Food Reward and Distance Influence the Foraging Pattern of Stingless Bee, Heterotrigona itama

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Experimental Design
2.2.1. Experiment 1: Sugar Concentration Preference
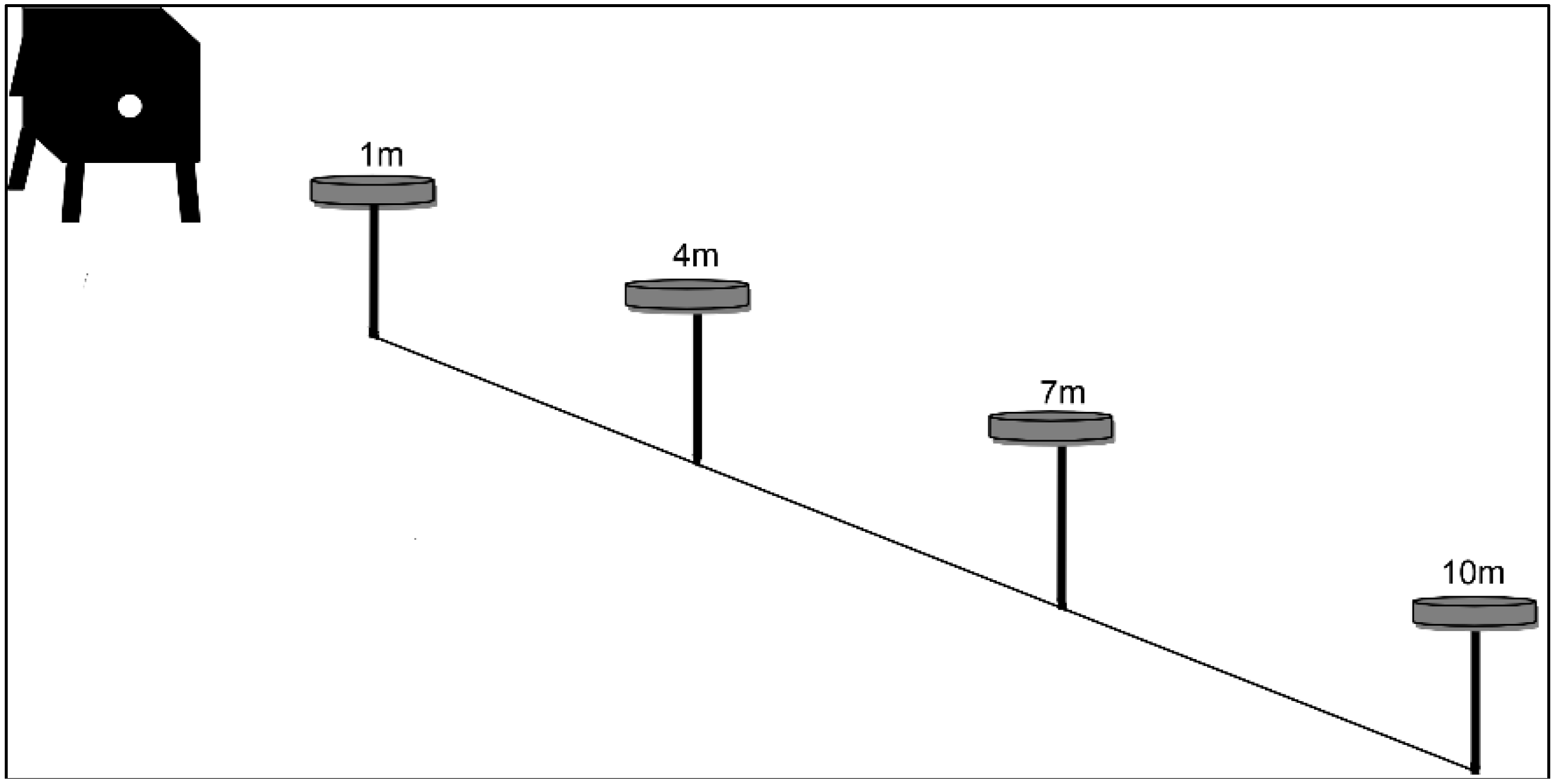
2.2.2. Experiment 2: Foraging Distance
2.3. Statistical Analysis
3. Results
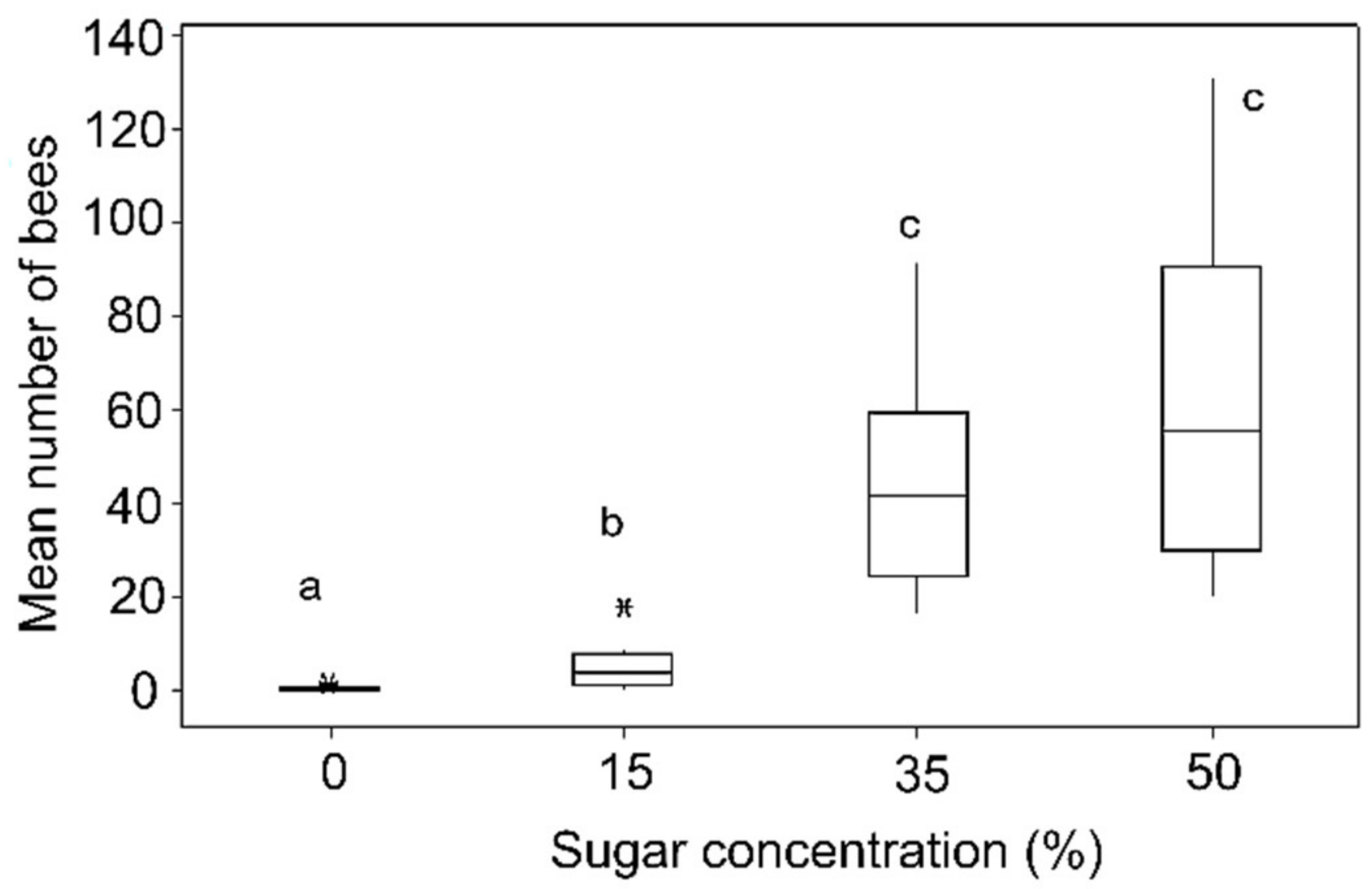
3.1. Sugar (Sucrose) Concentration Preference
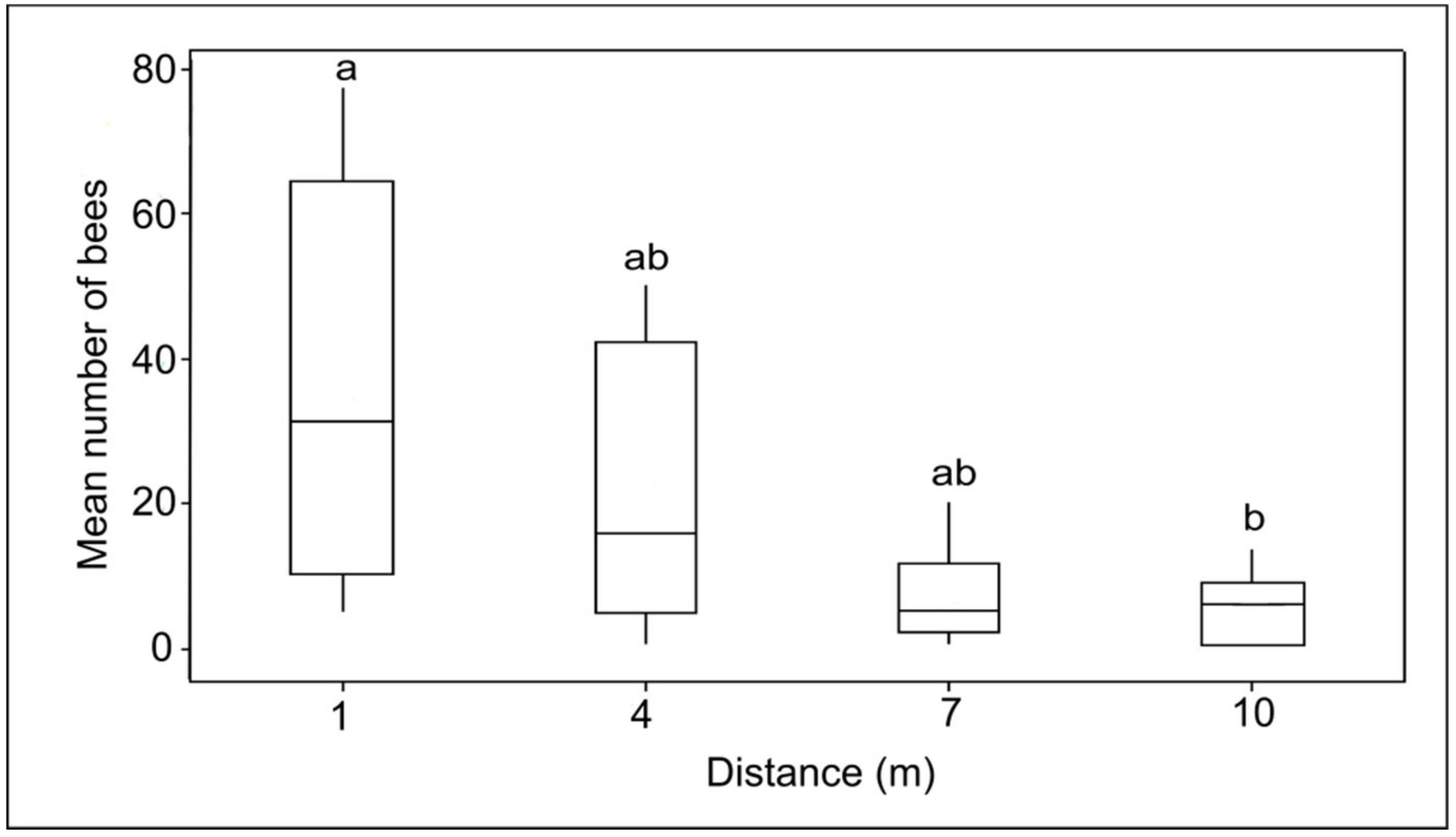
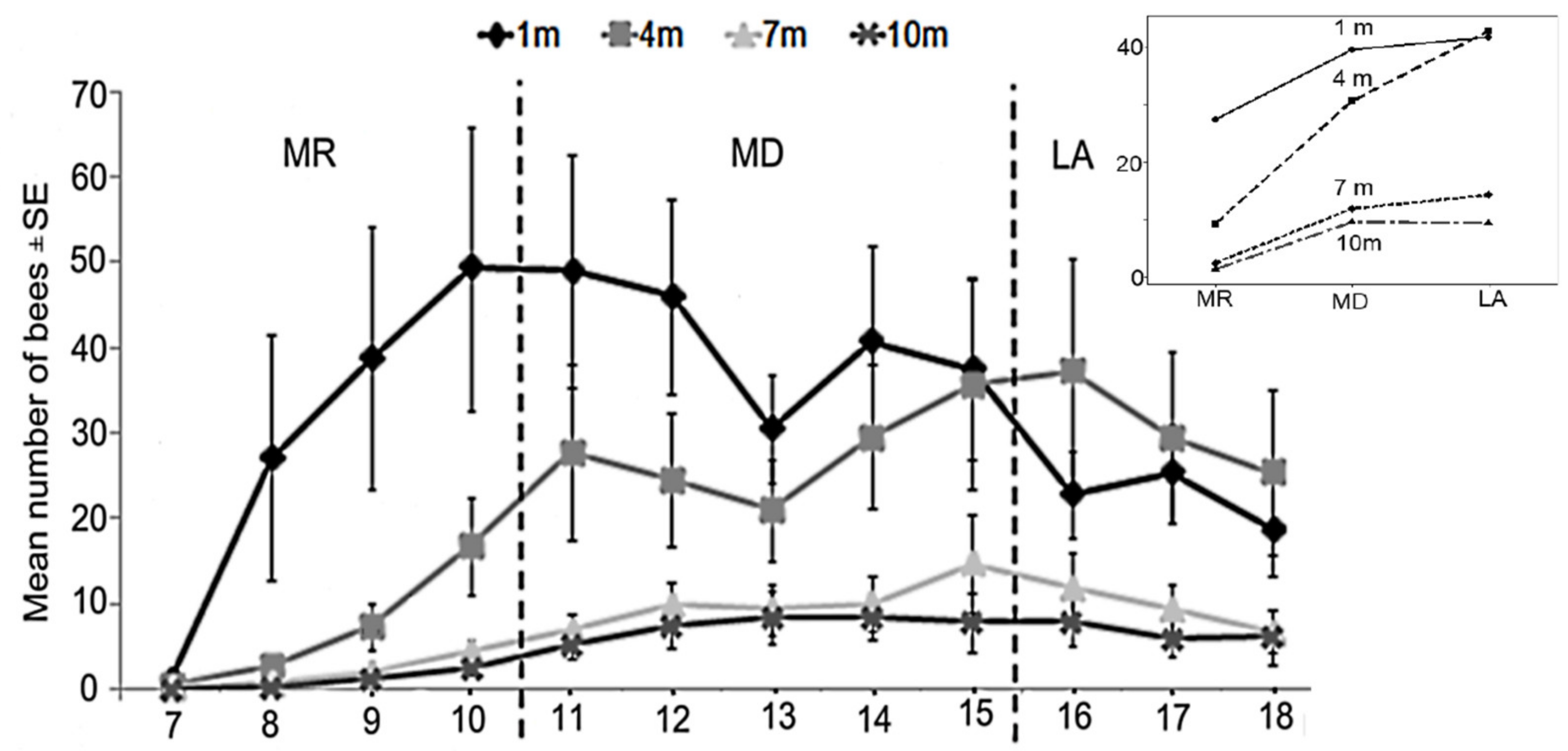
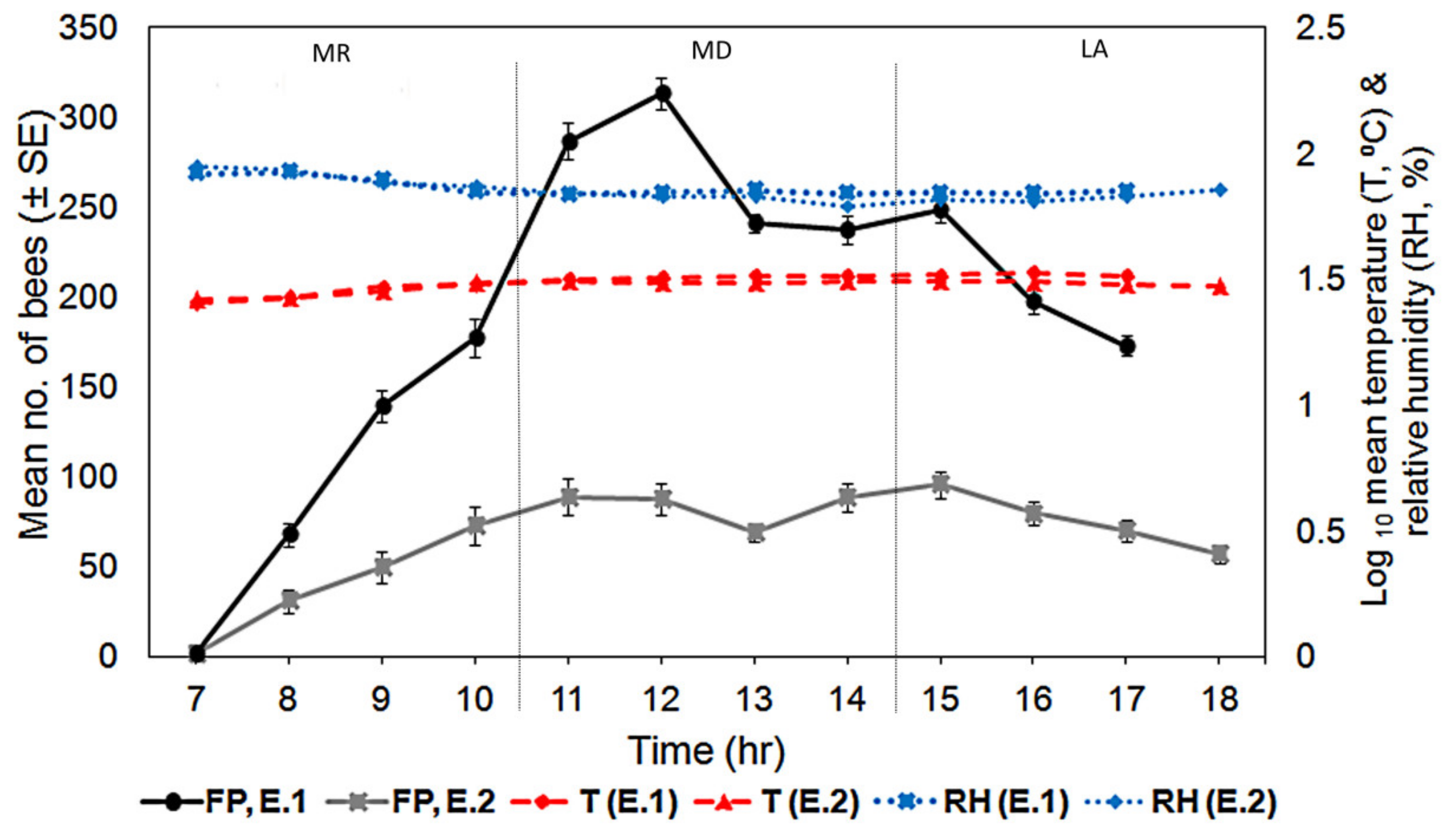
3.2. Food Distance
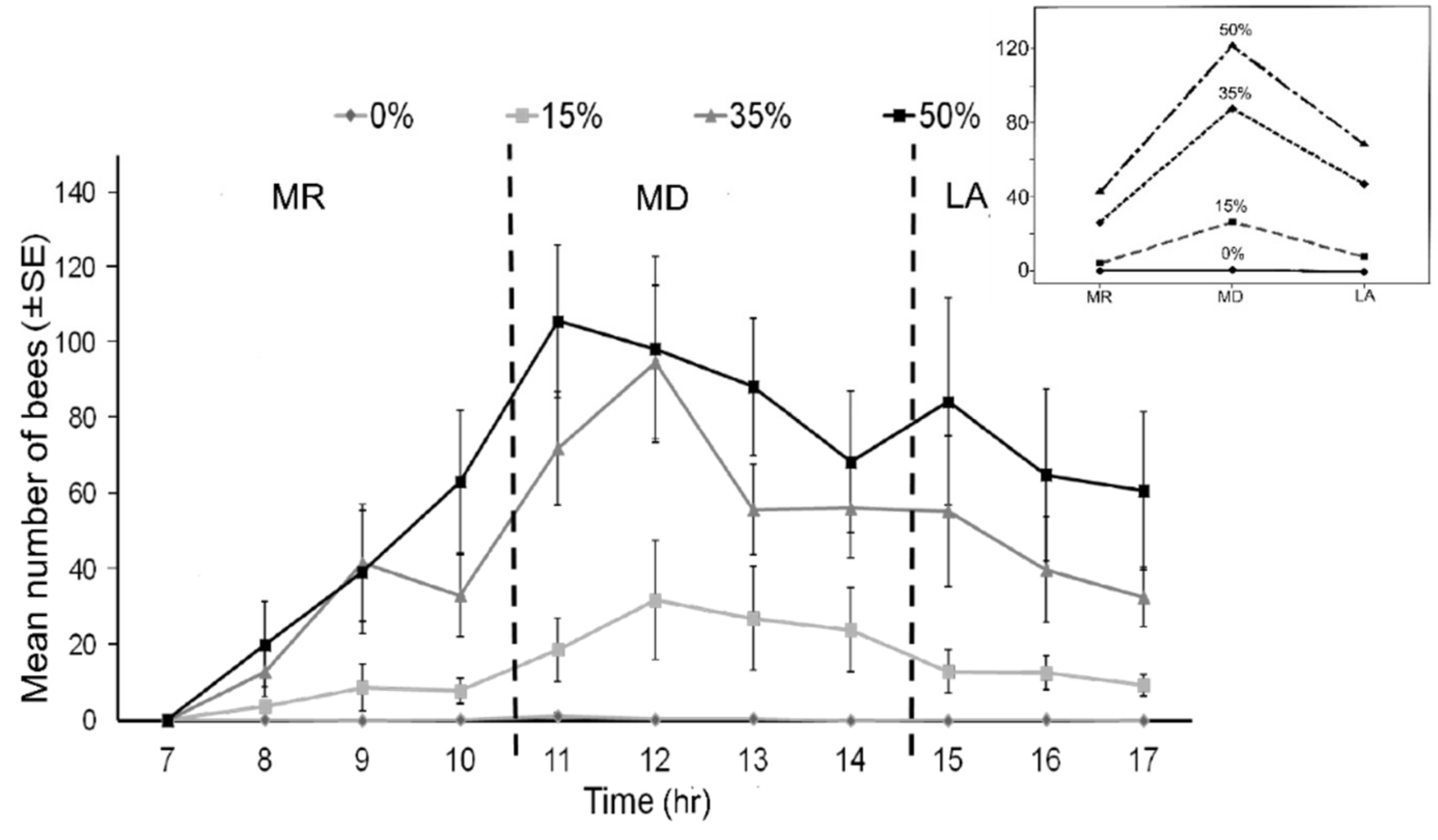
3.3. Foraging Pattern
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Jarau, S.; Hrncir, M.; Zucchib, R.G.; Bartha, F. Recruitment behavior in stingless bee, Melipona scutellaris and M. quadrifasciata. L. Foraging at food sources differeing in direction and distance. Apidologie 1999, 31, 81–91. [Google Scholar] [CrossRef]
- Nieh, J.C. Recruitment communication in stingless bee (Hymenoptera, Apidae, Meliponini). Apidologie 2004, 35, 159–182. [Google Scholar] [CrossRef]
- Layek, U.; Karmakar, P. Nesting characteristics, floral resources, and foraging activity of Trigona iridipennis Smith in Bankura district of West Bangal, India. Insectes Soc. 2017, 65, 117–132. [Google Scholar] [CrossRef]
- Heard, T.A. The role of stingless bee in crop pollination. Ann. Rev. Entomol. 1999, 44, 183–206. [Google Scholar] [CrossRef] [PubMed]
- Nunes-Silva, P.; Hilario, S.D.; de Souza Snatos Filho, P.; Imperatriz-Fonseca, V.L. Foraging activity in Plebeia remota, a stingless bee species, is influenced by reproductive stat of colony. Psyche 2010. [Google Scholar] [CrossRef]
- Roubik, D.W. Ecology and Natural History of Tropical Bees; Cambridge University Press: Cambridge, UK, 1989; 514p. [Google Scholar]
- Jalil, A.H. Beescape for Meliponines: Conservation of Indo-Malayan Stingless Bees; Partridge Publishing: Singapore, 2014; p. 214. [Google Scholar]
- Rasmussen, C.; Cameron, S.A. Global stingless bee phylogeny supports ancient divergence, vicariance, and long distance dispersal. Biol. J. Linn. Soc. 2010, 99, 206–232. [Google Scholar] [CrossRef]
- Jaapar, M.F.; Halim, M.; Mispan, M.R.; Jajuli, R.; Saranum, M.M.; Zainuddin, M.Y.; Ghani, R.; Abdul Ghani, I. The diversity and abundance of stingless bee (Hymenoptera: Meliponini) in Peninsular Malaysia. AEB 2016, 10, 1–7. [Google Scholar]
- Kelly, N.; Farisya, M.S.N.; Kumara, T.K.; Marcela, P. Species diversity and external best characteristics of stingless bees in Meliponiculture. Pertanika J. Trop. Agric. Sci. 2014, 37, 293–298. [Google Scholar]
- Kajobe, R. Botanical sources and sugar concentration of nectar collected by two stingless bee species in tropical African rainforest. Apidologie 2007, 38, 110–121. [Google Scholar] [CrossRef]
- Nieh, J.C.; Sánchez, D. Effect of food quality, distance and height on thoracic temperature in the stingless bee Melipona panamica. J. Exp. Biol. 2005, 208, 3933–3943. [Google Scholar] [CrossRef] [PubMed]
- Ciar, R.R.; Bonto, L.S.; Bayer, M.H.P.; Rabajante, J.F.; Lubag, S.P.; Fajardo, A.C.; Cervancia, C.R. Foraging Behavior of Singles Bees Teragonula Biroi Friese: Distance, Direction and Height of Preferred Food Source; Cornell University Library: Ithaca, NY, USA, 2013. [Google Scholar]
- Cortopassi-Laurino, M.; Imperatriz-Fonseca, V.L.; Roubik, D.W.; Dollin, A.; Heard, T.; Aguilar, I.B.; Venturieri, G.C.; Eardley, C.; Noguiera-Neto, P. Global meliponiculture: Challenges and opportunities. Apidologie 2006, 37, 1–18. [Google Scholar] [CrossRef]
- Chidi, O.H.; Odo, P.E. Meliponiculture for sustainable economy. In Proceeding of the 4th Delta State University Faculty of Science International Conference, Abraka, Delta State, Nigeria, 6 July 2017; pp. 131–137. [Google Scholar]
- Roubik, D.W.; Yanega, D.; Aluja, M.S.; Buchmann, S.L.; Inouye, D.W. On optimal nectar foraging by some tropical bees (Hymenoptera: Apidae). Apidologie 1995, 26, 197–211. [Google Scholar] [CrossRef]
- Roubik, D.W.; Buchmann, S.L. Nectar selection by Melipona and Apis mellifera (Hymenoptera: Apidae) and the ecology of nectar intake by bee colonies in a tropical forest. Oecologia 1984, 61, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Biesmeijer, J.C.; Ermers, M.C.W. Social foraging in stingless bees: How colonies of Melipona fasciata choose among nectar sources. Behav. Ecol. Sociobiol. 1999, 46, 129–140. [Google Scholar] [CrossRef]
- Barbosa, M.F.; Campos, L.A.D.O.; Paixao, J.F.D.; Alves, R.M.D.O. Foraging pattern and harvesting of resources of subterranean stingless bee: Geotrigona subterranean (Friese, 1901) (Hymenoptera: Apidae: Meliponini). Pap. Avulos. Zool. 2016, 56, 151–157. [Google Scholar] [CrossRef]
- Jaapar, M.F.; Jajuli, R.; Mispan, M.R.; Abd Ghani, I. Foraging Behavior of Stingless Bee Heterotrigona itama (Cockerell, 1918) (Hymenoptera: Apidae: Meliponini). AIP Conf. Proc. 2018, 1940. [Google Scholar] [CrossRef]
- Macías-Macías, J.O.; Tapia-Gonzalez, J.M.; Contreras-Escareño, F. Foraging behavior, environmental parameters and nests development of Melipona colimana Ayala (Hymenoptera: Meliponini) in temperate climate of Jalisco, México. Braz. J. Biol. 2017. [Google Scholar] [CrossRef] [PubMed]
- Heard, T. The Australian Bee Book; Sugarbag Bees: Brisbane, Queensland, Australia, 2016. [Google Scholar]
- Schmidt, V.M.; Zucchi, R.; Barth, F.G. Recruitment in a scent trail laying stingless bee (Scaptotrigona aff. depilis): Changes with reduction but not with increase of the energy gain. Apidologie 2006, 37, 487–500. [Google Scholar] [CrossRef]
- Kim, W.; Gilet, T.; Bush, J.W.M. Optimal concentrations in nectar feeding. Proc. Natl. Acad. Sci. USA 2011, 108, 16618–16621. [Google Scholar] [CrossRef] [PubMed]
- Kuhn-Neto, B.; Contrera, F.A.L.; Castro, M.S.; Nieh, J.C. Long distance foraging and recruitment by stingless bee Melipona mandacaia. Apidologie 2009, 40, 472–480. [Google Scholar] [CrossRef]
- Nicolson, S.W.; de Veer, L.; Köhler, A.; Pirk, C.W.W. Honeybees prefer warmer nectar and less viscous nectar, regardless of sugar concentration. Proc. R. Soc. B 2013, 280, 20131597. [Google Scholar] [CrossRef] [PubMed]
- Beekman, L.M.; Ratnieks, F.L.W. Long range foraging by honey bee, Apis mellifera. Funct. Ecol. 2000, 14, 490–496. [Google Scholar] [CrossRef]
- Smith, J.P.; Heard, T.A.; Beekman, M.; Gloag, R. Flight range of the Australian stingless bee Tetragonula carbonaria (Hymenoptera: Apidae). Austral Entomol. 2017, 56, 50–53. [Google Scholar] [CrossRef]
- Yucel, B.; Duman, I. Effects of foraging activity of honeybees (Apis mellifera L.) on onion (Allium cepa) seed production and quality. Pak. J. Biol. Sci. 2005, 8, 123–126. [Google Scholar]
- Danaraddi, C.S.; Hakkalappanavar, S.; Biradar, S.B.; Tattimani, M.; Vinod, S.K. Studieson foraging behaviour of stingless bee, Trigona iridipennis Smith at Dharwad, Karnataka. Int. J. For. Crop Improv. 2011, 2, 163–169. [Google Scholar]







© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Basari, N.; Ramli, S.N.; Mohd Khairi, N.'.S. Food Reward and Distance Influence the Foraging Pattern of Stingless Bee, Heterotrigona itama. Insects 2018, 9, 138. https://doi.org/10.3390/insects9040138
Basari N, Ramli SN, Mohd Khairi N'S. Food Reward and Distance Influence the Foraging Pattern of Stingless Bee, Heterotrigona itama. Insects. 2018; 9(4):138. https://doi.org/10.3390/insects9040138
Chicago/Turabian StyleBasari, Norasmah, Sarah Najiah Ramli, and Nur 'Aina Syakirah Mohd Khairi. 2018. "Food Reward and Distance Influence the Foraging Pattern of Stingless Bee, Heterotrigona itama" Insects 9, no. 4: 138. https://doi.org/10.3390/insects9040138
APA StyleBasari, N., Ramli, S. N., & Mohd Khairi, N. '. S. (2018). Food Reward and Distance Influence the Foraging Pattern of Stingless Bee, Heterotrigona itama. Insects, 9(4), 138. https://doi.org/10.3390/insects9040138