


Epigenomes, Volume 2, Issue 4 (December 2018) – 5 articles
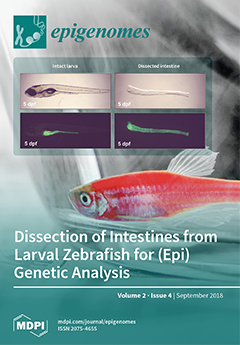
Cover Story (view full-size image):
The left panel of this figure shows a five-day-old zebrafish larva that expresses GFP specifically in the intestine. By using tweezers, the intestine can be removed from the body and non-intestinal tissue can be subsequently removed. The right panel of the figure shows the resulting isolated intestine. This isolated tissue is the starting point to study the genetic and epigenetic regulation of intestinal development. View this paper.
- Issues are regarded as officially published after their release is announced to the table of contents alert mailing list.
- You may sign up for e-mail alerts to receive table of contents of newly released issues.
- PDF is the official format for papers published in both, html and pdf forms. To view the papers in pdf format, click on the "PDF Full-text" link, and use the free Adobe Reader to open them.
Previous Issue
Next Issue