
Molecular Investigation of Klebsiella pneumoniae from Clinical Companion Animals in Beijing, China, 2017–2019

 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Pets Samples and K. pneumoniae Isolates
2.2. Antimicrobial Resistance
2.3. Antibiotic Resistance Genes and Virulence Genes
2.4. Hypervirulent, Hypermucoviscosity, Capsule Serotype, and O-Antigen of K. Pneumonia
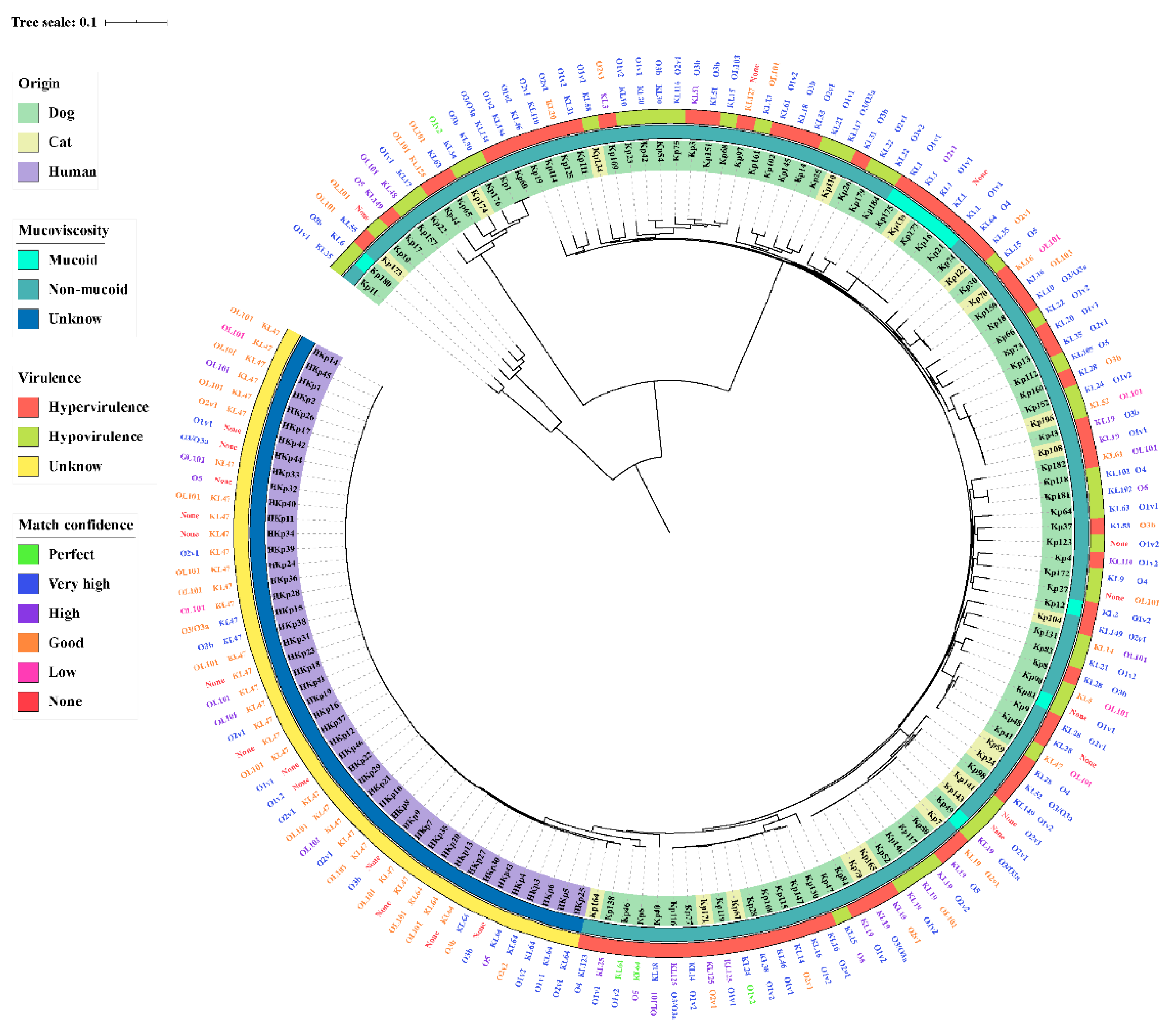
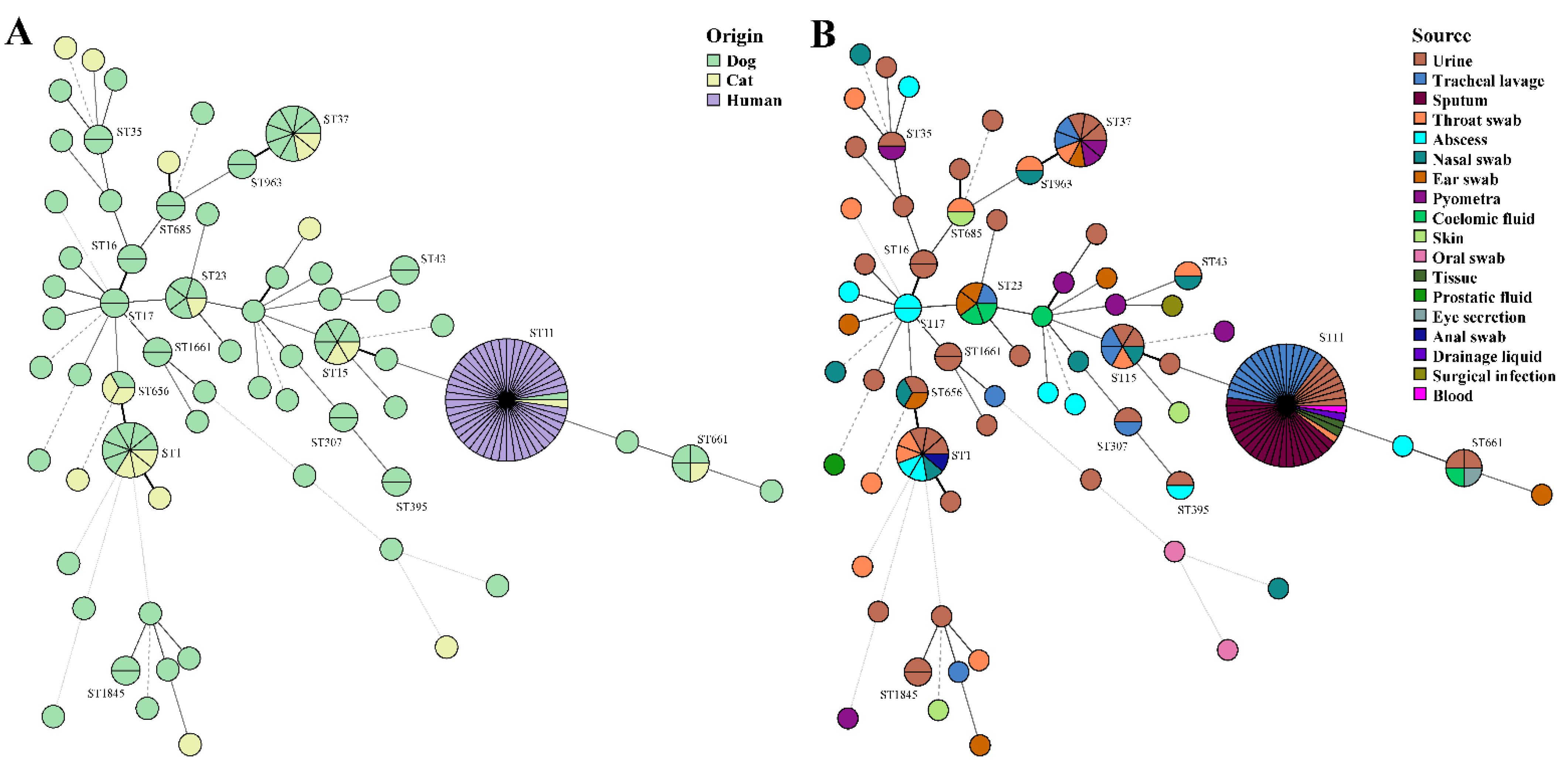
2.5. Diversity Genotypes of the K. pneumoniae Isolates
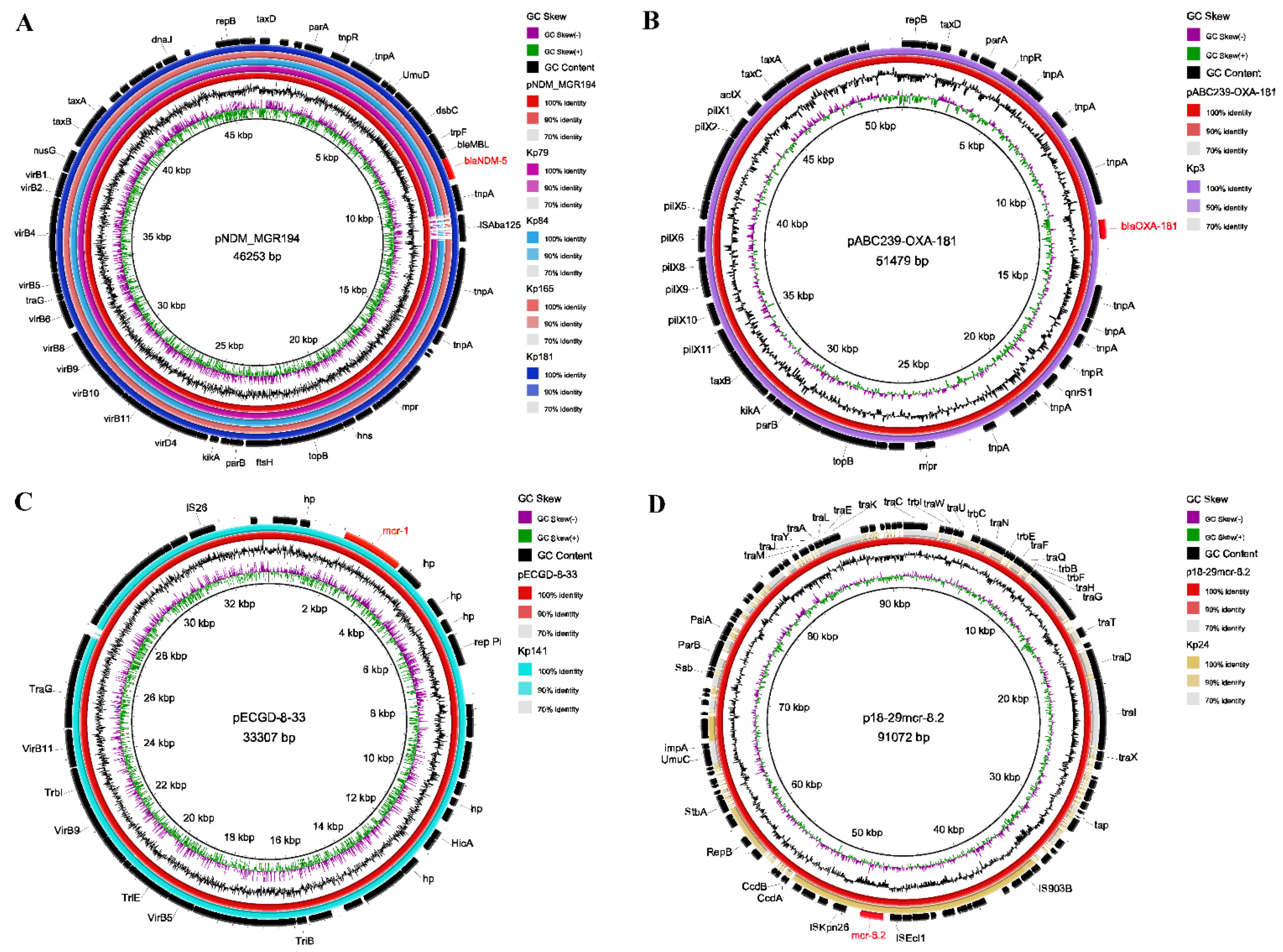
2.6. Characterization of Plasmids
3. Discussion
4. Materials and Methods
4.1. Samples Collection and Bacterial Characterization
4.2. Antimicrobial Susceptibility Testing
4.3. Mucoviscosity Assay
4.4. Galleria Mellonella Virulence Assay
4.5. Whole-Genome Sequencing and Molecular Analysis
4.6. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Holt, K.E.; Wertheim, H.; Zadoks, R.N.; Baker, S.; Whitehouse, C.A.; Dance, D.; Jenney, A.; Connor, T.R.; Hsu, L.Y.; Severin, J.; et al. Genomic analysis of diversity, population structure, virulence, and antimicrobial resistance in Klebsiella pneumoniae, an urgent threat to public health. Proc. Natl. Acad. Sci. USA 2015, 112, E3574–E3581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wyres, K.L.; Holt, K.E. Klebsiella pneumoniae Population Genomics and Antimicrobial-Resistant Clones. Trends Microbiol. 2016, 24, 944–956. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wyres, K.L.; Lam, M.M.C.; Holt, K.E. Population genomics of Klebsiella pneumoniae. Nat. Rev. Microbiol. 2020, 18, 344–359. [Google Scholar] [CrossRef] [PubMed]
- Parisi, S.G.; Bartolini, A.; Santacatterina, E.; Castellani, E.; Ghirardo, R.; Berto, A.; Franchin, E.; Menegotto, N.; De Canale, E.; Tommasini, T.; et al. Prevalence of Klebsiella pneumoniae strains producing carbapenemases and increase of resistance to colistin in an Italian teaching hospital from January 2012 To December 2014. BMC Infect. Dis. 2015, 15, 244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tacconelli, E.; Carrara, E.; Savoldi, A.; Harbarth, S.; Mendelson, M.; Monnet, D.L.; Pulcini, C.; Kahlmeter, G.; Kluytmans, J.; Carmeli, Y.; et al. Discovery, research, and development of new antibiotics: The WHO priority list of antibiotic-resistant bacteria and tuberculosis. Lancet Infect. Dis. 2018, 18, 318–327. [Google Scholar] [CrossRef]
- Wang, X.; Wang, Y.; Zhou, Y.; Li, J.; Yin, W.; Wang, S.; Zhang, S.; Shen, J.; Shen, Z.; Wang, Y. Emergence of a novel mobile colistin resistance gene, mcr-8, in NDM-producing Klebsiella pneumoniae. Emerg. Microbes Infect. 2018, 7, 122. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Zhang, R.; Li, J.; Wu, Z.; Yin, W.; Schwarz, S.; Tyrrell, J.M.; Zheng, Y.; Wang, S.; Shen, Z.; et al. Comprehensive resistome analysis reveals the prevalence of NDM and MCR-1 in Chinese poultry production. Nat. Microbiol. 2017, 2, 16260. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.-Y.; Wang, Y.; Walsh, T.R.; Yi, L.-X.; Zhang, R.; Spencer, J.; Doi, Y.; Tian, G.; Dong, B.; Huang, X.; et al. Emergence of plasmid-mediated colistin resistance mechanism MCR-1 in animals and human beings in China: A microbiological and molecular biological study. Lancet Infect. Dis. 2016, 16, 161–168. [Google Scholar] [CrossRef]
- Liu, L.; Feng, Y.; Zhang, X.; McNally, A.; Zong, Z. New Variant of mcr-3 in an Extensively Drug-Resistant Escherichia coli Clinical Isolate Carrying mcr-1 and blaNDM-5. Antimicrob. Agents Chemother. 2017, 61, e01757-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Yang, L.; Liu, Z.; Yin, W.; Liu, D.; Shen, Y.; Walsh, T.; Shao, B.; Wang, Y. Molecular Insights into Functional Differences between mcr-3- and mcr-1-Mediated Colistin Resistance. Antimicrob. Agents Chemother. 2018, 62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, Y.; Wang, S.; Zhan, L.; Jin, Y.; Duan, J.; Hao, Z.; Lv, J.; Qi, X.; Chen, L.; Kreiswirth, B.N.; et al. Microbiological and Clinical Characteristics of Hypermucoviscous Klebsiella pneumoniae Isolates Associated with Invasive Infections in China. Front. Cell. Infect. Microbiol. 2017, 7, 24. [Google Scholar] [CrossRef]
- Zhang, Y.; Jin, L.; Ouyang, P.; Wang, Q.; Wang, R.; Wang, J.; Gao, H.; Wang, X.; Wang, H.; Kang, H.; et al. Evolution of hypervirulence in carbapenem-resistant Klebsiella pneumoniae in China: A multicentre, molecular epidemiological analysis. J. Antimicrob. Chemother. 2020, 75, 327–336. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.-R.; Lee, J.H.; Park, K.S.; Jeon, J.H.; Kim, Y.B.; Cha, C.-J.; Jeong, B.C.; Lee, S.H. Antimicrobial Resistance of Hypervirulent Klebsiella pneumoniae: Epidemiology, Hypervirulence-Associated Determinants, and Resistance Mechanisms. Front. Cell. Infect. Microbiol. 2017, 7, 483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, N.; Yang, X.; Zhang, R.; Chan, E.W.; Chen, S. Tracking microevolution events among ST11 carbapenemase-producing hypervirulent Klebsiella pneumoniae outbreak strains. Emerg. Microbes Infect. 2018, 7, 146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marques, C.; Menezes, J.; Belas, A.; Aboim, C.; Cavaco-Silva, P.; Trigueiro, G.; Gama, L.T.; Pomba, C. Klebsiella pneumoniae causing urinary tract infections in companion animals and humans: Population structure, antimicrobial resistance and virulence genes. J. Antimicrob. Chemother. 2019, 74, 594–602. [Google Scholar] [CrossRef]
- Hu, Y.; Liu, C.; Shen, Z.; Zhou, H.; Cao, J.; Chen, S.; Lv, H.; Zhou, M.; Wang, Q.; Sun, L.; et al. Prevalence, risk factors and molecular epidemiology of carbapenem-resistant Klebsiella pneumoniae in patients from Zhejiang, China, 2008–2018. Emerg. Microbes Infect. 2020, 9, 1771–1779. [Google Scholar] [CrossRef]
- Donati, V.; Feltrin, F.; Hendriksen, R.S.; Svendsen, C.A.; Cordaro, G.; García-Fernández, A.; Lorenzetti, S.; Lorenzetti, R.; Battisti, A.; Franco, A. Extended-Spectrum-Beta-Lactamases, AmpC Beta-Lactamases and Plasmid Mediated Quinolone Resistance in Klebsiella spp. from Companion Animals in Italy. PLoS ONE 2014, 9, e90564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harada, K.; Shimizu, T.; Mukai, Y.; Kuwajima, K.; Sato, T.; Usui, M.; Tamura, Y.; Kimura, Y.; Miyamoto, T.; Tsuyuki, Y.; et al. Phenotypic and Molecular Characterization of Antimicrobial Resistance in Klebsiella spp. Isolates from Companion Animals in Japan: Clonal Dissemination of Multidrug-Resistant Extended-Spectrum β-Lactamase-Producing Klebsiella pneumoniae. Front. Microbiol. 2016, 7, 1021. [Google Scholar] [CrossRef] [Green Version]
- Hong, J.S.; Song, W.; Park, H.-M.; Oh, J.-Y.; Chae, J.-C.; Shin, S.; Jeong, S.H. Clonal Spread of Extended-Spectrum Cephalosporin-Resistant Enterobacteriaceae Between Companion Animals and Humans in South Korea. Front. Microbiol. 2019, 10, 1371. [Google Scholar] [CrossRef] [Green Version]
- Hartantyo, S.H.P.; Chau, M.L.; Fillon, L.; Ariff, A.; Kang, J.S.L.; Aung, K.T.; Gutiérrez, R.A. Sick pets as potential reservoirs of antibiotic-resistant bacteria in Singapore. Antimicrob. Resist. Infect. Control. 2018, 7, 106. [Google Scholar] [CrossRef]
- Sun, J.; Zhang, H.; Liu, Y.-H.; Feng, Y. Towards Understanding MCR-like Colistin Resistance. Trends Microbiol. 2018, 26, 794–808. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Tian, G.-B.; Zhang, R.; Shen, Y.; Tyrrell, J.M.; Huang, X.; Zhou, H.; Lei, L.; Li, H.-Y.; Doi, Y.; et al. Prevalence, risk factors, outcomes, and molecular epidemiology of mcr-1 -positive Enterobacteriaceae in patients and healthy adults from China: An epidemiological and clinical study. Lancet Infect. Dis. 2017, 17, 390–399. [Google Scholar] [CrossRef] [Green Version]
- Lei, L.; Wang, Y.; Schwarz, S.; Walsh, T.R.; Ou, Y.; Wu, Y.; Li, M.; Shen, Z. mcr-1 in Enterobacteriaceae from Companion Animals, Beijing, China, 2012–2016. Emerg. Infect. Dis. 2017, 23, 710–711. [Google Scholar] [CrossRef]
- Pulss, S.; Stolle, I.; Stamm, I.; Leidner, U.; Heydel, C.; Semmler, T.; Prenger-Berninghoff, E.; Ewers, C. Multispecies and Clonal Dissemination of OXA-48 Carbapenemase in Enterobacteriaceae From Companion Animals in Germany, 2009–2016. Front. Microbiol. 2018, 9, 1265. [Google Scholar] [CrossRef] [PubMed]
- Ewers, C.; Stamm, I.; Pfeifer, Y.; Wieler, L.H.; Kopp, P.A.; Schønning, K.; Prenger-Berninghoff, E.; Scheufen, S.; Stolle, I.; Günther, S.; et al. Clonal spread of highly successful ST15-CTX-M-15 Klebsiella pneumoniae in companion animals and horses. J. Antimicrob. Chemother. 2014, 69, 2676–2680. [Google Scholar] [CrossRef] [Green Version]
- Xia, J.; Fang, L.-X.; Cheng, K.; Xu, G.-H.; Wang, X.-R.; Liao, X.-P.; Liu, Y.-H.; Sun, J. Clonal Spread of 16S rRNA Methyltransferase-Producing Klebsiella pneumoniae ST37 with High Prevalence of ESBLs from Companion Animals in China. Front. Microbiol. 2017, 8, 529. [Google Scholar] [CrossRef] [Green Version]
- De Jong, A.; Muggeo, A.; El Garch, F.; Moyaert, H.; De Champs, C.; Guillard, T. Characterization of quinolone resistance mechanisms in Enterobacteriaceae isolated from companion animals in Europe (ComPath II study). Vet. Microbiol. 2018, 216, 159–167. [Google Scholar] [CrossRef]
- Struve, C.; Roe, C.C.; Stegger, M.; Stahlhut, S.G.; Hansen, D.S.; Engelthaler, D.M.; Andersen, P.S.; Driebe, E.M.; Keim, P.; Krogfelt, K.A. Mapping the Evolution of Hypervirulent Klebsiella pneumoniae. mBio 2015, 6, e00630-15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wacharotayankun, R.; Arakawa, Y.; Ohta, M.; Tanaka, K.; Akashi, T.; Mori, M.; Kato, N. Enhancement of extracapsular polysaccharide synthesis in Klebsiella pneumoniae by RmpA2, which shows homology to NtrC and FixJ. Infect. Immun. 1993, 61, 3164–3174. [Google Scholar] [CrossRef] [Green Version]
- Zou, Z.-Y.; Lei, L.; Chen, Q.-Y.; Wang, Y.-Q.; Cai, C.; Li, W.-Q.; Zhang, Z.; Shao, B.; Wang, Y. Prevalence and dissemination risk of antimicrobial-resistant Enterobacteriaceae from shared bikes in Beijing, China. Environ. Int. 2019, 132, 105119. [Google Scholar] [CrossRef]
- Insua, J.L.; Llobet, E.; Moranta, D.; Pérez-Gutiérrez, C.; Tomás, A.; Garmendia, J.; Bengoechea, J.A. Modeling Klebsiella pneumoniae Pathogenesis by Infection of the Wax Moth Galleria mellonella. Infect. Immun. 2013, 81, 3552–3565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A New Genome Assembly Algorithm and Its Applications to Single-Cell Sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Bi, Z.; Hulth, A.; Wang, Y.; Shen, Z.; Wang, S.; Wu, C.; Nilsson, L.E.; Walsh, T.R.; Börjesson, S.; et al. Inter-host Transmission of Carbapenemase-Producing Escherichia coli among Humans and Backyard Animals. Environ. Health Perspect. 2019, 127, 107009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Treangen, T.J.; Ondov, B.D.; Koren, S.; Phillippy, A.M. The Harvest suite for rapid core-genome alignment and visualization of thousands of intraspecific microbial genomes. Genome Biol. 2014, 15, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Alikhan, N.-F.; Petty, N.K.; Ben Zakour, N.L.; Beatson, S.A. BLAST Ring Image Generator (BRIG): Simple prokaryote genome comparisons. BMC Genom. 2011, 12, 402. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Category | No. of Samples (%) | Kp Isolates (%, 95% CI) |
|---|---|---|---|
| Origin | Dog | 3356 (62.6) | 85 (2.5, 2.0–3.1) |
| Cat | 2003 (37.4) | 20 (1.0, 0.6–1.5) | |
| Gender | Male | 3352 (62.5) | 58 (1.7, 1.3–2.2) |
| Female | 2007 (37.5) | 47 (2.3, 1.7–3.1) | |
| Source | Urine | 2879 (53.7) | 37 (1.3, 0.9–1.8) |
| Throat swabs | 200 (3.7) | 13 (6.5, 3.5–10.9) | |
| Nasal swabs | 194 (3.6) | 9 (4.6, 2.1–8.6) | |
| Abscess | 391 (7.3) | 10 (2.6, 1.2–4.7) | |
| Tracheal lavage | 126 (2.4) | 8 (6.3, 2.8–12.1) | |
| Ear swabs | 267 (5.0) | 8 (3.0, 1.3–5.8) | |
| Pyometra | 158 (2.9) | 7 (4.4, 1.8–8.9) | |
| Coelomic fluid | 367 (6.8) | 4 (1.1, 0.3–2.8) | |
| Skin | 452 (8.4) | 3 (0.7, 0.1–1.9) | |
| Oral swabs | 21 (0.4) | 2 (9.5, 1.2–30.4) | |
| Prostatic fluid | 22 (0.4) | 1 (4.5, 0.1–22.8) | |
| Surgical infection | 70 (1.3) | 1 (1.4, 0–7.7) | |
| Anal swabs | 32 (0.6) | 1 (3.1, 0.1–16.2) | |
| Eye secretion | 41 (0.8) | 1 (2.4, 0.1–12.9) | |
| Others | 139 (2.6) | 0 (0, 0–2.6) | |
| Total | —— | 5359 | 105 (2.0, 1.6–2.4) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Z.; Lei, L.; Zhang, H.; Dai, H.; Song, Y.; Li, L.; Wang, Y.; Xia, Z. Molecular Investigation of Klebsiella pneumoniae from Clinical Companion Animals in Beijing, China, 2017–2019. Pathogens 2021, 10, 271. https://doi.org/10.3390/pathogens10030271
Zhang Z, Lei L, Zhang H, Dai H, Song Y, Li L, Wang Y, Xia Z. Molecular Investigation of Klebsiella pneumoniae from Clinical Companion Animals in Beijing, China, 2017–2019. Pathogens. 2021; 10(3):271. https://doi.org/10.3390/pathogens10030271
Chicago/Turabian StyleZhang, Zhenbiao, Lei Lei, Haixia Zhang, Hegen Dai, Yu Song, Lei Li, Yang Wang, and Zhaofei Xia. 2021. "Molecular Investigation of Klebsiella pneumoniae from Clinical Companion Animals in Beijing, China, 2017–2019" Pathogens 10, no. 3: 271. https://doi.org/10.3390/pathogens10030271