
White-Toothed Shrews (Genus Crocidura): Potential Reservoirs for Zoonotic Leptospira spp. and Arthropod-Borne Pathogens?
 , , ,
, , ,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Collection and Dissection of Shrews and Hedgehogs
2.2. Nucleic Acid Extraction
2.3. Molecular Species Identification
2.4. Polymerase-Chain-Reaction-Based Screening for Leptospira spp. DNA
2.5. Multilocus Sequence Typing of Leptospira spp.
2.6. Amplification and Sequencing of the secY Gene of Leptospira spp.
2.7. Polymerase-Chain-Reaction-Based Screening for Arthropod-Borne Pathogens, Coxiella burnetii and Brucella spp.
2.8. Statistical Analysis
2.9. Generation of Maps
3. Results
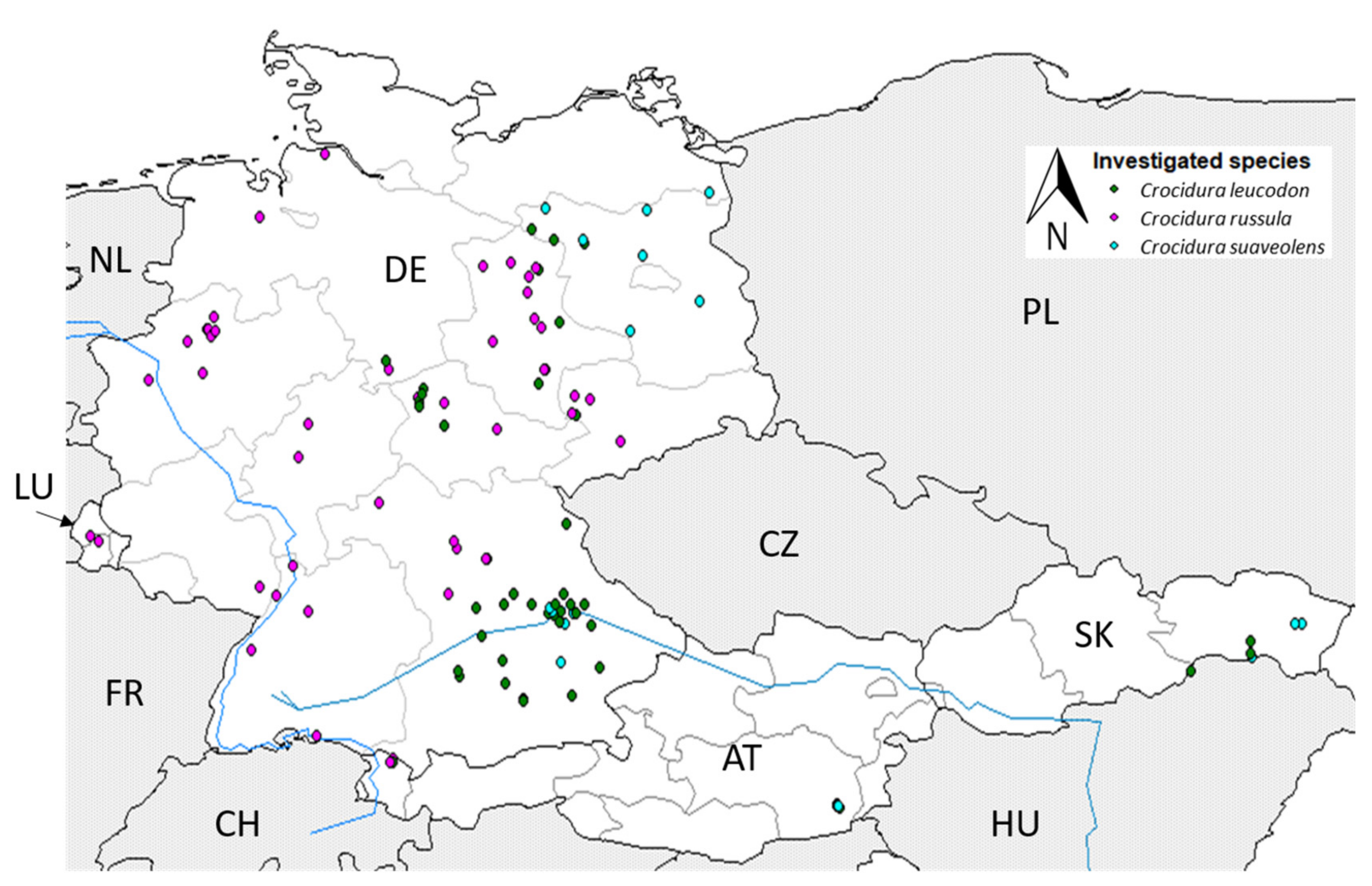
3.1. Distribution of White-Toothed Shrews
3.2. Detection and Sequence Type Identification of Leptospira spp.
3.3. PCR Analysis for Arthropod-Borne Pathogens, Coxiella burnetii and Brucella spp.
4. Discussion
4.1. Current Distribution of White-Toothed Shrews in Germany
4.2. Detection and Characterization of Leptospira spp. in White-Toothed Shrews
4.3. Identification of White-Toothed Shrews as Reservoirs for Arthropod-Borne Pathogens
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wilson, D.E.; Mittermaier, R.A. (Eds.) Handbook of the Mammals of the World: Volume 8. Insectivores, Sloths and Colugos; Lynx Edicions: Barcelona, Spain, 2017; ISBN 9788416728046. [Google Scholar]
- Esselstyn, J.A.; Maharadatunkamsi; Achmadi, A.S.; Siler, C.D.; Evans, B.J. Carving out turf in a biodiversity hotspot: Multiple, previously unrecognized shrew species co-occur on Java Island, Indonesia. Mol. Ecol. 2013, 22, 4972–4987. [Google Scholar] [CrossRef]
- Hutterer, R.; Balete, D.S.; Giarla, T.C.; Heaney, L.R.; Esselstyn, J.A. A new genus and species of shrew (Mammalia: Soricidae) from Palawan Island, Philippines. J. Mammal. 2018, 99, 518–536. [Google Scholar] [CrossRef] [Green Version]
- Esselstyn, J.A.; Achmadi, A.S.; Handika, H.; Swanson, M.T.; Giarla, T.C.; Rowe, K.C. Fourteen New, Endemic Species of Shrew (Genus Crocidura) from Sulawesi Reveal a Spectacular Island Radiation. Bull. Am. Mus. Nat. Hist. 2021, 454, 3–105. [Google Scholar] [CrossRef]
- Dubey, S.; Zaitsev, M.; Cosson, J.-F.; Abdukadier, A.; Vogel, P. Pliocene and Pleistocene diversification and multiple refugia in a Eurasian shrew (Crocidura suaveolens group). Mol. Phylogenet. Evol. 2006, 38, 635–647. [Google Scholar] [CrossRef] [Green Version]
- Dubey, S.; Salamin, N.; Ohdachi, S.D.; Barrière, P.; Vogel, P. Molecular phylogenetics of shrews (Mammalia: Soricidae) reveal timing of transcontinental colonizations. Mol. Phylogenet. Evol. 2007, 44, 126–137. [Google Scholar] [CrossRef] [Green Version]
- Dubey, S.; Salamin, N.; Ruedi, M.; Barrière, P.; Colyn, M.; Vogel, P. Biogeographic origin and radiation of the Old World crocidurine shrews (Mammalia: Soricidae) inferred from mitochondrial and nuclear genes. Mol. Phylogenet. Evol. 2008, 48, 953–963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spitzenberger, F. Die Weißzahnspitzmäuse (Crocidurinae) Österreichs: Mammalia austriaca 8 (Mammalia, Insectivora). Mitt. Abt. Zool. 1985, 35, 1–40. [Google Scholar]
- Kraft, R.; Klemmer, W.; Malec, F. Kleinsäugernachweise aus dem Südlichen Oberpfälzer Wald und angrenzenden Gebieten (Ostbayern). Säugetierkundliche Inf. Jena 2010, 7, 361–372. [Google Scholar]
- Mitchell-Jones, A.G.; Amori, G.; Bogdanowicz, W.; Krystufek, B.; Reijnders, P.J.; Spitzenberger, F.; Stubbe, M.; Thissen, J.B.; Vohralik, V.; Zima, J. The Atlas of European Mammals; T & A D Poyser: London, UK, 1999; ISBN 0856611301. [Google Scholar]
- Cosson, J.-F.; Hutterer, R.; Libois, R.; Sarà, M.; Taberlet, P.; Vogel, P. Phylogeographical footprints of the Strait of Gibraltar and Quaternary climatic fluctuations in the western Mediterranean: A case study with the greater white-toothed shrew, Crocidura russula (Mammalia: Soricidae). Mol. Ecol. 2005, 14, 1151–1162. [Google Scholar] [CrossRef]
- Tosh, D.G.; Lusby, J.; Montgomery, W.I.; O‘Halloran, J. First record of greater white-toothed shrew Crocidura russula in Ireland. Mammal Rev. 2008, 38, 321–326. [Google Scholar] [CrossRef]
- Bond, I.F.; Gilford, E.; McDevitt, A.D.; Young, M.A.; Coomber, F.G. First records of the greater white-toothed shrew Crocidura russula from Great Britain. Mammal Commun. 2022, 8, 23–28. [Google Scholar] [CrossRef]
- Borkenhagen, P. First record of the Greater white-toothed shrew (Crocidura russula) in Schleswig-Holstein, Northern Germany. Faun-Oekol-Mitt 1995, 7, 1–8. [Google Scholar]
- Kraft, R. Ehemalige und aktuelle Verbreitung von Hausspitzmaus, Crocidura russula (Hermann, 1780) und Gartenspitzmaus, Crocidura suaveolens (Pallas, 1811), in Bayern. Bonn.-Zool.-Beitraege 2000, 49, 115–129. [Google Scholar]
- Vogel, P.; Jutzeler, S.; Rulence, B.; Reutter, B.A. Range expansion of the greater white-toothed shrew Crocidura russula in Switzerland results in local extinction of the bicolored white-toothed shrew C. leucodon. Acta Theriol. 2002, 47, 15–24. [Google Scholar] [CrossRef]
- Lehmann, E.v.; Brücher, H. Zum Rückgang der Feld- und der Hausspitzmaus (Crocidura leucodon und russula) in Westeuropa. Bonn.-Zool.-Beitraege 1997, 28, 13–18. [Google Scholar]
- Wolf, R. Bestandsveränderungen und Arealverschiebungen bei den Wimperspitzmäusen (Crocidura Wagler, 1832) zwischen Wurzen und Grimma, Nordwestsachsen. Mitt. Für Sächsische Säugetierfreunde 2010, 2010, 37–44. [Google Scholar]
- Zhang, X.-A.; Li, H.; Jiang, F.-C.; Zhu, F.; Zhang, Y.-F.; Chen, J.-J.; Tan, C.-W.; Anderson, D.E.; Fan, H.; Dong, L.-Y.; et al. A Zoonotic Henipavirus in Febrile Patients in China. N. Engl. J. Med. 2022, 387, 470–472. [Google Scholar] [CrossRef]
- Johne, R.; Tausch, S.H.; Schilling-Loeffler, K.; Ulrich, R.G. Genome sequence analysis of a novel rotavirus strain indicates a broad genetic diversity of rotavirus A in shrews. Infect. Genet. Evol. 2022, 107, 105392. [Google Scholar] [CrossRef]
- Schlegel, M.; Radosa, L.; Rosenfeld, U.M.; Schmidt, S.; Triebenbacher, C.; Löhr, P.-W.; Fuchs, D.; Heroldová, M.; Jánová, E.; Stanko, M.; et al. Broad geographical distribution and high genetic diversity of shrew-borne Seewis hantavirus in Central Europe. Virus Genes 2012, 45, 48–55. [Google Scholar] [CrossRef]
- Radosa, L.; Schlegel, M.; Gebauer, P.; Ansorge, H.; Heroldová, M.; Jánová, E.; Stanko, M.; Mošanský, L.; Fričová, J.; Pejčoch, M.; et al. Detection of shrew-borne hantavirus in Eurasian pygmy shrew (Sorex minutus) in Central Europe. Infect. Genet. Evol. 2013, 19, 403–410. [Google Scholar] [CrossRef]
- Hilbe, M.; Herrsche, R.; Kolodziejek, J.; Nowotny, N.; Zlinszky, K.; Ehrensperger, F. Shrews as reservoir hosts of borna disease virus. Emerg. Infect. Dis. 2006, 12, 675–677. [Google Scholar] [CrossRef] [PubMed]
- Dürrwald, R.; Kolodziejek, J.; Weissenböck, H.; Nowotny, N. The bicolored white-toothed shrew Crocidura leucodon (HERMANN 1780) is an indigenous host of mammalian Borna disease virus. PLoS ONE 2014, 9, e93659. [Google Scholar] [CrossRef] [PubMed]
- Ayral, F.; Djelouadji, Z.; Raton, V.; Zilber, A.-L.; Gasqui, P.; Faure, E.; Baurier, F.; Vourc’h, G.; Kodjo, A.; Combes, B. Hedgehogs and Mustelid Species: Major Carriers of Pathogenic Leptospira, a Survey in 28 Animal Species in France (20122015). PLoS ONE 2016, 11, e0162549. [Google Scholar] [CrossRef] [PubMed]
- Jahfari, S.; Ruyts, S.C.; Frazer-Mendelewska, E.; Jaarsma, R.; Verheyen, K.; Sprong, H. Melting pot of tick-borne zoonoses: The European hedgehog contributes to the maintenance of various tick-borne diseases in natural cycles urban and suburban areas. Parasit. Vectors 2017, 10, 134. [Google Scholar] [CrossRef] [Green Version]
- Skuballa, J.; Petney, T.; Pfäffle, M.; Taraschewski, H. Molecular detection of Anaplasma phagocytophilum in the European hedgehog (Erinaceus europaeus) and its ticks. Vector Borne Zoonotic Dis. 2010, 10, 1055–1057. [Google Scholar] [CrossRef]
- Majerová, K.; Gutiérrez, R.; Fonville, M.; Hönig, V.; Papežík, P.; Hofmannová, L.; Lesiczka, P.M.; Nachum-Biala, Y.; Růžek, D.; Sprong, H.; et al. Hedgehogs and Squirrels as Hosts of Zoonotic Bartonella Species. Pathogens 2021, 10, 686. [Google Scholar] [CrossRef]
- Bharti, A.R.; Nally, J.E.; Ricaldi, J.N.; Matthias, M.A.; Diaz, M.M.; Lovett, M.A.; Levett, P.N.; Gilman, R.H.; Willig, M.R.; Gotuzzo, E.; et al. Leptospirosis: A zoonotic disease of global importance. Lancet Infect. Dis. 2003, 3, 757–771. [Google Scholar] [CrossRef]
- Fischer, S.; Mayer-Scholl, A.; Imholt, C.; Spierling, N.G.; Heuser, E.; Schmidt, S.; Reil, D.; Rosenfeld, U.M.; Jacob, J.; Nöckler, K.; et al. Leptospira Genomospecies and Sequence Type Prevalence in Small Mammal Populations in Germany. Vector Borne Zoonotic Dis. 2018, 18, 188–199. [Google Scholar] [CrossRef]
- Mayer-Scholl, A.; Hammerl, J.A.; Schmidt, S.; Ulrich, R.G.; Pfeffer, M.; Woll, D.; Scholz, H.C.; Thomas, A.; Nöckler, K. Leptospira spp. in rodents and shrews in Germany. Int. J. Environ. Res. Public Health 2014, 11, 7562–7574. [Google Scholar] [CrossRef] [Green Version]
- Jeske, K.; Jacob, J.; Drewes, S.; Pfeffer, M.; Heckel, G.; Ulrich, R.G.; Imholt, C. Hantavirus-Leptospira coinfections in small mammals from central Germany. Epidemiol. Infect. 2021, 149, e97. [Google Scholar] [CrossRef]
- Nally, J.E.; Arent, Z.; Bayles, D.O.; Hornsby, R.L.; Gilmore, C.; Regan, S.; McDevitt, A.D.; Yearsley, J.; Fanning, S.; McMahon, B.J. Emerging Infectious Disease Implications of Invasive Mammalian Species: The Greater White-Toothed Shrew (Crocidura russula) Is Associated With a Novel Serovar of Pathogenic Leptospira in Ireland. PLoS Negl. Trop. Dis. 2016, 10, e0005174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rizzoli, A.; Silaghi, C.; Obiegala, A.; Rudolf, I.; Hubálek, Z.; Földvári, G.; Plantard, O.; Vayssier-Taussat, M.; Bonnet, S.; Spitalská, E.; et al. Ixodes ricinus and Its Transmitted Pathogens in Urban and Peri-Urban Areas in Europe: New Hazards and Relevance for Public Health. Front. Public Health 2014, 2, 251. [Google Scholar] [CrossRef] [PubMed]
- Rar, V.; Golovljova, I. Anaplasma, Ehrlichia, and “Candidatus Neoehrlichia” bacteria: Pathogenicity, biodiversity, and molecular genetic characteristics, a review. Infect. Genet. Evol. 2011, 11, 1842–1861. [Google Scholar] [CrossRef]
- Bown, K.J.; Lambin, X.; Telford, G.; Heyder-Bruckner, D.; Ogden, N.H.; Birtles, R.J. The common shrew (Sorex araneus): A neglected host of tick-borne infections? Vector Borne Zoonotic Dis. 2011, 11, 947–953. [Google Scholar] [CrossRef] [Green Version]
- Obiegala, A.; Jeske, K.; Augustin, M.; Król, N.; Fischer, S.; Mertens-Scholz, K.; Imholt, C.; Suchomel, J.; Heroldova, M.; Tomaso, H.; et al. Highly prevalent bartonellae and other vector-borne pathogens in small mammal species from the Czech Republic and Germany. Parasit. Vectors 2019, 12, 332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krügel, M.; Król, N.; Kempf, V.A.J.; Pfeffer, M.; Obiegala, A. Emerging rodent-associated Bartonella: A threat for human health? Parasit. Vectors 2022, 15, 113. [Google Scholar] [CrossRef]
- Mediannikov, O.; El Karkouri, K.; Robert, C.; Fournier, P.-E.; Raoult, D. Non-contiguous finished genome sequence and description of Bartonella florenciae sp. nov. Stand. Genom. Sci. 2013, 9, 185–196. [Google Scholar] [CrossRef] [Green Version]
- Robert Koch-Institut. Infektionsepidemiologisches Jahrbuch meldepflichtiger Krankheiten für 2020. Berl. Ger. 2021, 1–212. [Google Scholar] [CrossRef]
- Körner, S.; Makert, G.R.; Ulbert, S.; Pfeffer, M.; Mertens-Scholz, K. The Prevalence of Coxiella burnetii in Hard Ticks in Europe and Their Role in Q Fever Transmission Revisited-A Systematic Review. Front. Vet. Sci. 2021, 8, 655715. [Google Scholar] [CrossRef]
- Meredith, A.L.; Cleaveland, S.C.; Denwood, M.J.; Brown, J.K.; Shaw, D.J. Coxiella burnetii (Q-Fever) Seroprevalence in Prey and Predators in the United Kingdom: Evaluation of Infection in Wild Rodents, Foxes and Domestic Cats Using a Modified ELISA. Transbound. Emerg. Dis. 2015, 62, 639–649. [Google Scholar] [CrossRef] [Green Version]
- Webster, J.P.; Lloyd, G.; Macdonald, D.W. Q fever (Coxiella burnetii) reservoir in wild brown rat (Rattus norvegicus) populations in the UK. Parasitology 1995, 110, 31–35. [Google Scholar] [CrossRef]
- Reusken, C.; van der Plaats, R.; Opsteegh, M.; de Bruin, A.; Swart, A. Coxiella burnetii (Q fever) in Rattus norvegicus and Rattus rattus at livestock farms and urban locations in the Netherlands; could Rattus spp. represent reservoirs for (re)introduction? Prev. Vet. Med. 2011, 101, 124–130. [Google Scholar] [CrossRef] [PubMed]
- Rehácek, J.; Zupancicová, M.; Ac, P.; Tarasevic, I.V.; Jablonskaja, V.A.; Pospísil, R.; Baloghovă, D. Ricettsioses studies. 2. Natural foci of rickettsioses in east Slovakia. Bull. World Health Organ. 1976, 53, 31–38. [Google Scholar]
- Enkelmann, J.; Stark, K.; Faber, M. Epidemiological trends of notified human brucellosis in Germany, 2006–2018. Int. J. Infect. Dis. 2020, 93, 353–358. [Google Scholar] [CrossRef]
- Scholz, H.C.; Hubalek, Z.; Sedlácek, I.; Vergnaud, G.; Tomaso, H.; Al Dahouk, S.; Melzer, F.; Kämpfer, P.; Neubauer, H.; Cloeckaert, A.; et al. Brucella microti sp. nov., isolated from the common vole Microtus arvalis. Int. J. Syst. Evol. Microbiol. 2008, 58, 375–382. [Google Scholar] [CrossRef] [Green Version]
- Scholz, H.C.; Hofer, E.; Vergnaud, G.; Le Fleche, P.; Whatmore, A.M.; Al Dahouk, S.; Pfeffer, M.; Krüger, M.; Cloeckaert, A.; Tomaso, H. Isolation of Brucella microti from mandibular lymph nodes of red foxes, Vulpes vulpes, in lower Austria. Vector Borne Zoonotic Dis. 2009, 9, 153–156. [Google Scholar] [CrossRef]
- Rónai, Z.; Kreizinger, Z.; Dán, Á.; Drees, K.; Foster, J.T.; Bányai, K.; Marton, S.; Szeredi, L.; Jánosi, S.; Gyuranecz, M. First isolation and characterization of Brucella microti from wild boar. BMC Vet. Res. 2015, 11, 147. [Google Scholar] [CrossRef] [Green Version]
- Hammerl, J.A.; Ulrich, R.G.; Imholt, C.; Scholz, H.C.; Jacob, J.; Kratzmann, N.; Nöckler, K.; Al Dahouk, S. Molecular Survey on Brucellosis in Rodents and Shrews—Natural Reservoirs of Novel Brucella Species in Germany? Transbound. Emerg. Dis. 2017, 64, 663–671. [Google Scholar] [CrossRef]
- Schulze, V.; Große, R.; Fürstenau, J.; Forth, L.F.; Ebinger, A.; Richter, M.T.; Tappe, D.; Mertsch, T.; Klose, K.; Schlottau, K.; et al. Borna disease outbreak with high mortality in an alpaca herd in a previously unreported endemic area in Germany. Transbound. Emerg. Dis. 2020, 67, 2093–2107. [Google Scholar] [CrossRef] [Green Version]
- Ulrich, R.G.; Schmidt-Chanasit, J.; Schlegel, M.; Jacob, J.; Pelz, H.-J.; Mertens, M.; Wenk, M.; Büchner, T.; Masur, D.; Sevke, K.; et al. Network “Rodent-borne pathogens” in Germany: Longitudinal studies on the geographical distribution and prevalence of hantavirus infections. Parasitol. Res. 2008, 103, 121–129. [Google Scholar] [CrossRef]
- Schlegel, M.; Ali, H.S.; Stieger, N.; Groschup, M.H.; Wolf, R.; Ulrich, R.G. Molecular identification of small mammal species using novel Cytochrome b gene-derived degenerated primers. Biochem. Genet. 2012, 50, 440–447. [Google Scholar] [CrossRef]
- Schmidt, E.; Obiegala, A.; Imholt, C.; Drewes, S.; Saathoff, M.; Freise, J.; Runge, M.; Jacob, J.; Mayer-Scholl, A.; Ulrich, R.G.; et al. Influence of Season, Population and Individual Characteristics on the Prevalence of Leptospira spp. in Bank Voles in North-West Germany. Biology 2021, 10, 933. [Google Scholar] [CrossRef]
- Nau, L.H.; Obiegala, A.; Król, N.; Mayer-Scholl, A.; Pfeffer, M. Survival time of Leptospira kirschneri serovar Grippotyphosa under different environmental conditions. PLoS ONE 2020, 15, e0236007. [Google Scholar] [CrossRef]
- Boonsilp, S.; Thaipadungpanit, J.; Amornchai, P.; Wuthiekanun, V.; Bailey, M.S.; Holden, M.T.G.; Zhang, C.; Jiang, X.; Koizumi, N.; Taylor, K.; et al. A single multilocus sequence typing (MLST) scheme for seven pathogenic Leptospira species. PLoS Negl. Trop. Dis. 2013, 7, e1954. [Google Scholar] [CrossRef] [Green Version]
- Norman, A.F.; Regnery, R.; Jameson, P.; Greene, C.; Krause, D.C. Differentiation of Bartonella-like isolates at the species level by PCR-restriction fragment length polymorphism in the citrate synthase gene. J. Clin. Microbiol. 1995, 33, 1797–1803. [Google Scholar] [CrossRef] [Green Version]
- Kosoy, M.Y.; Regnery, R.L.; Tzianabos, T.; Marston, E.L.; Jones, D.C.; Green, D.; Maupin, G.O.; Olson, J.G.; Childs, J.E. Distribution, diversity, and host specificity of Bartonella in rodents from the Southeastern United States. Am. J. Trop. Med. Hyg. 1997, 57, 578–588. [Google Scholar] [CrossRef]
- Silaghi, C.; Liebisch, G.; Pfister, K. Genetic variants of Anaplasma phagocytophilum from 14 equine granulocytic anaplasmosis cases. Parasit. Vectors 2011, 4, 161. [Google Scholar] [CrossRef] [Green Version]
- Silaghi, C.; Woll, D.; Mahling, M.; Pfister, K.; Pfeffer, M. Candidatus Neoehrlichia mikurensis in rodents in an area with sympatric existence of the hard ticks Ixodes ricinus and Dermacentor reticulatus, Germany. Parasit. Vectors 2012, 5, 285. [Google Scholar] [CrossRef] [Green Version]
- Galfsky, D.; Król, N.; Pfeffer, M.; Obiegala, A. Long-term trends of tick-borne pathogens in regard to small mammal and tick populations from Saxony, Germany. Parasit. Vectors 2019, 12, 131. [Google Scholar] [CrossRef] [Green Version]
- Obiegala, A.; Heuser, E.; Ryll, R.; Imholt, C.; Fürst, J.; Prautsch, L.-M.; Plenge-Bönig, A.; Ulrich, R.G.; Pfeffer, M. Norway and black rats in Europe: Potential reservoirs for zoonotic arthropod-borne pathogens? Pest Manag. Sci. 2019, 75, 1556–1563. [Google Scholar] [CrossRef]
- Klee, S.R.; Tyczka, J.; Ellerbrok, H.; Franz, T.; Linke, S.; Baljer, G.; Appel, B. Highly sensitive real-time PCR for specific detection and quantification of Coxiella burnetii. BMC Microbiol. 2006, 6, 2. [Google Scholar] [CrossRef] [Green Version]
- Probert, W.S.; Schrader, K.N.; Khuong, N.Y.; Bystrom, S.L.; Graves, M.H. Real-time multiplex PCR assay for detection of Brucella spp., B. abortus, and B. melitensis. J. Clin. Microbiol. 2004, 42, 1290–1293. [Google Scholar] [CrossRef] [Green Version]
- Kraft, R. (Ed.) Mäuse und Spitzmäuse in Bayern: Verbreitung, Lebensraum, Bestandssituation; Ulmer: Stuttgart, Germany, 2008; ISBN 3800156091. [Google Scholar]
- Niethammer, J. Zur gegenwärtigen Nordgrenze von Crocidura leucodon in Niedersachsen. Z. Säugetierkunde 1980, 45, 192. [Google Scholar]
- Roschen, A.; Hellbernd, L.; Nettmann, H.-K. Die Verbreitung von Crocidura russula und Crocidura leucodon in der Bremer Wesermarsch. Z. Säugetierkunde 1984, 49, 70–74. [Google Scholar]
- Güttinger, R.; Pfunder, M.; Wüst, M. Die Verbreitung von Feldspitzmaus Crodidura leucodon und Hausspitzmaus C. russula in der Ostschweiz: Eine spezielle Situation in ihrer zoogeografischen Kontaktzone. Gall. Nat. Ges. 2008, 91, 179–194. [Google Scholar] [CrossRef]
- Krämer, M.; Jentzsch, M. Kleinsäuger-Vorkommen aus dem Raum Zeitz—Eine vergleichende Studie. Mauritiana 2008, 20, 411–427. [Google Scholar]
- Dornberger, W. Fund einer Hausspitzmaus (Crocidura russula) in Niederstetten. Faun-Und-Flor-Mitt 1990, 9, 84. [Google Scholar]
- Klesser, R.; Jessen, F.; Ringenberg, J.; Preuß, M.; Kaiser, T.; Husemann, M. Return of the walking dead: First verified record of the shrew Crocidura leucodon (Hermann, 1780) in Hamburg, Germany. EvolSyst 2021, 5, 121–128. [Google Scholar] [CrossRef]
- Nationalatlas Bundesrepublik Deutschland: Verbreitung der Säugetierarten; Leibniz-Institut für Länderkunde: Leipzig, Germany, 2003.
- Frank, F. Zur Arealverschiebung zwischen Crocidura russula und C. leucodon in NW-Deutschland und zum wechselseitigen Verhältnis beider Arten. Z. Säugetierkunde 1984, 49, 65–70. [Google Scholar]
- Worschech, K. Ehemaliges und gegenwärtiges Vorkommen der Weißzahnspitzmäuse (Crocidura WAGLER, 1832) im Altenburger Land (Thüringen) (Mammalia:Soricidae). Mauritiana 2010, 21, 249–267. [Google Scholar]
- Paliocha, E.; Wilhelm, P. Forschungsprojekt “Wildlebende Säugetiere in Baden-Württemberg” Bericht über das Forschungsvorhaben: 1. Gewöllanalyse. MAUS. Mitt. Unserer Säugetierwelt 1993, 3, 3. [Google Scholar]
- Brünner, H. Erfassung von Säugetieren im LIFE-Projektgebiet “Lebendige Rheinauen bei Karlsruhe. 2010. Available online: https://rp.baden-wuerttemberg.de/fileadmin/RP-Internet/Karlsruhe/Abteilung_5/Referat_56/Lebendige_Rheinauen/_DocumentLibraries/ErgebnisseundVortrge/Endberichte/08_leb_rhein_endb_saeuger.pdf (accessed on 23 April 2023).
- Spitzenberger, F. Die Säugetierfauna Österreichs; Austria Medien Service GmbH: Graz, Austria, 2001; ISBN 3-85333-063-0. [Google Scholar]
- Jentzsch, M.; Trost, M. Zum Vorkommen der Gartenspitzmaus Crocidura suaveolens (Pallas, 1811) in Sachsen-Anhalt. Hercynia 2008, 41, 135–141. [Google Scholar]
- Schmidt, A. Reaktionen von Säugetierarten auf die Klimaerwärmung—Eine Auswahl von Beispielen, insbesondere aus der Fledermausfauna. Nyctalus (N.F.) 1998, 6, 609–613. [Google Scholar]
- Schmidt, A. Zur Verbreitungsgeschichte der Gartenspitzmaus Crocidura suaveolens in Ostdeutschland. Nat. Und Landsch. Brandenbg. 1998, 1998, 49–51. [Google Scholar]
- Vogel, P.; Cosson, J.-F.; López Jurado, L.F. Taxonomic status and origin of the shrews (Soricidae) from the Canary islands inferred from a mtDNA comparison with the European Crocidura species. Mol. Phylogenet. Evol. 2003, 27, 271–282. [Google Scholar] [CrossRef] [Green Version]
- Jaquiéry, J.; Guélat, J.; Broquet, T.; Berset-Brändli, L.; Pellegrini, E.; Moresi, R.; Hirzel, A.H.; Perrin, N. Habitat-quality effects on metapopulation dynamics in greater white-toothed shrews, Crocidura russula. Ecology 2008, 89, 2777–2785. [Google Scholar] [CrossRef] [Green Version]
- Torten, M.; Eliash, Z.; Lawrence, D.; Shenberg, E. Crocidura russula, a hitherto unknown carrier of leptospires. Isr. J. Med. Sci. 1972, 8, 1917–1920. [Google Scholar]
- Obiegala, A.; Albrecht, C.; Dafalla, M.; Drewes, S.; Oltersdorf, C.; Turni, H.; Imholt, C.; Jacob, J.; Wagner-Wiening, C.; Ulrich, R.G.; et al. Leptospira spp. in Small Mammals from Areas with Low and High Human Hantavirus Incidences in South-West Germany. Vector Borne Zoonotic Dis. 2017, 17, 312–318. [Google Scholar] [CrossRef]
- Obiegala, A.; Woll, D.; Karnath, C.; Silaghi, C.; Schex, S.; Eßbauer, S.; Pfeffer, M. Prevalence and Genotype Allocation of Pathogenic Leptospira Species in Small Mammals from Various Habitat Types in Germany. PLoS Negl. Trop. Dis. 2016, 10, e0004501. [Google Scholar] [CrossRef] [Green Version]
- Wright, T.F.; Eberhard, J.R.; Hobson, E.A.; Avery, M.L.; Russello, M.A. Behavioral flexibility and species invasions: The adaptive flexibility hypothesis. Ethol. Ecol. Evol. 2010, 22, 393–404. [Google Scholar] [CrossRef]
- Ferreira, A.S.; Ahmed, A.; Rocha, T.; Vieira, M.L.; Paiva-Cardoso, M.d.N.; Mesquita, J.R.; van der Linden, H.; Goris, M.; Thompson, G.; Hartskeerl, R.A.; et al. Genetic diversity of pathogenic leptospires from wild, domestic and captive host species in Portugal. Transbound. Emerg. Dis. 2020, 67, 852–864. [Google Scholar] [CrossRef]
- Stritof Majetic, Z.; Galloway, R.; Ruzic Sabljic, E.; Milas, Z.; Mojcec Perko, V.; Habus, J.; Margaletic, J.; Pernar, R.; Turk, N. Epizootiological survey of small mammals as Leptospira spp. reservoirs in Eastern Croatia. Acta Trop. 2014, 131, 111–116. [Google Scholar] [CrossRef] [PubMed]
- Renaud, C.; Andrews, S.; Djelouadji, Z.; Lecheval, S.; Corrao-Revol, N.; Buff, S.; Demont, P.; Kodjo, A. Prevalence of the Leptospira serovars bratislava, grippotyphosa, mozdok and pomona in French dogs. Vet. J. 2013, 196, 126–127. [Google Scholar] [CrossRef] [PubMed]
- Da Cunha, C.E.P.; Felix, S.R.; Neto, A.C.P.S.; Campello-Felix, A.; Kremer, F.S.; Monte, L.G.; Amaral, M.G.; de Oliveira Nobre, M.; Da Silva, É.F.; Hartleben, C.P.; et al. Infection with Leptospira kirschneri Serovar Mozdok: First Report from the Southern Hemisphere. Am. J. Trop. Med. Hyg. 2016, 94, 519–521. [Google Scholar] [CrossRef] [Green Version]
- Jolley, K.A.; Bray, J.E.; Maiden, M.C.J. Open-Access Bacterial Population Genomics: BIGSdb Software. Wellcome Open Res. 2018, Volume 3. The PubMLST.org Website and Their Applications. Available online: https://pubmlst.org/bigsdb?page=info&db=pubmlst_leptospira_isolates&id=503 (accessed on 23 April 2023). [CrossRef]
- Barandika, J.F.; Hurtado, A.; García-Esteban, C.; Gil, H.; Escudero, R.; Barral, M.; Jado, I.; Juste, R.A.; Anda, P.; García-Pérez, A.L. Tick-borne zoonotic bacteria in wild and domestic small mammals in northern Spain. Appl. Environ. Microbiol. 2007, 73, 6166–6171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silaghi, C.; Beck, R.; Oteo, J.A.; Pfeffer, M.; Sprong, H. Neoehrlichiosis: An emerging tick-borne zoonosis caused by Candidatus Neoehrlichia mikurensis. Exp. Appl. Acarol. 2016, 68, 279–297. [Google Scholar] [CrossRef] [PubMed]
- Bray, D.P.; Bown, K.J.; Stockley, P.; Hurst, J.L.; Bennett, M.; Birtles, R.J. Haemoparasites of common shrews (Sorex araneus) in Northwest England. Parasitology 2007, 134, 819–826. [Google Scholar] [CrossRef] [Green Version]
- Lin, J.-W.; Hsu, Y.-M.; Chomel, B.B.; Lin, L.-K.; Pei, J.-C.; Wu, S.-H.; Chang, C.-C. Identification of novel Bartonella spp. in bats and evidence of Asian gray shrew as a new potential reservoir of Bartonella. Vet. Microbiol. 2012, 156, 119–126. [Google Scholar] [CrossRef]
- Liyai, R.; Kimita, G.; Masakhwe, C.; Abuom, D.; Mutai, B.; Onyango, D.M.; Waitumbi, J. The spleen bacteriome of wild rodents and shrews from Marigat, Baringo County, Kenya. PeerJ 2021, 9, e12067. [Google Scholar] [CrossRef]
- Celebi, B.; Anani, H.; Zgheib, R.; Carhan, A.; Raoult, D.; Fournier, P.-E. Genomic Characterization of the Novel Bartonella refiksaydamii sp. Isolated from the Blood of a Crocidura suaveolens (Pallas, 1811). Vector Borne Zoonotic Dis. 2021, 21, 432–440. [Google Scholar] [CrossRef]
- Vieira-Damiani, G.; Diniz, P.P.V.d.P.; Pitassi, L.H.U.; Sowy, S.; Scorpio, D.G.; Lania, B.G.; Drummond, M.R.; Soares, T.C.B.; Barjas-Castro, M.d.L.; Breitschwerdt, E.B.; et al. Bartonella clarridgeiae Bacteremia Detected in an Asymptomatic Blood Donor. J. Clin. Microbiol. 2014, 53, 352–356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Špitalská, E.; Boldiš, V.; Mošanský, L.; Sparagano, O.; Stanko, M. Rickettsia species in fleas collected from small mammals in Slovakia. Parasitol. Res. 2015, 114, 4333–4339. [Google Scholar] [CrossRef] [PubMed]
- Morrone, J.J.; Acosta, R. A synopsis of the fleas (Insecta: Siphonaptera) parasitizing New World species of Soricidae (Mammalia: Insectivora). Zootaxa 2006, 1354, 1–30. [Google Scholar] [CrossRef]
- Heglasová, I.; Rudenko, N.; Golovchenko, M.; Zubriková, D.; Miklisová, D.; Stanko, M. Ticks, fleas and rodent-hosts analyzed for the presence of Borrelia miyamotoi in Slovakia: The first record of Borrelia miyamotoi in a Haemaphysalis inermis tick. Ticks Tick Borne Dis. 2020, 11, 101456. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Species | Number of Leptospira DNA-Positive/Total Number of Tested Individuals (Percentage, 95% CI *) | Number of N. mikurensis DNA-Positive/Total Number of Tested Individuals (Percentage, 95% CI *) | Number of A. phagocytophilum, C. burnetii, Brucella spp., Babesia spp. and Bartonella spp. DNA-Positive/Total Number of Tested Individuals (Percentage, 95% CI *) |
|---|---|---|---|
| Greater white-toothed shrew *** (Crocidura russula) | 28/227 (12.3%, 8.6–17.3) | 2/213 (0.9%, 0–3.6) | 0/213 (0%, 0–2.1) |
| Bicoloured white-toothed shrew *** (Crocidura leucodon) | 3/81 ** (3.7%, 0.8–10.7) | 0/80 (0%, 0–5.5) | 0/80 (0%, 0–5.5) |
| Lesser white-toothed shrew *** (Crocidura suaveolens) | 0/22 (0%, 0–17.6) | 0/21 (0%, 0–18.2) | 0/21 (0%, 0–18.2) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Haring, V.; Jacob, J.; Walther, B.; Trost, M.; Stubbe, M.; Mertens-Scholz, K.; Melzer, F.; Scuda, N.; Gentil, M.; Sixl, W.; et al. White-Toothed Shrews (Genus Crocidura): Potential Reservoirs for Zoonotic Leptospira spp. and Arthropod-Borne Pathogens? Pathogens 2023, 12, 781. https://doi.org/10.3390/pathogens12060781
Haring V, Jacob J, Walther B, Trost M, Stubbe M, Mertens-Scholz K, Melzer F, Scuda N, Gentil M, Sixl W, et al. White-Toothed Shrews (Genus Crocidura): Potential Reservoirs for Zoonotic Leptospira spp. and Arthropod-Borne Pathogens? Pathogens. 2023; 12(6):781. https://doi.org/10.3390/pathogens12060781
Chicago/Turabian StyleHaring, Viola, Jens Jacob, Bernd Walther, Martin Trost, Michael Stubbe, Katja Mertens-Scholz, Falk Melzer, Nelly Scuda, Michaela Gentil, Wolfdieter Sixl, and et al. 2023. "White-Toothed Shrews (Genus Crocidura): Potential Reservoirs for Zoonotic Leptospira spp. and Arthropod-Borne Pathogens?" Pathogens 12, no. 6: 781. https://doi.org/10.3390/pathogens12060781