
Role of Gut Microbiota in Breast Cancer and Drug Resistance
by
 ,
,
Sathiyapriya Viswanathan
1,
Sheetal Parida
2,
Bhuvana Teja Lingipilli
3,
Ramalingam Krishnan
4,
Devendra Rao Podipireddy
5 and
Nethaji Muniraj
6,* 1
Department of Biochemistry, ACS Medical College and Hospital, Chennai 600007, Tamil Nadu, India
2
Department of Oncology, Johns Hopkins University School of Medicine, Baltimore, MD 21231, USA
3
Gandhi Institute of Technology and Management (GITAM), Deemed University, Visakhapatnam 530045, Andhra Pradesh, India
4
Department of Biochemistry, Narayana Medical College, Nellore 524003, Andhra Pradesh, India
5
Rangaraya Medical College, Dr. YSR University of Health Sciences, Kakinada 533001, Andhra Pradesh, India
6
Center for Cancer and Immunology Research, Children’s National Hospital, 111, Michigan Ave NW, Washington, DC 20010, USA
*
Author to whom correspondence should be addressed.
Pathogens 2023, 12(3), 468; https://doi.org/10.3390/pathogens12030468
Submission received: 28 January 2023
/
Revised: 8 March 2023
/
Accepted: 13 March 2023
/
Published: 16 March 2023
(This article belongs to the Special Issue Role of Microorganisms in Breast Cancer)
Abstract
:Breast cancer is the most common malignancy in women worldwide. The cause of cancer is multifactorial. An early diagnosis and the appropriate treatment of cancer can improve the chances of survival. Recent studies have shown that breast cancer is influenced by the microbiota. Different microbial signatures have been identified in the breast microbiota, which have different patterns depending on the stage and biological subgroups. The human digestive system contains approximately 100 trillion bacteria. The gut microbiota is an emerging field of research that is associated with specific biological processes in many diseases, including cardiovascular disease, obesity, diabetes, brain disease, rheumatoid arthritis, and cancer. In this review article, we discuss the impact of the microbiota on breast cancer, with a primary focus on the gut microbiota’s regulation of the breast cancer microenvironment. Ultimately, updates on how immunotherapy can affect the breast cancer-based microbiome and further clinical trials on the breast and microbiome axis may be an important piece of the puzzle in better predicting breast cancer risk and prognosis.
1. Introduction
Cancer is the second most common cause of death in the world after heart disease. In 2022, approximately 287,850 cases of breast cancer were diagnosed in women. In addition, approximately 51,400 new cases of breast ductal carcinoma in situ were diagnosed in women [1]. Breast cancer is a heterogeneous disease that is considered to be complex, with great diversity within and between tumors [2]. Despite extensive research, the exact etiology of the disease has not yet been elucidated, as the identified genetic and epigenetic interactions cannot explain the cause of breast cancer in most cases [3]. Therefore, there must be an unexplored pathway that contributes to the development of breast cancer. Studies on cancer risk factors have shown that microorganisms can contribute to the development of cancer in 15–20% of cases [4]. Many of the associations between the gut microbiota and disease are related to both the composition of the microbiota and the specific types of microbes involved in disease development. The relationship between gut microbiota and cancer is unknown.
The human body is a complex symbiotic system of microbial cells and human host cells. There are 100 trillion microbes in our body, distributed everywhere, including the gastrointestinal tract, holding enormous colonies of microorganisms, with their genomes being 150 times larger than the genome of the host cell. These microorganisms comprise the second genome of the human body, and play an important role in health and disease [5,6,7,8]. With advances in sequencing techniques, dysbiotic microbial signatures have been identified that are involved in modulating the onset and progression of various diseases, including cancer [9]. Studies have strong evidence that microbiome composition and regulation, as well as the presence of specific microbes, can initiate tumor formation and promote tumor growth in vivo [10,11,12,13].
Most microbiomes induce cancer through three main mechanisms: 1. altering the balance between cell proliferation and death; 2. controlling the host’s immune system; 3. controlling the host’s metabolism. The microbiota shifts the balance toward cell proliferation by affecting host Wnt/β-catenin, a pathway that maintains cell polarity and growth, and is thus implicated in tumor progression. Proteins such as FadA (Fusobacterium nucleatum), Avra (Salmonella typhi) and CagA (Helicobacter pylori) activate β-catenin, and cause colorectal, hepatobiliary and stomach cancer, respectively [14]. Some bacterial toxins also mediate double-stranded DNA breaks, which the microbiota have developed as a survival mechanism to kill hostile bacteria in its environment. These toxins also mediate cellular DNA damage that leads to carcinogenesis [15]. Colibactin from Enterococaceae, Bacteroides fragilis toxin (BFT) from B. fragilis, and cytolethal expansive toxin (CDT) from ε/γ proteobacteria are some examples of toxins involved in DNA double-strand breaks [16]. The microbiota can control the host’s immune system and mediate inflammation, which plays an important role in cancer development. Both innate and adaptive immune responses are stimulated by the microbiota [17].
Gut microbiota can alter the efficacy and side effects of cancer treatments. It is clear that the gut microbiota is a double-edged sword in cancer immunotherapy. The gut microbiome contributes to tumor initiation and progression by inducing tumor-promoting inflammation, or by modulating the local tumor microenvironment through its effects on tissue remodeling and mucosal immunity [18]. In addition, it has been suggested that some gut microbes can protect the tumor microenvironment and regulate the anti-cancer immune response, as well as protect the host from inappropriate inflammation. In this review, we provide a broader perspective on the impact of the gut microbiome on the breast cancer microenvironment. Ultimately, the microbiome has direct and indirect effects on cancer immunotherapy, thereby influencing tumor growth and the therapeutic response. Finally, we describe clinical trials targeting the breast cancer and microbiome axis, which may be important for the better prediction of breast cancer risk.
2. Microbiome and Breast Cancer, the Connection
According to a 2018 global study, 13% of global cancer burden can be attributed to microbial infections, including both bacterial and viral infection, and this shows a clear geographical association [19]. While the causative agents of cancer were determined to be H. pylori, Human papilloma virus, Hepatitis B virus (HBV) and Hepatitis C virus (HCV) [19], it should be noted that the human microbiome is composed of 10–100 trillion microbial partners, most of which remain unidentified. The highest rate of infection associated with cancer development was identified as being in eastern Asia, followed by sub-Saharan Africa, and then by northern Europe and west Asia. China alone accounted for one-third of the cancers driven by H. pylori and human papilloma viruses [19]. Therefore, the role of microbes in cancer requires increased attention. Breast cancer is not a single disease, but a variety of different cancers all affecting the breast. Molecular subtyping based on the presence or absence of cell surface receptors, such as ER, PR and Her2, therefore, is used to determine the correct treatment strategy. Patients lacking all three receptors or markers, or triple-negative breast cancer (TNBC) patients, have many adverse outcomes, due to lack of targeted therapies. Breast cancer is a multifactorial disease, potentially impacted by age, lifestyle, parity, exposure to carcinogens and genetics. However, 70% of breast cancers are detected without any known risk factors, other than those of being a woman and being above 50 years of age. Owing to advanced early detection, modern treatment strategies and the public awareness of breast cancer incidence has increased for the past four decades, by 0.5% annually, while mortality has dramatically declined [20]. However, there is an inconsistency in predicting outcomes, particularly among younger and socially disadvantaged women, suggesting that other factors are at play. With the advent of modern sequencing technologies and multicentric techniques, the microbiota has emerged as a potential determinant of breast cancer severity and mortality. Breast cancer can be affected by the local breast microbiota or gut microbiota, either positively or negatively. The microbiota can increase/decrease the risk of breast cancer by regulating circulating steroid hormone levels, regulating energy intake and utilization, synthesizing metabolites, such as genotoxins, lipopolysaccharides, vitamins, and antibiotics, and modulating the immune system.
3. The Microbiota of Breast and Breast Tumor
The upsurge in human microbiome research was the direct consequence of the findings of the human microbiome project (HMP), which was initiated in 2007. By 2016, the microbiota of five different body sites of 300 healthy individuals, including the nasal cavity, oral cavity, skin, gastrointestinal tract, and urinogenital tract, was characterized and made publicly available. This marked the beginning of the second phase of the HMP, the integrated HMP (iHMP), which focuses on three non-infectious health conditions, pregnancy and pre-term birth, onset of inflammatory bowel disease and onset of Type 2 diabetes [21]. The breast was considered sterile until Urbaniak et al. proposed the presence of a distinct microbial population in the breast that persisted beyond lactation [22]. Eventually they and other groups, using deep sequencing techniques, such as 16s rRNA sequencing and shotgun sequencing, proved that there is indeed a microbial community living in breast tissue that is significantly altered in breast cancer [23,24]. Moreover, differences were evident between malignant and benign breast cancers and also between different subtypes of breast cancers [24]. Several bacterial genera have been significantly associated with breast cancer. Tzeng, A. et al. examined the 16S rRNA gene sequence of human breast tissues compared to controls. A distinct microbial profile was associated with each histologic tumor subtype; for example, invasive ductal carcinoma (IDC) was characterized by the presence of Tepidiphilus, Alkanindiges, and Stenotrophomonas, while samples of invasive lobular carcinomas (ILC) contained Peptostreptococcus, Micromonospora, Faecalibacterium, and Stenotrophomonas [25]. However, their bioinformatic analysis showed that Porphyromonas, Lacibacter, Ezakiella, and Fusobacterium were more abundant at a more advanced stage than in lower-stage tumors [25].
Parhi et al. showed that an oral pathogen, Fusobacterium nucleatum, could translocate via the blood stream and accumulate in breast tumors, progressively increasing with stages of breast cancer [26]. F. nucleatum has been shown to promote breast tumor growth and metastatic progression, possibly by preventing the accumulation of tumor-infiltrating T cells in the tumor microenvironment and colonizing breast tumors through D-galactose-β(1–3)-N-acetyl -D-galactosamine (Gal -GalNAc) which binds to Fap2, a surface lectin from F. nucleatum, involved in the colonization of breast cancer. Furthermore, antibiotic therapy with metronidazole suppresses F. nucleatum-induced breast tumor aggravation, indicating that targeting F. nucleatum may enhance breast cancer treatment [27]. In another detailed study, Parida et al. reported that toxin-producing strains of Bacteroides fragilis, when present in the gut or breast tissue, could increase the aggressiveness of breast cancers, induce self-renewal in breast cancer cells and initiate metastatic dissemination to distant organs [10].
The oral administration of Lactobacillus acidophilus results in anti-cancer activity in mice bearing breast tumors, via stimulating the Th1 response and enhancing cellular immunity [28]. Another study showed that Lactobacillus helveticus R389 increased IL-10 and decreased IL-6 levels in serum and mammary cells, thereby suppressing mammary tumor cells by activating the local immune response [29]. Oral administration of Lactobacillus casei significantly increased the production of IL-12 and IFN-γ, thereby improving the immune response in mice with invasive ductal carcinoma [30]. In addition, a population-based case–control study showed that the long-term exposure to probiotics, such as Lactobacillus casei Shirota and soy isoflavones, protected against breast cancer in Japanese women [31].
Improved imaging techniques, such as fluorescent in situ hybridization and modified PCR protocols, have allowed for the visualization of the spatial organization of microbial riders within the tumor, offering a sneak peek into their potential functions in shaping the tumor microenvironment. In a multicenter study of 1526 tumors and their adjacent normal tissues, Nejman et al. examined nine tumor types, including those in the breast, lung, ovary, pancreas, melanoma, bone, and brain. They demonstrated that tumor-specific microbes resided within tumors, as well as immune cells, in a cell-wall-deficient intracellular state [32]. In addition, breast tumors were found to be the richest and most biodiverse among the nine tumor types examined. In another seminal study, Cai et al. proposed that the internal tumor environment enhanced the metastatic dissemination of breast cancers. The intra-tumor bacteria induce cytoskeletal remodeling in circulating breast cancer cells, making them more resistant to the fluid sheer stress in the circulation, thereby helping them establish colonies in distant sites [33].
4. Gut Microbiome and Breast Cancer
A healthy human gut harbors between 300 and 500 bacterial species, predominantly composed of members belonging to four phyla, Actinobacteria, Bacteroidetes, Proteobacteria and Firmicutes [34]. The most important physiological functions, energy assimilation, immune regulation, and xenobiotic metabolism, take place in the gut, and are largely accomplished by the gut microbes [35]. In the context of breast cancer, the gut microbiota plays a complex yet crucial role. In addition to producing pro-carcinogenic toxins, such as BFT from B. fragilis and colibactin from pks+ E. coli, which can potentially reach the breast tissue via circulation, gut microbes produce metabolites such as cadaverine [36], indoxusulfate [37], and lithocolic acid [38], which are touted to hinder breast cancer progression.
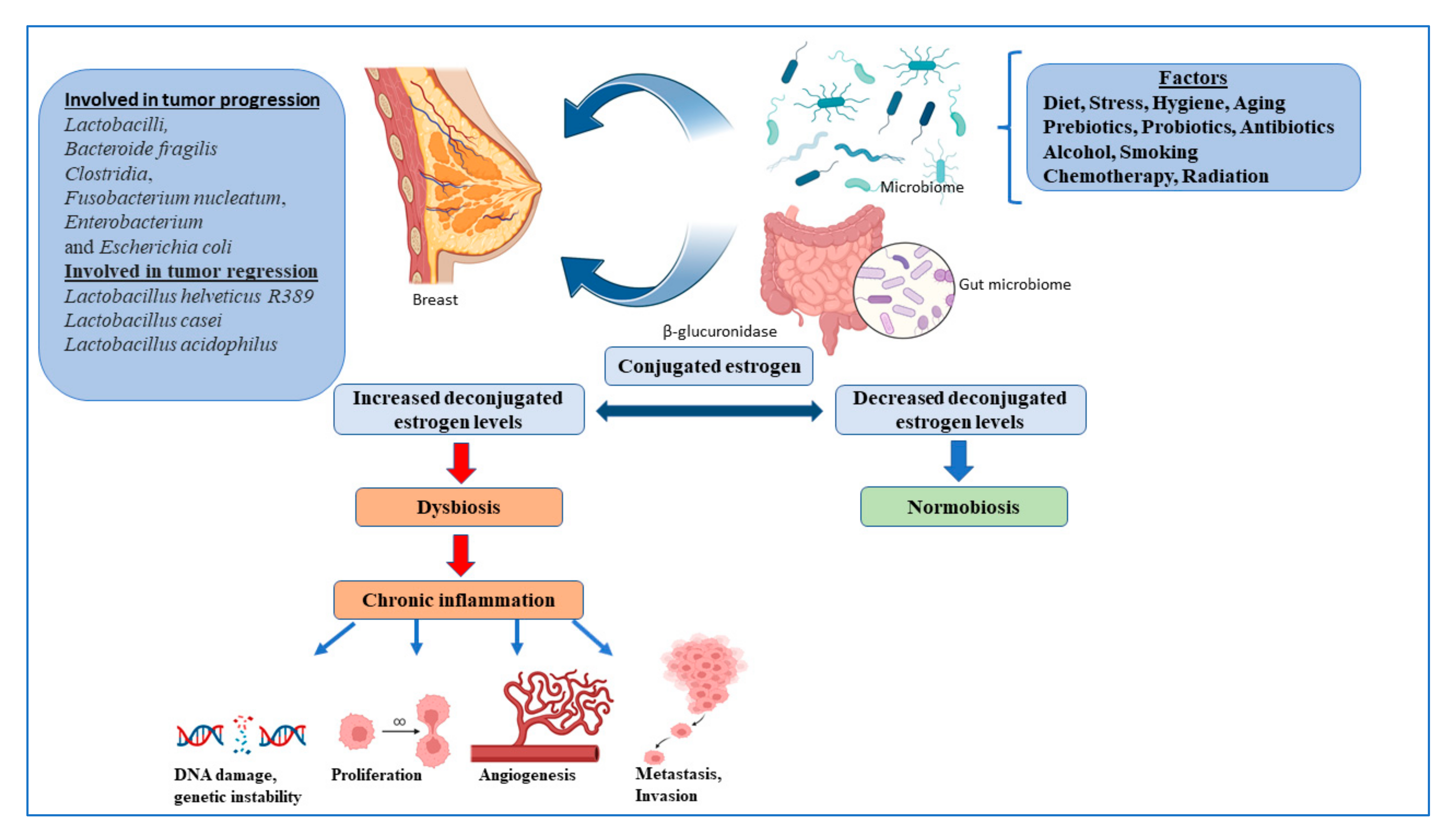
Multiple strains of the gut microbes are known to synthesize enzymes that deconjugate conjugated estrogen metabolites, preventing their excretion, and thereby regulating the levels of active estrogens in the circulation, one of the major promoters of breast cancer [39]. Many bacterial species are also known to synthesize estrogen mimics e.g., seasmin, eterolactone and enterodiol, by breaking down dietary lignans [39]. Gut microbial beta glucuronidases convert conjugated estrogen to deconjugated estrogen, which regulates breast dysbiosis, and leads to chronic inflammation, resulting in the alteration of the DNA breaks, proliferation, angiogenesis, metastasis, and invasion (Figure 1).
Gut bacteria act through pathogen-associated molecular patterns (PAMPs), which regulate Toll-like receptors (TLRs) that are responsible for host defense against invading pathogens, and that activate signaling pathways that lead to the induction of immune and inflammatory genes. PAMPs are also responsible for inducing T cells, B cells and CD4 T cells to differentiate into Treg and Th17 cells, which return to the gut or enter the systemic circulation, which can affect immunity at different levels [40]. The gut microbiota supports digestion, metabolism, and host immune responses, resulting in a symbiotic relationship between the host and microbiota, called the normobiosis, that maintains homeostasis [41]. Dysbiosis is caused by changes in the microbiome, leading to a decrease in microbial diversity. As a result, the inability of the microbiota to defend against pathogenic organisms ultimately leads to local and systemic diseases [41]. Obesity, an important breast cancer risk factor, is also closely associated with gut dysbiosis. Multiple studies, to date, have shown significant differences between the gut microbiota of healthy women compared to women with breast cancer, with some showing an overlap with obese microbiota [23,42]. Finally, multiple studies have shown that a healthy gut microbiota is indispensable for the effective utilization of drugs, chemotherapeutics, immunotherapy, and even radiotherapy [40,43].
5. Gut Microbiome and Hormone Therapy
Hormone treatment, commonly known as endocrine therapy, is used to treat hormone-sensitive breast cancer. There has been minimal research investigating the relationship between hormone therapy and the gut microbiome in breast cancer. The long-term effects on the microbiome must be determined. Studies reported that some types of breast cancer are hormone-dependent on estrogen and progesterone [44,45,46], but, given the significance of the gut microbiota in estrogen metabolism, this may be of relevance. The gut microbiota varies by race, ethnicity, nutrition, BMI, exposure to antibiotics, and the presence of infections, and it is a major factor in the development of breast cancer [39]. The gut microbiome plays a key role in regulating estrogens, through secretions of β-glucuronidase, an enzyme that deconjugates estrogens into their active forms [39]. The collection of bacteria in the gut responsible for metabolizing and modulating the system’s circulating estrogen comprises an estrobolome. Circulating estrogen levels decline when this process is hindered by gut microbiota dysbiosis, which is defined as a decrease in microbial diversity. Many studies linking dysbiosis of the gut microbiota to many kinds of cancer have been conducted in recent years [47]. Additionally, the circulating estrogens may contribute to pre-menstrual syndrome, obesity, metabolic syndrome, endometriosis, polycystic ovary syndrome (PCOS), infertility, and cardiovascular disease (CVD), due to estrogen dominance.
The consumption of soy products is a major source of isoflavones, which contain phytoestrogens that have been hypothesized to reduce breast cancer risk. Yamamoto S et al. showed that the consumption of miso soup and isoflavones was inversely associated with the risk of breast cancer, in a population-based prospective cohort study in Japan [48]. However, another population-based prospective cohort study suggested that soy and isoflavone intakes have a protective effect on postmenopausal breast cancer in Japan [49]. A probiotic drink containing Lactobacillus casei Shirota was found to be inversely associated with breast cancer incidence with the consumption of soy isoflavones [31]. The breast cancer prevention mechanism of soy may be attributed to the estrogenic and antiestrogenic effects of soy isoflavones, such as genistein and daidzein. Lactobacillus is a genus of Gram-positive bacteria with the probiotic ability to reduce the incidence of estrogen receptor-positive (ER+) breast cancers, by increasing the anti-cancer activity of tamoxifen and other endocrine system-targeting drugs [50].
6. Microbiome and Cancer Immunotherapy
The microbiome has a significant impact on local and systemic host immunity. The microbiota can control the host’s immune system and mediate inflammation, which plays an important role in cancer development. Both innate and adaptive immune responses are stimulated by the microbiota. Cancer immunotherapy has proven to be a promising approach in the treatment of cancer patients. Several studies have demonstrated that the microbiome influences the effectiveness of cancer immunotherapies, especially immune checkpoint inhibitors (ICIs) and CTLA-4 [51]. Microbiome toxicity in response to immune blockades has been studied in animal models. Vetizou M. et al. demonstrated that the anti-tumor efficacy of CTLA blockade depends on the presence of different Bacteroides species. T-cell-specific responses to B. thetaiotaomicron or B. fragilis correlate with the efficacy of CTLA blockade in mice and patients [13]. Other studies have shown that Bifidobacterium spp., Ruminococcaceae and Faecalibacterium, found in the gut microbiota, can influence the efficacy of anti-PD-1 immunotherapy in melanoma patients [52,53,54,55]. Supplementation with specific strains of Bifidobacterium breve enhances lymphocyte-mediated anti-cancer immunity, thereby inducing efficacy in MC38 colon cancer mice [56].
Patients with metastatic melanoma showed a relative abundance of Lactobacillales in the oral microbiome and Bacteroidetes in the fecal microbiome [52]. Patients with a high diversity and abundance of Ruminococcaceae/Faecalibacterium have enhanced systemic and anti-tumor immune responses, mediated by increased antigen presentation, as well as improved effector T cell function in the tumor microenvironment [52]. Patients with a higher frequency of Bacteroidetes in the gut microbiome had higher levels of regulatory T cells (Tregs) and myeloid-derived suppressor cells (MDSCs) in the systemic circulation, and impaired cytokine responses. The oral administration of Bifidobacteria alone improved tumor control with PD-L1-specific antibody treatment, and combination therapy almost suppressed tumor growth, which may influence the therapeutic response to anti-PD-1 at the level of the tumor microenvironment [54]. Patients with epithelial cancer who were not treated with antibiotics had significantly better overall survival and progression-free survival than those who were treated with antibiotics containing anti-PD-1/PD-L1 [12]. Some challenges to improving the gut microbiome in immunotherapy have been noted, including the presence of unfavorable gut bacteria that may affect the efficacy of immunotherapy, optimal fecal microbial transplantation (FMT) donor selection, and other factors, such as diet, sleep habits, exercise, and medications [51].
An innovative adoptive cell treatment, known as chimeric antigen receptor (CAR) cell therapy, has the potential to alter and instruct immune cells to target certain tumor cells. These CAR-T or CAR-NK cells have a synthetic receptor that is specific to tumor cells expressed on their surface. CARs are divided into four main components, based on their structure and function, including an extracellular ligand-binding domain, most commonly a single-chain variable fragment (scFv), a hinge domain, a transmembrane domain, and an intracellular signaling domain. Over the years, several studies have demonstrated promising therapeutic targets for CAR cell therapy in breast cancer [40,57,58]. Meili Sun et al. demonstrated a novel CAR-T cell therapy for HER2-positive breast and ovarian cancer cells [59]. In another study, Priceman J.S et al. showed that either the intravenous or intraventricular administration of HER2-CAR T cells decreases anti-tumor activity in an orthotropic xenograft model of breast cancer [60]. HER2-specific mouse CAR-T cells increase anti-tumor activity against HER2-positive, transtuzumab-resistant tumor cells in vitro and in vivo [57,58]. A recent study showed that EGFR-CAR-T cell treatment induced a set of immunosuppressive genes through IFN-γ signaling in triple-negative breast cancer cells [61,62]. Zhiwei Hu showed that tissue factor (TF)-CAR-NK enhanced the treatment of TNBC in a xenograft mouse model [63].
7. Microbiota Role in Drug Resistance
Chemoresistance is one of the major causes of breast cancer deaths [64]. The microbiota has been shown to play a role in chemoresistance. Chemotherapy and radiation play an integral role in the treatment of almost all types of cancer. Shio et al. reported that the bacterial and fungal microbiota differentially regulate tumor responses to radiation therapy in mouse models of breast cancer. Targeting commensal fungi enhanced the response to radiation therapy and reduced the expression of the C-type lectin receptor Dectin-1, a key innate immune receptor for sensing fungi that contributes to survival in breast cancer [65]. Loss of Dectin-1 abrogated the effect of antifungal agents on radiation therapy. Depleting the bacteria significantly reduces the effect of radiation on tumor growth by curtailing the anti-tumor immune response. Commensal bacteria are required for efficient anti-tumor immune responses, while commensal fungi regulate the immunosuppressive microenvironment following treatment [65]. Several bacterial subsets, including those from the phylum Bacteroidetes and the genera Bifidobacterium and Akkermansia, have been implicated in regulating the anti-tumor immunity induced by oxaliplatin in colon cancer, and by cyclophosphamide in in vivo fibrosarcoma models [66,67]. Furthermore, E. coli was shown to regulate the cytotoxicity of gemcitabine, doxorubicin, etoposide phosphate, etc., in the cancer cell lines [68].
The microbiome particularly affects drug metabolism. Irinotecan (CPT-11), a topoisomerase 1 inhibitor, in combination with fluorouracil and leucovorin, is one of three first-line treatments for metastatic colorectal cancer. FDA approved the use of liposomal irinotecan, as well as 5-fluorouracil and leucovorin, for the treatment of patients with metastatic pancreatic cancer following previous gemcitabine treatment. It is being actively investigated for its use various cancers, including breast cancer [69,70]. Irinotecan is administered to patients intravenously and converted to its active form (SN-38) by carboxylesterases in the liver. SN-38 is inactivated by UDP-glucuronosyltransferases, creating the glucuronidated form (SN-38G), where it enters the intestine via biliary excretion. SN-38G could be reactivated by microbial β-glucuronidases in the gut, which recognize the glucuronidated drug as a carbon source. As a result, adverse drug responses, such as diarrhea, could occur in patients [71]. Breast cancer drugs were shown to regulate microbiota. HER2+ breast cancer patients treated with neoadjuvant trastuzumab (targets HER2) achieved a pathological complete response (pCR), and were characterized by a higher abundance of Clostridiales bacteria and a lower abundance of Bacteroidales. The aromatase inhibitor letrozole, used in treating hormone-positive cancer, resulted in a decrease in the number of Bacteroidales operational taxonomic units (OTUs) and an increase in a majority of Firmicutes OTUs [72,73]. Taken altogether, the potential of microbiota in terms of chemotherapy and radiation needs to be further explored in breast cancer.
8. Clinical Trials-Microbiota and Breast Cancer
Currently, several clinical studies are investigating the effect of the microbiota in relation to breast cancer treatment and the quality of life of breast cancer patients (Table 1). However, conclusive clinical trials are needed to demonstrate the benefits of targeting the microbiota in breast cancer.
9. Microbiota as a Potential Biomarker in Breast Cancer
The analysis of fecal samples in a pilot breast cancer study revealed that postmenopausal women with newly diagnosed breast cancer had a fecal microbiota that was less diverse and compositionally different when compared with similar women without breast cancer [74]. A comparison of microbiota from 70 women who had breast cancer to healthy individuals revealed that breast cancer patients had a higher relative abundance of Bacillus, Enterobacteriaceae and Staphylococcus when compared to health individuals. Further, Escherichia coli and Staphylococcus epidermidis, isolated from breast cancer patients, induced DNA double-stranded breaks in HeLa cells [75]. A comparison of 48 postmenopausal breast cancer cases (75% stage 0–I, 88% estrogen-receptor positive) to 48 contemporaneous, postmenopausal, normal-mammogram, age-matched controls revealed that breast cancer cases had significant estrogen-independent associations with the IgA-positive and IgA-negative gut microbiota [76]. An evaluation of 50 ER/PR+, 34 HER2+, 24 ER/PR/HER+, 40 TNBC and 20 heathy breast cancer tissues showed a unique viral, bacterial, fungal, and parasitic signature between the sub-types [77]. Similarly, other studies have shown different microbiota patterns in breast cancer patients [43,78].
10. Conclusions
Breast cancer is a multifactorial disease that can be affected by age, lifestyle, parity, exposure to carcinogens and genetic factors. However, 70% of breast cancer cases were found to have no known risk factors, other than being female and over 50 years of age. Accumulating evidence points to a new role for the local immune microbiome in breast cancer. Large-scale studies, including animal models, retrospective and prospective studies, and clinical studies, should be designed to determine the role of the microbiota in breast cancer. This could lead to the identification of anti-tumor microbiomes for the treatment or prevention of breast cancer. All of these investigations, confirming the presence of the microbiota in breast tissue, concentrated on distinct cohorts that included healthy controls vs. breast cancer survivors, benign versus malignant illness, and normal breast versus breast cancer.
Author Contributions
Conceptualization and supervision: N.M.; Writing-Original draft preparation: S.V. and S.P.; writing-review editing: B.T.L., R.K. and D.R.P. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Acknowledgments
All figures have been constructed using BioRender (https://.biorender.com).
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| BFT | Bacteroides fragilis toxin |
| HBV | Hepatitis B virus |
| BCV | Hepatitis C virus |
| ER | Estrogen receptor |
| PR | progesterone receptor |
| TNBC | Triple negative breast cancer |
| HMP | Human microbiome project |
| iHMP | Integrated Human microbiome project |
| IDC | Invasive ductal carcinoma |
| ILC | Invasive ductal carcinoma |
| PAMPs | Pathogen-associated molecular patterns |
| ICIS | Immune checkpoint inhibitors |
| T-regs | Regulatory T cells |
| FMT | Fecal microbial transplantation |
| pCR | Pathological complete response |
| OTUs | Operational Taxonomic Units |
| PCOS | Polycystic ovary syndrome |
| CVD | Cardiovascular disease |
References
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer statistics, 2022. CA Cancer J. Clin. 2022, 72, 7–33. [Google Scholar] [CrossRef] [PubMed]
- Turashvili, G.; Brogi, E. Tumor Heterogeneity in Breast Cancer. Front. Med. 2017, 4, 227. [Google Scholar] [CrossRef] [Green Version]
- Lacey, J.V., Jr.; Kreimer, A.R.; Buys, S.S.; Marcus, P.M.; Chang, S.C.; Leitzmann, M.F.; Hoover, R.N.; Prorok, P.C.; Berg, C.D.; Hartge, P.; et al. Breast cancer epidemiology according to recognized breast cancer risk factors in the Prostate, Lung, Colorectal and Ovarian (PLCO) Cancer Screening Trial Cohort. BMC Cancer 2009, 9, 84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morales-Sanchez, A.; Fuentes-Panana, E.M. Human viruses and cancer. Viruses 2014, 6, 4047–4079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sender, R.; Fuchs, S.; Milo, R. Revised Estimates for the Number of Human and Bacteria Cells in the Body. PLoS Biol. 2016, 14, e1002533. [Google Scholar] [CrossRef] [Green Version]
- Gilbert, J.A.; Blaser, M.J.; Caporaso, J.G.; Jansson, J.K.; Lynch, S.V.; Knight, R. Current understanding of the human microbiome. Nat. Med. 2018, 24, 392–400. [Google Scholar] [CrossRef] [PubMed]
- Lynch, S.V.; Pedersen, O. The Human Intestinal Microbiome in Health and Disease. N. Engl. J. Med. 2016, 375, 2369–2379. [Google Scholar] [CrossRef] [Green Version]
- Allen, J.; Sears, C.L. Impact of the gut microbiome on the genome and epigenome of colon epithelial cells: Contributions to colorectal cancer development. Genome Med. 2019, 11, 11. [Google Scholar] [CrossRef] [Green Version]
- Kadosh, E.; Snir-Alkalay, I.; Venkatachalam, A.; May, S.; Lasry, A.; Elyada, E.; Zinger, A.; Shaham, M.; Vaalani, G.; Mernberger, M.; et al. The gut microbiome switches mutant p53 from tumour-suppressive to oncogenic. Nature 2020, 586, 133–138. [Google Scholar] [CrossRef]
- Parida, S.; Wu, S.; Siddharth, S.; Wang, G.; Muniraj, N.; Nagalingam, A.; Hum, C.; Mistriotis, P.; Hao, H.; Talbot, C.C., Jr.; et al. A Procarcinogenic Colon Microbe Promotes Breast Tumorigenesis and Metastatic Progression and Concomitantly Activates Notch and beta-Catenin Axes. Cancer Discov. 2021, 11, 1138–1157. [Google Scholar] [CrossRef]
- Hwang, S.; Yi, H.C.; Hwang, S.; Jo, M.; Rhee, K.J. Dietary Salt Administration Decreases Enterotoxigenic Bacteroides fragilis (ETBF)-Promoted Tumorigenesis via Inhibition of Colonic Inflammation. Int. J. Mol. Sci. 2020, 21, 8034. [Google Scholar] [CrossRef]
- Routy, B.; Le Chatelier, E.; Derosa, L.; Duong, C.P.M.; Alou, M.T.; Daillere, R.; Fluckiger, A.; Messaoudene, M.; Rauber, C.; Roberti, M.P.; et al. Gut microbiome influences efficacy of PD-1-based immunotherapy against epithelial tumors. Science 2018, 359, 91–97. [Google Scholar] [CrossRef] [Green Version]
- Vetizou, M.; Pitt, J.M.; Daillere, R.; Lepage, P.; Waldschmitt, N.; Flament, C.; Rusakiewicz, S.; Routy, B.; Roberti, M.P.; Duong, C.P.; et al. Anticancer immunotherapy by CTLA-4 blockade relies on the gut microbiota. Science 2015, 350, 1079–1084. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, P.; Tian, Z.; Kong, X.; Yang, L.; Shan, X.; Dong, B.; Ding, X.; Jing, X.; Jiang, C.; Jiang, N.; et al. FadA promotes DNA damage and progression of Fusobacterium nucleatum-induced colorectal cancer through up-regulation of chk2. J. Exp. Clin. Cancer Res. 2020, 39, 202. [Google Scholar] [CrossRef]
- Mellemgaard, A.; Gaarslev, K. Risk of hepatobiliary cancer in carriers of Salmonella typhi. J. Natl. Cancer Inst. 1988, 80, 288. [Google Scholar] [CrossRef] [PubMed]
- Oliero, M.; Hajjar, R.; Cuisiniere, T.; Fragoso, G.; Calve, A.; Dagbert, F.; Loungnarath, R.; Sebajang, H.; Schwenter, F.; Wassef, R.; et al. Prevalence of pks + bacteria and enterotoxigenic Bacteroides fragilis in patients with colorectal cancer. Gut Pathog. 2022, 14, 51. [Google Scholar] [CrossRef]
- Grivennikov, S.I. IL-11: A prominent pro-tumorigenic member of the IL-6 family. Cancer Cell 2013, 24, 145–147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fulbright, L.E.; Ellermann, M.; Arthur, J.C. The microbiome and the hallmarks of cancer. PLoS Pathog. 2017, 13, e1006480. [Google Scholar] [CrossRef]
- De Martel, C.; Georges, D.; Bray, F.; Ferlay, J.; Clifford, G.M. Global burden of cancer attributable to infections in 2018: A worldwide incidence analysis. Lancet Glob. Health 2020, 8, e180–e190. [Google Scholar] [CrossRef] [Green Version]
- Giaquinto, A.N.; Sung, H.; Miller, K.D.; Kramer, J.L.; Newman, L.A.; Minihan, A.; Jemal, A.; Siegel, R.L. Breast Cancer Statistics, 2022. CA Cancer J. Clin. 2022, 72, 524–541. [Google Scholar] [CrossRef]
- The Integrative HMP (iHMP) Research Network Consortium. The Integrative Human Microbiome Project. Nature 2019, 569, 641–648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Urbaniak, C.; Cummins, J.; Brackstone, M.; Macklaim, J.M.; Gloor, G.B.; Baban, C.K.; Scott, L.; O’Hanlon, D.M.; Burton, J.P.; Francis, K.P.; et al. Microbiota of human breast tissue. Appl. Environ. Microbiol. 2014, 80, 3007–3014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parida, S.; Sharma, D. The power of small changes: Comprehensive analyses of microbial dysbiosis in breast cancer. Biochim. Biophys. Acta Rev. Cancer 2019, 1871, 392–405. [Google Scholar] [CrossRef] [PubMed]
- Smith, A.; Pierre, J.F.; Makowski, L.; Tolley, E.; Lyn-Cook, B.; Lu, L.; Vidal, G.; Starlard-Davenport, A. Distinct microbial communities that differ by race, stage, or breast-tumor subtype in breast tissues of non-Hispanic Black and non-Hispanic White women. Sci. Rep. 2019, 9, 11940. [Google Scholar] [CrossRef] [Green Version]
- Tzeng, A.; Sangwan, N.; Jia, M.; Liu, C.C.; Keslar, K.S.; Downs-Kelly, E.; Fairchild, R.L.; Al-Hilli, Z.; Grobmyer, S.R.; Eng, C. Human breast microbiome correlates with prognostic features and immunological signatures in breast cancer. Genome Med. 2021, 13, 60. [Google Scholar] [CrossRef]
- Parhi, L.; Alon-Maimon, T.; Sol, A.; Nejman, D.; Shhadeh, A.; Fainsod-Levi, T.; Yajuk, O.; Isaacson, B.; Abed, J.; Maalouf, N.; et al. Breast cancer colonization by Fusobacterium nucleatum accelerates tumor growth and metastatic progression. Nat. Commun. 2020, 11, 3259. [Google Scholar] [CrossRef]
- He, Z.; Tian, W.; Wei, Q.; Xu, J. Involvement of Fusobacterium nucleatum in malignancies except for colorectal cancer: A literature review. Front. Immunol. 2022, 13, 968649. [Google Scholar] [CrossRef]
- Yazdi, M.H.; Soltan Dallal, M.M.; Hassan, Z.M.; Holakuyee, M.; Agha Amiri, S.; Abolhassani, M.; Mahdavi, M. Oral administration of Lactobacillus acidophilus induces IL-12 production in spleen cell culture of BALB/c mice bearing transplanted breast tumour. Br. J. Nutr. 2010, 104, 227–232. [Google Scholar] [CrossRef] [Green Version]
- De Moreno de LeBlanc, A.; Matar, C.; Theriault, C.; Perdigon, G. Effects of milk fermented by Lactobacillus helveticus R389 on immune cells associated to mammary glands in normal and a breast cancer model. Immunobiology 2005, 210, 349–358. [Google Scholar] [CrossRef]
- Soltan Dallal, M.M.; Yazdi, M.H.; Holakuyee, M.; Hassan, Z.M.; Abolhassani, M.; Mahdavi, M. Lactobacillus casei ssp.casei induced Th1 cytokine profile and natural killer cells activity in invasive ductal carcinoma bearing mice. Iran. J. Allergy Asthma Immunol. 2012, 11, 183–189. [Google Scholar]
- Toi, M.; Hirota, S.; Tomotaki, A.; Sato, N.; Hozumi, Y.; Anan, K.; Nagashima, T.; Tokuda, Y.; Masuda, N.; Ohsumi, S.; et al. Probiotic Beverage with Soy Isoflavone Consumption for Breast Cancer Prevention: A Case-control Study. Curr. Nutr. Food Sci. 2013, 9, 194–200. [Google Scholar] [CrossRef] [PubMed]
- Nejman, D.; Livyatan, I.; Fuks, G.; Gavert, N.; Zwang, Y.; Geller, L.T.; Rotter-Maskowitz, A.; Weiser, R.; Mallel, G.; Gigi, E.; et al. The human tumor microbiome is composed of tumor type-specific intracellular bacteria. Science 2020, 368, 973–980. [Google Scholar] [CrossRef] [PubMed]
- Fu, A.; Yao, B.; Dong, T.; Chen, Y.; Yao, J.; Liu, Y.; Li, H.; Bai, H.; Liu, X.; Zhang, Y.; et al. Tumor-resident intracellular microbiota promotes metastatic colonization in breast cancer. Cell 2022, 185, 1356–1372.e1326. [Google Scholar] [CrossRef]
- Bodai, B.I.; Nakata, T.E. Breast Cancer: Lifestyle, the Human Gut Microbiota/Microbiome, and Survivorship. Perm. J. 2020, 24. [Google Scholar] [CrossRef]
- Parida, S.; Sharma, D. Microbial Alterations and Risk Factors of Breast Cancer: Connections and Mechanistic Insights. Cells 2020, 9, 1091. [Google Scholar] [CrossRef] [PubMed]
- Kovacs, T.; Miko, E.; Vida, A.; Sebo, E.; Toth, J.; Csonka, T.; Boratko, A.; Ujlaki, G.; Lente, G.; Kovacs, P.; et al. Cadaverine, a metabolite of the microbiome, reduces breast cancer aggressiveness through trace amino acid receptors. Sci. Rep. 2019, 9, 1300. [Google Scholar] [CrossRef] [Green Version]
- Sari, Z.; Miko, E.; Kovacs, T.; Boratko, A.; Ujlaki, G.; Janko, L.; Kiss, B.; Uray, K.; Bai, P. Indoxylsulfate, a Metabolite of the Microbiome, Has Cytostatic Effects in Breast Cancer via Activation of AHR and PXR Receptors and Induction of Oxidative Stress. Cancers 2020, 12, 2915. [Google Scholar] [CrossRef]
- Kovacs, P.; Csonka, T.; Kovacs, T.; Sari, Z.; Ujlaki, G.; Sipos, A.; Karanyi, Z.; Szeocs, D.; Hegedus, C.; Uray, K.; et al. Lithocholic Acid, a Metabolite of the Microbiome, Increases Oxidative Stress in Breast Cancer. Cancers 2019, 11, 1255. [Google Scholar] [CrossRef] [Green Version]
- Parida, S.; Sharma, D. The Microbiome-Estrogen Connection and Breast Cancer Risk. Cells 2019, 8, 1642. [Google Scholar] [CrossRef] [Green Version]
- Vitorino, M.; Baptista de Almeida, S.; Alpuim Costa, D.; Faria, A.; Calhau, C.; Azambuja Braga, S. Human Microbiota and Immunotherapy in Breast Cancer—A Review of Recent Developments. Front. Oncol. 2021, 11, 815772. [Google Scholar] [CrossRef]
- Rea, D.; Coppola, G.; Palma, G.; Barbieri, A.; Luciano, A.; Del Prete, P.; Rossetti, S.; Berretta, M.; Facchini, G.; Perdona, S.; et al. Microbiota effects on cancer: From risks to therapies. Oncotarget 2018, 9, 17915–17927. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geng, J.; Ni, Q.; Sun, W.; Li, L.; Feng, X. The links between gut microbiota and obesity and obesity related diseases. Biomed. Pharmacother. 2022, 147, 112678. [Google Scholar] [CrossRef] [PubMed]
- Eslami, S.Z.; Majidzadeh, A.K.; Halvaei, S.; Babapirali, F.; Esmaeili, R. Microbiome and Breast Cancer: New Role for an Ancient Population. Front. Oncol. 2020, 10, 120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brooks, P.G. The relationship of estrogen and progesterone to breast disease. J. Reprod. Med. 1984, 29, 530–538. [Google Scholar]
- Jerry, D.J. Roles for estrogen and progesterone in breast cancer prevention. Breast Cancer Res. 2007, 9, 102. [Google Scholar] [CrossRef] [Green Version]
- Truin, W.; Roumen, R.M.H.; Siesling, S.; van de Vijver, K.K.; Tjan-Heijnen, V.C.G.; Voogd, A.C. Estrogen and progesterone receptor expression levels do not differ between lobular and ductal carcinoma in patients with hormone receptor-positive tumors. Breast Cancer Res. Treat. 2017, 164, 133–138. [Google Scholar] [CrossRef] [Green Version]
- Helmink, B.A.; Khan, M.A.W.; Hermann, A.; Gopalakrishnan, V.; Wargo, J.A. The microbiome, cancer, and cancer therapy. Nat. Med. 2019, 25, 377–388. [Google Scholar] [CrossRef]
- Yamamoto, S.; Sobue, T.; Kobayashi, M.; Sasaki, S.; Tsugane, S.; Japan Public Health Center-Based Prospective Study on Cancer Cardiovascular Diseases (JPHC Study) Group. Soy, isoflavones, and breast cancer risk in Japan. J. Natl. Cancer Inst. 2003, 95, 906–913. [Google Scholar] [CrossRef] [Green Version]
- Wada, K.; Nakamura, K.; Tamai, Y.; Tsuji, M.; Kawachi, T.; Hori, A.; Takeyama, N.; Tanabashi, S.; Matsushita, S.; Tokimitsu, N.; et al. Soy isoflavone intake and breast cancer risk in Japan: From the Takayama study. Int. J. Cancer 2013, 133, 952–960. [Google Scholar] [CrossRef]
- Katherine, L.C. Probiotic Bacteria May Enhance Tamoxifen Effectiveness in Treatment of ER+ Breast Cancer; Press Release; Endocrine Society: Atlanta, GA, USA, 2022. [Google Scholar]
- Li, W.; Deng, Y.; Chu, Q.; Zhang, P. Gut microbiome and cancer immunotherapy. Cancer Lett. 2019, 447, 41–47. [Google Scholar] [CrossRef]
- Gopalakrishnan, V.; Spencer, C.N.; Nezi, L.; Reuben, A.; Andrews, M.C.; Karpinets, T.V.; Prieto, P.A.; Vicente, D.; Hoffman, K.; Wei, S.C.; et al. Gut microbiome modulates response to anti-PD-1 immunotherapy in melanoma patients. Science 2018, 359, 97–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Killock, D. Immunotherapy: Gut bacteria modulate responses to PD-1 blockade. Nat. Rev. Clin. Oncol. 2018, 15, 6–7. [Google Scholar] [CrossRef]
- Sivan, A.; Corrales, L.; Hubert, N.; Williams, J.B.; Aquino-Michaels, K.; Earley, Z.M.; Benyamin, F.W.; Lei, Y.M.; Jabri, B.; Alegre, M.L.; et al. Commensal Bifidobacterium promotes antitumor immunity and facilitates anti-PD-L1 efficacy. Science 2015, 350, 1084–1089. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matson, V.; Fessler, J.; Bao, R.; Chongsuwat, T.; Zha, Y.; Alegre, M.L.; Luke, J.J.; Gajewski, T.F. The commensal microbiome is associated with anti-PD-1 efficacy in metastatic melanoma patients. Science 2018, 359, 104–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoon, Y.; Kim, G.; Jeon, B.N.; Fang, S.; Park, H. Bifidobacterium Strain-Specific Enhances the Efficacy of Cancer Therapeutics in Tumor-Bearing Mice. Cancers 2021, 13, 957. [Google Scholar] [CrossRef] [PubMed]
- Toth, G.; Szollosi, J.; Abken, H.; Vereb, G.; Szoor, A. A Small Number of HER2 Redirected CAR T Cells Significantly Improves Immune Response of Adoptively Transferred Mouse Lymphocytes against Human Breast Cancer Xenografts. Int. J. Mol. Sci. 2020, 21, 1039. [Google Scholar] [CrossRef] [Green Version]
- Szoor, A.; Toth, G.; Zsebik, B.; Szabo, V.; Eshhar, Z.; Abken, H.; Vereb, G. Trastuzumab derived HER2-specific CARs for the treatment of trastuzumab-resistant breast cancer: CAR T cells penetrate and eradicate tumors that are not accessible to antibodies. Cancer Lett. 2020, 484, 1–8. [Google Scholar] [CrossRef]
- Sun, M.; Shi, H.; Liu, C.; Liu, J.; Liu, X.; Sun, Y. Construction and evaluation of a novel humanized HER2-specific chimeric receptor. Breast Cancer Res. 2014, 16, R61. [Google Scholar] [CrossRef] [Green Version]
- Priceman, S.J.; Tilakawardane, D.; Jeang, B.; Aguilar, B.; Murad, J.P.; Park, A.K.; Chang, W.C.; Ostberg, J.R.; Neman, J.; Jandial, R.; et al. Regional Delivery of Chimeric Antigen Receptor-Engineered T Cells Effectively Targets HER2(+) Breast Cancer Metastasis to the Brain. Clin. Cancer Res. 2018, 24, 95–105. [Google Scholar] [CrossRef] [Green Version]
- Xia, L.; Zheng, Z.; Liu, J.Y.; Chen, Y.J.; Ding, J.; Hu, G.S.; Hu, Y.H.; Liu, S.; Luo, W.X.; Xia, N.S.; et al. Targeting Triple-Negative Breast Cancer with Combination Therapy of EGFR CAR T Cells and CDK7 Inhibition. Cancer Immunol. Res. 2021, 9, 707–722. [Google Scholar] [CrossRef]
- Xia, L.; Zheng, Z.Z.; Liu, J.Y.; Chen, Y.J.; Ding, J.C.; Xia, N.S.; Luo, W.X.; Liu, W. EGFR-targeted CAR-T cells are potent and specific in suppressing triple-negative breast cancer both in vitro and in vivo. Clin. Transl. Immunol. 2020, 9, e01135. [Google Scholar] [CrossRef]
- Hu, Z. Tissue factor as a new target for CAR-NK cell immunotherapy of triple-negative breast cancer. Sci. Rep. 2020, 10, 2815. [Google Scholar] [CrossRef] [Green Version]
- Makena, M.R.; Ranjan, A.; Thirumala, V.; Reddy, A.P. Cancer stem cells: Road to therapeutic resistance and strategies to overcome resistance. Biochim. Biophys. Acta Mol. Basis Dis. 2020, 1866, 165339. [Google Scholar] [CrossRef] [PubMed]
- Shiao, S.L.; Kershaw, K.M.; Limon, J.J.; You, S.; Yoon, J.; Ko, E.Y.; Guarnerio, J.; Potdar, A.A.; McGovern, D.P.B.; Bose, S.; et al. Commensal bacteria and fungi differentially regulate tumor responses to radiation therapy. Cancer Cell 2021, 39, 1202–1213.e1206. [Google Scholar] [CrossRef] [PubMed]
- Daillere, R.; Vetizou, M.; Waldschmitt, N.; Yamazaki, T.; Isnard, C.; Poirier-Colame, V.; Duong, C.P.M.; Flament, C.; Lepage, P.; Roberti, M.P.; et al. Enterococcus hirae and Barnesiella intestinihominis Facilitate Cyclophosphamide-Induced Therapeutic Immunomodulatory Effects. Immunity 2016, 45, 931–943. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iida, N.; Dzutsev, A.; Stewart, C.A.; Smith, L.; Bouladoux, N.; Weingarten, R.A.; Molina, D.A.; Salcedo, R.; Back, T.; Cramer, S.; et al. Commensal bacteria control cancer response to therapy by modulating the tumor microenvironment. Science 2013, 342, 967–970. [Google Scholar] [CrossRef]
- Dieleman, S.; Aarnoutse, R.; Ziemons, J.; Kooreman, L.; Boleij, A.; Smidt, M. Exploring the Potential of Breast Microbiota as Biomarker for Breast Cancer and Therapeutic Response. Am. J. Pathol. 2021, 191, 968–982. [Google Scholar] [CrossRef]
- Bailly, C. Irinotecan: 25 years of cancer treatment. Pharmacol. Res. 2019, 148, 104398. [Google Scholar] [CrossRef]
- Kang, M.H.; Wang, J.; Makena, M.R.; Lee, J.S.; Paz, N.; Hall, C.P.; Song, M.M.; Calderon, R.I.; Cruz, R.E.; Hindle, A.; et al. Activity of MM-398, nanoliposomal irinotecan (nal-IRI), in Ewing’s family tumor xenografts is associated with high exposure of tumor to drug and high SLFN11 expression. Clin. Cancer Res. 2015, 21, 1139–1150. [Google Scholar] [CrossRef] [Green Version]
- Guthrie, L.; Gupta, S.; Daily, J.; Kelly, L. Human microbiome signatures of differential colorectal cancer drug metabolism. NPJ Biofilms Microbiomes 2017, 3, 27. [Google Scholar] [CrossRef] [Green Version]
- Di Modica, M.; Gargari, G.; Regondi, V.; Bonizzi, A.; Arioli, S.; Belmonte, B.; De Cecco, L.; Fasano, E.; Bianchi, F.; Bertolotti, A.; et al. Gut Microbiota Condition the Therapeutic Efficacy of Trastuzumab in HER2-Positive Breast Cancer. Cancer Res. 2021, 81, 2195–2206. [Google Scholar] [CrossRef] [PubMed]
- Kelley, S.T.; Skarra, D.V.; Rivera, A.J.; Thackray, V.G. The Gut Microbiome Is Altered in a Letrozole-Induced Mouse Model of Polycystic Ovary Syndrome. PLoS ONE 2016, 11, e0146509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goedert, J.J.; Jones, G.; Hua, X.; Xu, X.; Yu, G.; Flores, R.; Falk, R.T.; Gail, M.H.; Shi, J.; Ravel, J.; et al. Investigation of the association between the fecal microbiota and breast cancer in postmenopausal women: A population-based case-control pilot study. J. Natl. Cancer Inst. 2015, 107, djv147. [Google Scholar] [CrossRef] [PubMed]
- Urbaniak, C.; Gloor, G.B.; Brackstone, M.; Scott, L.; Tangney, M.; Reid, G. The Microbiota of Breast Tissue and Its Association with Breast Cancer. Appl. Environ. Microbiol. 2016, 82, 5039–5048. [Google Scholar] [CrossRef] [Green Version]
- Goedert, J.J.; Hua, X.; Bielecka, A.; Okayasu, I.; Milne, G.L.; Jones, G.S.; Fujiwara, M.; Sinha, R.; Wan, Y.; Xu, X.; et al. Postmenopausal breast cancer and oestrogen associations with the IgA-coated and IgA-noncoated faecal microbiota. Br. J. Cancer 2018, 118, 471–479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Banerjee, S.; Tian, T.; Wei, Z.; Shih, N.; Feldman, M.D.; Peck, K.N.; DeMichele, A.M.; Alwine, J.C.; Robertson, E.S. Distinct Microbial Signatures Associated With Different Breast Cancer Types. Front. Microbiol. 2018, 9, 951. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Altemus, J.; Niazi, F.; Green, H.; Calhoun, B.C.; Sturgis, C.; Grobmyer, S.R.; Eng, C. Breast tissue, oral and urinary microbiomes in breast cancer. Oncotarget 2017, 8, 88122–88138. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
The microbiome and regulation of estrogen in the breast cancer. Figures created with BioRender.com.
Figure 1.
The microbiome and regulation of estrogen in the breast cancer. Figures created with BioRender.com.


{kind=link}
Table 1.
Ongoing clinical trials targeting the microbiome of breast cancer patients.
| Title | Clinical Trial No. | Study Design | Status |
|---|---|---|---|
| Gut Microbiome Components Predict Response to Neoadjuvant Therapy in HER2-positive Breast Cancer Patients: A Prospective Study | NCT05444647 | Observational model: Cohort | Recruiting |
| Exercise, Gut Microbiome, and Breast Cancer: Increasing Reach to Underserved Populations (EMBRACE) | NCT05000502 | Randomized | Recruiting |
| Assessing the Impact of the Microbiome on Breast Cancer Radiotherapy Toxicity | NCT04245150 | Observational model: Cohort | Recruiting |
| Gut and Intratumoral Microbiome Effect on the Neoadjuvant Chemotherapy-induced Immunosurveillance in Triple Negative Breast Cancer | NCT03586297 | Observational model: Cohort | Recruiting |
| The Association Between Radiation Dermatitis and Skin Microbiome in Breast Cancer Patients | NCT05032768 | Observational model: Cohort | Recruiting |
| Engineering Gut Microbiome to Target Breast Cancer | NCT03358511 | Intervention model: Single group assignment | Completed |
| Evaluating the Association Between Changes in the Gut Microbiome and Chemotherapy-Induced Nausea in Women Receiving Chemotherapy for Stage I-III Breast Cancer | NCT05417867 | Observational model: Case-only | Recruiting |
| Evaluating Mepitel in Post-mastectomy Patients and the Role of the Skin Microbiome in Radiation Dermatitis | NCT03519438 | Observational model: Cohort | Completed |
| Determinants of Acquired Endocrine Resistance in Metastatic Breast Cancer: A Pilot Study (ENDO-RESIST) | NCT04579484 | Observational model: Cohort | Recruiting |
| Oral Aromatase Inhibitors Modify the Gut Microbiome | NCT05030038 | Observational model: Cohort | Recruiting |
| The Breast Cancer Personalized Nutrition Study (BREACPNT) | NCT04079270 | Interventional: Randomized | Recruiting |
| Microbiome and Association With Implant Infections | NCT05020574 | Interventional: Randomized (phase 2) | Recruiting |
| Gut Microbe Composition, Exercise, and Breast Breast Cancer Survivors (ROME) | NCT04088708 | Interventional: Randomized | Recruiting |
| Effect of Radiotherapy Variables on Circulating Effectors of Immune Response and Local Microbiome | NCT03383107 | Observational | Completed |
| Study to Investigate Efficacy of a Novel Probiotic on the Bacteriome and Mycobiome of Breast Cancer | NCT04362826 | Interventional: Randomized | Not yet recruited |
| ARGONAUT: Stool and Blood Sample Bank for Cancer Patients | NCT04638751 | Observational model: Cohort | Recruiting |
| The Gut Microbiome and Immune Checkpoint Inhibitor Therapy in Solid Tumors (PARADIGM) | NCT05037825 | Observational model: Cohort | Recruiting |
| Anti-anxiety Biotics for Breast Cancer Survivors (ABBCS) | NCT04784182 | Interventional: Randomized | Completed |
| Adaptive Nutrition and Exercise Weight Loss (A-NEW) Study (A-NEW) | NCT04499950 | Interventional: non-randomized | Recruiting |
| Effects of Probiotics on the Gut Microbiome and Immune System in Operable Stage I-III Breast or Lung Cancer | NCT04857697 | Interventional | Recruiting |
| Probiotics and Breast Health | NCT03290651 | Interventional | Completed |
| Intestinal Microbiota Impact for Prognosis and Treatment Outcomes in Early Luminal Breast Cancer and Pancreatic Cancer Patients | NCT05580887 | Observational model: Cohort | Recruiting |
| Intestine Bacteria and Breast Cancer Risk | NCT01461070 | Observational model: Case-only | Completed |
| Neoadjuvant Treatment of Locally-advanced Breast Cancer Patients With Ribociclib and Letrozole (NEOLETRIB) | NCT05163106 | Interventional | Recruiting |
| Persistent Post-Surgical Pain in Women With BrCA | NCT02266082 | Observational model: Cohort | Completed |
| Study of Moderate Dose Omega 3 Fatty Acid Supplement in Premenopausal Women at High Risk for Breast Cancer | NCT03383835 | Interventional | Un-Known |
| GRACE-trial: a Randomized Active-controlled Trial for vulvovaginal atrophy in breast Cancer Patients on Endocrine Therapy. (GRACE) | NCT05562518 | Interventional: Randomized | Recruiting |
| Comprehensive Outcomes for After Cancer Health (COACH) | NCT05349227 | Interventional: Randomized | Recruiting |
| Rifaximin for the Treatment of Gastrointestinal Toxicities Related to Pertuzumab-Based Therapy in Patients With Stage I-III HER2 Positive Breast Cancer | NCT04249622 | Interventional: Non-Randomized | Recruiting |
| Impact of Vitamin D Supplementation on the Rate of Pathologic Complete Response in Vitamin D Deficient Patients | NCT04677816 | Interventional: Non-Randomized | Recruiting |
| Weight Loss Plus Omega-3 Fatty Acids or Placebo in High Risk Women | NCT02101970 | Interventional: Randomized | Recruiting |
| Comprehensive Lifestyle Change To Prevent Breast Cancer | NCT03448003 | Interventional: Randomized | Recruiting |
| Avera/Sema4 Oncology and Analytics Protocol (ASAP) | NCT05142033 | Interventional: Randomized | Recruiting |
| Gender Difference in Side effects of Immunotherapy: A Possible Clue to Optimize Cancer Treatment (G-DEFINER) | NCT04435964 | Observational | Recruiting |
| Neoadjuvant Pembrolizumab(Pbr)/Nab-Paclitaxel Followed by Pbr/Epirubicin/Cyclophosphamide in TNBC (NIB) | NCT03289819 | Interventional | Completed |
| Abemaciclib in Treating Patients With Surgically Resectable, Chemotherapy Resistant, Triple Negative Breast Cancer | NCT03979508 | Interventional: Non-Randomized | Recruiting |
The clinical trials information obtained from https://clinicaltrials.gov/.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Viswanathan, S.; Parida, S.; Lingipilli, B.T.; Krishnan, R.; Podipireddy, D.R.; Muniraj, N. Role of Gut Microbiota in Breast Cancer and Drug Resistance. Pathogens 2023, 12, 468. https://doi.org/10.3390/pathogens12030468
AMA Style
Viswanathan S, Parida S, Lingipilli BT, Krishnan R, Podipireddy DR, Muniraj N. Role of Gut Microbiota in Breast Cancer and Drug Resistance. Pathogens. 2023; 12(3):468. https://doi.org/10.3390/pathogens12030468
Chicago/Turabian StyleViswanathan, Sathiyapriya, Sheetal Parida, Bhuvana Teja Lingipilli, Ramalingam Krishnan, Devendra Rao Podipireddy, and Nethaji Muniraj. 2023. "Role of Gut Microbiota in Breast Cancer and Drug Resistance" Pathogens 12, no. 3: 468. https://doi.org/10.3390/pathogens12030468
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.