
Epigenetic Features in Newborns Associated with Preadolescence Lung Function and Asthma Acquisition during Adolescence


Abstract
:1. Introduction
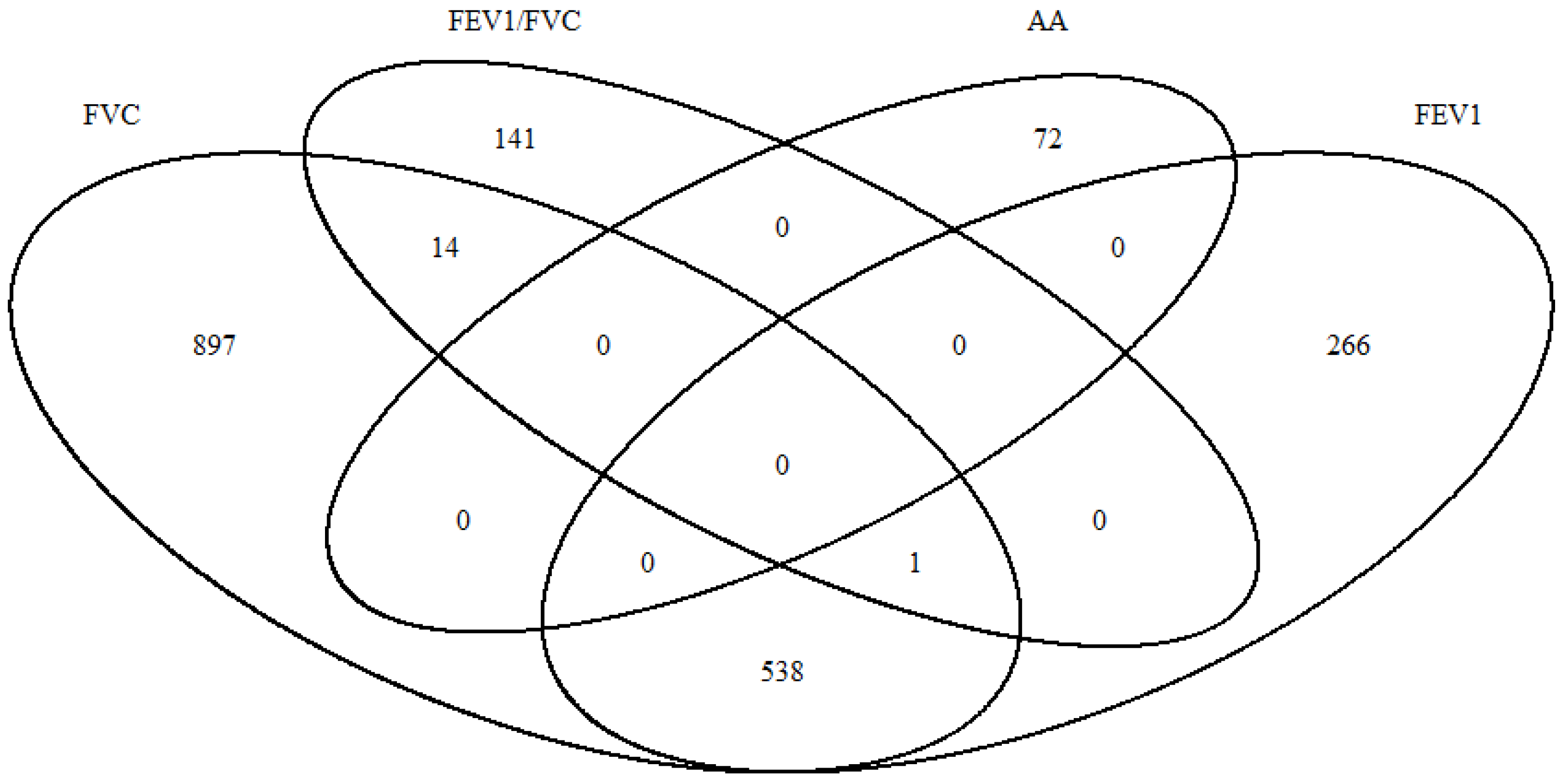
2. Results
3. Discussion
4. Materials and Methods
4.1. Study Population
4.2. Asthma Acquisition
4.3. Height Adjusted Lung Function
4.4. DNA Methylation at Birth
4.5. Statistical Analysis
4.6. Replication Study
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Peters, S.P.; Ferguson, G.; Deniz, Y.; Reisner, C. Uncontrolled asthma: A review of the prevalence, disease burden and options for treatment. Respir. Med. 2006, 100, 1139–1151. [Google Scholar] [CrossRef] [PubMed]
- Soriano, J.B.; Kendrick, P.J.; Paulson, K.R.; Gupta, V.; Abrams, E.M.; Adedoyin, R.A.; Adhikari, T.B.; Advani, S.M.; Agrawal, A.; Ahmadian, E.; et al. Prevalence and attributable health burden of chronic respiratory diseases, 1990–2017: A systematic analysis for the Global Burden of Disease Study 2017. Lancet Respir. Med. 2020, 8, 585–596. [Google Scholar] [CrossRef] [PubMed]
- Sheikhpour, M.; Maleki, M.; Vargoorani, M.E.; Amiri, V. A review of epigenetic changes in asthma: Methylation and acetylation. Clin. Epigenet. 2021, 13, 65. [Google Scholar] [CrossRef] [PubMed]
- Ho, S.-M. Environmental epigenetics of asthma: An update. J. Allergy Clin. Immunol. 2010, 126, 453–465. [Google Scholar] [CrossRef]
- Maor, G.L.; Yearim, A.; Ast, G. The alternative role of DNA methylation in splicing regulation. Trends Genet. 2015, 31, 274–280. [Google Scholar] [CrossRef] [PubMed]
- Reese, S.E.; Xu, C.-J.; Dekker, H.T.D.; Lee, M.K.; Sikdar, S.; Ruiz-Arenas, C.; Merid, S.K.; Rezwan, F.I.; Page, C.M.; Ullemar, V.; et al. Epigenome-wide meta-analysis of DNA methylation and childhood asthma. J. Allergy Clin. Immunol. 2019, 143, 2062–2074. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.-J.; Söderhäll, C.; Bustamante, M.; Baïz, N.; Gruzieva, O.; Gehring, U.; Mason, D.; Chatzi, L.; Basterrechea, M.; Llop, S.; et al. DNA methylation in childhood asthma: An epigenome-wide meta-analysis. Lancet Respir. Med. 2018, 6, 379–388. [Google Scholar] [CrossRef]
- Rathod, A.; Zhang, H.; Arshad, S.H.; Ewart, S.; Relton, C.L.; Karmaus, W.; Holloway, J.W. DNA methylation and asthma acquisition during adolescence and post-adolescence, an epigenome-wide longitudinal study. J. Pers. Med. 2022, 12, 202. [Google Scholar] [CrossRef]
- Young, R.; Hopkins, R. Lung function predicts lung cancer. Eur. Respir. J. 2010, 35, 1421–1422. [Google Scholar] [CrossRef]
- Baughman, P.; Marott, J.L.; Lange, P.; Andrew, M.; Hnizdo, E. Health outcomes associated with lung function decline and respiratory symptoms and disease in a community cohort. COPD J. Chronic Obstr. Pulm. Dis. 2011, 8, 103–113. [Google Scholar] [CrossRef]
- Maestú, L.; de Pedro, J.G. Lung function tests in clinical decision-making. Arch. Bronconeumol. 2012, 48, 161–169. [Google Scholar] [CrossRef]
- Ranu, H.; Wilde, M.; Madden, B. Pulmonary function tests. Ulst. Med. J. 2011, 80, 84. [Google Scholar]
- Jaakkola, J.J.K.; Hernberg, S.; Lajunen, T.K.; Sripaijboonkij, P.; Malmberg, L.P.; Jaakkola, M.S. Smoking and lung function among adults with newly onset asthma. BMJ Open Respir. Res. 2019, 6, e000377. [Google Scholar] [CrossRef] [PubMed]
- Dai, X.; Dharmage, S.C.; Lowe, A.J.; Allen, K.J.; Thomas, P.S.; Perret, J.; Waidyatillake, N.; Matheson, M.C.; Svanes, C.; Welsh, L.; et al. Early smoke exposure is associated with asthma and lung function deficits in adolescents. J. Asthma 2017, 54, 662–669. [Google Scholar] [CrossRef]
- Li, L.; Holloway, J.W.; Ewart, S.; Arshad, S.H.; Relton, C.L.; Karmaus, W.; Zhang, H. Newborn DNA methylation and asthma acquisition across adolescence and early adulthood. Clin. Exp. Allergy 2022, 52, 658–669. [Google Scholar] [CrossRef]
- Rathod, A.; Zhang, H.; Arshad, S.; Ewart, S.; Karmaus, W.; Holloway, J. DNA Methylation at Birth Is Associated with Asthma Acquisition from Pre-to Post-Adolescence Mediated by Atopy. In A32. Asthma Clinical Studies; American Thoracic Society: New York, NY, USA, 2020; p. A1334. [Google Scholar]
- Turner, S. Lung Function Tracking: Does It Wobble during Adolescence? American Thoracic Society: New York, NY, USA, 2018; pp. 1470–1471. [Google Scholar]
- Mukherjee, N.; Arathimos, R.; Chen, S.; Rahimabad, P.K.; Han, L.; Zhang, H.; Holloway, J.W.; Relton, C.; Henderson, A.J.; Arshad, S.H.; et al. DNA methylation at birth is associated with lung function development until age 26 years. Eur. Respir. J. 2021, 57, 2003505. [Google Scholar] [CrossRef]
- Sunny, S.K.; Zhang, H.; Relton, C.L.; Ring, S.; Kadalayil, L.; Mzayek, F.; Ewart, S.; Holloway, J.W.; Arshad, S.H. Sex-specific longitudinal association of DNA methylation with lung function. ERJ Open Res. 2021, 7, 00127-2021. [Google Scholar] [CrossRef]
- de Vries, M.; Plaat, D.V.D.; Nedeljković, I.; Amin, N.; Boezen, M. DNA methylation is associated with lung function in never smokers. Respir. Res. 2019, 20, 1–11. [Google Scholar] [CrossRef]
- Mu, G.; Nie, X.; Yang, S.; Ye, Z.; Cheng, M.; Fan, L.; Qiu, W.; Tan, Q.; Zhou, M.; Guo, Y.; et al. PM2.5-related DNA methylation and the association with lung function in non-smokers. Environ. Pollut. 2023, 316, 120700. [Google Scholar] [PubMed]
- Wei, P.; Li, Y.; Wu, L.; Wu, J.; Wu, W.; Chen, S.; Qin, S.; Feng, J. Serum cortisol levels and adrenal gland size in patients with chronic obstructive pulmonary disease. Am. J. Transl. Res. 2021, 13, 8150. [Google Scholar] [PubMed]
- Numa, A.H.; Williams, G.D.; Dakin, C.J. The effect of nebulized epinephrine on respiratory mechanics and gas exchange in bronchiolitis. Am. J. Respir. Crit. Care Med. 2001, 164, 86–91. [Google Scholar] [CrossRef]
- Gólczewski, T.; Lubiński, W.; Chciałowski, A. A mathematical reason for FEV1/FVC dependence on age. Respir. Res. 2012, 13, 1–7. [Google Scholar] [CrossRef]
- Peebles, R.S.; Aronica, M.A. Proinflammatory pathways in the pathogenesis of asthma. Clin. Chest Med. 2019, 40, 29–50. [Google Scholar] [CrossRef]
- Fuentes, N.; Silveyra, P. Endocrine regulation of lung disease and inflammation. Exp. Biol. Med. 2018, 243, 1313–1322. [Google Scholar] [CrossRef]
- Liu, G.; Summer, R. Cellular metabolism in lung health and disease. Annu. Rev. Physiol. 2019, 81, 403–428. [Google Scholar] [CrossRef] [PubMed]
- Neve, V.; Girard, F.; Flahault, A.; Boule, M. Lung and thorax development during adolescence: Relationship with pubertal status. Eur. Respir. J. 2002, 20, 1292–1298. [Google Scholar] [CrossRef]
- Arshad, S.H.; Patil, V.; Mitchell, F.; Potter, S.; Zhang, H.; Ewart, S.; Mansfield, L.; Venter, C.; Holloway, J.W.; Karmaus, W.J. Cohort profile update: The Isle of Wight whole population birth cohort (IOWBC). Int. J. Epidemiol. 2020, 49, 1083–1084. [Google Scholar] [CrossRef] [PubMed]
- Mahmoud, O.; Granell, R.; Tilling, K.; Minelli, C.; Garcia-Aymerich, J.; Holloway, J.W.; Custovic, A.; Jarvis, D.; Sterne, J.; Henderson, J. Association of height growth in puberty with lung function. A longitudinal study. Am. J. Respir. Crit. Care Med. 2018, 198, 1539–1548. [Google Scholar] [CrossRef]
- Hepper, N.G.G.; Fowler, W.S.; Helmholz, F. Relationship of height to lung volume in healthy men. Dis. Chest 1960, 37, 314–320. [Google Scholar] [CrossRef]
- Beyan, H.; Down, T.A.; Ramagopalan, S.V.; Uvebrant, K.; Nilsson, A.; Holland, M.L.; Gemma, C.; Giovannoni, G.; Boehm, B.O.; Ebers, G.C.; et al. Guthrie card methylomics identifies temporally stable epialleles that are present at birth in humans. Genome Res. 2012, 22, 2138–2145. [Google Scholar] [CrossRef] [PubMed]
- Johnson, W.E.; Li, C.; Rabinovic, A. Adjusting batch effects in microarray expression data using empirical Bayes methods. Biostatistics 2007, 8, 118–127. [Google Scholar] [CrossRef]
- Aryee, M.J.; Jaffe, A.E.; Corrada-Bravo, H.; Ladd-Acosta, C.; Feinberg, A.P.; Hansen, K.D.; Irizarry, R.A. Minfi: A flexible and comprehensive Bioconductor package for the analysis of Infinium DNA methylation microarrays. Bioinformatics 2014, 30, 1363–1369. [Google Scholar] [CrossRef]
- Ray, M.A.; Tong, X.; Lockett, G.A.; Zhang, H.; Karmaus, W.J.J. An efficient approach to screening epigenome-wide data. BioMed Res. Int. 2016, 2016, 2615348. [Google Scholar] [CrossRef]
- Karolchik, D. The UCSC genome browser database. Nucleic Acids Res. 2003, 31, 51–54. [Google Scholar] [CrossRef] [PubMed]
- Rosseel, Y. lavaan: An R package for structural equation modeling. J. Stat. Softw. 2012, 48, 1–36. [Google Scholar] [CrossRef]
- Phipson, B.; Maksimovic, J.; Oshlack, A. missMethyl: An R package for analyzing data from Illumina’s HumanMethylation450 platform. Bioinformatics 2015, 32, 286–288. [Google Scholar] [CrossRef] [PubMed]
- Boyd, A.; Golding, J.; Macleod, J.; Smith, G.D. Cohort profile: The ‘children of the 90s’—The index offspring of the Avon Longitudinal Study of Parents and Children. Int. J. Epidemiol. 2013, 42, 111–127. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
| IOWBC (n = 405) | Sample (n = 249) | p-Value | |
|---|---|---|---|
| Variables | Count (%) | Count (%) | |
| Asthma acquisition | 30 (7.41%) | 19 (7.63%) | 0.8943 |
| Mean (SD) | Mean (SD) | ||
| FEV1 (L) | 2.01(0.29) | 2.01(0.31) | 0.8340 |
| FVC (L) | 2.253 (0.33) | 2.25(0.34) | 0.9572 |
| FEV1/FVC Ratio | 0.89 (0.056) | 0.90(0.054) | 0.1668 |
| Height (cm) | 139.33 (6.51 | 139.39(6.55) | 0.8947 |
| FVC | FEV1 | Ratio | Asthma Acquisition |
|---|---|---|---|
| positive regulation of peptide hormone secretion | mannosyltransferase complex | ketone biosynthetic process | postsynaptic density membrane |
| positive regulation of peptide secretion | dorsal/ventral axon guidance | corticotropin secretion | flavonoid glucuronidation |
| glycosylphosphatidylinositol-N-acetylglucosaminyltransferase (GPI-GnT) complex | MHC class II protein complex binding | basolateral plasma membrane | trans-synaptic signaling by lipid modulating synaptic transferase |
| positive regulation of insulin secretion | MHC protein complex binding | C21-steroid hormone biosynthetic process | trans-synaptic signaling by endocannabinoid |
| positive regulation of cytosolic calcium ion concentration involved in phospholipase C-activating G protein-coupled signaling pathway | intrinsic component of endosome membrane | Ctf18 RFC-like complex | xenobiotic glucuronidation |
| structural constituent of muscle | regulation of synaptic transmission, cholinergic | olefinic compound biosynthetic process | postsynaptic specialization membrane |
| positive regulation of hormone secretion | polymeric immunoglobulin receptor activity | response to osmotic stress | catalytic activity, acting on a protein |
| signaling receptor regulator activity | regulation of elastin biosynthetic process | cell–cell contact zone | ionotropic glutamate receptor binding |
| phospholipase C-activating G protein-coupled receptor signaling pathway | negative regulation of elastin biosynthetic process | basal plasma membrane | ubiquitin-like protein transferase activity |
| positive regulation of secretion by cell | negative regulation of synapse assembly | DNA clamp loader activity | integral component of postsynaptic density membrane |
| IOW (CpG Sites from Ttscreening) | No. of Common CpG Sites among IOW and ALSPAC | No. of Coefficients with Same Direction (All Common CpG Sites) | No of Significant CpG Sites from ALSPAC (Raw p-Value) | No of Coefficients with Same Direction (All Significant CpGs from ALSPAC) | |
|---|---|---|---|---|---|
| FEV1 | 514 | 282 | 132 (46.81%) | 21 | 7 (33.33%) |
| FVC | 436 | 222 | 108 (48.65%) | 24 | 12 (50%) |
| Ratio | 408 | 226 | 134 (59.29%) | 11 | 8 (72.73%) |
| Asthma acquisition | 292 | 174 | 76 (43.68%) | 8 | 0 (0%) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abrar, M.N.F.; Jiang, Y.; Zhang, H.; Li, L.; Arshad, H. Epigenetic Features in Newborns Associated with Preadolescence Lung Function and Asthma Acquisition during Adolescence. Epigenomes 2024, 8, 12. https://doi.org/10.3390/epigenomes8020012
Abrar MNF, Jiang Y, Zhang H, Li L, Arshad H. Epigenetic Features in Newborns Associated with Preadolescence Lung Function and Asthma Acquisition during Adolescence. Epigenomes. 2024; 8(2):12. https://doi.org/10.3390/epigenomes8020012
Chicago/Turabian StyleAbrar, Mohammad Nahian Ferdous, Yu Jiang, Hongmei Zhang, Liang Li, and Hasan Arshad. 2024. "Epigenetic Features in Newborns Associated with Preadolescence Lung Function and Asthma Acquisition during Adolescence" Epigenomes 8, no. 2: 12. https://doi.org/10.3390/epigenomes8020012
APA StyleAbrar, M. N. F., Jiang, Y., Zhang, H., Li, L., & Arshad, H. (2024). Epigenetic Features in Newborns Associated with Preadolescence Lung Function and Asthma Acquisition during Adolescence. Epigenomes, 8(2), 12. https://doi.org/10.3390/epigenomes8020012