation of bollworm migration across southcentral Texas in 1994–1996. Southwest Entomol. 1998, 23, 209–220. [Google Scholar]
- Wood, C.R.; Chapman, J.W.; Reynolds, D.R.; Barlow, J.F.; Smith, A.D.; Woiwod, I.P. The influence of the atmospheric boundary layer on nocturnal layers of noctuids and other moths migrating over southern Britain. Int. J. Biometeorol. 2006, 50, 193–204. [Google Scholar] [CrossRef] [PubMed]
- Anthes, R.A.; Warner, T.T. Development of hydrodynamic models suitable for air pollution and other mesometerological studies. Mon. Weather Rev. 1978, 106, 1045–1078. [Google Scholar] [CrossRef]
- Ishikawa, H.; Chino, M. Development of regionally extended Worldwide Version of System for Prediction of Environmental Emergency Dose Information: WSPEEDI, (II) Long-range transport model and its application to dispersion of Cesium-137 from Chernobyl. J. Nucl. Sci. Technol. 1991, 28, 642–655. [Google Scholar] [CrossRef]
- Otuka, A.; Watanabe, T.; Suzuki, Y.; Matsumura, M.; Furuno, A.; Chino, M. Real-time prediction system for migration of rice planthoppers Sogatella furcifera (Horváth) and Nilaparvata lugens (Stål) (Homoptera: Delphacidae). Appl. Entomol. Zool. 2005, 40, 221–229. [Google Scholar] [CrossRef]
- Draxler, R.R.; Rolph, G.D. HYSPLIT (HYbrid Single-Particle Lagrangian Integrated Trajectory) Model; NOAA Air Resources Laboratory: Silver Spring, MD, USA. Available online: https://ready.arl.noaa.gov/HYSPLIT.php (accessed on 11 January 2018).
- Zhu, M.; Radcliffe, E.B.; Ragsdale, D.W.; MacRae, I.V.; Seeley, M.W. Low-level jet streams associated with spring aphid migration and current season spread of potato viruses in the US northern Great Plains. Agric. For. Meteorol. 2006, 138, 192–202. [Google Scholar] [CrossRef]
- Miao, J.; Wu, Y.Q.; Gong, Z.J.; He, Y.Z.; Duan, Y.; Jiang, Y.L. Long-distance wind-borne dispersal of Sitodiplosis mosellana Géhin (Diptera: Cecidomyiidae) in northern china. J. Insect Behav. 2013, 26, 120–129. [Google Scholar] [CrossRef]
- Westbrook, J.K.; Nagoshi, R.N.; Meagher, R.L.; Fleischer, S.J.; Jairam, S. Modeling seasonal migration of fall armyworm moths. Int. J. Biometeorol. 2016, 60, 255–267. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Wu, Y.; Li, X.J.; Hu, G.; Lu, M.H.; Zhong, L.; Duan, D.K.; Zhai, B.P. Annual fluctuations of early immigrant populations of Sogatella furcifera (Hemiptera: Delphacidae) in Jiangxi Province, China. J. Econ. Entomol. 2016, 109, 1636–1645. [Google Scholar] [CrossRef] [PubMed]
- Lingren, P.D.; Raulston, J.R.; Popham, T.W.; Wolf, W.W.; Lingren, P.S.; Esquivel, J.F. Flight behavior of corn earworm (Lepidoptera: Noctuidae) moths under low wind speed conditions. Environ. Entomol. 1995, 24, 851–860. [Google Scholar] [CrossRef]
- Wolf, W.W.; Westbrook, J.K.; Raulston, J.R.; Pair, S.D.; Lingren, P.D. Radar observations of orientation of noctuids migrating from corn fields in the Lower Rio Grande Valley. Southwest Entomol. Suppl. 1995, 18, 45–61. [Google Scholar]
- Schaefer, G.W. Radar observations of insect flight. In Insect Flight. Symposia of the Royal Entomological Society of London No. 7; Rainey, R.C., Ed.; Blackwell Scientific: Oxford, UK, 1976; pp. 157–197. [Google Scholar]
- Chapman, J.W.; Reynolds, D.R.; Mouritsen, H.; Hill, J.K.; Riley, J.R.; Sivell, D.; Smith, A.D.; Woiwod, I.P. Wind selection and drift compensation optimize migratory pathways in a high-flying moth. Curr. Biol. 2008, 18, 514–518. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, A.M.; Reynolds, D.R.; Smith, A.D.; Chapman, J.W. A single wind-mediated mechanism explains high-altitude ‘non-goal oriented’ headings and layering of nocturnally migrating insects. Proc. R. Soc. Lond. B Biol. Sci. 2009, 277, 765–772. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.Q.; Liao, D.X. An investigation for computing three-dimensional trajectory. Q. J. Appl. Meteorol. 1992, 3, 328–333. [Google Scholar]
- Hu, G.; Lu, F.; Lu, M.H.; Liu, W.C.; Xu, W.G.; Jiang, X.H.; Zhai, B.P. The influence of typhoon Khanun on the return migration of Nilaparvata lugens (Stål) in eastern China. PLoS ONE 2013, 8, e57277. [Google Scholar] [CrossRef] [PubMed]
- Michalakes, J.; Chen, S.; Dudhia, J.; Hart, L.; Klemp, J.; Middlecoff, J.; Skamarock, W. Development of a next generation regional weather research and forecast model. In Developments in Teracomputing, Proceedings of the Ninth ECMWF Workshop on the Use of High Performance Computing in Meteorology; World Scientific: Singapore, 2001; pp. 269–276. [Google Scholar]
- Hu, G.; Wu, Q.L.; Wu, X.W.; Jiang, Y.Y.; Zeng, J.; Zhai, B.P. Outbreak mechanism of second generation armyworms in northeastern China: A case study in 1978. Chin. J. Appl. Entomol. 2014, 51, 927–942. [Google Scholar]
- Wang, F.Y.; Yang, F.; Lu, M.H.; Luo, S.Y.; Zhai, B.P.; Lim, K.S.; McInerney, C.E.; Hu, G. Determining the migration duration of rice leaf folder (Cnaphalocrocis medinalis (Guenée)) moths using a trajectory analytical approach. Sci. Rep. 2017, 7, 39853. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.L.; Hu, G.; Lu, M.H.; Wang, B.; Zhu, X.X.; Su, F.; Zhang, Z.G.; Zhai, B.P. Mesoscale source areas and landing mechanisms of the early immigrant white-backed planthopper Sogatella furcifera (Horváth) in Hunan Province, China. Acta Ecol. Sin. 2015, 35, 7397–7417. [Google Scholar]
- Wu, Q.L.; Westbrook, J.K.; Hu, G.; Lu, M.H.; Liu, W.C.; Sword, G.A.; Zhai, B.P. Multiscale analyses on a massive immigration process of Sogatella furcifera (Horváth) in south-central China: Influences of synoptic-scale meteorological conditions and topography. Int. J. Biometeorol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Wolf, W.W.; Sparks, A.N.; Pair, S.D.; Westbrook, J.K.; Truesdale, F.M. Radar observations and collections of insects in the Gulf of Mexico. In Insect Flight: Dispersal and Migration; Danthanarayana, W., Ed.; Springer: Berlin/Heidelberg, Germany, 1986; pp. 221–234. [Google Scholar]
- Hendrix, W.H., III; Mueller, T.F.; Phillips, J.R.; Davis, O.K. Pollen as an indicator of long-distance movement of Heliothis zea (Lepidoptera: Noctuidae). Environ. Entomol. 1987, 16, 1148–1151. [Google Scholar] [CrossRef]
- Bryant, V.M.; Pendleton, M.; Murry, R.E.; Lingren, P.D.; Raulston, J.R. Techniques for studying pollen adhering to nectar-feeding corn earworm (Lepidoptera: Noctuidae) moths using scanning electron microscopy. J. Econ. Entomol. 1991, 84, 237–240. [Google Scholar] [CrossRef]
- Lingren, P.D.; Westbrook, J.K.; Bryant, V.M.; Raulston, J.R.; Esquivel, J.F.; Jones, G.D. Origin of corn earworm (Lepidoptera: Noctuidae) migrants as determined by citrus pollen markers and synoptic weather systems. Environ. Entomol. 1994, 23, 562–570. [Google Scholar] [CrossRef]
- Esquivel, J.F.; Lingren, P.D. Citrus pollen retention by adult Helicoverpa zea (Lepidoptera: Noctuidae) after exposure to citrus blooms. J. Econ. Entomol. 2002, 95, 1174–1178. [Google Scholar] [CrossRef] [PubMed]
- Raulston, J.R.; Summy, K.R.; Loera, J.; Pair, S.D.; Sparks, A.N. Population dynamics of corn earworm larvae (Lepidoptera: Noctuidae) on corn in the Lower Rio Grande Valley. Environ. Entomol. 1990, 19, 274–280. [Google Scholar] [CrossRef]
- Raulston, J.R.; Pair, S.D.; Loera, J.; Sparks, A.N.; Wolf, W.W.; Westbrook, J.K.; Fitt, G.P.; Rogers, C.E. Helicoverpa zea (Lepidoptera: Noctuidae) pupa production in fruiting com in Northeast Mexico and South Texas. Environ. Entomol. 1992, 21, 1393–1397. [Google Scholar] [CrossRef]
- Hartstack, A.W.; Lopez, J.D., Jr.; Muller, R.A.; Sterling, W.L.; King, E.G. Evidence of long range migration of Heliothis zea (Boddie) into Texas and Arkansas. Southwest Entomol. 1982, 7, 188–201. [Google Scholar]
- Beerwinkle, K.R.; Lopez, J.D., Jr.; Cheng, D.; Lingren, P.D.; Meola, R.W. Flight potential of feral Helicoverpa zea (Lepidoptera: Noctuidae) males measured with a 32-channel, computer-monitored, flight-mill system. Environ. Entomol. 1995, 24, 1122–1130. [Google Scholar] [CrossRef]
- Westbrook, J.K.; Eyster, R.S.; Wolf, W.W.; Lingren, P.D.; Raulston, J.R. Migration pathways of corn earworm (Lepidoptera: Noctuidae) indicated by tetroon trajectories. Agric. For. Meteorol. 1995, 73, 67–87. [Google Scholar] [CrossRef]
- Skamarock, W.C.; Klemp, J.B.; Dudhia, J.; Gill, D.O.; Barker, D.M.; Duda, M.G.; Huang, X.Y.; Wang, W.; Powers, J.G. A Description of the Advanced Research WRF Version 3; NCAR Technical Note. NCAR/TN-475+STR; Mesoscale and Microscale Meteorology Division, NCAR: Boulder, CO, USA, 2008; p. 125. [Google Scholar]
- Olden, J.D.; Jackson, D.A.; Peres-Neto, P.R. Spatial isolation and fish communities in drainage lakes. Oecologia 2001, 127, 572–585. [Google Scholar] [CrossRef] [PubMed]
- Wang, F. Quantitative Methods and Applications in GIS; Taylor & Francis: Boca Raton, FL, USA, 2006; pp. 19–34. [Google Scholar]
- JMP®, Version 13.2.0; SAS Institute Inc.: Cary, NC, USA, 1989–2007.
- Drake, V.A.; Farrow, R.A. The influence of atmospheric structure and motions on insect migration. Annu. Rev. Entomol. 1988, 33, 183–210. [Google Scholar] [CrossRef]
- Wallin, J.R.; Loonan, D.V. Low-level jet winds, aphid vectors, local weather, and barley yellow dwarf virus outbreaks. Phytopathology 1971, 61, 1068. [Google Scholar] [CrossRef]
- Seino, H.; Shiotsuki, Y.; Oya, S.; Hirai, Y. Prediction of long distance migration of rice planthoppers to northern Kyushu considering low-level jet stream. J. Agric. Meteorol. 1987, 43, 203–208. [Google Scholar] [CrossRef]
- Feng, C.H.; Zhai, B.P.; Zhang, X.X.; Tang, J.Y. Climatology of low-level jet and northward migration of rice planthoppers. Acta Ecol. Sin. 2002, 22, 559–565. [Google Scholar]
- Drake, V.A. Radar observations of moths migrating in a nocturnal low-level jet. Ecol. Entomol. 1985, 10, 259–265. [Google Scholar] [CrossRef]
- Wood, C.R. The Biometeorology of High-Altitude Insect Layers. Ph.D. Thesis, University of Reading, Reading, UK, 2007. [Google Scholar]
- Johnson, C.G. Migration and Dispersal of Insects by Flight; Methuen and Co. Ltd.: London, UK, 1969; p. 727. [Google Scholar]
- Dingle, H. Migration strategies of insects. Science 1972, 175, 1327–1335. [Google Scholar] [CrossRef] [PubMed]
- Jones, C.M.; Papanicolaou, A.; Mironidis, G.K.; Vontas, J.; Yang, Y.; Lim, K.S.; Oakeshott, J.G.; Bass, C.; Chapman, J.W. Genomewide transcriptional signatures of migratory flight activity in a globally invasive insect pest. Mol. Ecol. 2015, 24, 4901–4911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carlson, J.D.; Whalon, M.E.; Landis, D.A.; Gage, S.H. Springtime weather patterns coincident with long-distance migration of potato leafhopper into Michigan. Agric. For. Meteorol. 1992, 59, 183–206. [Google Scholar] [CrossRef]
- Westbrook, J.K.; López, J.D., Jr. Long-distance migration in Helicoverpa zea: What we know and need to know. Southwest Entomol. 2010, 35, 355–360. [Google Scholar] [CrossRef]
- Del Socorro, A.P.; Gregg, P.C. Sunflower (Helianthus annuus L.) pollen as a marker for studies of local movement in Helicoverpa armigera (Hübner) (Lepidoptera: Noctuidae). Aust. J. Entomol. 2001, 40, 257–263. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Date | Location | Position | Number of Trap Catches | Number of H. zea Examined for Pollen | Number (Percentage (%)) of H. zea with Citrus Pollen |
|---|---|---|---|---|---|
| 20 March | Falfurrias | 98.15° W, 27.17° N | 6 | 4 | 1 (25%) |
| Three Rivers | 98.18° W, 28.43° N | 2 | 2 | 1 (50%) | |
| Karnes City | 97.85° W, 28.92° N | 19 | 2 | 2 (100%) | |
| Belmont | 97.68° W, 29.52° N | 9 | 3 | 2 (66.67%) | |
| Rockdale | 96.68° W, 30.66° N | 11 | 1 | 1 (100%) | |
| Richland | 96.43° W, 31.91° N | 3 | 3 | 3 (100%) | |
| 21 March | Falfurrias | 98.15° W, 27.17° N | 10 | 2 | 2 (100%) |
| Alice | 98.08° W, 27.80° N | 2 | 1 | 1 (100%) | |
| 22 March | Falfurrias | 98.15° W, 27.17° N | 8 | 4 | 1 (25%) |
| Bastrop | 97.35° W, 30.11° N | 14 | 2 | 1 (50%) | |
| Kosse | 96.64° W, 31.29° N | 14 | 2 | 1 (50%) | |
| Richland | 96.43° W, 31.91° N | 8 | 3 | 3 (100%) | |
| Rogers | 97.22° W, 30.93° N | 21 | 5 | 1 (20%) | |
| Karnes City | 97.85° W, 28.92° N | 21 | 2 | 1 (50%) |
| Item | Domain 1 | Domain 2 |
|---|---|---|
| location | 31.16° N, 99.21° W | 31.16° N, 99.21° W |
| the number of grid points | 100 × 96 | 256 × 232 |
| distance between grid points (km) | 17 | 5.667 |
| layers | 45 | 45 |
| map projection | lambert | lambert |
| microphysics scheme | WSM3 | WSM3 |
| longwave radiation scheme | RRTM | RRTM |
| shortwave radiation scheme | Dudhia | Dudhia |
| surface layer scheme | Monin-Obukhov | Monin-Obukhov |
| land/water surface scheme | Noah | Noah |
| planetary boundary layer scheme | YSU | YSU |
| cumulus parameterization | Kain-Fritsch (new Eta) | Kain-Fritsch (new Eta) |
| simulated time | every 24 h | every 24 h |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, Q.-L.; Hu, G.; Westbrook, J.K.; Sword, G.A.; Zhai, B.-P. An Advanced Numerical Trajectory Model Tracks a Corn Earworm Moth Migration Event in Texas, USA. Insects 2018, 9, 115. https://doi.org/10.3390/insects9030115
Wu Q-L, Hu G, Westbrook JK, Sword GA, Zhai B-P. An Advanced Numerical Trajectory Model Tracks a Corn Earworm Moth Migration Event in Texas, USA. Insects. 2018; 9(3):115. https://doi.org/10.3390/insects9030115
Chicago/Turabian StyleWu, Qiu-Lin, Gao Hu, John K. Westbrook, Gregory A. Sword, and Bao-Ping Zhai. 2018. "An Advanced Numerical Trajectory Model Tracks a Corn Earworm Moth Migration Event in Texas, USA" Insects 9, no. 3: 115. https://doi.org/10.3390/insects9030115
APA StyleWu, Q.-L., Hu, G., Westbrook, J. K., Sword, G. A., & Zhai, B.-P. (2018). An Advanced Numerical Trajectory Model Tracks a Corn Earworm Moth Migration Event in Texas, USA. Insects, 9(3), 115. https://doi.org/10.3390/insects9030115