1. Introduction
Research into the use of microorganisms to perform limited self-maintenance and repair in construction materials, known as engineered living materials (ELMs), has received substantial interest in recent years [
1,
2,
3]. Bacteria with specific phenotypes have the potential to repair damage and prolong the operational life of such materials [
1,
3,
4,
5]. To date, research related to ELMs in construction repair has been confined almost exclusively to concrete, where two mechanisms of microbially induced calcite precipitation (MICP) have been used to repair cracks and maintain the structural integrity of concrete [
6,
7,
8]. The first approach uses ureolytic bacteria to catalyze the hydrolysis of urea to carbon dioxide and ammonia, increasing the pH and the subsequent saturation index for calcium carbonate (CaCO
3) [
1,
7,
9]. In the second approach, an organic calcium salt (such as calcium lactate) [
10], or inorganic calcium salt (such as calcium chloride) [
3], are embedded along with the bacteria in the concrete to provide nutrients for growth and metabolic CaCO
3 precipitation, although the high amount of organic carbon that must be embedded along with the bacteria (up to 50% weight) dramatically reduces the tensile strength of the concrete [
11]. Extensive work examining the potential for microbial self-healing in concrete and other solid building materials such as polymers has been conducted; however, comparatively little work has examined the potential for microbially mediated repair in surface coatings [
12,
13].
It is estimated that ~3.3 × 10
10 L of surface coatings are manufactured worldwide [
14]. Coating is the general term for film-forming materials that provide environmental protection, and encompasses high-performing products for concrete, metal, and other non-architectural surfaces. Paint refers to latex-based coatings suitable for the interiors and exteriors of homes and commercial buildings [
14] and are applied to decorative surfaces with the use of color and texture. In construction, coatings are primarily used for protection, shielding metal and wood surfaces from environmental factors, such as UV radiation, moisture (precipitation), and physical wear and tear [
14]. These coatings generally contain three major components that vary depending on the application: a polymeric binder (typically acrylic or styrene–acrylic latex) that serves as glue to hold together the pigments, such as CaCO
3, titanium dioxide, or silicate clays, which provide color and enhance durability, and the solvent (typically water for latex-based paints) in which the components are dispersed [
14]. Additional functional additives include a thickener to provide viscosity appropriate for the application (brushes, rolling, or spraying), surfactants to limit foaming, and biocides to prevent microbial degradation of the emulsion and maintain shelf-life [
14]. This mix is used as a protective coating over the surface, although when damaged (through scratching, gouging, abrasion, cracking, or age) this protection is compromised, and a new coating must be applied. If a self-healing ELM coating could be developed, where minor damage to the film could be repaired without adding the additional layers needed, this could provide considerable cost and maintenance savings, while reducing structural monitoring.
Self-healing mechanisms have been studied for many years as a way to extend the life of coatings and reduce the maintenance costs associated with re-application [
15]. Current self-healing technologies for paints are based on two approaches: the first involves the incorporation of capsules in the paint that, when broken, fill the damage [
16,
17,
18], and the second involves a more complicated vascular structure that, when broken, allows the flow of material to fill cracks [
19,
20]. Both approaches are based on the mass movement of material from one place to another, weakening the coating at locations other than the site of original damage [
19].
High levels of CaCO
3 (up to 30% solids in many coatings) as a pigment provide the potential for MICP to be used to repair damage in a microbially mediated self-healing paint; however, the use of existing approaches, such as ureolysis, is limited due to the already high pH in formulated coatings, along with the malodor issues that would be associated with the production of ammonia in an interior paint. Bacteria can facilitate precipitation of CaCO
3 through a variety of mechanisms other than ureolysis [
21,
22]. Our past work on bacteria isolated from caves has demonstrated the ability of bacteria to excrete potentially toxic levels of Ca
2+ using the ChaA transporter protein [
23]. To overcome the thermodynamic barrier against pumping out Ca
2+, the cell modifies the extracellular environment through buffering with HCO
3−, altering the pH and leading to spontaneous precipitation of CaCO
3 [
23,
24]. If microbial CaCO
3 precipitation could be incorporated into a paint or coating, it may be possible for the precipitated material to heal cracks in the coating. In this paper, we examine the ability of bacteria with strong precipitation phenotypes, isolated from Ca
2+-rich cave environments, to survive in the water-based acrylate and styrene emulsions commonly used in household paints [
14]. Of the species screened, we examined the ability of a long-surviving
Bacilli species embedded within a model paint film to grow and repair damaged paint films. By exploring some of the phenotypes that would promote survival in the model bacterial species,
E. coil, our results allow for a preliminary understanding of how survival in paint formulations could be affected by cellular architecture and metabolism.
2. Materials and Methods
Cultures and growth conditions. All environmental strains used in this study were originally collected and identified as described in Banks et al. [
23], with all incubations carried out at room temperature (21 °C). The
E. coli knockout strains used in this study were obtained from the Keio collection, a collection of
E. coli strains where the target gene is replaced by a kanamycin resistance cassette [
25]. Liquid cultures were shaken at 85 rpm on a Jeio Tech Lab Companion SKC 6200 orbital shaker (Billerica, MA, USA). Unless otherwise stated, growth was carried out on half-strength TSA, containing 15 g/L trypticase soy broth and 15 g/L agar, or half-strength TSB containing 15 g/L trypticase soy broth (BD Difco, Franklin Lakes, NJ, USA). The B4 media contained 4 g/L yeast extract, 10 g/L glucose, and 15 g/L agar, pH 7.2, with 2.5 g/L calcium acetate added after autoclaving to prevent precipitation [
26,
27]. The minimal B4 media was made the same way but lacked the added calcium acetate. All media were sterilized in a SR24D SV autoclave at 121 °C for 15 min (Consolidated Sterilizer Systems, Boston, MA, USA).
Spore suspensions of strain B. simplex GGC-P6A were prepared by growth in TSB for 9 days. The mixed cell/spore suspension was then pelleted at 2700× g for 10 min in a Sorvall XTR centrifuge (Thermo Fisher Scientific, Whaltam, MA, USA) and resuspended in 25 mL sterile H2O with shaking for an additional 7 days. Microscopic cell counting using an Olympus BX53 light microscope (Shinjuku, Japan), following malachite green (0.5%) staining with a safranin (2.5%) counterstain, confirmed that the final suspension contained > 99.8% spores. Vegetative cells and spore suspension CFUs were confirmed by serial dilution and plating on TSA.
Binder chemistry and model coating composition. Unless stated, all reagents were purchased from Sigma Aldrich and used as received. Stock solutions of Acrysol TT-935 were purchased from Dow Chemical (Midland, MI, USA). ALBAFIL S10 (70% CaCO
3, 700 nm particle size) was obtained from Specialty Minerals (Adams, MA, USA). Upon arrival, the ALBAFIL was autoclaved at 15 psi for 15 min prior to use. The general scheme of binder synthesis is shown in
Supplemental Figure S3.
MBF1 was prepared using a semi-continuous emulsion polymerization with an ammonium persulfate initiator and a monomer mixture including 41.0% styrene, 10% methyl methacrylate, 43.5% 2-ethyl hexyl acrylate, 3.0% methacrylic acid, and 2.5% diacetone acrylamide. The monomers and initiator were fed into a reactor containing an additional initiator at 80 °C over a period of 3 h and stirred for an additional 30 min. Then, 20% terbutylhydroperoxide and 20% sodium metabisulfate were added as separate solutions over 45 min at 65 °C to ensure monomer consumption. The liquid was cooled to room temperature and adjusted to a pH of 8.5 with ammonium hydroxide. A mixture of 20% adipic dihyrazide was added and then the solution was filtered.
MBF2 was prepared using a semi-continuous emulsion polymerization with an ammonium persulfate initiator and a monomer mixture including 58% methyl methacrylate, 37% butyl acrylate, 2.5% methacrylic acid, and 2.5% diacetone acrylamide. The monomers and initiator were fed into a reactor at 80 °C containing an additional initiator over a period of 2.5 h and stirred for an additional 30 min. Then, 20% terbutylhydroperoxide and 20% sodium metabisulfate were added as separate solutions over 45 min at 65 °C to ensure monomer consumption. The liquid was cooled to room temperature and adjusted to a pH of 8.5 with ammonium hydroxide. A mixture of 20% adipic dihyrazide was added and then the solution was filtered.
MBF3 was prepared using a semi-continuous emulsion polymerization with an ammonium persulfate initiator and a monomer mixture including 37.0% styrene, 9.5% methyl methacrylate, 45.5% butyl acrylate, and 8.0% methacrylic acid. The monomers and initiator were fed into a reactor containing an additional initiator at 78 °C over a period of 3 h and stirred for an additional 30 min. Then, 20% terbutylhydroperoxide and 20% sodium metabisulfate were added as separate solutions over 45 min at 65 °C to ensure monomer consumption. The liquid was cooled to room temperature and adjusted to a pH of 8.5 with ammonium hydroxide. A mixture of 20% zinc oxide was added and then the solution was filtered.
Bacterial survival assays. To examine survival in raw materials, 100 μL of a 3 mL overnight culture in TSB was used to inoculate 3 mL of MBF1, MBF2, and MBF3 before shaking at 85 rpm for 24 h, whereupon 100 μL samples were spread on TSA. Survival was recorded as positive/negative growth after 5 days. Long-term survival of GGC-P6A in binders was carried out in triplicate, although initial cell numbers were recorded by serial dilution of the inoculated binder on day 0, with cell survival recorded at 2, 7, 14, 21, and 28 days. The percent survival was calculated by averaging cell survival numbers at each time point, compared to the calculated CFU/mL at day 0 (recorded as % survival).
The model paint was prepared using 20 mL of the MBF1 binder, 1.6 mL ALBAFIL S10, 0.2 mL TT-935, and 18.2 mL sterile H2O and inoculated on day 0 with either GGC-P6A vegetative cells or spores. CFU values were measured. CFU values were measured on day 2, 7, 14, 21, and 28. The paint films were prepared using the model paint, inoculated as described, with 10 mL volumes pulled down immediately onto non-stick polypropylene mats using a Leneta WC-1670 Wire-Cator (Mahwah, NJ, USA). The film was allowed to dry overnight in a laminar flow hood and cut into 25 mm-diameter coupons using a sterile hole punch (Fiskars, Helsinki, Finland). Coupons were then stored in sterile Petri dishes before plating on B4 media for growth. CFUs were calculated as the total cell number in the inoculated volume of the disk (22 μL) and emergent colonies counted by backlighting of the film. Survival was calculated as percentage survival compared to the initial inoculum on day 0.
Microscopy. Carbonate precipitation was monitored using an Olympus BX60 microscope using an DP73 camera attachment and cellSens v1.11 image capture (Olympus, Shinjuku, Japan). For scanning electron microscopy (SEM) analysis, 1 cm2 squares were excised from dry paint films and coated with iridium using a Cressington 208HR Sputter Coater (Watford, UK). The thickness of the coating was estimated at 3 nm using the internal thickness monitor included with the coater. Images were captured using a Hitachi S-4700 Field Emission SEM (Chiyoda, Japan) in the SE (secondary electron) mode at 5 kV.
Zeta Potential Assays. Strains were grown in 3 mL of LB (which supports the growth of E. coli) overnight. Cells were pelleted by spinning the cultures at 3500 rpm for five minutes at 4 °C. Cell pellets were washed three times with 1 mM potassium phosphate buffer (PB) before being resuspended in 3 mL of PB buffer. The samples were diluted 1:10 before 1 mL was loaded into a Zetasizer Nano (Malvern Panalytical) for measurement.
3. Results
To determine the ability of bacteria to survive in paint, we first examined cell viability in paint binders. As commercially available emulsion polymers cannot be purchased without preservatives (biocide additives), three polymeric paint binders were synthesized (see materials and methods). The three emulsion polymers were chosen to include the major types of monomers used in paint binders [styrene–acrylic (MBF1 and MBF3) and acrylic (MBF2)] and cross-linking chemistries (
Table 1) [
14].
Fifteen bacterial strains previously isolated from caves that display a strong calcifying phenotype were selected for analysis (summarized in
Table 2), including Gram-negative/positive and spore-forming species [
23]. Given the large number of strains examined in three separate latex binders, we used a rapid, qualitative screen for survival. Vegetative cells from an overnight culture were assayed for survival for 24 h at room temperature (
Table 2). All the tested strains survived in binder MBF1, ten survived in MBF2, and nine species survived in MBF3 (
Table 2). It was interesting to note that survival in MBF1, a styrene–acrylic polymer, did not necessarily correlate with survival in the other styrene–acrylic polymer MBF3 (
Table 2). There was also no consistency in species survivability; for example, only two of three
Brevundimonas strains (GGC-D2A and GGC-D2B) survived in MBF2, while none survived in MBF3. Finally, the ability of the tested species to form spores did not demonstrate any increase in survival (
Table 2).
We decided to quantify survival of
Bacillus simplex str. GGC-P6A, which demonstrated the best survival after 24 h in both MBF1 and MBF2 (MBF3 was not used due to the loss of GGC-P6A viability at <24 h;
Table 2). Commercial paints are often a complex mixture, which can consist of up to 60 different additives, including biocides (for example, see ref. [
28]). We therefore developed a simplified formula to assay survival of GGC-P6A containing only latex binder, CaCO
3 pigment, thickener, and water. We exclusively used binder MBF1, which provided the best recovery rates for GGC-P6A. The CaCO
3 slurry was ALBAFIL S10, a commercial slurry that negated the need for additional dispersants or surfactants [
14]. The commercial thickener, Acrysol TT-935, is only available containing the biocides methylchloroisothiazolinone (MCI) and methylisothiazolinone (MI) at ~0.3% wt/wt [
29]. To limit the impact of the biocides, TT-935 was diluted with water to a working concentration of 1% vol/vol, giving a final concentration of 0.0015% MCI/MI. As adequate films could be created with this simple formulation, no coalescent was required [
14]. The model paint was inoculated with GGC-P6A vegetative cells and viability was examined for 15 days (
Figure 1).
Given that GGC-P6A is a spore former, we wondered whether this extended survival was due to sporulation and used purified spores in our assays to measure long-term survival. Unsurprisingly, spore survival was higher compared to vegetative cells (
Figure 1). Spores do not require nutrients, since they are in a dormant state, and there was no decrease in spore number over the 28 days compared to the vegetative cells (
Figure 1). We observed an increase in overall cell numbers above the initial inoculum (
Figure 1); however, there was more variability in spore number than vegetative cells. We believe that these variations may be due to clumping of spores within the paint, although this would be challenging to examine. Nonetheless, these data indicate that GGC-P6A spores were clearly able to remain viable at high levels for at least four weeks post-inoculation (
Figure 1) and are the ideal method of inoculation to retain large amounts of viable cells in our artificial paint formulation.
In order to be functional within an engineered coating, bacteria must be able to survive the process of drying and formation of a surface film. To determine the survival of GGC-P6A through this process while maintaining functionality (CaCO
3 precipitation), the liquid coating composition was inoculated with either GGC-P6A vegetative cells or spores. The coating was then immediately poured to create a film and allowed to dry for 24 h in sterile conditions to create coupons that could be cracked and placed on B4 media to facilitate growth, CaCO
3 precipitation, and examine potential crack filling. In the paint coupons inoculated with vegetative cells, bacterial growth was seen emerging across the surface and along the edges of the paint film within 3 days (
Figure 2). After 7 days, the paint coupons demonstrated CaCO
3 precipitation, both on the surface of the film and along the edges of the cracks, with a noticeable carbonate front advancing ahead of bacterial growth (
Figure 2). In places, this precipitated carbonate was able to occlude introduced cracks in the film (
Figure 2B). When paint coupons inoculated with GGC-P6A spores were placed on B4 media, a similar pattern of growth and CaCO
3 precipitation was observed, suggesting that sporulation did not interfere with the precipitation phenotype.
The CaCO
3 precipitation assays on the paint films were carried out on B4 media, which contains Ca(C
2H
2OOH)
2 as a calcium source; however, our original hypothesis was that the CaCO
3 within the coating would be sufficient to serve as a source of Ca
2+ for CaCO
3 precipitation. To determine if this was the case, we created a B4 media that lacked Ca(C
2H
2OOH)
2 (
Figure 2D). When the paint coupons containing GGC-P6A vegetative cells were grown on this Ca
2+-free media, CaCO
3 precipitation could also be observed at the growth front, albeit at a reduced rate than observed on regular B4 media (
Figure 2D). SEM images demonstrated vegetative cells emerging from, and following, cracks in the paint, or tracking along the edge of the paint film and onto the support media (
Supplemental Figure S1) during the crack-filling process.
Once it was clear that the bacteria were able to survive the film-forming process and calcify, we wanted to examine their ability to survive long term in dried paint films. To do this, we inoculated the paint films with both vegetative cells and spores at a comparable level to our work in the liquid formulation: ~1 × 10
6 CFU/mL. The films were then cut in 25 mm circles to standardize the volume of coupon used to 22 μm
3 (the coupons were measured to be ~45 μm in thickness;
Supplemental Figure S2). These coupons were then either examined immediately or stored in empty, sterile Petri plates to assess long-term survival. When the coupons were incubated on B4 support media, there was significant microbial growth, and the film became confluent (
Figure 3A). This made it difficult to quantify the actual number of colonies emerging from the film (
Figure 3A), so we repeated the experiment by reducing the inoculum to 3.1 × 10
3 CFU/mL. While containing significantly fewer cells than the liquid formulations, this did allow us to observe quantifiable colony growth (
Figure 3A). The coupons were plated on B4 support media at one-week intervals and the emergent colonies counted (
Figure 3B). At day 0, the CFU/mL values obtained were comparable to the total number spores added (4.9 × 10
3 CFU/mL ± 0.4 × 10
3), suggesting that the creation of paint films did not have an immediate effect on viability. These data also validated this assay as a way of observing bacterial survival within the paint film. Over the course of the experiment, we saw a 50% reduction in the recovery of GGC-P6A spores in paint films by day 7, a number that appeared to remain stable over 28 days of the experiment, with no significant additional reduction (
Figure 3B).
It was unsurprising that spore formers demonstrated long-term survival in the paint formulations; however, the use of spore formers somewhat limits the bacterial species that can be incorporated into coatings, or that are tractable for genetic enhancement. We had limited Gram-negative species in our cave isolates so decided to use
Escherichia coli as a model species. We inoculated samples of model paint containing MBF1 with
E. coli from a number of genetic backgrounds, including K-12, BL21 (DE3), and BW25113. To our surprise, not only did
E. coli strains K-12 and BW25113 demonstrate low but persistent survival (
Figure 4), but there was an inconsistency in the survival between strains, with
E. coli strain BL21 (DE3) becoming unculturable after 3 d. Periodic sampling after 15 d demonstrated that cells remained culturable for at least 32 days for
E. coli K-12 and BW25113, equivalent to the survival seen in GGC-P6A.
The
E. coli strain BW25113 is a K-12 derivative, which lacks the ability to synthesize the complete
E. coli O-antigen of LPS due to an insertion element present in
wbbL [
30,
31]. BL21 (DE3) is a B strain, which also has a disruption in the O-antigen synthesis pathway, albeit at a different locus [
30]. Genetic differences between the two backgrounds contribute to an outer membrane surface with significant structural differences [
32], which could explain the difference in survival and affect cell interactions with the charged components of the polymer emulsions. While it was not possible to test any varying chemistry within the model paint formulation, we could compare the cellular zeta potential between strains as a general measure of surface chemistry in each
E. coli strain (
Figure 4). The largest difference in zeta potential was observed between BW25113 and K-12 (−28.7 and −64.4 mV, respectively), although survival between these strains was not significantly different after 6 d of growth (Student’s
t-test: df = 2,
p = 0.37,
Figure 2). Overall, no clear difference in viability was observed between the relative cell surface charge (
Figure 4).
Given that LPS is known to stabilize the Gram-negative outer membrane against environmental stresses, the lack of survival in BL21 (DE3) may be attributed to a truncated O-antigen. To determine if LPS altered the survival phenotype, we assayed survival in
E. coli MAJ033, where a functional
wbbL has been cloned into the K-12 background [
33], and it demonstrated an overall change in surface charge (
Figure 4). When this strain was assayed for survival in MBF1, the restoration of LPS synthesis demonstrated significantly higher culturability after inoculation compared to the other
E. coli strains (
Figure 4); however, survival was not significantly different from other strains after 9 days of incubation. This loss of viability after 9 days may be due to the lack of nutrients in the MBF1 paint formulation. Since the purpose of these experiments was to model commercial paint, we did not investigate whether the addition of nutrients promoted extended survival in this strain.
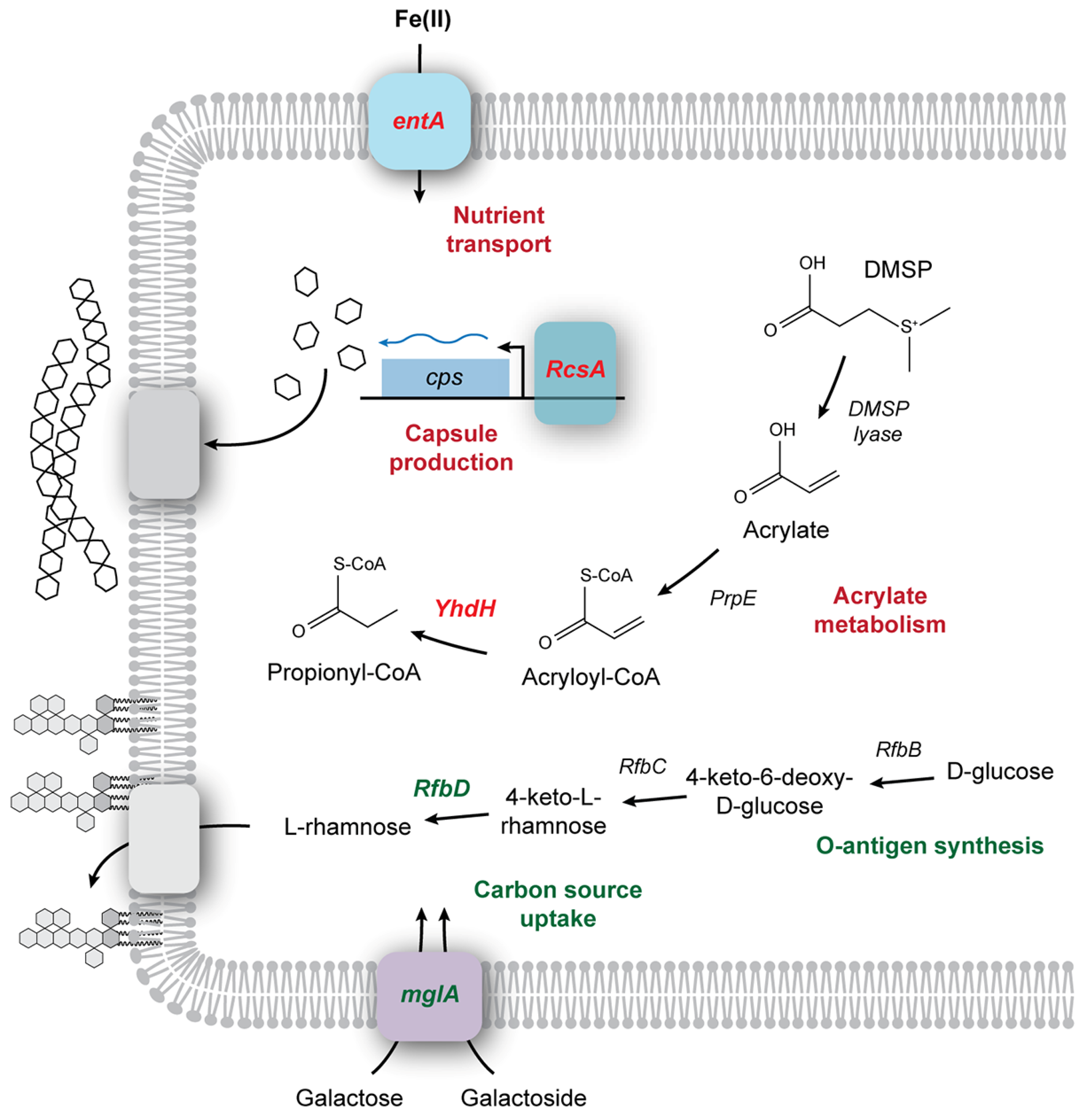
Since a robust genetic system exists for
E. coli, using this strain gives us an opportunity to probe some of the other phenotypic determinants may contribute to the survival of bacteria in the paint environment using the Keio library (
Figure 5) [
25]. As screening for survival in the presence of an opaque model paint makes optical density measurements impossible, rather than screening the entirety of the Keio mutant strain library, we screened genes of interest using dilution plating. These genes included Δ
rcsA, as the B strain (BL21) lacks the ability to produce capsules due to the loss of the RcsA regulatory protein [
34], leading to a complete loss of BL21 viability at 3 d in our initial survival assay (
Figure 4). We selected Δ
yhdH due to its role in acryloyl-CoA metabolism; the MBF1 solution is a styrene–acrylic latex, which could potentially be toxic due to the production of acryloyl-CoA during acrylate metabolism. Acryloyl-CoA is an electrophile that can interfere with electron transport [
35], and mutants lacking the enzyme acryloyl-CoA reductase (
yhdH) that converts acryloyl-CoA to the less toxic propionyl-CoA are more sensitive to acrylate [
36]. Finally, we included Δ
entA, which lacks a gene necessary for synthesis of siderophores [
37]. Since our model paint is a nutrient-poor environment, we wondered whether scavenging systems played an important role in the extended survival of
E. coli strains. As controls, we used the WT host strain BW25113, along with two mutants we predicted would have no impact on survival: Δ
mglA, a subunit of the D-galactose/D-galactoside ABC transporter, and Δ
rfbD of the operon responsible for synthesizing the rhamnose component of the O-antigen,
rfbABCDX. As we are not growing in the presence of galactose, and the K-12 background lacks a complete O-antigen, the deletion of either
mglA or
rfbD should have no observable phenotype in this assay. These mutant strains were incubated for 15 d at room temperature and the percentage survival (versus initial inoculum) was recorded (
Figure 6). No significant difference in viability was observed between Δ
mglA or Δ
rfbD and BW25113. The survival of all three mutant strains (Δ
rcsA, Δ
yhdH, and Δ
entA) was significantly lower than WT, with the lowest occurring in Δ
yhdH, which is a mutation that affects acrylate metabolism (
Figure 5). A one-directional ANOVA analysis [F(11,24) = 2.22,
p < 0.001] indicated that the loss in viability of Δ
entA, Δ
rscA, and Δ
yhdH was significant (
Figure 6), indicating that that the poor survival of
E. coli strains may be due to cell surface chemistry, polymer toxicity, and nutrient limitation.
4. Discussion
In our past work, we observed that limestone (CaCO
3) cave environments contain bacterial species that use CaCO
3 precipitation to maintain Ca
2+ homeostasis under what would otherwise be toxic levels [
23]. The impact of this CaCO
3 precipitation in situ is the accumulation of microbially induced, centimeter-scale, secondary deposits known as cave popcorn [
23]. Such secondary deposits can accumulate to many meters in length (stalactites and stalagmites) and occlude the original rock surface [
23]. Coatings, including paints, often include CaCO
3 as a filler, which provides both color and appearance [
14]. We therefore wondered whether added CaCO
3 could provide a source of Ca
2+ ions to drive a similar precipitation process in coatings to generate a limited self-repair mechanism: a living paint. Toward this goal, we carried out proof-of-concept experiments to determine whether such bacteria could be successfully added to paints and coatings, remain viable through the formulation and film-forming process, and maintain a functional CaCO
3 precipitation phenotype to repair induced damage.
Water-based paint formulations contain a variety of organic polymers capable of supporting microbial growth. When sealed into a paint can, microbial anaerobic growth causes the degradation and spoilage of the paint formulation, including the release of unpleasant odors and potentially dangerous microbially derived volatile organic compounds [
38,
39,
40]. The contamination of liquid formulations can also lead to substantial contamination of the final paint film, which can subsequently result in accelerated deterioration and discoloration [
41,
42]. While much work has attempted to identify the species responsible for this growth, a range of species have been observed, and no work has attempted to identify the factors that allow for survival and growth in these chemistries [
43]. Instead, most research (and patents) examining microbial survival in paint formulations has been geared toward limiting microbial (bacterial and fungal) survival and growth, either through the addition of biocides or formulation [
41,
44,
45]. Our approach was to determine whether we could leverage microbial growth to generate a unique, engineered living material approach to improve the functionality of paint, while attempting to understand some of the factors that allow survival in a type bacterial species.
Our first step was to determine whether the bulk chemistry of paint would allow for the survival of our calcifying species. We examined the ability of 15 cave isolates (that demonstrated a strong calcifying phenotype [
23]) to remain viable in three synthesized latex emulsions: MBF1, MBF2, and MBF3 (
Table 1). Our original hypothesis was that the polymer chemistry would be the primary factor affecting cell viability, either through osmotic stress or charge effects on the surface of the polymer emulsions [
46,
47]. We expected a strong pattern of inhibition related to either the polymer chemistry or cellular structure of the target species (for example, binding effects related to Gram-negative or Gram-positive surface chemistries [
46,
47]), but no such pattern was observed. Rather, the data suggest that the impact of binder chemistry on viability may be related to species/strain genotype. Unsurprisingly, spore-forming bacteria maintained the greatest viability, with MBF1 (styrene–acrylic) being the most permissive to survival (
Table 2). MBF3 is also a styrene–acrylic polymer, and this formulation was the least permissive to survival. In the monomeric form, styrene acts as an organic solvent that can reduce bacterial viability in the low mM range [
48]. The mechanisms of styrene toxicity are unclear, although research suggests that styrene can remodel membrane fatty acids from the
cis to the
trans form via a general solvent stress response [
48]. Given that we added cross-linking agents in excess (20%) during polymer synthesis, it is unlikely that unbound monomers reach the μM range (well below the toxicity observed for styrene), suggesting that styrene monomers alone are responsible for the observed loss of viability (
Table 2; [
49,
50]). Another difference between MBF1 and MBF3 was the polymer cross-linking chemistry; MBF1 used diacetone acrylamide while Zn(II) was used for MBF3.
While acrylamide is an acute mutagen in eukaryotic cells, it has a reduced toxicity in bacteria [
51], having been used successfully to encapsulate bacteria for a variety of industrial processes [
52]. On the other hand, Zn(II) is known to be toxic [
53], outcompeting Mg(II) cofactor binding sites and bringing about cell death by limiting essential cellular processes [
54]. There are several ATP-driven transport pumps that detoxify cellular Zn(II); however, these tend to be plasmid-encoded, leading to strain and species variability (
Table 2; [
54,
55]). We have previously demonstrated that cave-adapted microbial species display different sensitivities to transition metals [
56], so the strain-specific sensitivity to MBF3 may reflect a differential sensitivity to Zn(II) rather than polymer chemistry.
Acrylates (CH
2=CHCOO
−) are found in relatively high abundance in nature, where they are formed from the catabolism of dimethylsulfonioproprionate (DMSP), an important organic compound in the sulfur cycle (
Figure 5) [
57]. As a result, the inhibitory levels of acrylates are relatively high (~30 mM; [
58]). The ability to catabolize DMSP is dependent on the presence of the
ddd genes, a group of DMSP lyases that cleave DMSP into 3-hydroxyproprionate (3HP) and acrylate [
58]. The next step in catabolism is the conversion of 3HP and acrylate to acryloyl-CoA, a strong electrophile capable of interfering with electron transport [
36,
57]. As a result, DMSP catabolism requires the cell to rapidly convert acryloyl-CoA to propionyl-CoA via acetyl-CoA reductase [
36]. Genes involved in DMSP catabolism are widespread within bacterial species, even those species unlikely to be exposed to naturally occurring acrylates, including the genera
Bacilli and
Microbacterium (
Table 2; [
57,
58]). Such pathways may therefore catabolize trace acrylate, with or without the cellular detoxification pathways necessary to remove acryloyl-CoA intermediates, as suggested by the Δ
yhdH mutant [
57]. The mutant Δ
yhdH lacks a gene which encodes a protein with structural similarity to acryloyl-CoA reductase,
acuI of
Ruegeria pomeroyi [
36], and survival was dramatically reduced in Δ
yhdH (
Figure 6), suggesting that the ability to metabolize potentially toxic secondary products of acrylamide may be an important phenotype for strain viability in acrylate binders (
Figure 5).
Given their chemically resistant nature, the GGC-P6A spores demonstrated greater long-term survivability than vegetative cells (
Figure 2), and subsequent sampling demonstrated culturability in excess of 200 d. Nonetheless, any microbe added to a paint must be able to survive not only in the paint mixture, but the drying process once applied to a surface. When the liquid formula was inoculated and immediately drawn down into a paint film, both vegetative cells and spores remained viable through the drying process, but only the spore-containing formulations remained viable for 30+ days in films. When the dried films were placed on agar, microbial growth could be seen emerging from the edges of the film and along small cracks, suggesting that damage to the acrylic polymer may be necessary for the release of cells from entombment within the polymer, although direct access to nutrients in the agar matrix did not appear to be essential (
Figure 3). This growth was quantified for GGC-P6A for 28 d (
Figure 3), while the coupons contained viable spores for >1 year. Over time, a combination of microbial growth and CaCO
3 precipitation did lead to the occlusion of small cracks introduced into the paint film (
Figure 3) with sufficient structural integrity that these films remained intact across the zone of repair when lifted from the plate. While the strength of this repair was not quantified, it does provide a level of self-healing that may be sufficient to protect the surface underneath and is a direction for future research. In all instances, microbial CaCO
3 precipitation continued to be observed in films, suggesting that the formulation, film-forming, and drying processes are permissive to maintaining this phenotype.
Very few of our original calcifying strains were Gram-negative bacteria. We thought that the outer lipid membrane would reduce viability in our formulations; however, when we used
E. coli as a model organism, the data demonstrated viability for >15 d in MBF1. Nonetheless, there was variability in survival of the different
E. coli strains. Given that polymer emulsions have a surface charge, we wondered whether the surface chemistry of the tested bacterial species was important for survival. To dissect the properties that could influence this survival, we examined the viability of several
E. coli strains, along with mutants from the Keio library that differ significantly in membrane chemistry (
Figure 4; K strains lack the O-antigen and B strains lack the ability to produce capsules) [
34]. Our data did demonstrate a pattern of viability, and the restoration of LPS O-antigen synthesis (using
wbbL
+) improved survival, while the Δ
rcsA mutant lacks ability to generate a capsule (similar to BL21;
Figure 5) and had no detectable survival in our model paint, suggesting that the chemistry of the outer membrane is important for viability within paint. The gene
entA is necessary for synthesis of the siderophore enterobactin [
37] and Δ
entA had a reduced survival rate in paint. While this could indicate that nutrient limitation may prevent extended survival, enterobactin also has a role in resistance to oxidative stress [
59]. It is difficult to experimentally dissect these two interpretations in our system, especially as the addition of cationic Fe
2+ ions will disrupt the emulsion chemistry. Nonetheless, the addition of nutrients to a formulated paint, which is often sealed for long-term storage, would in the very least be an extremely problematic mechanism of maintaining bacterial survival.
Our work has demonstrated the potential for an engineered living material that incorporates functional bacterial species into coatings and films. These data suggest that the impact of binder composition appeared to be strain-specific and that the polymer chemistry, cross-linking chemistry, and genotype of target species should all be considered when designing a coating that incorporates bacterial species. It is no surprise that spores survive for much longer in binders and liquid coatings, although it is unclear as to whether germination, growth, and subsequent germination occurs within the liquid paint formulation.
While our experiments suggest a self-healing paint that incorporates bacterial activity is possible, there are many obstacles that remain before a functional solution is possible. For example, the growth of microorganisms would need to be limited until self-healing is initiated, preventing unwanted growth in stored paint, and Ca2+ ions within the paint film sufficient to support CaCO3 precipitation and growth without the presence of a growth media/substrate remain elusive. Ongoing research will be required to examine ways to overcome these limitations.
Anthropogenic emissions of greenhouse gases represent one of the greatest threats to human society, with impacts on our climate that are expected to increase extreme weather events, reduce food production, reduce biodiversity, and damage important ecosystem services [
60,
61,
62,
63,
64]. As society continues to shift toward sustainable production, there are many industries, such as construction, that are tied to products with few ‘green’ alternatives. While a self-healing coating does not remove the need for polymer synthesis, it could provide a cost-effective approach to reduce the number of applications and volume of paint needed to protect and maintain coated surfaces. The carbonates produced in our method are also fixed from atmospheric CO
2, which does not require the movement of material from one site within a film to another to mediate repair that has been problematic in other proposed self-repair systems. Such CO
2 removal also provides for the possibility of carbon fixation in our approach, which has not been explored in any other microbially mediated self-healing approach.









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}