1. Introduction
Since the beginnings of agriculture, there has been a struggle to avoid pest populations in order to guarantee our food supply [
1,
2]. The more abundant, severe and crop-threatening a pest is, the more effort is needed to solve the pest problem [
3]. Over time, however, environmental conditions can change, and some pests may become more frequent or have higher population densities [
4], as is the case for the sugar beet weevil (
Asproparthenis punctiventris Germar, previously
Bothynoderes punctiventris Germar, Coleoptera: Curculionidae) [
5,
6]. Problems caused by this pest as well as its control have been the focus of discussions since the 19th century [
7]. In Europe, the pest has occurred regularly, especially locally. Ukraine, Croatia, Romania, Bulgaria, Serbia, Hungary, Greece, Austria, Germany, and Italy are among the most affected regions [
8,
9,
10]. It is well known that the weevil prefers warm, dry weather to emerge from the soil and start emigrating [
11]. Therefore, the effects of climate change on environmental conditions–particularly in the months of May/June, which are crucial for the weevil’s mass appearance–have led to an increase in incidences of the insect pest and its spread to new regions. Furthermore, changes in the approval and availability of chemical insecticides have further exacerbated the problem [
6].
Metarhizium spp. Sorokin (Hypocreales: Clavicipitaceae) have long been known as a group of fungi with insecticidal potential. In Europe, the successful commercial use of
Metarhizium against the June and garden chafer (
Amphimallon solstitiale L. and
Phyllopertha horticola L., Coleoptera: Scarabaeidae), the black vine weevil (
Otiorhynches sulcatus F., Coleoptera: Curculionidae), wireworms (
Agriotes spp. L., Coleoptera: Elateridae) and the grape phylloxera (
Daktulosphaira vitifoliae Fitch, Hemiptera: Sternorryncha) is already well-established, and is now being tested to control the Japanese beetle (
Popillia japonica Newman, Coleoptera: Scarabaeidae; [
12]) or the western corn root worm (
Diabrotica virgifera virgifera LeConte, Coleoptera: Chrisomelidae; [
13]).
Biological control of the sugar beet weevil with entomopathogenic fungi has been neglected for the last 100 years. Only a few studies have reported the use of
Beauveria bassiana against the sugar beet weevil [
14,
15],
Metarhizium spp., however, was hardly mentioned. Interestingly, the first fungal infections of the weevil caused by
Metarhizium spp. were already reported at the end of the 19th century, and the first large-scale treatment with
Metarhizium was also first carried out against this pest around this time [
7,
16]. Despite Krassilstchik’s reference [
7] to the great potential of this treatment approach, further trials were discontinued in the 20th century. This may be due to the different mode of action of fungal biopesticides compared to the chemically synthesized insecticides. Whereas chemical insecticides are supposed to kill shortly upon contact with the insect, the fungal spores have to adhere, germinate, penetrate the cuticula of the insect and multiply within the insect to have an effect. These key stages in the invasive and developmental processes of the fungus are time-consuming [
17]. Nevertheless, there are major advantages in using entomopathogenic fungi against insects. The fungus can be applied to the soil as well as to surfaces. This enables the control not only of the adult weevil but of all developmental stages of the pest, including its larvae. As the spores have a certain resistance to environmental influences, an application does not have to directly coincide with an invasion of the insects [
18].
Integrated pest management in sugar beet cultivation is still oriented towards crop-specific requirements. It attempts to continue the use of so-called effective and “intelligent” solutions [
19], such as seed dressing with the insecticidal active ingredient clothianidin from the group of neonicotinoids, permitted under an emergency use authorization under Article 53 of Regulation (EC) No 1107/2009. This is what happened in numerous European member state countries in 2022, including Austria, where a temporary emergency authorization for the neonicotinoid treatment of beet seed was granted anew. The further development of integrated pest management and alternative methods or procedures is increasingly in focus in order to reduce the dependence on repeated emergency authorizations. The overall goal should be the reduction of chemical synthetic plant protection products and replacing these with alternative products such as fungal biopesticides. These fulfil the requirement of a sustainable control approach based on natural antagonists. Our strategy is to establish a holistic, alternative control option for all developmental stages of
A. punctiventris as well as to monitor its emigration to better predict and plan further control measurements.
The major goals of this work were to (i) assess the pathogenicity of the commercially used fungus Metarhizium brunneum Petch Ma 43 (previously M. anisopliae var. anisopliae (Metsch.) Sorokin BIPESCO 5/F52) products against the sugar beet weevil; (ii) test different application methods in the sugar beet field for a sustainable establishment of the fungal active agent; (iii) evaluate the rate of fungal infection of the weevils by locally occurring pathogens under optimal fungal growth conditions (25 °C, 60% rH) and (iv) determine the migration of the weevil from the sugar beet fields in Lower Austria.
2. Materials and Methods
2.1. Trial Sites
Field trials were performed in the most important sugar beet growing area of Lower Austria, which was severely affected by the pest
A. punctiventris in 2018 and 2019. Seven field sites were chosen (
Table 1) to perform duplicates of four different treatment approaches (2.2). In addition, a further trial site was chosen for the spray treatments on sugar beet leaves (field J, n = 24 plots). To determine the natural mycosis rate of the beetles by entomopathogenic fungi, two additional fields were selected that had never been treated with
Metarhizium spp. The field sites were selected according to routine crop rotation of sugar beet cultivation and predicted weevil infestation (based on monitoring in the previous autumn of pupae/beetles in the fields).
Pot experiments were performed at the University Innsbruck in an open greenhouse. Pots of 40 cm length, 15 cm width and 15 cm depth were filled with a
Metarhizium-free (the soil was tested for the presence of entomopathogenic fungi before use [
20]) standardized soil mixture containing peat, sand and lava granules (ratio 3.5:1:1). All pots were planted with six sugar beet seedlings (organic quality) per replication and left undisturbed until the treatment (n = 4 per treatment). Water was not applied separately. The soil was not further fertilized.
In addition, 13 field sites (further referred to as “Emigration fields”,
Table S1), were selected to evaluate the emigration of weevils (2.3). The fields were chosen based on the results obtained from autumn monitoring of overwintering weevils the previous year, which predicted a high number of migrating weevils. No application was performed on the field sites.
2.2. Fungal Application Strategies
The strain
M. brunneum Ma 43 was applied as granulare (GranMet
TM-P, Agrifutur Srl., Italy) or spore dispersion (GranMet
TM-WP, Agrifutur Srl., Italy). The quality of the used products was assessed in the laboratory before application. Assessed parameters were spore density, purity, colonization ability (for granular product) and strain identity [
20]. Different application strategies were used to gain insight into advantages and disadvantages of the strategies against the sugar beet weevil.
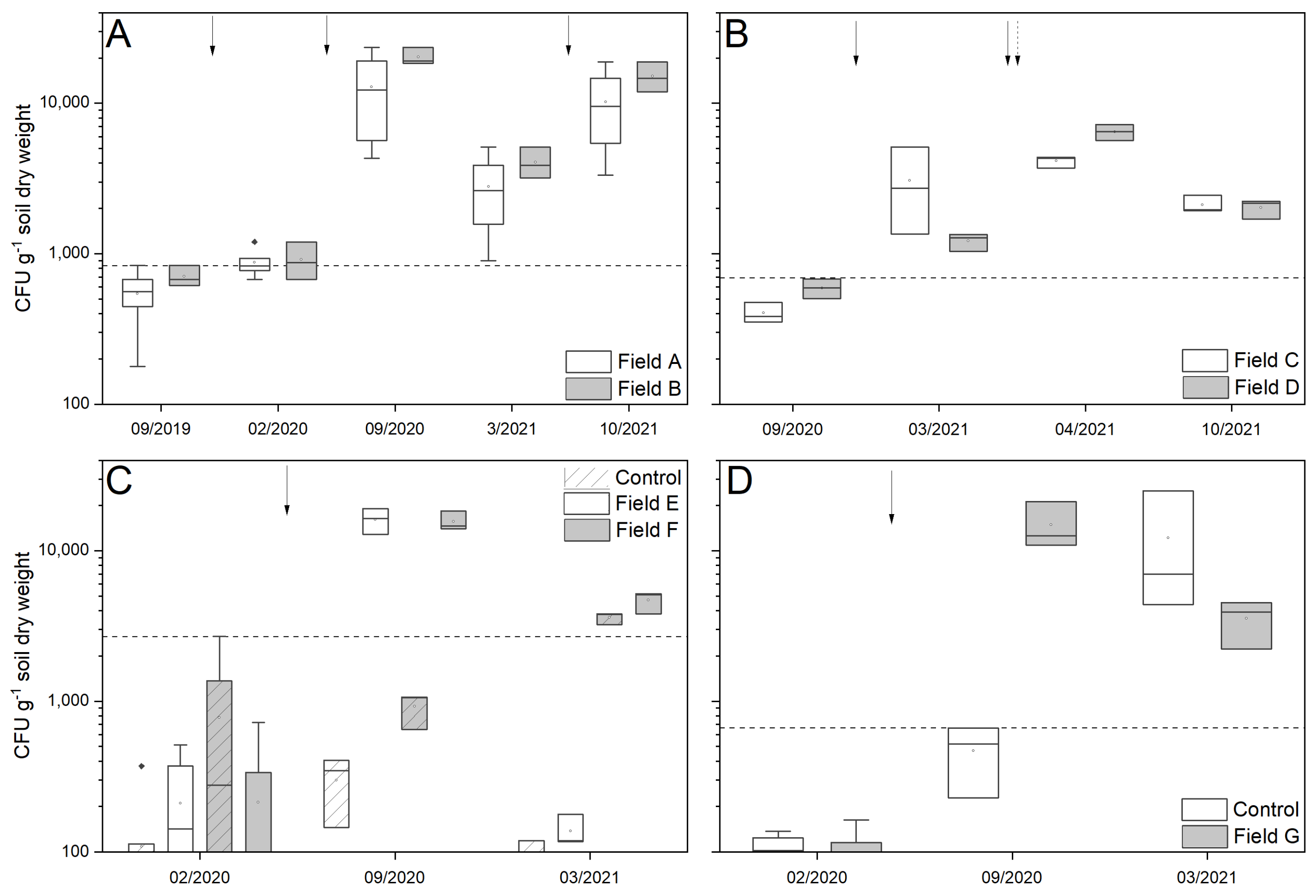
In a long-term trial, the field sites A and B were treated in autumn 2019, spring 2020 and spring 2021 with 100 kg ha−1 GranMetTM-P. The application of the product was performed by the farmers directly before the sewing of sugar beet seed using a fertilizer spreader and incorporating it into the soil with a power harrow. With the single annual application, the persistence of the fungus in the treatment areas was assessed.
Field sites C and D were treated with a combination of granular and liquid application. In autumn 2020 and spring 2021, 100 kg ha
−1 of the granular product GranMet
TM-P was applied into the soil as previously described. To determine the start of sugar beet immigration, sugar weevil migration was regularly monitored by the authors and farmers. Furthermore, the current weather situation was taken into account, as it influences the behavior of the weevils and the migration time. In addition, the depth at which the weevils were located in the soil was determined by digging. Just before weevil immigration started, the field sites were surrounded by a trap ditch (12 cm width and 40 cm depth,
Figure S2A) produced with a cable trencher (Cosmeco V1). This trap ditch plus a strip of in total 12 m (due to the working width of the machine) was treated with GranMet
TM-WP with a spore density of 1 × 10
12 spores ha
−1. The liquid formulation was applied with a saddle tank sprayer, which is normally used for the application of liquid chemicals. This dual application should ensure contact of the adult weevils as well as of the newly laid eggs or the emerging larvae with the infectious fungal spores, enabling the control of all developmental stages.
In addition to the large-scale trials, plot trials were performed at three field sites (Field E-G). At the field sites E and F, sugar beet was grown before the application in 2019, whereas field site G was treated before cultivation of sugar beet in 2020. In each field, 12 plots with a size of 5 × 5 m were created, of which half were randomly selected and manually treated with GranMet
TM-P in spring 2020. The remaining plots were addressed as controls. On each plot, a cage with 2 × 2 m was installed (
Figure S1). At field site G, adult
A. punctiventris (n = 20) were introduced into each cage. In all other fields, treatments were conducted with the naturally occurring population. One trap per cage was used in the plot trials in 2020, as well as in 2021.
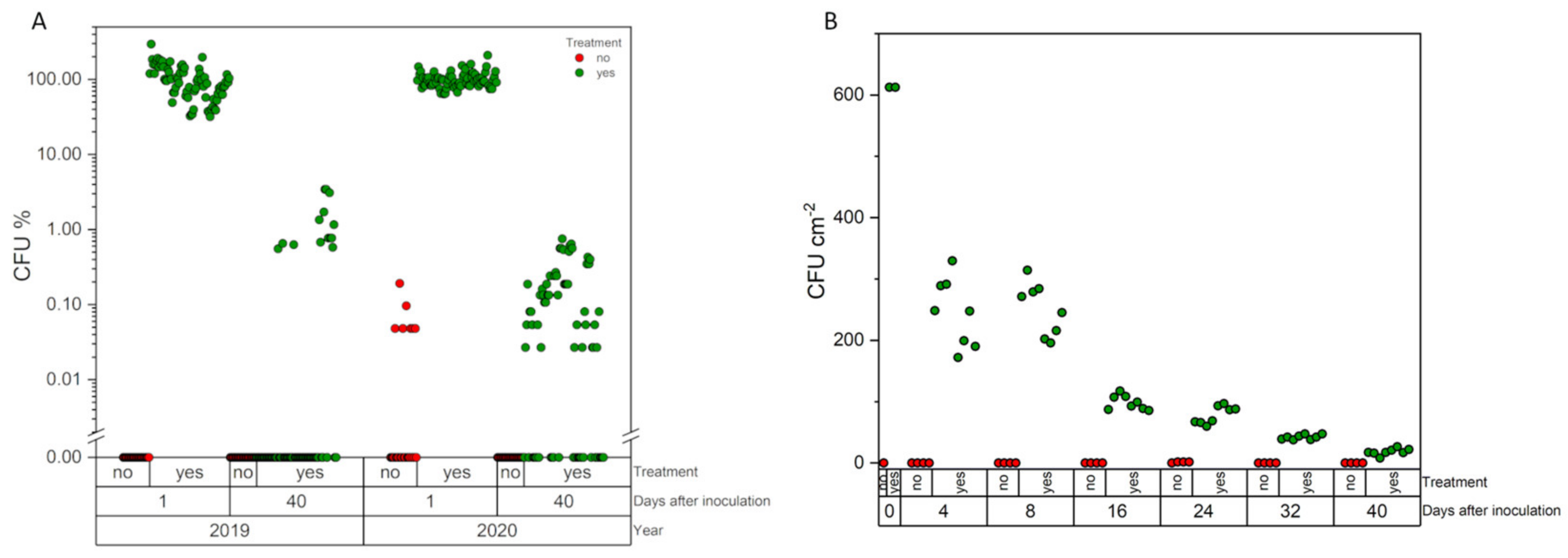
On field J, 24 (2 × 2 m) cages (12 control, 12 treatment) were set up. The sugar beet plants were spray treated with 1 × 1013 spores ha−1 M. brunneum Ma 43 as a dispersion formulation with 0.05% (v/v) NeoWett® (wetting agent based on isotridecanol-polyglycolether, Kwizda Holding GmbH, Vienna, Austria) and 1.5% (v/v) PRESS® Antischiuma (foam reducing agent based on silicone oil, Cifo Srl Bologna, Italy). Before treatment, 20 weevils were introduced into these plots. The same treatment was performed on the sugar beet plants of the open greenhouse experiment, but without the weevils.
2.3. Pest Monitoring
Pheromone traps were used to assess the infestation density of the weevil population. The traps consisted of a bucket (volume 3 L) and a piece of natural rubber mounted in the center with wire, to which an aggregation pheromone (Grandlure III&IV; Bedoukian Research Inc., Danbury, CT, USA) was added. Traps were buried in the soil (
Figure S2B) around the field borders with a distance of 10 to 15 m. The beetles are attracted by the pheromone, fall into the bucket and cannot escape due to the smooth walls. All traps were evaluated once a week, and the caught weevils were counted. Weevils from the plot trial were separated for further analysis (i.e., to determine the prevalence of weevil infection).
To evaluate whether A. punctiventris can be infected with the fungus by treated trap ditches, 300 weevils were collected from fields C and D shortly after they had entered the fields. Ten days after the spray application, weevils were manually collected from the trap ditch and up to 2 m in the field site and separated for quarantine. For comparison, additional control fields, which were untreated, were sampled. Two different approaches were performed: 180 weevils were manually caught from field H and individually kept for quarantine. For comparative sampling, pheromone traps were placed around the borders of field site I in the morning and all weevils were collected from the bucket in the afternoon, and then separated for quarantine. This should highlight whether the direct contact of weevils increases endemic mycosis. All weevils were kept at conditions optimal for fungal growth, 25 °C and 60% rH, and fed regularly with organic grown sugar beet seedlings. Upon death, cadavers were evaluated for fungal growth. Fungal isolates were taken from mycosed weevils and identified by morphology. Metarhizium spp. isolates were further prepared for molecular genetic identification.
At the Emigration field sites, the number of weevils leaving the field was assessed to determine whether there was selective directional migration of the beetles that could be attributed to for instance wind direction, position of the sun or neighboring sugar beet fields to better adjust future control measurements. For this purpose, the fields were closely bordered with buried pheromone traps (10 m distance between traps) and the weevils in the traps were counted weekly.
2.4. Evaluation of Metarhizium Abundance
Soil samples of all field sites were taken once before treatment and twice a year after application of the products with a core borer (for two years in a row). Samples were processed after Laengle et al. [
20], plated onto selective Sabouraud-4%-Glucose agar media and were incubated for 14 days at 25 °C and 60% rH before counting colonies. Final abundance was calculated as a colony forming unit (CFU) per gram soil dry weight.
The abundance of
Metarhizium on sugar beet leaves was determined by “washing” the spores off the leaves (10 leaves per plot were pooled). For this purpose, they were shaken for one hour in 0.1% (
w/
v) Tween 80 solution (0.6 to 10 cm
2 mL
−1), on a rotary shaker at 200 rpm in an Erlenmeyer flask. The solution was filtrated through a cotton plug which was inserted into a sterile 10 mL pipette (used upside down, [
21]) to avoid leaf fragments in the spore suspension. From this suspension, 50 µL were plated in fourfold onto selective Sabouraud agar media. Incubation of plates was performed as mentioned above for the agar plates from soil samples. Abundance was calculated as CFU per square cm leaf surface.
2.5. Bioassay
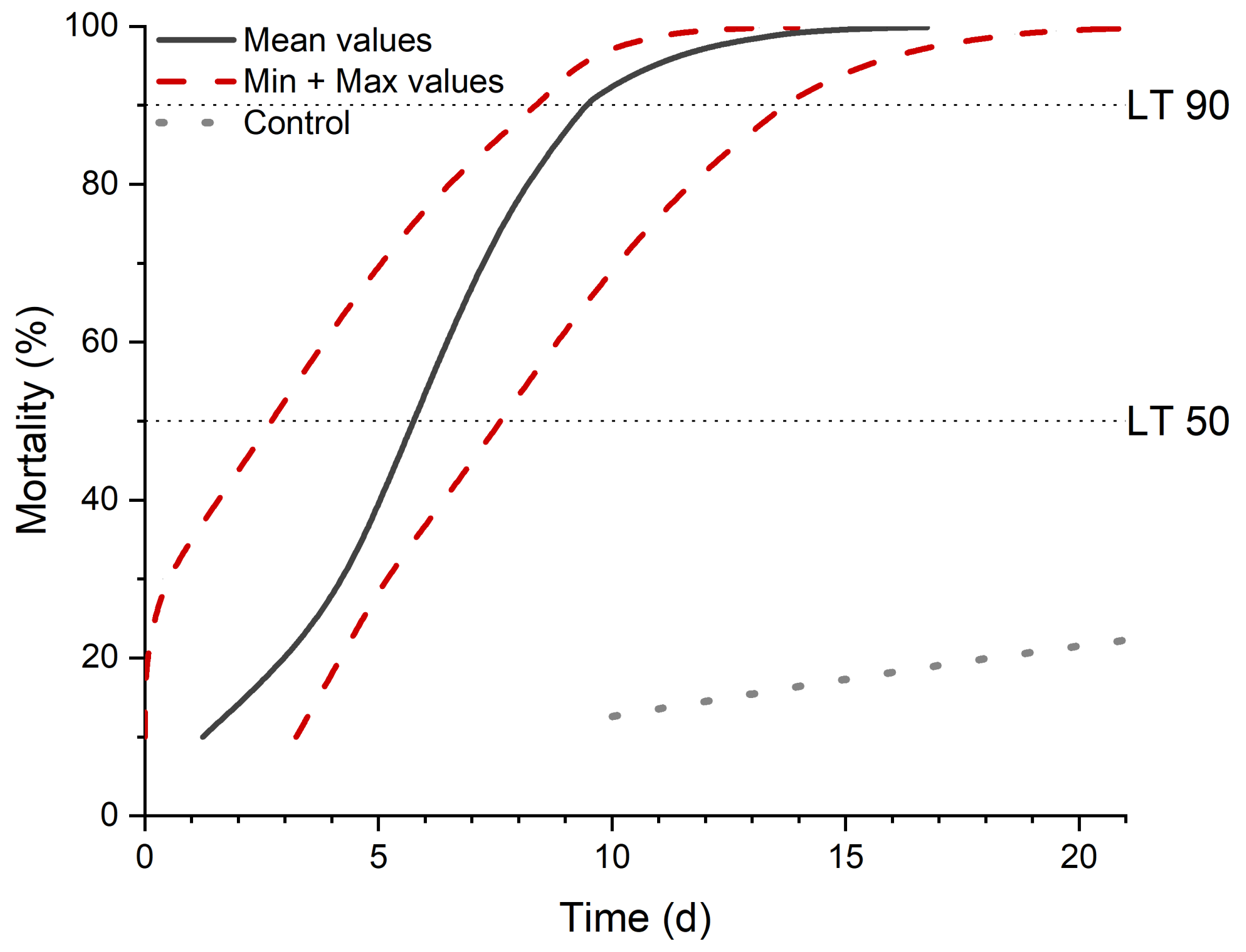
For the bioassay (n = 4), sugar beet weevils were collected from an untreated field site and separated into 50 mL tubes containing entomopathogen free soil. After a 14-day quarantine, 18 weevils per treatment were chosen and put into 5 cm Petri dishes containing a moist paper towel. The weevils were directly inoculated with the spore suspension to ensure contact of host and pathogen. They were either treated with 50 µL 0.1% (w/v) Tween 80 solution (control) or with M. brunneum Ma 43, spore density of 5 × 106 spores per weevil. The bioassay was incubated at 25 °C in a moist chamber, and the mortality of A. punctiventris was evaluated every day until all fungal treated weevils died.
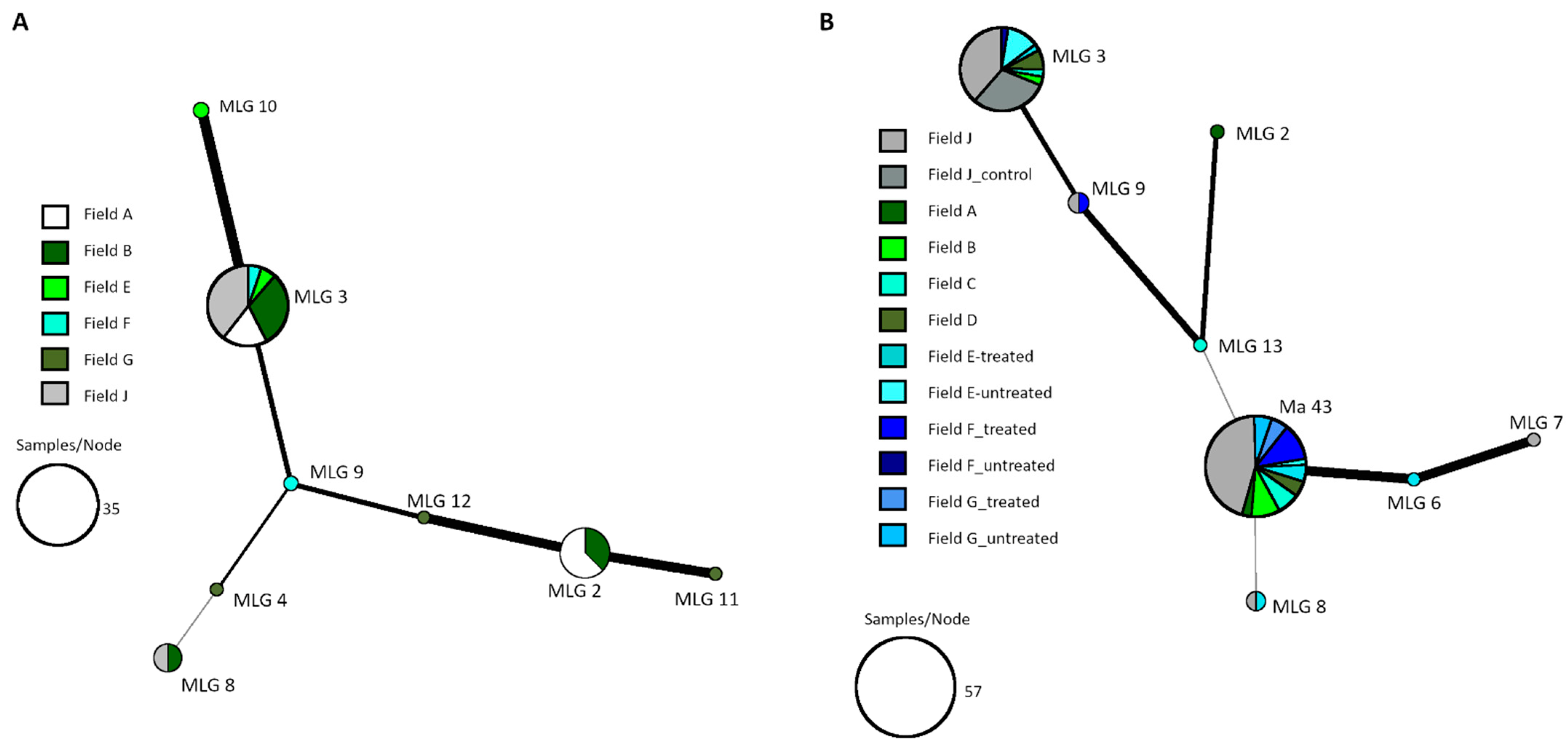
2.6. Genetic Analysis of Fungal Isolates
To evaluate successful application of the product, isolates from the soil samples were identified with simple satellite repeat (SSR) PCR analysis. Therefore, from each sample two isolates belonging to the most common morphotype and one isolate with another morphology were selected. In addition,
Metarhizium isolates from the mycosed sugar beet weevils, collected during the monitoring, were analyzed. The PCR was performed after Mayerhofer et al. [
22] with the two marker sets I and V (Ma 2049, Ma2054, Ma2063, Ma195, Ma327, Ma2287). Species were assigned either based on comparison to already known genotypes from our previous work, or sequencing of the nuclear translation elongation factor-1α (EF1a) [
23].
2.7. Data Analysis
Statistical analyses were performed with R version 4.1.0 (Free Software Foundation, Boston, MA, USA). The minimum spanning network (MSN) was created using the “poppr” package of R. Graphics were plotted with OriginPro 2020 (version 9.7.0.185; OriginLab Corporation, Northampton, MA, USA). SSR raw data were analyzed with GeneMarker (SoftGenetics, State College, PA, USA), sequencing data with BioEdit [
24]. Phylogenetic tree was calculated with MEGA [
25]. The bioassay was evaluated using Mathematica (Wolfram Research, Inc., Champaign, IL, USA) and a time- mortality analysis developed by Throne et al. [
26], in which a formula is included to correct the treatment mortality with the control mortality.
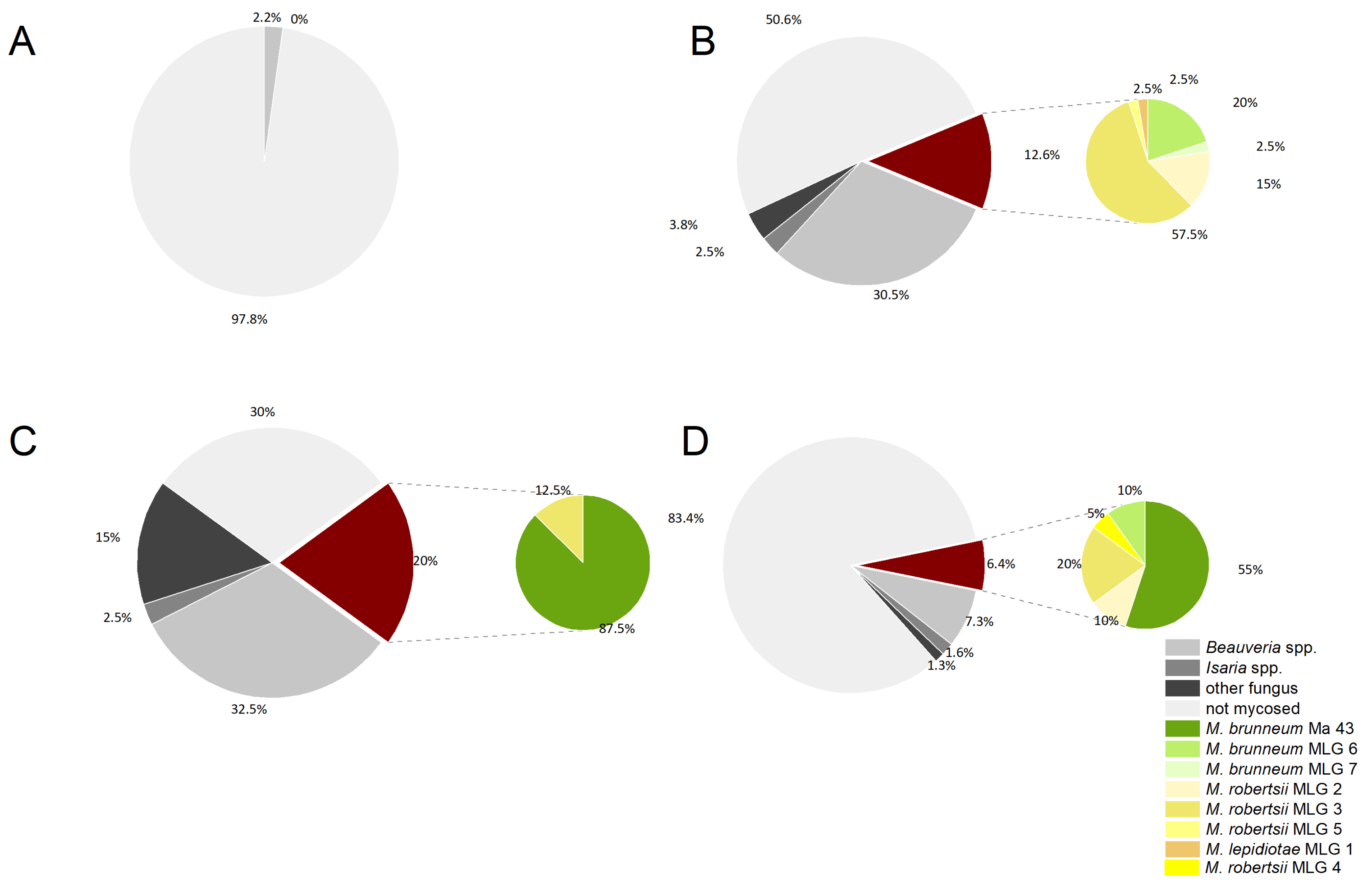
4. Discussion
Apart from the use of nematodes [
27,
28], which were only used against the larvae of the sugar beet weevil
A. punctiventris in the soil, this large-scale study is one of the first in almost a hundred years (except a few studies using
Beauveria bassiana [
14,
15]) to test the use of a biologically based insecticide against the weevil pest. We aimed to test the effectiveness of a preventive use of the entomopathogenic fungus
Metarhizium brunneum against the weevil in all stages of its development. To directly control the adult beetles, a foliar spray application containing the same fungus was carried out (field J). In addition, a spray treatment of the trap ditches with
M. brunneum was performed when the adult beetles were immigrating into the sugar beet field (field C and D).
For the spray treatment trials, the spore viability of the powder product GranMet
TM-WP was tested, and accordingly a maximum of 4.4 kg per hectare was used. Although this dosage was lower than that mentioned in older literature, our production strain
M. brunneum Ma 43 could nevertheless be established in the treated areas over a period of several months and remain at a fungal density of >1000 CFU g
−1 soil dry weight. This persistence was above the maximum background level defined by Scheepmaker et al. [
29]. The conclusion that
Metarhizium spp. seems to be suitable for in situ use in sugar beet cultivation is further supported by the fact that more than a hundred years ago, the fungus was already shown to successfully infect and kill the sugar beet weevil in a sugar-beet field [
9]. However, a direct comparison of older studies from the nineteenth and twentieth centuries on the use of
Metarhizium spp. against the sugar beet weevil is only useful and possible to a limited extent, due to the small number of conducted trials as well as the lack of direct pathological findings. For example, Krassilstchik [
7] achieved a 55–80% reduction in beetle density after about 2 weeks with a spore application of 8 kg per hectare, but there is no information on the actual number of vital spores the author used per hectare. Furthermore, abundance data for this application was not published either, so that we can only speculate about the actual spore density and its persistence. In addition, our abundance values of more than 1000 CFU g
−1 soil dry weight after the application of the products were in contradiction with the results published by Pospelov [
30], who performed similar experiments. The author could only prove
Metarhizium infections in the trap ditches, but no accumulation of the fungus in the soil, nor an effect of the fungus on adult beetles when applied directly in the field. The reasons for this cannot be determined, because no information on the used spore concentrations, the application methods, the quality of the spore product nor the product formulation could be gained from Pospelov’s report [
30].
In their studies, Krassilchik [
7], Ivanov [
31] and Pospelov [
30] observed mycosis rates of 20–80% and 55–80%, respectively. The authors indirectly attributed the strong deviations in the abundance values to the pH value of the soil in the experimental fields: Pyatnitzkiï [
32] reported that
Metarhizium successfully infects insects, especially in more acidic soils. Pospelov [
30] advocated the use of
Metarhizium, especially in the area around Kiev, because he found that the “white muscardine”, i.e., the infestation with the insect-killing fungus of the genus
Beauveria spp. was not successful in this region. The soil pH around Kiev is slightly acidic, which would also confirm the authors’ theory that acidic soils can improve the persistence and development of
Metarhizium. However, the sites investigated in our study were characterized by neutral to alkaline soils [
33]. Nevertheless, our abundance values showed a maximum density of nearly 20,000 CFU g
−1 soil dry weight, which were directly attributable to the presence of our applied production strain
M. brunneum Ma 43. Interestingly, we also found a high indigenous density of
Beauveria spp. in our soil samples. In this study, we focused on
Metarhizium spp. density and genetic composition, but analyzing the
Beauveria spp. population composition and virulence might be addressed in further studies. Previous experiments have indicated that the abundance of applied biological control agent (BCA) strains is promoted by pest infestation and application frequency of the active ingredient. Abiotic and edaphic factors are principally important test criteria but are usually of secondary importance, and their influence on the fungus is often not yet well understood [
34,
35]. Unfortunately, due to the very inhomogeneous distribution of pest infestation, we could not observe a direct effect on the pest density. In our first control approaches, the weevils were already present or were just about to infest newly planted sugar beet fields. As such, the fungus only had a short time to infect the host before the plants were damaged. To prolong the possible infection time frame and increase efficacy, one should consider applying the fungus in spring, before the start of migration, on fields where sugar beet was grown in the previous year. Furthermore, weevils emigrating from these fields should be trapped in treated trap ditches.
For successful biological control, the importance of knowledge of the biology of
Asproparthenis should not be neglected. We were able to show that the weevils not only hatch and emerge in the year following sugar beet planting, but also two years after the first infestation as a migratory beetle: even in the second year of crop change, about 3 percent of the beetles emerged from the soil of the experimental plots. Regarding a sustainable control of the
Asproparthenis population, this observation is of utmost importance, because despite a supposedly low number of weevils, the population of the beetle in the region can increase significantly. Assuming that each female lays between 80 and 100 eggs on average [
11,
36], the damage threshold of the weevil in the cultivation regions (i.e., damage threshold for sugar beet weevils is 0.1 insects m-
2 during emergence of beet seedlings, [
37]) can be reached again by the surviving weevils within the next generation (considering optimal conditions for egg weevil development). This active emigration of adults from old sugar beet fields over several years was also described by Pyatnitzkiï [
32,
38] and could also explain the increase in population density in the cultivation region in Lower Austria despite crop change (annual crop change with a four-year gap between two sugar beet plantings). In contrast, Tielecke [
11] did not observe any weevils that hibernated for two years, but attributed this to the low overwintering depth of the pest population (75% at 10 cm depth). This is in accordance with the findings of Pyatnitzkiï [
32], who was able to establish a connection between the overwintering depth and the hibernation length of the beetles (dormant weevils in depths of 40–50 cm). However, while Pyatnitzkiï’s studies reported a relatively high proportion of dormant beetles (up to 20% for two years, up to 10% for three years; [
38]), in the second year the proportion of emerging beetles in our experimental plots was six times lower (overwintering depth 10–50 cm).
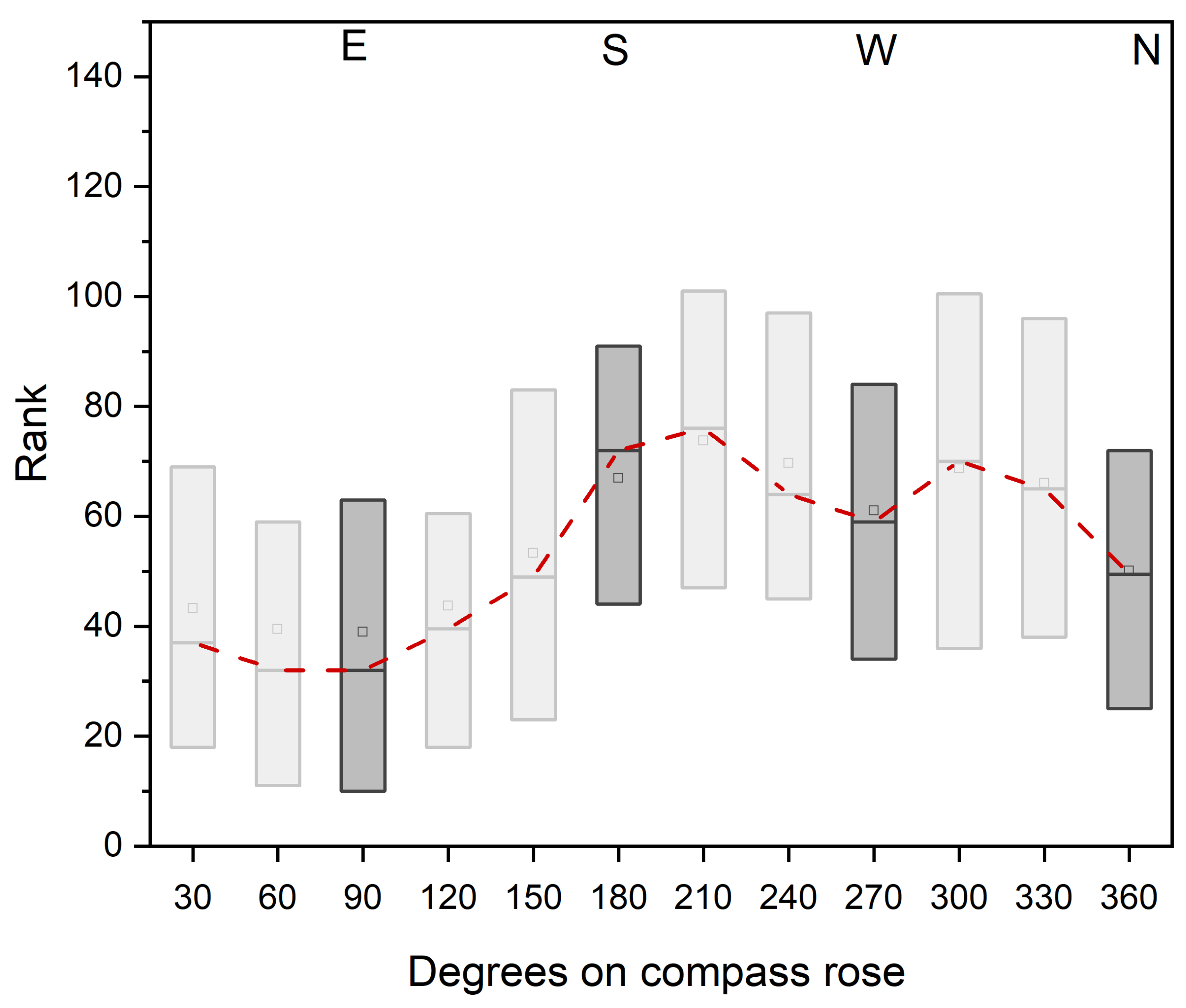
The direction of the beetles’ migration is also of importance for a pest prognosis and, consequently, for preventive control measures. For this reason, the migration of the weevil from thirteen different field sites across the entire sugar beet cultivation area in Lower Austria was investigated. Varying observations from previous investigations (e.g., no direction of migration, migration according to location of sun, migration according to soil grooves) do not allow a clear conclusion to be drawn [
10,
11,
32]. Although at present no significant cardinal direction of emigration could be proven in this study either, a tendency to migrate towards the south and west could be detected. For further studies it is necessary to pay attention to other environmental factors (including location of nearest beet field, vegetation surrounding the field, wind direction, other topological obstacles) as well as their combined effect. If the direction of emigration can be predicted, this will enable a more targeted
Metarhizium treatment. Similarly, to an application technique mentioned by Hunter [
39], barrier strips in the migration direction of the weevil can be established, thus increasing the possibility of a successful fungal infection and leading to a higher mortality of the emigrating weevils.
This first large-scale field trial will serve as the promising basis for further research into the right combination of IPM measures and additional studies of the biology of the weevil. We also consider it important to apply the fungus repeatedly as well as before the weevil starts emigrating. This increases the likelihood of controlling all development stages of the weevil. Sugar beet fields which were cultivated in the previous year should be treated in spring shortly before the start of migration, in combination with other IPM measures (e.g., also in combination with trap ditches). Initial spray treatments of the trap-ditches have been effective, and are now being investigated in more detail for sustainable barrier system support as part of a follow-up project.

 ,
, 







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}