1. Introduction
Drosophila suzukii Matsumura 1931 (Diptera: Drosophilidae), commonly called spotted wing
Drosophila, is an invasive insect pest of soft-skinned fruits native to Southeast Asia. As a highly polyphagous pest species, adults are attracted to a wider range of odors of ripening and ripe fruits than most drosophilids [
1]. In contrast to most Drosophilidae, female
D. suzukii possess a saw-like sclerotized ovipositor, enabling oviposition into healthy, ripening and ripened fruits [
2,
3], resulting in heavy economic losses to fruit growers [
4,
5]. Many economically important soft and stone fruits (e.g., raspberries and blackberries:
Rubus spp., strawberries:
Fragaria ananassa, cherries:
Prunus avium,
P. cerasus, plums:
P. domestica, elderberry:
Sambucus spp.), some susceptible grapevine varieties, as well as numerous wild or ornamental fruits (e.g.,
P. serotina,
Viscum album,
Rubus spp.,
Basella alba, [
6,
7,
8,
9,
10,
11] are known hosts. While a high number of reproduction hosts is known from fruit sampling and lab studies, the complete feeding ecology is yet to be described. Early attempts to identify the ingested plant DNA and microbes in the guts of adult individuals have been successful [
12,
13]. Further studies using these techniques will help to understand the whole feeding ecology of this pest species.
Continental USA (California) and Southern Europe (Italy and Spain) recorded
D. suzukii in 2008 for the first time [
2,
14,
15]. It spread rapidly the following years, with periodic reports of economic damage in cultivated soft and stone fruits around the world [
4,
5,
6].
In 2011, the Julius Kühn-Institut (JKI) established a monitoring program in southwest Germany in the vicinity of the JKI in Dossenheim, Siebeldingen and Darmstadt expecting the arrival of
D. suzukii. Monitoring traps were randomly installed in different fruit crops, forested areas and hedges, in experimental fields and in the direct vicinity of the institutes. First recorded observations of
D. suzukii in Germany date from late summer 2011 from private observations in Bavaria and from the JKI monitored traps [
16,
17].
In spring 2016, the JKI launched the online platform DrosoMon (
http://drosomon.julius-kuehn.de/), which serves as a database and visualization platform for monitoring the occurrence and activity of
D. suzukii. Currently, several other institutes and plant protection services from all over Germany, as well as institutions from other European countries have joined this project. DrosoMon helps with the harmonization of the datasets and simplifies the comparability of trap data between monitoring systems. DrosoMon started with the data of the three JKI sites located in the northern part of the upper Rhine valley, an important fruit- and wine-growing region. Many of the fruit crops are susceptible to
D. suzukii infestation, and the resulting damages lead to substantial economic losses. This region is characterized by small-scale agriculture thriving under the humid Atlantic climate with warm summers and moderate winters [
18,
19] and provides diverse habitats and retreats for
D. suzukii. The climate differs significantly from that of other fruit- and wine-growing regions like the Pacific Northwest with hot dry summers and cold winters or the coastal climates of North America with their mild winters [
20,
21,
22,
23,
24,
25]. However, it is quite similar to the moderate summer and winter climate of Northern Italy, where
D. suzukii’s presence since 2009 has been related to episodic damages to fruit harvests [
25,
26,
27,
28].
When
D. suzukii was first recorded in Europe, it was suggested that cold winters and hot dry summers are unsuitable for this species [
15,
21]. Consequently, it was supposed that only low numbers of individuals might overwinter at sheltered sites [
3,
6]. Previously published studies already showed that
D. suzukii is well adapted to various weather conditions [
29] and defined weather indices for their development, mortality and population dynamics [
20,
27,
30,
31]. Seasonal dimorphism of winter and summer morphs has been put forth as an adaptation to survive hot and dry summers and overwinter colder winters [
6,
32].
In the present study, a number of the above-mentioned weather indices were evaluated for their impacts on D. suzukii trap captures in the upper Rhine Valley. We describe for the first time the seasonal trap-activity of D. suzukii in various habitats of the highly diverse and fragmented upper Rhine valley.
The objectives of this study were (i) to study the spatio-temporal trap activity of D. suzukii in cropped and wild habitats, (ii) the relationship between land use and the seasonal and annual capture rates and (iii) the impact of weather on capture rates.
3. Results
The number of traps monitored varied across the years from 25 in 2011 to 93 in 2016–2017 (
Table 4). The traps were exposed for a total of 116,602 days (min = 4, max = 48) and captured 756,768 adult
D. suzukii individuals. The first captures of
D. suzukii during the monitoring were between September and November, 2011 in a plum orchard, in the experimental field in Dossenheim (10 ♂, 1 ♀) and in a vineyard in Siebeldingen (3 ♂, 1 ♀). The JKI Dar captured the first individuals (12 ♂, 6 ♀) in August 2013 in a commercial cherry orchard near Nieder-Beerbach, Germany.
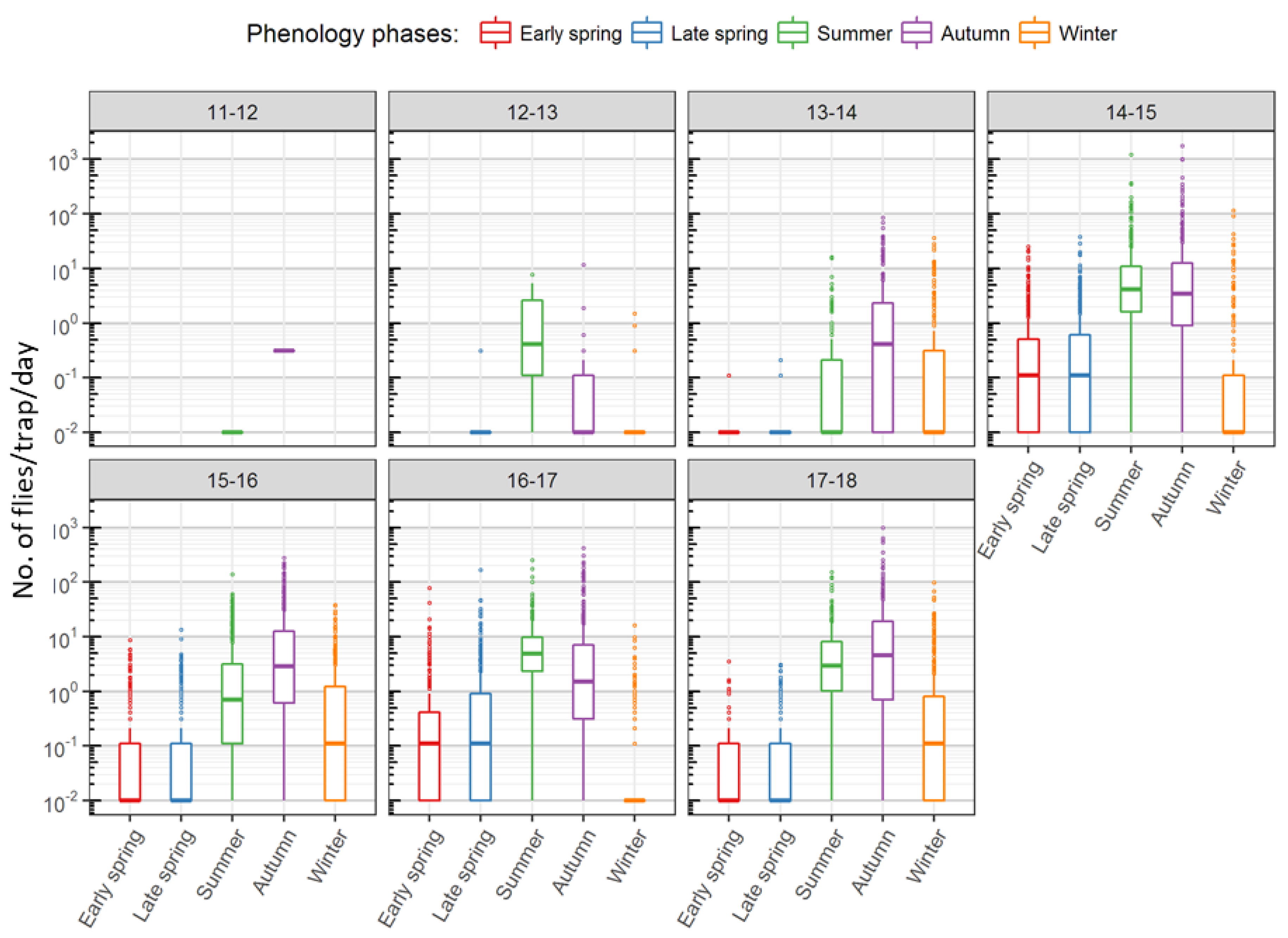
The mean annual trap activity followed population build-up starting in late spring and reaching a peak in summer. Forests and hedges recorded a second peak in autumn, after a brief decline in October (
Figure 3). Capture rates steadily declined in winter to the very low numbers seen in early spring and the first half of late spring with a brief increase in April (
Figure 3 and
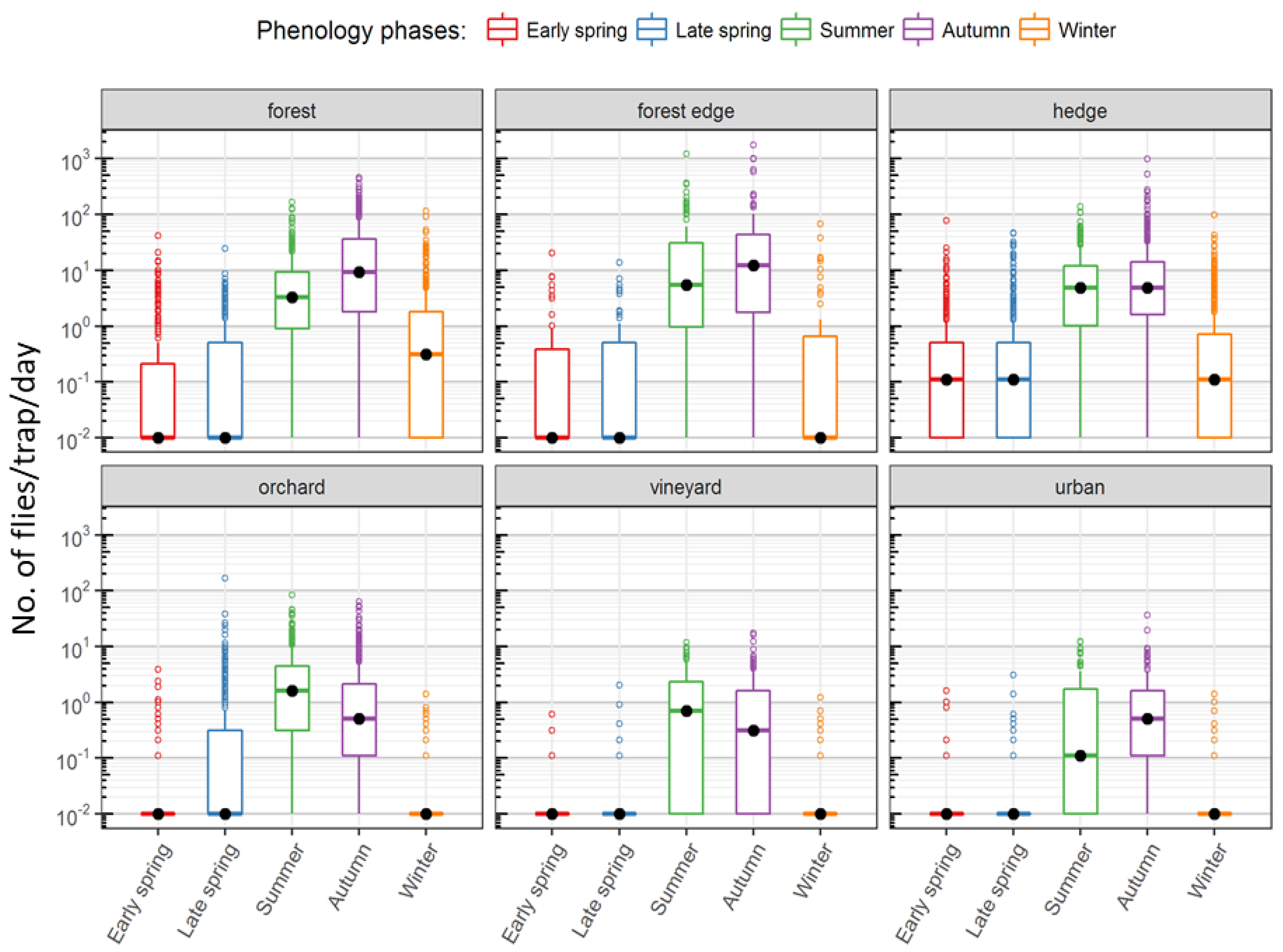
Figure S3). They also varied greatly between the sampled habitat types. Traps in forests, hedges and forest borders recorded the highest numbers, in peak seasons, over 1000 flies per day. In contrast, traps in fruit crops and vineyards recorded no more than a maximum of 50–100 flies per day (
Figure 4). The plot of all trap captures together showed a distinct two-peak dynamic with higher captures in summer and autumn. Upon closer scrutiny, the two-peak dynamic is to be seen only in the case of forested areas and hedge traps and also not in all years, while those in vineyards and orchards show a decline already from the onset of cold weather (
Figure 3). Though the highest number of captures in forested areas and hedges was in autumn, in some years, the median captures in summer were higher than in autumn (
Figure 5). In most years, traps in forests and hedges captured consistently more flies in autumn than in summer, whereas traps in orchards captured more flies in summer than in autumn. Hedges and forests accounted for a much higher share of winter captures compared to other locations (
Figure 4 and
Figure S4).
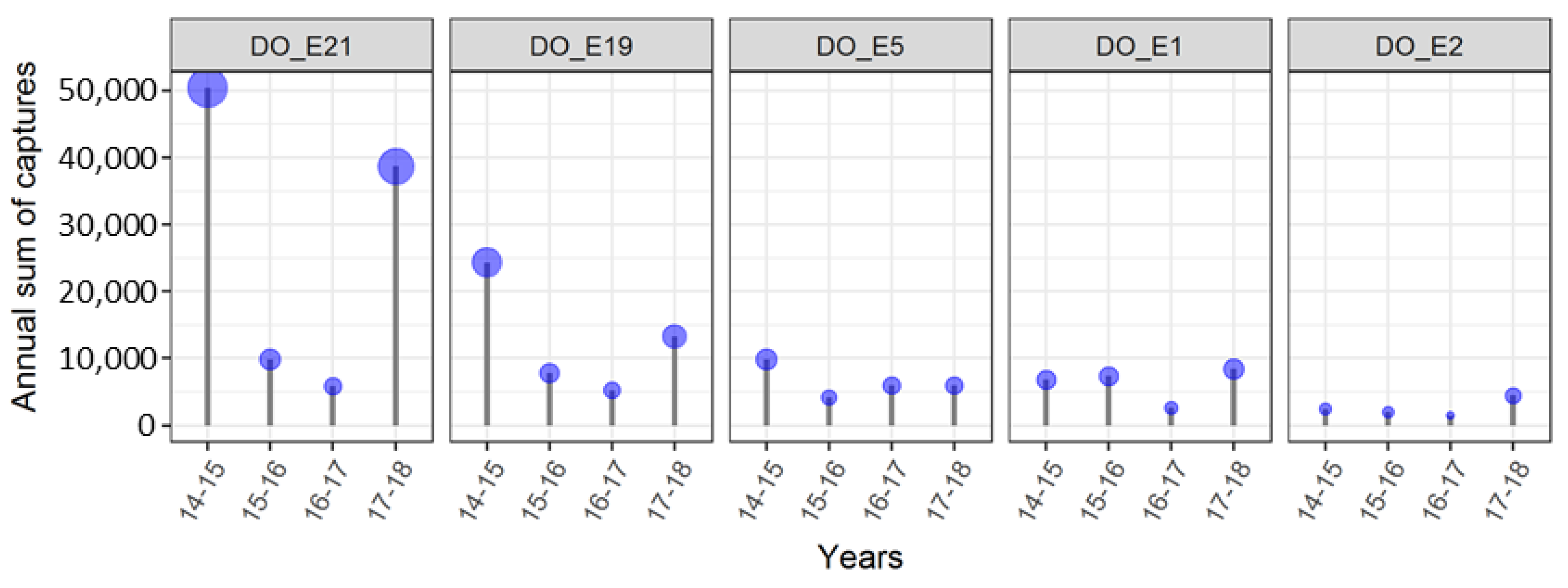
Five traps with very high capture rates were selected for a detailed analysis of the effects of land use on their capture rates (
Figure 6 and
Table S1). These traps were located on the borders of the Forest of Odes next to the Rhine valley with the highest captures in 2014–2015 and 2017–2018. Two traps (DO_E21 and DO_E19) were located prominently on a small mountain ledge, close to orchards/vineyards and the forest. Both traps captured between five- and 10-times more individuals over the years than trap DO_E5, which is also close to orchards/vineyards and shrubberies. The trap DO_E1 on the forest border, next to the urban area (Dossenheim) and a quarry captured less individuals than the above-mentioned traps, whereas the trap DO_E2, located deeper in the woods with only trees and no shrubberies nearby, and hence farther than the others from forest borders, orchards or urban areas, captured the lowest numbers.
Overall, an equal number of females and males were captured (50.9:49.1%), with small variations over the seasons. Slightly more females were captured in early spring, late spring and winter compared to summer or autumn (
Table 5). The pattern was fairly similar across the different locations except in vineyards where more females than males were captured in summer.
Though
D. suzukii was observed already in 2011, the next two years, 2012 and 2013, registered far less captures compared to the years after. Following the winter of 2013–2014, the warmest in the period with zero days below freezing, there was a comparable surge in the captures afterwards, never reaching values as low as those in 2012 and 2013. For studying the relationship between weather and captures, only capture data from 2013 onwards were used as
D. suzukii appears to have successfully established in the monitored areas during the warm winter of 2013–2014. While the low temperatures in autumn were not accompanied by a corresponding change in capture rates, the higher number of ice days in the winter of 2016/2017 was reflected in the lowest winter captures from this year (
Figure 7).
During the monitoring period, the summer of 2017 was the hottest followed by that of 2015, resulting in the low late spring capture rates compared to those of 2014 and 2016. The 2014 and 2016 late springs also received more precipitation than those of 2015 or 2017 (
Figure 8). The low summer captures in 2015 also corresponded to the high number of days when maximum temperature exceeded 30 °C this year (
Figure 9).
The significant parameters from regressing seasonal captures against weather parameters are given in
Table 6. Both late spring and summer captures were negatively associated with maximum temperatures above 30 °C. Temperatures in the range of 20 °C–30 °C defined as optimal weather for
D. suzukii were positively associated with captures in late spring, but negatively associated for summer captures. Both frost and ice days were correlated with a decrease in capture rates in the cold seasons, while daily mean temperature (mean) and precipitation (sum) were positively correlated.
4. Discussion
In this study, we analyzed the monitoring data of the JKI in the upper Rhine valley in southwest Germany. The study started a few months before D. suzukii appeared for the first time in Germany, in August 2011, and followed its establishment in the succeeding years.
Drosophila suzukii has been present since at least late 2011 in southwest Germany. Currently, it is widespread and overwinters successfully in the upper Rhine valley. We suppose that the mild winter of 2013–2014 helped to establish a stable population in this part of Germany. The trap activity of this region was comparable to Northern Italy with a similar climate [
27,
28].
We split the typical annual population cycle of D. suzukii into five phenology phases (early spring, late spring, summer, autumn and winter) to better identify factors that affect the different phases of trap activity. Both weather conditions and habitat type influenced trap activity. The influence of habitat type on the trap captures is manifested in the regular, seasonal changes in the trap activity of D. suzukii.
The summer increase of trap captures and the following peak coincide with the ripening of most wild and cultivated host fruits of
D. suzukii [
11,
41]. The single annual peak in summer in orchard and vineyard traps suggests that those sites are not suitable overwintering habitats and
D. suzukii has to re-immigrate each year, whereas forested areas appear to be the favored habitat year-round. The increase of trap captures from late spring onwards until the autumn peak in forests, forest borders and hedges has also been noted in other temperate regions [
20,
22,
24,
42]. The single peak in late spring/summer in fruit crops and vineyards is quite similar to the findings in northwestern Italy [
28]. We suppose that when attractive food sources are present, the traps are less attractive to the flies, such as in orchards, but in forests, the same baits are more attractive to the flies due to less competition between ACV-baited traps and attractive food sources. Further, forests may provide better habitat conditions when it is too hot in summer or too cold in winter. The presence of berries in hedges or in the forested areas [
11,
36,
43] at various stages of ripeness partly all throughout the year could be an additional factor why forested areas and hedges always recorded very high capture rates and microorganisms growing on the surface of evergreen trees in the cold bottleneck seasons may also serve as food resources, supporting
D. suzukii populations [
13]. Thus, we cannot conclude whether higher trap captures in forests result from a higher
D. suzukii abundance or a higher attractivity of traps. The winter decline and low captures in early spring have also been noted in California [
22,
44], northern Italy [
28,
45] and Canada [
20].
The five traps at the forest border of the Forest of Odes suggest a distinct importance of forest borders to
D. suzukii. Traps with the highest captures, located prominently on a small mountain ledge, were especially attractive in autumn, when most trees in orchards and vineyards have already lost their leaves. We assume that
D. suzukii migrates from orchards/vineyards in the plains to forested areas in autumn in search of nutrition and overwintering habitats. DO_E21, in particular, offers such habitats since it is located on a
Robinia pseudoacacia overgrown with
Hedera helix. Our findings are in agreement with Tait et al. (2018) [
46] that
D. suzukii migrates between different habitats, depending on the season, searching for suitable overwintering habitats. Since DO_E2 captured less individuals than DO_E1, we assume that
D. suzukii migrates into sheltered overwintering sites such as hedges and forested areas as long as it can find shrubbery. The attractiveness of the traps appears to be strongly influenced by the immediate habitat and the seasonal changes in the habitat with respect to the availability of nourishment and shelter. We recommend taking this into consideration when estimating in-field fly density based on trap captures or when trap capture data are treated as representative of fly density.
Based on our seven-year monitoring data with much higher capture rates in forests than in fruit orchards or vineyards even in the fruiting season, we question the reliability of the estimation of the actual density of
D. suzukii in the field using ACV traps. When natural food sources are available, the flies seem to prefer the ripe fruits markedly more than the ACV mixture, resulting in an under-representation of the fly density in the fruit orchards and vineyards during the fruiting season. For economic reasons and to have comparable data over the monitoring period, we did not change our trap designs during the study. Our trap designs may have resulted in relatively conservative estimates of the abundance compared to other traps and/or baits that are now available [
47,
48,
49]. However, it was already suggested that every individual fly captured in an ACV-baited trap represents 198 flies in a 2.7-ha cherry orchard [
50]. These findings strengthen the argument that trap captures should be interpreted with caution [
5,
29,
44,
51].
We tested the capture data against temperature thresholds that were found to affect
D. suzukii individuals and populations significantly in field and laboratory studies. The decline in trap captures when tmax > 30 °C in late spring was noted previously [
22,
30]. Further, it was already suggested that in lab assays, temperatures above 30 °C reduce reproduction or even inhibit it [
52,
53]. Although our weather data were of quite high resolution (1 × 1 km
2), it would be impossible to simulate the authentic field conditions that could vary in much smaller units that provide suitable shelters for
D. suzukii. We also found that the years with high late spring captures correlated with high precipitation. Whether this is due to the effect of precipitation on
D. suzukii host plants or the higher relative humidity favoring
D. suzukii was beyond the scope of our study.
Since the climate in southwestern Germany is more akin to that of Northern Italy [
28] than to California [
22] or Canada [
20], we expected
D. suzukii population dynamics to be similar to that observed in northern Italy. The winter of 2013/2014, with zero days below freezing and only a few cold days, was the mildest during the whole monitoring period. Survival success during this mild winter could have played a key role in the excessive damages observed in the following spring in this region. Next, low temperatures in autumn do not seem to be correlated with low capture rates, whereas ice-days in winter are followed by a decrease in trap captures over a longer period (e.g., winter of 2016/2017). In contrast to previously published findings [
20], we did not find any conclusive relationships between mild winter days and earlier fly appearance in the traps the following spring.
Both females and males were captured all throughout the year, but the number of males was slightly higher during the peak seasons (summer and autumn), and previously published studies showed less attractiveness of ACV-baited traps to female
D. suzukii the more mature eggs their ovaries contained [
1,
54]. However, we assume that a combination of this and a higher dispersal of males to ensure gene flow results in higher capture rates of males during the peak seasons. More females were captured in the colder seasons when trap captures were lower, suggesting the better adaptability of female winter morphs to colder periods with insufficient food sources. The higher cold tolerance of females was observed in many other studies, as well [
20,
21,
42,
55,
56]. However, reports are not conclusive. Ryan et al. [
52] did not find any difference between sexes in their response to low temperatures, while Enriquez and Colinet [
57] even suggested that males are better adapted to survive through winters.
5. Conclusions
Through our monitoring program we were able to record the first appearance of D. suzukii in Germany in 2011 and follow its establishment over the following years. Despite its first occurrence in 2011, it was the mild winter of 2013/2014 that helped D. suzukii the firm establishment in southwest Germany. The successful establishment in this region is proof of its adaptability to cold winters, hot summers, and fluctuating temperatures, in a small structured landscape.
Further, our analysis confirms that temperature thresholds for in-field development and survival should be re-adjusted with respect to further meteorological (e.g., global radiation) and habitat parameters as conclusions based on laboratory experiments cannot be easily juxtaposed with those based on field studies. While laboratory studies are carried out in a controlled environment, numerous factors are at play in field studies. Next, trap captures do not reflect actual in-field fly densities and we still lack information about realistic in-field population densities since the attractiveness of ACV baited traps varies significantly between habitats and seasons.
A detailed analysis of the effect of landscape on D. suzukii captures and determination of weather conditions was beyond the scope of the study. However, it must be noted that the traps were spatially quite clustered and did not map large variations in weather. An analysis of a country-wide monitoring is necessary to validate the results on a bigger scale.

 ,
, 









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}