
The Potential of Probiotics as Ingestible Adjuvants and Immune Modulators for Antiviral Immunity and Management of SARS-CoV-2 Infection and COVID-19



Abstract
:1. Introduction
2. Gut Mucosal Immune Function and Influence of Commensal Microbiome and Probiotics
2.1. The Gut Microbiome
2.2. Structure–Functionality of the Gut Mucosa
2.3. Probiotic-Derived Metabolites Modulate Host Immunity
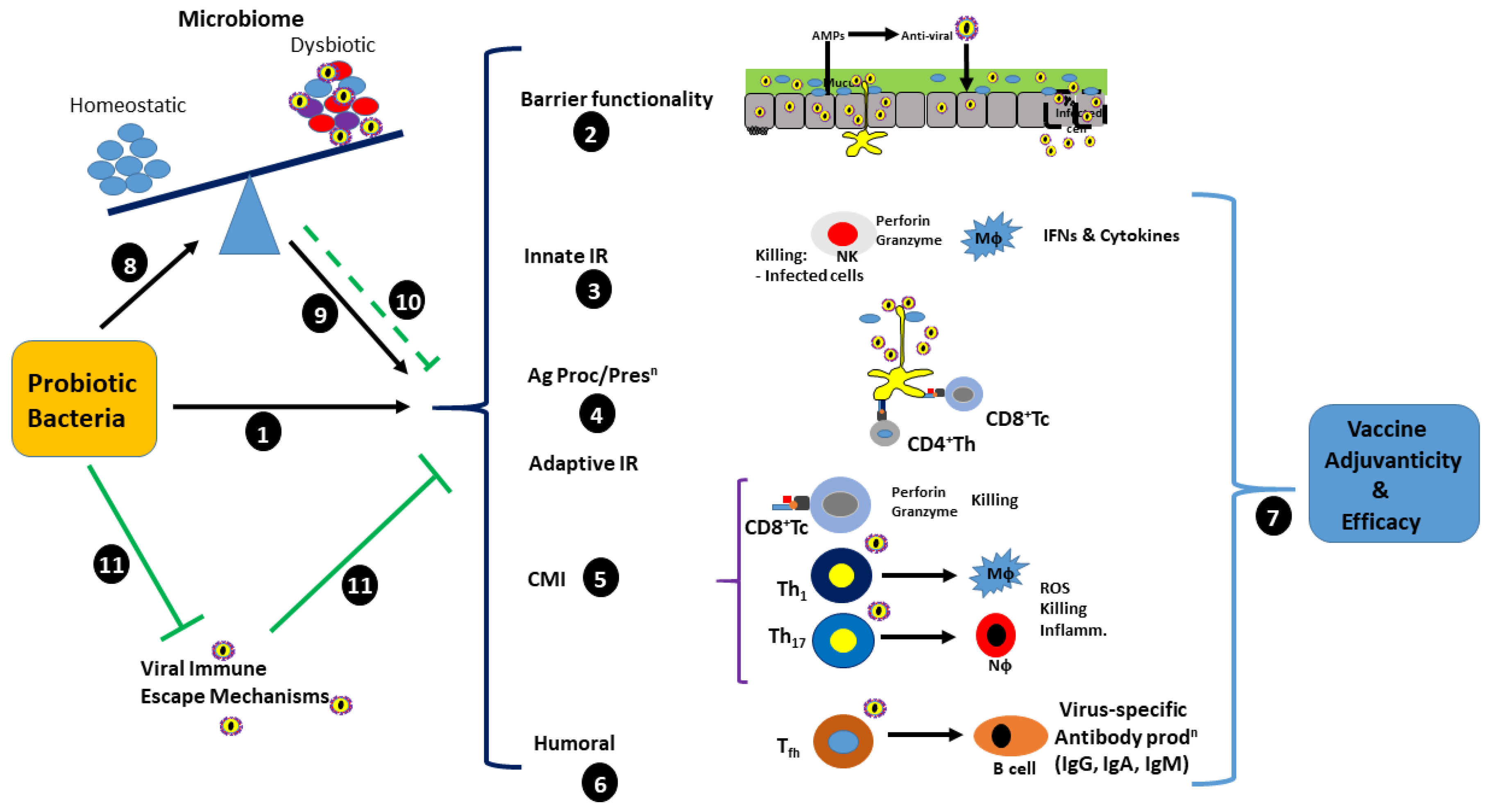
3. Immunomodulatory and Antiviral Capability of Probiotics
4. COVID-19: Infection by SARS-CoV-2
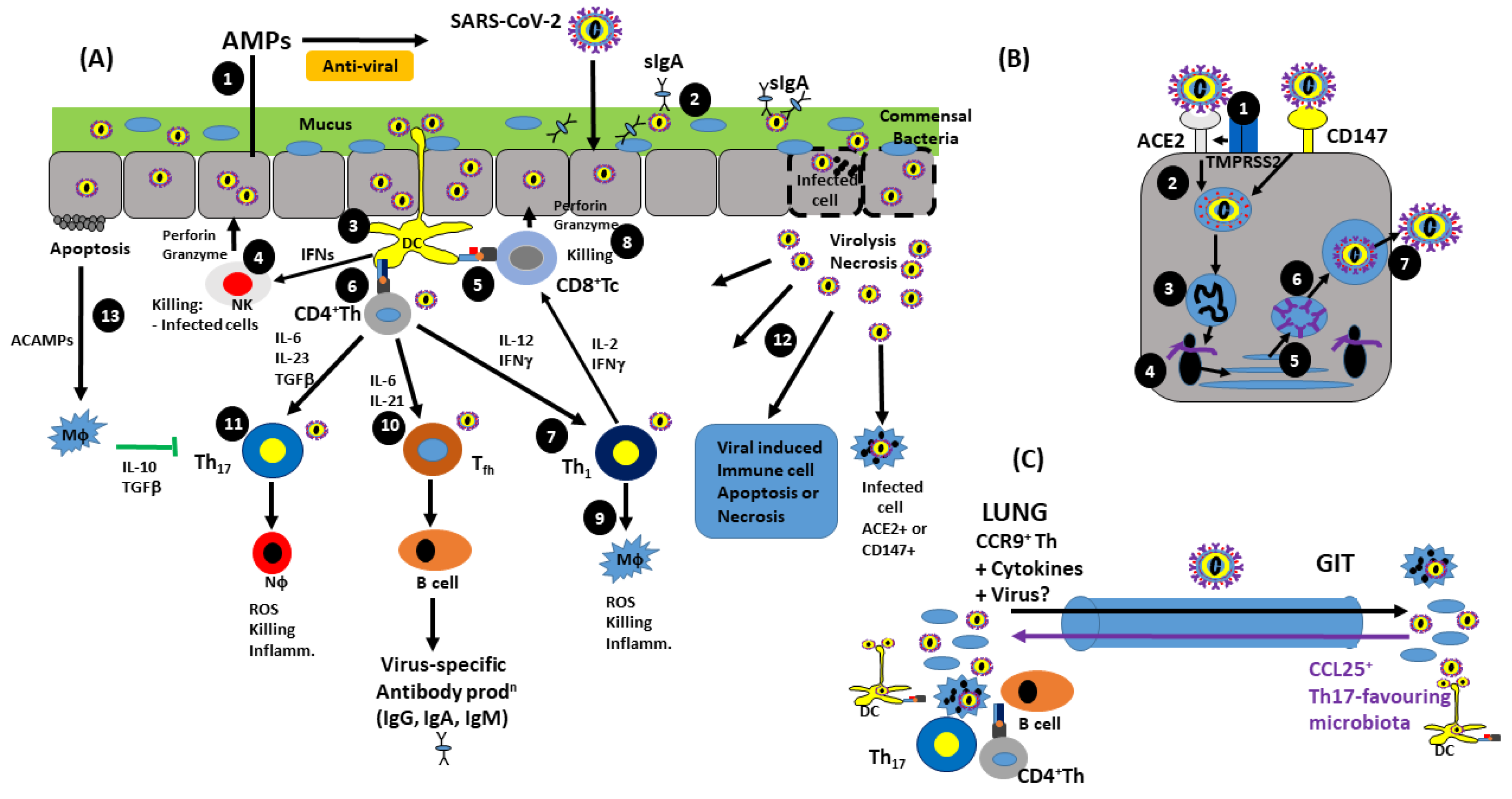
5. Antiviral Immunity and SARS-CoV-2 Immunopathology
5.1. Innate Immunity
5.2. Adaptive Immunity
5.3. Viral Immune Evasion Mechanisms

{kind=link}
{kind=link}
| Viral Immune Evasion Response | Infection/Pathology | Reference |
|---|---|---|
| Recognition by PRRs | M inhibits RIG-1,MDA5, and MAVS 9b inhibits interaction between RIG-1 and MAVS Nsps-3,-4,-6 encode double-membrane vesicles, hiding dsRNA from RLRs N protein binds TRIM25, preventing RIG-1 actn | [134] |
| Type I IFN production and signalling | Nsp-1 degrades IFN mRNA Nsp-1blocks STAT-1 phosphorylation—delaying type I IFN production Nsp6 inhibits TBK1 phosphorylation of IRF7 7a destabilises TBK1, inhibiting IRF3 phosphorylation SARS-CoV-2 ORF6 inhibits downstream IFNα signalling Nsps-1,-3,-13,-14, ORFs-6,-8, M and N inhibit IFN type I-induced ISG gene expression N protein binds TRIM25, preventing RIG-1 actn and reduction in IFNβ production IAV Nsp1 inhibits RIG-1-IFNβ production SARS-CoV-2 ORF3b—potent antagonist of IFN prodn via suppression of IRF3 nuclear translocation | [134,135,136,137,138] |
| Anti-inflammatory cytokine production | Nsp3 ORF9b and M inhibit NFκB activation SARS-CoV-2 strongly induces AM IL-10 prodn | [88] |
| Suppression of MHC expression | MHC I expression inhibited by ORF3a and ORF7a SARS downregulates DC MHC II expression MERS downregulates Mϕ MHC II SARS-CoV-2 ORF8 downregulates T cell MHC I | [87,139] |
| Suppression of NK cell activation | NKG2A upregulation—inhibits NK-mediated cell cytotoxicity (also on CD8+ Tc) Increased IL-6 and IL-10 inhibit STAT-3-dependent NK activation Both IL-6 and IL-10 increase NKG2A expression | [140,141] |
Inhibition of Cell-mediated immunity
| SARS-CoV-2 ORF3B antagonism of IFN production. NKG2A upregulation—inhibits CD8+ Tc-mediated cell cytotoxicity (also on NKs) SARS downregulates DC MHC II and B7 expression Antigenic mutation of M protein | [100,142,143] |
Inhibition of Humoral Immunity
| Omicron variant—high mutational burden in S protein: increased Ab evasion. SARS downregulates DC MHC II and B7 expression | [144,145] |
| Inhibition of receptor binding | Flexible RBD in S trimers: RBD exposed in standing state. Lying state—RBD not exposed, hence reduced binding, infection and immunogenicity | [146] |
5.4. SARS-CoV-2 Infects and Affects Tissues/Organs Distal to the Gut–Lung Axis
6. Probiotics Reduce COVID-19 Symptoms of the Gut Mucosa via the Gut–Lung Axis

6.1. The Potential Role of Probiotics in the Prophylaxis of SARS-CoV-2 Infection and Pathology
6.2. The Potential Role of Probiotics in the Treatment of SARS-CoV-2 Associated Secondary Infection
7. Summary and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yan, F.; Polk, D. Probiotics and Immune Health. Curr. Opin. Gastroenterol. 2014, 27, 496–501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manohar, P.; Loh, B.; Nachimuthu, R.; Hua, X.; Welburn, S.C.; Leptihn, S. Secondary Bacterial Infections in Patients With Viral Pneumonia. Front. Med. 2020, 7, 420. [Google Scholar] [CrossRef] [PubMed]
- Raoult, D.; Zulma, A.; Locatelli, F.; Ippolito, G.; Kroemer, G. Coronavirus Infections: Epidemiological, Clinical, and Immunological Features and Hypotheses. Cell Stress 2020, 4, 66–75. [Google Scholar] [CrossRef] [PubMed]
- Richman, D.D.; Whitley, R.J.; Hayden, F.G. Clinical Virology, 4th ed.; ASM Press: Washington, DC, USA, 2016. [Google Scholar]
- Melley, L.E.; Bress, E.; Polan, E. Hypogeusia as the initial presenting symptom of COVID-19. BMJ Case Rep. 2020, 13, e236080. [Google Scholar] [CrossRef] [PubMed]
- Zaim, S.; Chong, J.H.; Sankaranarayanan, V.; Harky, A. COVID-19 and Multiorgan Response. Curr. Probl. Cardiol. 2020, 45, 100618. [Google Scholar] [CrossRef]
- Mirzaei, R.; Goodarzi, P.; Asadi, M.; Soltani, A.; Aljanabi, H.; Jeda, A.S.; Dastbin, S.; Jalalifar, S.; Mohammadzadeh, R.; Teimoori, A.; et al. Bacterial Co-infections with SARS-CoV-2. IUBMB Life 2020, 72, 2097–2111. [Google Scholar] [CrossRef]
- Hojyo, S.; Uchida, M.; Tanaka, K.; Hasebe, R.; Tanaka, Y.; Murakami, M.; Hirano, T. How COVID-19 Induces Cytokine Storm with High Mortality. Inflamm. Regen. 2020, 40, 37. [Google Scholar] [CrossRef]
- Guaraldi, G.; Meschiari, M.; Cozzi-Lepri, A.; Milic, J.; Tonelli, R.; Menozzi, M.; Franceschini, E.; Cuomo, G.; Orlando, G.; Borghi, V.; et al. Tocilizumab in Patients with Severe COVID-19: A Retrospective Cohort Study. Lancet Rheumatol. 2020, 2, e474–e484. [Google Scholar] [CrossRef]
- Horby, P.; Lim, W.S.; Emberson, J.R.; Mafham, M.; Bell, J.L.; Linsell, L.; Staplin, N.; Brightling, C.; Ustianowski, A.; Elmahi, E.; et al. Dexamethasone in Hospitalized Patients with COVID-19. New Engl. J. Med. 2021, 384, 693–704. [Google Scholar] [CrossRef]
- Hardy, H.; Harris, J.; Lyon, E.; Beal, J.; Foey, A.D. Probiotics, Prebiotics and Immunomodulation of Gut Mucosal Defences: Homeostasis and Immunopathology. Nutrients 2013, 5, 1869–1912. [Google Scholar] [CrossRef] [Green Version]
- Esakandari, H.; Nabi-Afjadi, M.; Fakkari-Afjadi, J.; Farahmandian, N.; Miresmaeili, S.M.; Bahreini, E. A Comprehensive Review of COVID-19 Characteristics. Biol. Proced. Online 2020, 22, 19. [Google Scholar] [CrossRef] [PubMed]
- Gill, S.R.; Pop, M.; Deboy, R.T.; Eckburg, P.B.; Turnbaugh, P.J.; Samuel, B.S.; Gordon, J.I.; Relman, D.A.; Fraser-Liggett, C.M.; Nelson, K.E. Metagenomic Analysis of the Human Distal Gut Microbiome. Science 2006, 312, 1355–1359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.; Pu, J.; Lu, S.; Bai, X.; Wu, Y.; Jin, D.; Cheng, Y.; Zhang, G.; Zhu, W.; Luo, X.; et al. Species-Level Analysis of Human Gut Microbiota with Metataxonomics. Front. Microbiol. 2020, 11, 2029. [Google Scholar] [CrossRef] [PubMed]
- Senico, V.; Machado, M.G.; Trottein, F. The Lung-Gut Axis during Viral Respiratory Infections: The Impact of Gut Dysbiosis on Secondary Disease Outcomes. Mucosal Immunol. 2021, 14, 296–304. [Google Scholar] [CrossRef]
- Cameron, E.A.; Curtis, M.M.; Kumar, A.; Dunny, G.M.; Sperandio, V. Microbiota and Pathogen Proteases Modulate Type III Secretion Activity in Enterohemorrhagic Escherichia coli. mBio 2018, 9, e02204-18. [Google Scholar] [CrossRef] [Green Version]
- Laitinen, K.; Mokkala, K.; Kalliomaki, M. Chapter 8—Impact of Early Nutrition on Intestinal Microbiome: Effects on Immunity and Long-Term Health. In Early Nutrition and Long-Term Health–Mechanisms, Consequences, and Opportunities; Saavedra, J.M., Dattilo, A.M., Eds.; Woodhead Publishing: Cambridge, UK, 2017; pp. 212–213. [Google Scholar]
- McDermott, A.J.; Huffnagle, G.B. The Microbiome and Regulation of Mucosal Immunity. Immunology 2014, 142, 24–31. [Google Scholar] [CrossRef] [Green Version]
- Shi, N.; Li, N.; Duan, X.; Niu, H. Interaction between the Gut Microbiome and Mucosal Immune System. Mil. Med. Res. 2017, 4, 14. [Google Scholar] [CrossRef] [Green Version]
- Zhu, G.; Hu, J.; Xi, R. The Cellular Niche for Intestinal Stem Cells: A Team Effort. Cell Regen. 2021, 10, 1. [Google Scholar] [CrossRef]
- Tong, Y.; Tang, J. Candida albicans Infection and Intestinal Immunity. Microbiol. Res. 2017, 198, 27–35. [Google Scholar] [CrossRef]
- Singh, N.; Gallagher, H.C.; Song, R.; Dhinsa, J.K.; Ostroff, G.R.; De Jesus, M. RNA Isolation from Peyer’s Patch Lymphocytes and Mononuclear Phagocytes to Determine Gene Expression Profiles Using NanoString Technology. J. Biol. Methods 2018, 5, e95. [Google Scholar] [CrossRef] [Green Version]
- Kogan, A.N.; von Andrian, U.H. Chapter 10—Lymphocyte Trafficking. In Microcirculation, 2nd ed.; Tuma, R.F., Durán, W.N., Ley, K., Eds.; Academic Press: San Diego, CA, USA, 2008; pp. 449–482. [Google Scholar]
- Littman, D.R.; Pamer, E.G. Role of the Commensal Microbiota in Normal and Pathogenic Host Immune Responses. Cell Host Microbe 2011, 10, 311–323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, E.M.; Sadarangani, M.; Finlay, B.B. The Role of the Immune System in Governing Host-Microbiome Interactions in the Intestine. Nat. Immunol. 2013, 14, 660–667. [Google Scholar] [CrossRef] [PubMed]
- Chunxi, L.; Haiyue, L.; Yanxia, L.; Jianbing, P.; Jin, S. The Gut Microbiota and Respiratory Diseases: New Evidence. J. Immunol. Res. 2020, 2020, 2340670. [Google Scholar] [CrossRef] [PubMed]
- Enaud, R.; Prevel, R.; Ciarlo, E.; Beaufils, F.; Wieёrs, G.; Guery, B.; Delhaes, L. The Gut-Lung Axis in Health and Respiratory Diseases: A Place for Inter-Organ and Inter-Kingdom Crosstalks. Front. Cell. Infect. Microbiol. 2020, 10, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schuijt, T.J.; Lankelma, J.M.; Scicluna, B.P.; de Sousa e Melo, F.; Roelofs, J.J.; de Boer, J.D.; Hoogendijk, A.J.; de Beer, R.; de Vos, A.; Belzer, C.; et al. The Gut Microbiota Plays a Protective Role in the Host Defence Against Pneumococcal Pneumonia. Gut 2016, 65, 575–583. [Google Scholar] [CrossRef] [Green Version]
- Shu, Y.; Li, C.K.F.; Gao, R.; Liang, Q.; Zhang, Y.; Dong, L.; Zhou, J.; Dong, J.; Wang, D.; Wen, L.; et al. Avian Influenza A (H5N1) Viruses can Directly Infect and Replicate in Human Gut Tissues. J. Infect. Dis. 2010, 201, 1173–1177. [Google Scholar] [CrossRef]
- Scaldaferri, F.; Pugliese, D.; Privitera, G.; Onali, S.; Lopetuso, L.R.; Rizzatti, G.; Settanni, C.R.; Pizzoferrato, M.; Schiavoni, E.; Turchini, L.; et al. Impact of COVID-19 Pandemic on the Daily Management of Biotechnological Therapy in Inflammatory Bowel Disease Patients: Reorganisational Response in a High-Volume Italian Inflammatory Bowel Disease Centre. United Eur. Gastroenterol. J. 2020, 8, 775–781. [Google Scholar] [CrossRef]
- Abt, M.C.; Osborne, L.C.; Monticelli, L.A.; Doering, T.A.; Alenghat, T.; Sonnenberg, G.F.; Paley, M.A.; Antenus, M.; Williams, K.L.; Erikson, J.; et al. Commensal Bacteria Calibrate the Activation Threshold of Innate Antiviral Immunity. Immunity 2013, 37, 158–170. [Google Scholar] [CrossRef] [Green Version]
- Smith, K.; McCoy, K.D.; Macpherson, A.J. Use of Axenic Animals in Studying the Adaptation of Mammals to Their Commensal Intestinal Microbiota. Semin. Immunol. 2007, 19, 59–69. [Google Scholar] [CrossRef]
- Mazmanian, S.K.; Liu, C.H.; Tzianabos, A.O.; Kasper, D.L. An Immunomodulatory Molecule of Symbiotic Bacteria Directs Maturation of the Host Immune System. Cell 2005, 122, 107–118. [Google Scholar] [CrossRef] [Green Version]
- Samuelson, D.R.; Welsh, D.A.; Shellito, J.E. Regulation of Lung Immunity and Host Defence by the Intestinal Microbiota. Front. Microbiol. 2015, 6, 1085. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonçalves, P.; Araújo, J.R.; Di Santo, J.P. A Cross-Talk Between Microbiota-Derived Short-Chain Fatty Acids and the Host Mucosal Immune System Regulates Intestinal Homeostasis and Inflammatory Bowel Disease. Inflamm. Bowel Dis. 2018, 24, 558–572. [Google Scholar] [CrossRef]
- Sun, M.; Wu, W.; Liu, Z.; Cong, Y. Microbiota Metabolite Short Chain Fatty Acids, GPCR, and Inflammatory Bowel Diseases. J. Gastroenterol. 2017, 52, 1–8. [Google Scholar] [CrossRef]
- Ji, J.; Shu, D.; Zheng, M.; Wang, J.; Luo, C.; Wang, Y.; Guo, F.; Zou, X.; Lv, X.; Li, Y.; et al. Microbial Metabolite Butyrate Facilitates M2 Macrophage Polarisation and Function. Sci. Rep. 2016, 6, 24838. [Google Scholar] [CrossRef]
- Foey, A.D. Butyrate Regulation of Distinct Macrophage Subsets: Opposing Effects on M1 and M2 Macrophages. Int. J. Probiotics Prebiotics 2011, 6, 147–158. [Google Scholar]
- Venegas, D.P.; De la Fuente, M.K.; Landskron, G.; González, M.J.; Quera, R.; Dijkstra, G.; Harmsen, H.J.M.; Faber, K.N.; Hermoso, M.A. Short Chain Fatty Acids (SCFAs)-Mediated Gut Epithelial and Immune Regulation and Its Relevance for Inflammatory Bowel Diseases. Front. Immunol. 2019, 10, 277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Macia, L.; Tan, J.; Vieira, A.T.; Leach, K.; Stanley, D.; Luong, S.; Maruya, M.; McKenzie, C.I.; Hijikata, A.; Wong, C.; et al. Metabolite-Sensing Receptors GPR43 and GPR109A Facilitate Dietary Fibre-Induced Gut Homeostasis Through Regulation of the Inflammasome. Nat. Commun. 2015, 6, 6734. [Google Scholar] [CrossRef] [Green Version]
- Chang, P.V.; Hao, L.; Offermanns, S.; Medzhitov, R. The Microbial Metabolite Butyrate Regulates Intestinal Macrophage Function via Histone Deacetylase Inhibition. Proc. Natl. Acad. Sci. USA 2014, 111, 2247–2252. [Google Scholar] [CrossRef] [PubMed]
- Luu, M.; Weigand, K.; Wedi, F.; Breidenbend, C.; Leister, H.; Pautz, S.; Adhikary, T.; Visekruna, A. Regulation of the Effector Function of CD8+ T Cells by Gut Microbiota-Derived Metabolite Butyrate. Sci. Rep. 2018, 8, 14430. [Google Scholar] [CrossRef] [Green Version]
- Azad, M.A.K.; Sarker, M.; Li, T.; Yin, J. Probiotic Species in the Modulation of Gut Microbiota: An Overview. BioMed Res. Int. 2018, 2018, 9478630. [Google Scholar] [CrossRef] [Green Version]
- Sundararaman, A.; Ray, M.; Ravindra, P.V.; Halami, P.M. Role of Probiotics to Combat Viral Infections with Emphasis on COVID-19. Appl. Microbiol. Biotechnol. 2020, 104, 8089–8104. [Google Scholar] [CrossRef] [PubMed]
- Ashraf, R.; Shah, N.P. Immune System Stimulation by Probiotic Microorganisms. Crit. Rev. Food Sci. Nutr. 2014, 54, 938–956. [Google Scholar] [CrossRef] [PubMed]
- Galdeano, C.M.; Cazorla, S.I.; Lemme Dumit, J.M.; Vélez, E.; Perdigón, G. Beneficial Effects of Probiotic Consumption on the Immune System. Ann. Nutr. Metab. 2019, 74, 115–124. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Hu, W.; Zhu, L.; Yang, Q. Bacillus subtilis and Surfactin Inhibit the Transmissible Gastroenteritis Virus from Entering the Intestinal Epithelial Cells. Biosci. Rep. 2017, 37, BSR20160557. [Google Scholar] [CrossRef] [Green Version]
- Li, D.; Breiman, A.; le Pendu, J.; Uyttendaele, M. Anti-viral Effect of Bifidobacterium adolescentis Against Noroviruses. Front. Microbiol. 2016, 7, 864. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guillemard, E.; Tondu, F.; Lacoin, F.; Schrezenmeir, J. Consumption of a Fermented Dairy Product Containing the Probiotic Lactobacillus casei DN-114001 Reduces the Duration of Respiratory Infections in the Elderly in a Randomised Controlled Trial. Br. J. Nutr. 2010, 103, 58–68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arimori, Y.; Nakamura, R.; Hirose, Y.; Murosaki, S.; Yamamoto, Y.; Shidara, O.; Ichikawa, H.; Yoshikai, Y. Daily Intake of Heat-Killed Lactobacillus plantarum L-137 Enhances Type I Interferon Production in Healthy Humans and Pigs. Immunopharmacol. Immunotoxicol. 2012, 34, 937–943. [Google Scholar] [CrossRef]
- Kawase, M.; He, F.; Kubota, A.; Harata, G.; Hiramatsu, M. Oral Administration of Lactobacilli from Human Intestinal Tract Protects Mice against Influenza Virus Infection. Lett. Appl. Microbiol. 2010, 51, 6–10. [Google Scholar] [CrossRef]
- Tomosada, Y.; Chiba, E.; Zelaya, H.; Takahashi, T.; Tsukida, K.; Kitazawa, H.; Alvarez, S.; Villena, J. Nasally Administered Lactobacillus rhamnosus Strains Differentially Modulate Respiratory Antiviral Immune Responses and Induce Protection Against Respiratory Syncytial Virus Infection. BMC Immunol. 2013, 14, 40. [Google Scholar] [CrossRef] [Green Version]
- Hori, T.; Kiyoshima, J.; Shida, K.; Yasui, H. Effect of Intranasal Administration of Lactobacillus casei Shirota on Influenza Virus Infection of Upper Respiratory Tract in Mice. Clin. Diagn. Lab. Immunol. 2001, 8, 593–597. [Google Scholar] [CrossRef] [Green Version]
- Waki, N.; Matsumoto, M.; Fukui, Y.; Suganuma, H. Effects of Probiotic Lactobacillus brevis KB290 on Incidence of Influenza Infection among Schoolchildren: An Open-Label Pilot Study. Lett. Appl. Microbiol. 2014, 59, 565–571. [Google Scholar] [CrossRef] [Green Version]
- Kiso, M.; Takano, R.; Sakabe, S.; Katsura, H.; Shinya, K.; Uraki, R.; Watanabe, S.; Saito, H.; Toba, M.; Kohda, N.; et al. Protective Efficacy of Orally Administered, Heat-Killed Lactobacillus pentosus b240 Against Influenza A Virus. Sci. Rep. 2013, 3, 1563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sugimura, T.; Jounai, K.; Ohshio, K.; Takana, T.; Suwa, M.; Fujiwara, D. Immunomodulatory Effect of Lactococcus lactis JCM5805 on Human Plasmacytoid Dendritic Cells. Clin. Immunol. 2013, 149, 509–518. [Google Scholar] [CrossRef] [PubMed]
- Sugimura, T.; Takahashi, H.; Jounai, K.; Ohshio, K.; Kanayama, M.; Tazumi, K.; Tanihata, Y.; Miura, Y.; Fujiwara, D.; Yamamoto, N. Effects of Oral Intake of Plasmacytoid Dendritic Cells-Stimulative Lactic Acid Bacterial Strain on Pathogenesis of Influenza-Like Illness and Immunological Response to Influenza Virus. Br. J. Nutr. 2015, 114, 727–733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shibata, T.; Kanayama, M.; Haida, M.; Fujimoto, S.; Oroguchi, T.; Sata, K.; Mita, N.; Kutsuzawa, T.; Ikeuchi, M.; Kondo, M.; et al. Lactococcus lactis JCM5805 Activates Anti-Viral Immunity and Reduces Symptoms of Common Cold and Influenza in Healthy Adults in a Randomised Controlled Trial. J. Funct. Foods 2016, 24, 492–500. [Google Scholar] [CrossRef]
- Suzuki, H.; Kanayama, M.; Fujii, T.; Fujiwara, D.; Sugimura, H. Effects of the Beverage Containing Lactococcus lactis subsp. lactis JCM5805 on Anti-Viral Immune Responses and Maintenance of Physical Conditions. Jpn. Pharmacol. Ther. 2015, 43, 106–111. [Google Scholar]
- Sakata, K.; Sasaki, Y.; Jounai, K.; Fujii, T.; Fujiwara, D. Preventive Effect of Lactococcus lactis subsp. lactis JCM5805 Yogurt Intake on Influenza Infection Among Schoolchildren. Health 2017, 9, 756–762. [Google Scholar] [CrossRef]
- Jounai, K.; Ikado, K.; Sugimura, T.; Ano, Y.; Braun, J.; Fujiwara, D. Spherical Lactic Acid Bacteria Activate Plasmacytoid Dendritic Cells Immunomodulatory Function via TLR9-Dependent Crosstalk with Myeloid Dendritic Cells. PLoS ONE 2012, 7, e32588. [Google Scholar] [CrossRef]
- van Puyenbroeck, K.; Hens, N.; Coenen, S.; Michiels, B.; Beunckens, C.; Molenberghs, G.; Van Royen, P.; Verhoeven, V. Efficacy of Daily Intake of Lactobacillus casei Shirota on Respiratory Symptoms and Influenza Vaccination Immune Response: A Randomised, Double-Blind, Placebo-Controlled Trial in Healthy Elderly Nursing Home Residents. Am. J. Clin. Nutr. 2012, 95, 1165–1171. [Google Scholar] [CrossRef] [Green Version]
- Makino, S.; Ikegami, S.; Kume, A.; Horiuchi, H.; Sasaki, H.; Orii, N. Reducing the risk of infection in the elderly by dietary intake of yogurt fermented with Lactobacillus delbrueckii ssp. bulgaricus OLL1073R-1. Br. J. Nutr. 2010, 104, 998–1006. [Google Scholar] [CrossRef] [Green Version]
- Nagai, T.; Makino, S.; Ikegami, S.; Itoh, H.; Yamada, H. Effects of Oral Administration of Yogurt Fermented with Lactobacillus delbrueckii ssp. bulgaricus OLL1073R-1 and its Exopolysaccharides Against Influenza Virus Infection in Mice. Int. Immunopharmacol. 2011, 11, 2246–2250. [Google Scholar] [CrossRef] [PubMed]
- Makino, S.; Sato, A.; Goto, A.; Nakamura, M.; Ogawa, M.; Chiba, Y.; Hemmi, J.; Kano, H.; Takeda, K.; Okumura, K.; et al. Enhanced Natural Killer Cell Activation by Exopolysaccharides Derived from Yogurt Fermented with Lactobacillus delbrueckii ssp. bulgaricus OLL1073R-1. J. Dairy Sci. 2016, 99, 915–923. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gleeson, M.; Bishop, N.C.; Struszczak, L. Effects of Lactobacillus casei Shirota Ingestion on Common Cold Infection and Herpes Virus Antibodies in Endurance Athletes: A Placebo-Controlled, Randomised Trial. Eur. J. Appl. Physiol. 2016, 116, 1555–1563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murosaki, S.; Yamamoto, Y.; Ito, K.; Inokuchi, T.; Kusaka, H.; Ikeda, H.; Yoshikai, Y. Heat-killed Lactobacillus plantarum L-137 Suppresses Naturally Fed Antigen-Specific IgE Production by Stimulation of IL-12 Production in Mice. J. Allergy Clin. Immunol. 1998, 102, 57–64. [Google Scholar] [CrossRef]
- Maeda, N.; Nakamura, R.; Hirose, Y.; Murosaki, S.; Yamamoto, Y.; Kase, T.; Yoshikai, Y. Oral Administration of Heat-Killed Lactobacillus plantarum L-137 Enhances Protection Against Influenza Virus Infection by Stimulation of Type I Interferon Production in Mice. Int. Immunopharmacol. 2009, 9, 1122–1125. [Google Scholar] [CrossRef]
- Kechaou, N.; Chain, F.; Gratadoux, J.J.; Blugeon, S.; Bertho, N.; Chevalier, C.; Le Goffic, R.; Courau, S.; Molimard, P.; Chatel, J.M.; et al. Identification of One Novel Candidate Probiotic Lactobacillus plantarum Strain Active Against Influenza Virus Infection in Mice by a Large-Scale Screening. Appl. Environ. Microbiol. 2013, 79, 1491–1499. [Google Scholar] [CrossRef] [Green Version]
- Kawashima, T.; Hayashi, K.; Kosaka, A.; Kawashima, M.; Igarashi, T.; Tsutsui, H.; Tsuji, N.M.; Nishimura, I.; Hayashi, T.; Obata, A. Lactobacillus plantarum strain YU from fermented foods activates Th1 and protective immune responses. Int. Immunopharmacol. 2011, 11, 2017–2024. [Google Scholar] [CrossRef]
- Chiba, Y.; Shida, K.; Nagata, S.; Wada, M.; Bian, L.; Wang, C.; Shimizu, T.; Yamashiro, Y.; Kiyoshima-Shibata, J.; Nanno, M.; et al. Well-Controlled Proinflammatory Cytokine Responses of Peyer’s Patch Cells to Probiotic Lactobacillus casei. Immunology 2010, 130, 352–362. [Google Scholar] [CrossRef]
- Hirose, Y.; Murosaki, S.; Yamamoto, Y.; Yoshikai, Y.; Tsuru, T. Daily Intake of Heat-Killed Lactobacillus plantarum L-137 Augments Acquired Immunity in Healthy Adults. J. Nutr. 2006, 136, 3069–3073. [Google Scholar] [CrossRef] [Green Version]
- Hirose, Y.; Yamamoto, Y.; Yoshikai, Y.; Murosaki, S. Oral Intake of Heat-Killed Lactobacillus plantarum L-137 Decreases the Incidence of Upper Respiratory Tract Infection in Healthy Subjects with High Levels of Psychological Stress. J. Nutr. Sci. 2013, 2, e39. [Google Scholar] [CrossRef] [Green Version]
- Yeo, J.M.; Lee, H.J.; Kim, J.W.; Lee, J.B.; Park, S.Y.; Choi, I.S.; Song, C.S. Lactobacillus fermentum CJL-112 Protects Mice Against Influenza Virus Infection by Activating T-helper 1 and Eliciting a Protective Immune Response. Int. Immunopharmacol. 2014, 18, 50–54. [Google Scholar] [CrossRef] [PubMed]
- Youn, H.N.; Lee, D.H.; Lee, Y.N.; Park, J.K.; Yuk, S.S.; Yang, S.Y.; Lee, H.J.; Woo, S.H.; Kim, H.M.; Lee, J.B.; et al. Intranasal Administration of Live Lactobacillus species Facilitates Protection Against Influenza Virus Infection in Mice. Antivir. Res. 2012, 93, 138–143. [Google Scholar] [CrossRef] [PubMed]
- Rizzardini, G.; Eskesen, D.; Calder, P.C.; Capetti, A.; Jespersen, L.; Clerici, M. Evaluation of the Immune Benefits of Two Probiotic Strains Bifidobacterium animalis ssp. lactis, BB-12® and Lactobacillus paracasei ssp. paracasei, L. casei 431® in an Influenza Vaccination Model: A Randomised, Double-Blind, Placebo-Controlled Study. Br. J. Nutr. 2012, 107, 876–884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borthakur, A.; Anbazhagan, A.N.; Kumar, A.; Raheja, G.; Singh, V.; Ramaswamy, K.; Dudeja, P.K. The Probiotic Lactobacillus Plantarum Counteracts TNF-α-Induced Downregulation of SMCT1 Expression and Function. Am. J. Physiol. Gastrointest. Liver Physiol. 2010, 299, G928–G934. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zelaya, H.; Alvarez, S.; Kitazawa, H.; Villena, J. Respiratory Antiviral Immunity and Immunobiotics: Beneficial Effects on Inflammation-Coagulation Interaction during Influenza Virus Infection. Front. Immunol. 2016, 7, 633. [Google Scholar] [CrossRef] [Green Version]
- Kourelis, A.; Zinonos, I.; Kakagianni, M.; Christidou, A.; Christoglou, N.; Yiannaki, E.; Testa, T.; Kotzamanidis, C.; Litopoulou-Tzanetaki, E.; Tzanetakis, N.; et al. Validation of the Dorsal Air Pouch Model to Predict and Examine Immunostimulatory Responses in the Gut. J. Appl. Microbiol. 2010, 108, 274–284. [Google Scholar] [CrossRef]
- Fu, L.; Wang, B.; Yuan, T.; Chen, X.; Ao, Y.; Fitzpatrick, T.; Li, P.; Zhou, Y.; Lin, Y.F.; Duan, Q.; et al. Clinical Characteristics of Coronavirus Disease 2019 (COVID-19) in China: A Systematic Review and Meta-Analysis. J. Infect. 2020, 80, 656–665. [Google Scholar] [CrossRef]
- Zhong, P.; Xu, J.; Yang, D.; Shen, Y.; Wang, L.; Feng, Y.; Du, C.; Song, Y.; Wu, C.; Hu, X.; et al. COVID-19-Associated Gastrointestinal and Liver Injury: Clinical Features and Potential Mechanisms. Signal Transduct. Target. Ther. 2020, 5, 256. [Google Scholar] [CrossRef]
- Zhou, P.; Yang, X.L.; Wang, X.G.; Hu, B.; Zhang, L.; Zhang, W.; Si, H.R.; Zhu, Y.; Li, B.; Huang, C.L.; et al. A Pneumonia Outbreak associated with a New Coronavirus of Probable Bat Origin. Nature 2020, 579, 270–273. [Google Scholar] [CrossRef] [Green Version]
- Wu, F.; Zhao, S.; Yu, B.; Chen, Y.M.; Wang, W.; Song, Z.G.; Hu, Y.; Tao, Z.W.; Tian, J.H.; Pei, Y.Y.; et al. A New Coronavirus Associated with Human Respiratory Disease in China. Nature 2020, 579, 265–269. [Google Scholar] [CrossRef] [Green Version]
- D’Amico, F.; Baumgart, D.C.; Danese, S.; Peyrin-Biroulet, L. Diarrhea during COVID-19 Infection: Pathogenesis, Epidemiology, Prevention, and Management. Clin. Gastroenterol. Hepatol. 2020, 18, 1663–1672. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, M.; Kleine-Weber, H.; Schroeder, S.; Krüger, N.; Herrler, T.; Erichsen, S.; Schiergens, T.S.; Herrler, G.; Wu, N.H.; Nitsche, A.; et al. SARS-CoV-2 Cell Entry Depends on ACE2 and TMPRSS2 and Is Blocked by a Clinically Proven Protease Inhibitor. Cell 2020, 181, 271–280.e8. [Google Scholar] [CrossRef]
- Wang, K.; Chen, W.; Zhang, Z.; Deng, Y.; Lian, J.-Q.; Du, P.; Wei, D.; Zhang, Y.; Sun, X.-X.; Gong, L.; et al. CD147-spike protein is a novel route for SARS-CoV-2 infection to host cells. Signal Transduct. Target. Ther. 2020, 5, 283. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Li, H.B.; Lyu, J.R.; Lei, X.M.; Li, W.; Wu, G.; Lyu, J.; Dai, Z.M. Specific ACE2 Expression in Small Intestinal Enterocytes May Cause Gastrointestinal Symptoms and Injury After 2019-nCoV Infection. Int. J. Infect. Dis. 2020, 96, 19–24. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Hu, B.; Hu, C.; Zhu, F.; Liu, X.; Zhang, J.; Wang, B.; Xiang, H.; Cheng, Z.; Xiong, Y.; et al. Clinical Characteristics of 138 Hospitalized Patients With 2019 Novel Coronavirus-Infected Pneumonia in Wuhan, China. JAMA 2020, 323, 1061–1069. [Google Scholar] [CrossRef]
- Li, J.; Guo, M.; Tian, X.; Wang, X.; Yang, X.; Wu, P.; Liu, C.; Xiao, Z.; Qu, Y.; Yin, Y.; et al. Virus-Host Interactome and Proteomic Survey Reveal Potential Virulence Factors Influencing SARS-CoV-2 Pathogenesis. Medicine 2021, 2, 99–112.e7. [Google Scholar] [CrossRef] [PubMed]
- Ren, X.; Glende, J.; Al-Falah, M.; de Vries, V.; Schwegmann-Wessels, C.; Qu, X.; Tan, L.; Tschernig, T.; Deng, H.; Naim, H.Y.; et al. Analysis of ACE2 in Polarized Epithelial Cells: Surface Expression and Function as Receptor for Severe Acute Respiratory Syndrome-Associated Coronavirus. J. Gen. Virol. 2006, 87 Pt 6, 1691–1695. [Google Scholar] [CrossRef]
- He, J.; Tao, H.; Yan, Y.; Huang, S.Y.; Xiao, Y. Molecular Mechanisms of Evolution and Human Infection with SARS-CoV-2. Viruses 2020, 12, 428. [Google Scholar] [CrossRef] [Green Version]
- Subbarao, K.; Mahanty, S. Respiratory Virus Infections: Understanding COVID-19. Immunity 2020, 52, 905–909. [Google Scholar] [CrossRef]
- Bhattacharyya, P.; Das, S.; Aich, S.; Sarkar, J. COVID-19: Morphology and Mechanism of the SARS-CoV-2, Global Outbreak, Medication, Vaccines and Future of the Virus. Front. Biosci. 2021, 13, 272–290. [Google Scholar] [CrossRef]
- Zeppa, S.D.; Agostini, D.; Piccoli, G.; Stocchi, V.; Sestili, P. Gut Microbiota Status in COVID-19: An Unrecognised Player? Front. Cell. Infect. Microbiol. 2020, 10, 576551. [Google Scholar] [CrossRef]
- Tripathi, S.; Tecle, T.; Verma, A.; Crouch, E.; White, M.; Hartshorn, K.L. The Human Cathelicidin LL-37 Inhibits Influenza A Viruses through a Mechanism Distinct from that of Surfactant Protein D or Defensins. J. Gen. Virol. 2013, 94 Pt 1, 40–49. [Google Scholar] [CrossRef]
- Pinkenburg, O.; Meyer, T.; Bannert, N.; Norley, S.; Bolte, K.; Czudai-Matwich, V.; Herold, S.; Gessner, A.; Schnare, M. The Human Antimicrobial Protein Bactericidal/Permeability-Increasing Protein (BPI) Inhibits the Infectivity of Influenza A Virus. PLoS ONE 2016, 11, e0156929. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, H.; Zhou, J.; Zhang, K.; Chu, H.; Liu, D.; Poon, V.K.M.; Chan, C.C.-S.; Leung, H.-C.; Fai, N.; Lin, Y.-P.; et al. A novel peptide with potent and broad-spectrum antiviral activities against multiple respiratory viruses. Sci. Rep. 2016, 6, 22008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wohlford-Lenane, C.L.; Meyerholz, D.K.; Perlman, S.; Zhou, H.; Tran, D.; Selsted, M.E.; McCray, P.B. Rhesus theta-Defensin prevents death in a mouse model of severe acute respiratory syndrome Coronavirus pulmonary disease. J. Virol. 2009, 83, 11385–11390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niv, Y. Defensin 5 for Protection of SARS-CoV-2 Invasion and COVID-19 Disease. Med. Hypotheses 2020, 143, 110244. [Google Scholar] [CrossRef]
- Astuti, I.; Ysrafil. Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2): An Overview of Viral Structure and Host Response. Diabetes Metab. Syndr. 2020, 14, 407–412. [Google Scholar] [CrossRef]
- Song, P.; Li, W.; Xie, J.; Hou, Y.; You, C. Cytokine Storm Induced by SARS-CoV-2. Clin. Chim. Acta 2020, 509, 280–287. [Google Scholar] [CrossRef]
- Choudhury, A.; Muckherjee, S. In Silico Studies on the Comparative Characterisation of the Interactions of SARS-CoV-2 Spike Glycoprotein with ACE-2 Receptor Homologs and Human TLRs. J. Med. Virol. 2020, 92, 2105–2113. [Google Scholar] [CrossRef]
- Choudhury, A.; Das, N.C.; Patra, R.; Mukherjee, S. In Silico Analyses of the Comparative Sensing of SARS-CoV-2 mRNA by the Intracellular TLRs of Humans. J. Med. Virol. 2021, 93, 2476–2486. [Google Scholar] [CrossRef]
- Gorden, K.B.; Gorski, K.S.; Gibson, S.J.; Kedl, R.M.; Keiper, W.C.; Qui, X.; Tomai, M.A.; Alkan, S.S.; Vasilakos, J.P. Synthetic TLR Agonists Reveal Functional Differences between Human TLR7 and TLR8. J. Immunol. 2005, 174, 1259–1268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, S.; Jiang, Z.Y.; Sun, Y.F.; Yu, B.; Chen, J.; Dai, C.Q.; Wu, X.L.; Tang, X.L.; Chen, X.Y. Microbiota Regulates the TLR7 Signaling Pathway Against Respiratory Tract Influenza A Virus Infection. Curr. Microbiol. 2013, 67, 414–422. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Zhao, S.; Teng, T.; Abdalla, A.E.; Zhu, W.; Xie, L.; Wang, Y.; Guo, X. Systematic Comparison of Two Animal-to-Human Transmitted Human Coronaviruses: SARS-CoV-2 and SARS-CoV. Viruses 2020, 12, 244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, N.; Zhou, M.; Dong, X.; Qu, J.; Gong, F.; Han, Y.; Qui, Y.; Wang, J.; Liu, Y.; Wei, Y.; et al. Epidemiological and Clinical Characteristics of 99 Cases of 2019 Novel Coronavirus Pneumonia in Wuhan, China: A Descriptive Study. Lancet 2020, 395, 507–513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siddiqi, H.K.; Mehra, M.R. COVID-19 Illness in Native and Immunosuppressed States: A Clinical-Therapeutic Staging Proposal. J. Heart Lung Transplant. 2020, 39, 405–407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dalskov, L.; Møhlenberg, M.; Thyrsted, J.; Blay-Cadanet, J.; Poulsen, E.T.; Folkersen, B.H.; Skaarup, S.H.; Olagnier, D.; Reinert, L.; Enghild, J.J.; et al. SARS-CoV-2 Evades Immune Detection in Alveolar Macrophages. EMBO Rep. 2020, 21, e51252. [Google Scholar] [CrossRef]
- Mordstein, M.; Neugebauer, E.; Ditt, V.; Jessen, B.; Rieger, T.; Falcone, V.; Sorgeloos, F.; Ehl, S.; Mayer, D.; Kochs, G.; et al. Lambda Interferon Renders Epithelial Cells of the Respiratory and Gastrointestinal Tracts Resistant to Viral Infections. J. Virol. 2010, 84, 5670–5677. [Google Scholar] [CrossRef] [Green Version]
- Banerjee, S.; Chakrabarti, A.; Jha, B.K.; Weiss, S.R.; Silverman, R.H. Cell-Type-Specific Effects of RNaseL on Viral Induction of Beta-Interferon. mBio 2014, 5, e00856-14. [Google Scholar] [CrossRef] [Green Version]
- Stegemann-Koniszewski, S.; Behrens, S.; Boehme, J.D.; Hochnadel, I.; Riese, P.; Guzmán, C.A.; Kröger, A.; Schreiber, J.; Gunzer, M.; Bruder, D. Respiratory Influenza A Virus Infection Triggers Local and Systemic Natural Killer Cell Activation via Toll-Like Receptor 7. Front. Immunol. 2018, 9, 245. [Google Scholar] [CrossRef] [Green Version]
- Hwang, I.; Scott, J.M.; Kakarla, T.; Duriancik, D.M.; Choi, S.; Cho, C.; Lee, T.; Park, H.; French, A.R.; Beli, E.; et al. Activation Mechanisms of Natural Killer Cells during Influenza Virus Infection. PLoS ONE 2012, 7, e51858. [Google Scholar] [CrossRef] [Green Version]
- Bortolotti, D.; Gentili, V.; Rizzo, S.; Rotola, A.; Rizzo, R. SARS-CoV-2 Spike 1 Protein Controls Natural Killer Cell Activation via the HLA-E/NKG2A Pathway. Cells 2020, 9, 1975. [Google Scholar] [CrossRef] [PubMed]
- To, E.E.; Broughton, B.R.S.; Hendricks, K.S.; Vlahos, R.; Selemidis, S. Influenza A Virus and TLR7 Activation Potentiate NOX2 Oxidase-Dependent ROS Production in Macrophages. Free Radic. Res. 2014, 48, 940–947. [Google Scholar] [CrossRef] [PubMed]
- Jamaluddin, M.; Tian, B.; Boldogh, I.; Garofalo, R.P.; Brasier, A.R. Respiratory Syncytial Virus Infection Induces a Reactive Oxygen Species-MSK1-Phospho-Ser-276 RelA Pathway Required for Cytokine Expression. J. Virol. 2009, 83, 10605–10615. [Google Scholar] [CrossRef] [Green Version]
- Tincati, C.; Cannizzo, E.S.; Giacomelli, M.; Badolato, R.; d’Arminio Monforte, A.; Marchetti, G. Heightened Circulating Interferon-Inducible Chemokines, and Activated Pro-Cytolytic Th1-Cell Phenotype Features COVID-19 Aggravation in the Second Week of Illness. Front. Immunol. 2020, 11, 580987. [Google Scholar] [CrossRef] [PubMed]
- Carvelli, J.; Demaria, O.; Vély, F.; Batista, L.; Chouaki Benmansour, N.; Fares, J.; Carpentier, S.; Thibult, M.L.; Morel, A.; Remark, R.; et al. Association of COVID-19 Inflammation with Activation of the C5a-C5aR1 Axis. Nature 2020, 588, 146–150. [Google Scholar] [CrossRef] [PubMed]
- Ong, E.Z.; Chan, Y.F.Z.; Leong, W.Y.; Lee, N.M.Y.; Kalimuddin, S.; Mohideen, H.S.M.; Chan, K.S.; Tan, A.T.; Bertoletti, A.; Ooi, E.E.; et al. A Dynamic Immunee Response Shapes COVID-19 Progression. Cell Host Microbe 2020, 27, 879–882. [Google Scholar] [CrossRef]
- Mahmoud, A.B.; Tu, M.M.; Wight, A.; Zein, H.S.; Rahim, M.M.; Lee, S.H.; Sekhon, H.S.; Brown, E.G.; Makrigiannis, A.P. Correction: Influenza Virus Targets Class I MHC-Educated NK Cells for Immunoevasion. PLoS Pathog. 2016, 12, e1005446. [Google Scholar] [CrossRef]
- Rahim, M.M.A.; Parsons, B.D.; Price, E.L.; Slaine, P.D.; Chilvers, B.L.; Seaton, G.S.; Wight, A.; Medina-Luna, D.; Dey, S.; Grandy, S.L.; et al. Defective Influenza A Virus RNA Products Mediate MAVS-Dependent Upregulation of Human Leukocyte Antigen ClassIProteins. J. Virol. 2020, 94, e00165–e001620. [Google Scholar] [CrossRef] [Green Version]
- Sekine, T.; Perez-Potti, A.; Rivera-Ballesteros, O.; Strålin, K.; Gorin, J.B.; Olsson, A.; Llewellyn-Lacey, S.; Kamal, H.; Bogdanovic, G.; Muschiol, S.; et al. Robust T Cell Immunity in Convalescent Individuals with Asymptomatic or Mild COVID-19. Cell 2020, 183, 158–168. [Google Scholar] [CrossRef]
- Chein, J.Y.; Hsueh, P.R.; Cheng, W.C.; Yu, C.J.; Yang, P.C. Temporal Changes in Cytokine/Chemokine Profiles and Pulmonary Involvement in Severe Acute Respiratory Syndrome. Respirology 2006, 11, 715–722. [Google Scholar] [CrossRef] [Green Version]
- Chi, Y.; Ge, Y.; Wu, B.; Zhang, W.; Wu, T.; Wen, T.; Liu, J.; Guo, X.; Huang, C.; Jiao, Y.; et al. Serum Cytokine and Chemokine Profile in Relation to the Severity of Coronavirus Disease 2019 in China. J. Infect. Dis. 2020, 222, 746–754. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Wang, Y.; Li, X.; Ren, L.; Zhao, J.; Hu, Y.; Zhang, L.; Fan, G.; Xu, J.; Gu, X.; et al. Clinical Features of Patients Infected with 2019 Novel Coronavirus in Wuhan, China. Lancet 2020, 395, 497–506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Long, Q.X.; Liu, B.Z.; Deng, H.J.; Wu, G.C.; Deng, K.; Chen, Y.K.; Liao, P.; Qiu, F.; Lin, Y.; Cai, X.F.; et al. Antibody Responses to SARS-CoV-2 in Patients with COVID-19. Nat. Med. 2020, 26, 845–848. [Google Scholar] [CrossRef] [PubMed]
- Cervia, C.; Nilsson, J.; Zurbuchen, Y.; Valaperti, A.; Schreiner, J.; Wolfensberger, A.; Raeber, M.E.; Adamo, S.; Weigang, S.; Emmenegger, M.; et al. Systemic and Mucosal Antibody Responses Specific to SARS-CoV-2 During Mild versus Severe COVID-19. J. Allergy Clin. Immunol. 2021, 147, 545–557. [Google Scholar] [CrossRef]
- Azkur, A.K.; Akdis, M.; Azkur, D.; Sokolowska, M.; van de Veen, W.; Brüggen, M.C.; O’Mahony, L.; Gao, Y.; Nadeau, K.; Akdis, C.A. Immune Response to SARS-CoV-2 and Mechanisms of Immunopathological Changes in COVID-19. Allergy 2020, 75, 1564–1581. [Google Scholar] [CrossRef]
- Tso, F.Y.; Lidenge, S.J.; Poppe, L.K.; Peña, P.B.; Privatt, S.R.; Bennett, S.J.; Ngowi, J.R.; Mwaiselage, J.; Belshan, M.; Siedlik, J.A.; et al. Presence of Antibody-Dependent Cellular Cytotoxicity (ADCC) Against SARS-CoV-2 in COVID-19 Plasma. PLoS ONE 2021, 16, e0247640. [Google Scholar] [CrossRef]
- Chen, X.; Rostad, C.A.; Anderson, L.J.; Sun, H.Y.; Lapp, S.A.; Stephens, K.; Hussaini, L.; Gibson, T.; Rouphael, N.; Anderson, E.J. The Development and Kinetics of Functional Antibody-Dependent Cell-Mediated Cytotoxicity (ADCC) to SARS-CoV-2 Spike Protein. Virology 2021, 559, 1–9. [Google Scholar] [CrossRef]
- Garcia, L.F. Immune response, inflammation, and the clinical spectrum of COVID-19. Front. Immunol. 2020, 11, 1441. [Google Scholar] [CrossRef]
- Turner, M.D.; Nedjai, B.; Hurst, T.; Pennington, D.J. Cytokines and Chemokines: At the Crossroads of Cell Signalling and Inflammatory Disease. Biochim. Biophys. Acta 2014, 1843, 2563–2582. [Google Scholar] [CrossRef] [Green Version]
- Bouayad, A. Innate Immune Evasion by SARS-CoV-2: Comparison with SARS-CoV. Rev. Med. Virol. 2020, 30, 1–9. [Google Scholar] [CrossRef]
- Hu, Y.; Li, W.; Gao, T.; Cui, Y.; Jin, Y.; Li, P.; Ma, Q.; Liu, X.; Cao, C. The Severe Acute Respiratory Syndrome Coronavirus Nucleocapsid Inhibits Type I Interferon Production by Interfering with TRIM25-Mediated RIG-I Ubiquitination. J. Virol. 2017, 91, e02143-16. [Google Scholar] [CrossRef] [Green Version]
- Oh, S.J.; Shin, O.S. SARS-CoV-2 Nucleocapsid Protein Targets RIG-I-Like Receptor Pathways to Inhibit the Induction of Interferon Response. Cells 2021, 10, 530. [Google Scholar] [CrossRef]
- Opitz, B.; Rejaibi, A.; Dauber, B.; Eckhard, J.; Vinzing, M.; Schmeck, B.; Hippenstiel, S.; Suttorp, N.; Wolff, T. IFNbeta Induction by Influenza A Virus is Mediated by RIG-I which is Regulated by the Viral NS1 Protein. Cell. Microbiol. 2007, 9, 930–938. [Google Scholar] [CrossRef] [PubMed]
- Konno, Y.; Kimura, I.; Uriu, K.; Fukushi, M.; Irie, T.; Koyanagi, Y.; Sauter, D.; Gifford, R.J.; USFQ-COVID19 Consortium; Nakagawa, S.; et al. SARS-CoV-2 ORF3b Is a Potent Interferon Antagonist Whose Activity Is Increased by a Naturally Occurring Elongation Variant. Cell Rep. 2020, 32, 108185. [Google Scholar] [CrossRef]
- Narayanan, K.; Huang, C.; Lokugamage, K.; Kamitani, W.; Ikegami, T.; Tseng, C.T.; Makino, S. Severe Acute Respiratory Syndrome Coronavirus nsp1 Suppresses Host Gene Expression, Including That of Type I Interferon, in Infected Cells. J. Virol. 2008, 82, 4471–4479. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Chu, H.; Li, C.; Wong, B.H.; Cheng, Z.S.; Poon, V.K.; Sun, T.; Lau, C.C.; Wong, K.K.; Chan, J.Y.; et al. Active Replication of Middle East Respiratory Syndrome Coronavirus and Aberrant Induction of Inflammatory Cytokines and Chemokines in Human Macrophages: Implications for Pathogenesis. J. Infect. Dis. 2014, 209, 1331–1342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, M.; Gao, Y.; Wang, G.; Song, G.; Liu, S.; Sun, D.; Xu, Y.; Tian, Z. Functional Exhaustion of Antiviral Lymphocytes in COVID-19 Patients. Cell. Mol. Immunol. 2020, 17, 533–535. [Google Scholar] [CrossRef] [Green Version]
- Wilk, A.J.; Rustagi, A.; Zhao, N.Q.; Roque, J.; Martínez-Colón, G.J.; McKechnie, J.L.; Ivison, G.T.; Ranganath, T.; Vergara, R.; Hollis, T.; et al. A Single-Cell Atlas of the Peripheral Immune Response in Patients with Severe COVID-19. Nat. Med. 2020, 26, 1070–1076. [Google Scholar] [CrossRef] [PubMed]
- Diao, B.; Wang, C.; Tan, Y.; Chen, X.; Liu, Y.; Ning, L.; Chen, L.; Li, M.; Liu, Y.; Wang, G.; et al. Reduction and Functional Exhaustion of T Cells in Patients with Coronavirus Disease 2019 (COVID-19). Front. Immunol. 2020, 11, 827. [Google Scholar] [CrossRef]
- Gil-Etayo, F.J.; Suàrez-Fernández, P.; Cabrera-Marante, O.; Arroyo, D.; Garcinuño, S.; Naranjo, L.; Pleguezuelo, D.E.; Allende, L.M.; Mancebo, E.; Lalueza, A.; et al. T-Helper Cell Subset Response Is a Determining Factor in COVID-19 Progression. Front. Cell. Infect. Microbiol. 2021, 11, 624483. [Google Scholar] [CrossRef]
- Rosa, A.; Pye, V.E.; Graham, C.; Muir, L.; Seow, J.; Ng, K.W.; Cook, N.J.; Rees-Spear, C.; Parker, E.; Dos Santos, M.S.; et al. SARS-CoV-2 Can Recruit a Haem Metabolite to Evade Antibody Immunity. Sci. Adv. 2021, 7, eabg7607. [Google Scholar] [CrossRef]
- Andreano, E.; Piccini, G.; Licastro, D.; Casalino, L.; Johnson, N.V.; Paciello, I.; Dal Monego, S.; Pantano, E.; Manganaro, N.; Manenti, A.; et al. SARS-CoV-2 Escape from a Highly Neutralizing COVID-19 Convalescent Plasma. Proc. Natl. Acad. Sci. USA 2021, 18, e2103154118. [Google Scholar] [CrossRef]
- Yuan, Y.; Cao, D.; Zhang, Y.; Ma, J.; Qi, J.; Wang, Q.; Lu, G.; Wu, Y.; Yan, J.; Shi, Y.; et al. Cryo-EM Structures of MERS-CoV and SARS-CoV Spike Glycoproteins Reveal the Dynamic Receptor Binding Domains. Nat. Commun. 2017, 8, 15092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, S.; Chen, Y.; Wu, Z.; Chen, Y.; Gao, H.; Lv, L.; Guo, F.; Zhang, X.; Luo, R.; Huang, C.; et al. Alterations of the gut microbiota in patients with COVID-19 or H1N1 influenza. Clin. Infect. Dis. 2020, 71, 2669–2678. [Google Scholar] [CrossRef]
- Rueca, M.; Fontana, A.; Bartolini, B.; Piselli, P.; Mazzarelli, A.; Copetti, M.; Binda, E.; Perri, F.; Gruber, C.E.M.; Nicastri, E.; et al. Investigation of nasal/oropharyngeal microbial community of COVID-19 patients by 16S rDNA sequencing. Int. J. Environ. Res. Public Health 2021, 18, 2174. [Google Scholar] [CrossRef]
- Criado, P.R.; Abdalla, B.M.Z.; de Assis, I.C.; van Blarcum de Graaff Mello, C.; Caputo, G.C.; Vieira, I.C. Are the Cutaneous Manifestations During or Due to SARS-CoV-2 Infection/COVID-19 Frequent or Not? Revision of Possible Pathophysiologic Mechanisms. Inflamm. Res. 2020, 69, 745–756. [Google Scholar] [CrossRef] [PubMed]
- Paliwal, V.K.; Garg, R.K.; Gupta, A.; Tejan, N. Neuromuscular Presentations in Patients with COVID-19. Neurol. Sci. 2020, 41, 3039–3056. [Google Scholar] [CrossRef]
- Lozano-Sepulveda, S.A.; Galan-Huerta, K.; Martínez-Acuña, N.; Arellanos-Soto, D.; Rivas-Estilla, A.M. SARS-CoV-2 Another Kind of Liver Aggressor, How Does it Do That? Ann. Hepatol. 2020, 19, 592–596. [Google Scholar] [CrossRef] [PubMed]
- Achua, J.K.; Chu, K.Y.; Ibrahim, E.; Khodamoradi, K.; Delma, K.S.; Lakymenko, O.A.; Kryvenko, O.N.; Arora, H.; Ramasamy, R. Histopathology and Ultrastructural Findings of Fatal COVID-19 Infections on Testis. World J. Men’s Health 2021, 39, 65–74. [Google Scholar] [CrossRef]
- Iwasaki, M.; Saito, J.; Zhao, H.; Sakamoto, A.; Hirota, K.; Ma, D. Inflammation Triggered by SARS-CoV-2 and ACE2 Augment Drives Multiple Organ Failure of Severe COVID-19: Molecular Mechanisms and Implications. Inflammation 2021, 44, 13–34. [Google Scholar] [CrossRef] [PubMed]
- Saponaro, F.; Rutigliano, G.; Sestito, S.; Bandini, L.; Storti, B.; Bizzarri, R.; Zucchi, R. ACE2 in the Era of SARS-CoV-2: Controversies and Novel Perspectives. Front. Mol. Biosci. 2020, 7, 588618. [Google Scholar] [CrossRef] [PubMed]
- Tang, L.; Yin, Z.; Hu, Y.; Mei, H. Controlling Cytokine Storm Is Vital in COVID-19. Front. Immunol. 2020, 11, 570993. [Google Scholar] [CrossRef]
- Li, M.Y.; Li, L.; Zhang, Y.; Wang, X.S. Expression of the SARS-CoV-2 Cell Receptor Gene ACE2 in a Wide Variety of Human Tissues. Infect. Dis. Poverty 2020, 9, 45. [Google Scholar] [CrossRef] [PubMed]
- Fara, A.; Mitrev, Z.; Rosalia, R.A.; Assas, B.M. Cytokine Storm and COVID-19: A Chronicle of Pro-Inflammatory Cytokines. Open Biol. 2020, 10, 200160. [Google Scholar] [CrossRef]
- Wan, S.; Yi, Q.; Fan, S.; Lv, J.; Zhang, X.; Guo, L.; Lang, C.; Xiao, Q.; Xiao, K.; Yi, Z.; et al. Characteristics of Lymphocyte Subsets and Cytokines in Peripheral Blood of 123 Hospitalized Patients with 2019 Novel Coronavirus Pneumonia (NCP). MedRxiv 2020, 19, 57. [Google Scholar] [CrossRef] [Green Version]
- Zuo, T.; Zhang, F.; Lui, G.C.Y.; Yeoh, Y.K.; Li, A.Y.L.; Zhan, H.; Wan, Y.; Chung, A.C.K.; Cheung, C.P.; Chen, N.; et al. Alterations in Gut Microbiota of Patients With COVID-19 During Time of Hospitalization. Gastroenterology 2020, 159, 944–955. [Google Scholar] [CrossRef] [PubMed]
- Patra, S.; Saxena, S.; Sahu, N.; Pradhan, B.; Roychowdhury, A. Systematic Network and Meta-analysis on the Antiviral Mechanisms of Probiotics: A Preventive and Treatment Strategy to Mitigate SARS-CoV-2 Infection. Probiotics Antimicrob. Proteins 2021, 13, 1138–1156. [Google Scholar] [CrossRef]
- Liu, Z.; Li, N.; Fang, H.; Chen, X.; Guo, Y.; Gong, S.; Niu, M.; Zhou, H.; Jiang, Y.; Chang, P.; et al. Enteric dysbiosis is associated with sepsis in patients. FASEB J. 2019, 33, 12299–12310. [Google Scholar] [CrossRef] [Green Version]
- de Oliveira, G.L.V.; Leite, A.Z.; Higuchi, B.S.; Gonzaga, M.I.; Mariano, V.S. Intestinal Dysbiosis and Probiotic Applications in Autoimmune Diseases. Immunology 2017, 152, 1–12. [Google Scholar] [CrossRef]
- Zamani, B.; Golkar, H.R.; Farshbaf, S.; Emadi-Baygi, M.; Tajabadi-Ebrahimi, M.; Jafari, P.; Akhavan, R.; Taghizadeh, M.; Memarzadeh, M.R.; Asemi, Z. Clinical and Metabolic Response to Probiotic Supplementation in Patients with Rheumatoid Arthritis: A Randomized, Double-Blind, Placebo-Controlled Trial. Int. J. Rheum. Dis. 2016, 19, 869–879. [Google Scholar] [CrossRef]
- Alipour, B.; Homayouni-Rad, A.; Vaghef-Mehrabany, E.; Sharif, S.K.; Vaghef-Mehrabany, L.; Asghari-Jafarabadi, M.; Nakhjavani, M.R.; Mohtadi-Nia, J. Effects of Lactobacillus casei Supplementation on Disease Activity and Inflammatory Cytokines in Rheumatoid Arthritis Patients: A Randomized Double-Blind Clinical Trial. Int. J. Rheum. Dis. 2014, 17, 519–527. [Google Scholar] [CrossRef]
- Jin, X.; Lian, J.S.; Hu, J.H.; Gao, J.; Zheng, L.; Zhang, Y.M.; Hao, S.R.; Jia, H.Y.; Cai, H.; Zhang, X.L.; et al. Epidemiological, Clinical and Virological Characteristics of 74 Cases of Coronavirus-Infected Disease 2019 (COVID-19) with Gastrointestinal Symptoms. Gut 2020, 69, 1002–1009. [Google Scholar] [CrossRef] [Green Version]
- Brown, R.L.; Sequeira, R.P.; Clarke, T.B. The Microbiota Protects Against Respiratory Infection via GM-CSF Signaling. Nat. Commun. 2017, 8, 1512. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Yeh, C.; Jin, Z.; Ding, L.; Liu, B.Y.; Zhang, L.; Dannelly, H.K. Prospective Study of Probiotic Supplementation Results in Immune Stimulation and Improvement of Upper Respiratory Infection Rate. Synth. Syst. Biotechnol. 2018, 3, 113–120. [Google Scholar] [CrossRef] [PubMed]
- Ye, Q.; Wang, B.; Zhang, T.; Xu, J.; Shang, S. The Mechanism and Treatment of Gastrointestinal Symptoms in Patients with COVID-19. Am. J. Physiol. Gastrointest. Liver Physiol. 2020, 319, G245–G252. [Google Scholar] [CrossRef]
- Crowe, C.R.; Chen, K.; Pociask, D.A.; Alcorn, J.F.; Krivich, C.; Enelow, R.I.; Ross, T.M.; Witztum, J.L.; Kolls, J.K. Critical Role of IL-17RA in Immunopathology of Influenza Infection. J. Immunol. 2009, 183, 5301–5310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, F.; Yu, T.; Du, R.; Fan, G.; Liu, Y.; Liu, Z.; Xiang, J.; Wang, Y.; Song, B.; Gu, X.; et al. Clinical Course and Risk Factors for Mortality of Adult Inpatients with COVID-19 in Wuhan, China: A Retrospective Cohort Study. Lancet 2020, 395, 1054–1062. [Google Scholar] [CrossRef] [PubMed]
- National Health Commission; National Administration of Traditional Chinese Medicine. Diagnosis and Treatment Protocol for Novel Coronavirus Pneumonia (Trial Version 7). Chin. Med. J. 2020, 133, 1087–1095. [Google Scholar] [CrossRef]
- Hallersund, P.; Elfvin, A.; Helander, H.F.; Fandriks, L. The expression of renin-angiotensin system components in the human gastric mucosa. J. Renin-Angiotensin-Aldosterone Syst. 2011, 12, 54–64. [Google Scholar] [CrossRef]
- Hamming, I.; Timens, W.; Bulthuis, M.L.; Lely, A.T.; Navis, G.; van Goor, H. Tissue Distribution of ACE2 Protein, the Functional Receptor for SARS Coronavirus. A First Step in Understanding SARS Pathogenesis. J. Pathol. 2004, 203, 631–637. [Google Scholar] [CrossRef]
- Wang, X.-L.; Iwanami, J.; Min, L.-J.; Tsukuda, K.; Nakaoka, H.; Bai, H.-Y.; Shan, B.-S.; Kan-no, H.; Kukida, M.; Chisaka, T.; et al. Deficiency of angiotensin-converting enzyme 2 causes deterioration of cognitive function. NPJ Aging Mech. Dis. 2016, 2, 16024. [Google Scholar] [CrossRef] [Green Version]
- Verma, A.; Xu, K.; Du, T.; Zhu, P.; Liang, Z.; Liao, S.; Zhang, J.; Raizada, M.K.; Grant, M.B.; Li, Q. Expression of Human ACE2 in Lactobacillus and Beneficial Effects in Diabetic Retinopathy in Mice. Mol. Ther. Methods Clin. Dev. 2019, 14, 161–170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pasquereau, S.; Kumar, A.; Herbein, G. Targeting TNF and TNF Receptor Pathway in HIV-1 Infection: From Immune Activation to Viral Reservoirs. Viruses 2017, 9, 64. [Google Scholar] [CrossRef] [PubMed]
- Feldmann, M.; Maini, R.N.; Woody, J.N.; Holgate, S.T.; Winter, G.; Rowland, M.; Richards, D.; Hussell, T. Trials of Anti-Tumour Necrosis Factor Therapy for COVID-19 are Urgently Needed. Lancet 2020, 395, 1407–1409. [Google Scholar] [CrossRef] [PubMed]
- Freeman, T.L.; Swartz, T.H. Targeting the NLRP3 Inflammasome in Severe COVID-19. Front. Immunol. 2020, 11, 1518. [Google Scholar] [CrossRef]
- Chung, I.C.; OuYang, C.N.; Yuan, S.N.; Lin, H.C.; Huang, K.Y.; Wu, P.S.; Liu, C.Y.; Tsai, K.J.; Loi, L.K.; Chen, Y.J.; et al. Pretreatment with a Heat-Killed Probiotic Modulates the NLRP3 Inflammasome and Attenuates Colitis-Associated Colorectal Cancer in Mice. Nutrients 2019, 11, 516. [Google Scholar] [CrossRef] [Green Version]
- Evans, S.S.; Repasky, E.A.; Fisher, D.T. Fever and the Thermal Regulation of Immunity: The Immune System Feels the Heat. Nat. Rev. Immunol. 2015, 15, 335–349. [Google Scholar] [CrossRef]
- Plaza-Díaz, J.; Ruiz-Ojeda, F.J.; Vilchez-Padial, L.M.; Gil, A. Evidence of the Anti-Inflammatory Effects of Probiotics and Synbiotics in Intestinal Chronic Diseases. Nutrients 2017, 9, 555. [Google Scholar] [CrossRef] [Green Version]
- Taddonio, M.A.; Dolgachev, V.; Bosmann, M.; Ward, P.A.; Su, G.; Wang, S.C.; Hemmila, M.R. Influence of Lipopolysaccharide Binding Protein on Pulmonary Inflammation in Gram-negative Pneumonia. Shock 2015, 43, 612–619. [Google Scholar] [CrossRef] [Green Version]
- Onofrio, L.; Caraglia, M.; Facchini, G.; Margherita, V.; Placido, S.; Buonerba, C. Toll-Like Receptors and COVID-19: A Two-Faced Story with an Exciting Ending. Future Sci. OA 2020, 6, FSO605. [Google Scholar] [CrossRef]
- Kitazawa, H.; Villena, J. Modulation of Respiratory TLR3-Anti-Viral Response by Probiotic Microorganisms: Lessons Learned from Lactobacillus rhamnosus CRL1505. Front. Immunol. 2014, 5, 201. [Google Scholar] [CrossRef] [PubMed]
- Claverys, J.-P.; Havarstein, L.S. Extracellular-peptide control of competence for genetic transformation in Streptococcus pneumonia. Front. Biosci. 2002, 7, d1798–d1814. [Google Scholar] [CrossRef] [Green Version]
- Licciardi, P.V.; Toh, Z.Q.; Dunne, E.; Wong, S.S.; Mulholland, E.K.; Tang, M.; Robins-Browne, R.M.; Satzke, C. Protecting Against Pneumococcal Disease: Critical Interactions Between Probiotics and the Airway Microbiome. PLoS Pathog. 2012, 8, e1002652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, S.S.; Quan Toh, Z.; Dunne, E.M.; Mulholland, E.K.; Tang, M.L.; Robins-Browne, R.M.; Licciardi, P.V.; Satzke, C. Inhibition of Streptococcus pneumoniae Adherence to Human Epithelial Cells in Vitro by the Probiotic Lactobacillus rhamnosus GG. BMC Res. Notes 2013, 6, 135. [Google Scholar] [CrossRef] [Green Version]
- Mak, J.W.Y.; Chan, F.K.L.; Ng, S.C. Probiotics and COVID-19: One size does not fit all. Lancet Gastroenterol. Hepatol. 2020, 5, 644–645. [Google Scholar] [CrossRef]
- NCT01782755. Probiotics: Prevention of Severe Pneumonia and Endotracheal Colonisation Trial (PROSPECT): A Feasibility Clinical Trial. 2020. Available online: https://www.clinicaltrials.gov/study/NCT01782755?term=NCT01782755&rank=1 (accessed on 2 July 2023).
- Baud, D.; Dimopoulou Agri, V.; Gibson, G.R.; Reid, G.; Gianoni, E. Using probiotics to flatten the curve of Coronavirus Disease COVID-2019 Pandemic. Front. Public Health 2020, 8, 186. [Google Scholar] [CrossRef]
- Morone, G.; Palomba, A.; Iosa, M.; Caporaso, T.; De Angelis, D.; Venturiero, V.; Savo, A.; Coiro, P.; Carbone, D.; Gimigliano, F.; et al. Incidence and persistence of viral shedding in COVID-19 post-acute patients with negativized pharyngeal swab: A systematic review. Front. Med. 2020, 7, 562. [Google Scholar] [CrossRef]
- Guarino, A.; Guandalini, S.; Lo Vecchio, A. Probiotics for Prevention and Treatment of Diarrhea. J. Clin. Gastroenterol. 2015, 49 (Suppl. S1), S37–S45. [Google Scholar] [CrossRef]
- NCT05080244. WHO COVID-19—Evaluation of the Efficacy of Probiotics to Reduce Occurrence of Long COVID (PROVID-LD). 2021. Available online: https://www.clinicaltrials.gov/study/NCT05080244?term=NCT05080244&rank=1 (accessed on 2 July 2023).
- NCT04621071. Efficacy of Probiotics in Reducing Duration and Symptoms of COVID-19 (PROVID-19). 2021. Available online: https://www.clinicaltrials.gov/study/NCT04621071?term=NCT04621071&rank=1 (accessed on 2 July 2023).
- NCT05474144. Monitoring the Efficacy of Probiotic Dietary Supplement SmartProbio C in Patients with Severe COVID-19 Infection. 2021. Available online: https://www.clinicaltrials.gov/study/NCT05474144?term=NCT05474144&rank=1 (accessed on 2 July 2023).
- NCT04390477. Study to Evaluate the Effect of a Probiotic in COVID-19. 2020. Available online: https://www.clinicaltrials.gov/study/NCT04390477?term=NCT04390477&rank=1 (accessed on 2 July 2023).
- NCT04458519. Efficacy of Intranasal Probiotic Treatment to Reduce Severity of Symptoms in COVID-19 Infection. 2020. Available online: https://www.clinicaltrials.gov/study/NCT04458519?term=NCT04458519&rank=1 (accessed on 2 July 2023).
- NCT04937556. Evaluation of a Probiotic Supplementation in the Immune Response of Participants with COVID-19 (Coronavirus Disease). 2021. Available online: https://www.clinicaltrials.gov/study/NCT04937556?term=NCT04937556&rank=1 (accessed on 2 July 2023).
- NCT04734886. The Effect of Probiotic Supplementation on SARS-CoV-2 Antibody Response after COVID-19. 2020. Available online: https://clinicaltrials.gov/study/NCT04734886?term=NCT04734886&rank=1 (accessed on 2 July 2023).
- NCT05043376. Study to Investigate the Treatment Benefits of Probiotic Streptococcus salivarius K12 for Hospitalised (Non-ICU) Patients with COVID-19. 2021. Available online: https://clinicaltrials.gov/study/NCT05043376?term=NCT05043376&rank=1 (accessed on 2 July 2023).
- NCT05175833. Oral Probiotics and Secondary Bacterial Pneumonia in Severe COVID-19. 2020. Available online: https://clinicaltrials.gov/study/NCT05175833?term=NCT05175833&rank=1 (accessed on 2 July 2023).
- NCT04847349. Live Microbials to Boost Anti-Severe Acute Respiratory Syndrome Coronavirus-2 (SARS-CoV-2) Immunity Clinical Trial. 2021. Available online: https://clinicaltrials.gov/study/NCT04847349?term=NCT04847349&rank=1 (accessed on 2 July 2023).
- NCT04462627. Reduction of COVID-19 Transmission to Healthcare Professionals. 2020. Available online: https://clinicaltrials.gov/study/NCT04462627?term=NCT04462627&rank=1 (accessed on 2 July 2023).
- NCT04798677. Efficacy and Tolerability of ABBC1 in Volunteers Receiving the Influenza or COVID-19 Vaccine. 2020. Available online: https://clinicaltrials.gov/study/NCT04798677?term=NCT04798677&rank=1 (accessed on 2 July 2023).
- NCT04517422. Efficacy of L. plantarum and P. acidilactici in Adults with SARS-CoV-2 and COVID-19. 2020. Available online: https://clinicaltrials.gov/study/NCT04517422?term=NCT04517422&rank=1 (accessed on 2 July 2023).
- NCT04399252. Effect of Lactobacillus on the Microbiome of Household Contacts Exposed to COVID-19. 2020. Available online: https://clinicaltrials.gov/study/NCT04399252?term=NCT04399252&rank=1 (accessed on 2 July 2023).
- NCT04507867. Effect of a NSS to Reduce Complications in Patients with COVID-19 and Comorbidities in Stage III. 2020. Available online: https://clinicaltrials.gov/study/NCT04507867?term=NCT04507867&rank=1 (accessed on 2 July 2023).
- Baindara, P.; Chakraborty, R.; Holliday, Z.M.; Mandal, S.M.; Schrum, A.G. Oral Probiotics in Coronavirus Disease 2019: Connecting the Gut-Lung Axis to Viral Pathogenesis, Inflammation, Secondary Infection and Clinical Trials. New Microbes New Infect. 2021, 40, 100837. [Google Scholar] [CrossRef]
- Mirzaei, R.; Attar, A.; Papizadeh, S.; Jeda, A.S.; Hosseini-Fard, S.R.; Jamasbi, E.; Kazemi, S.; Amerkani, S.; Talei, G.R.; Moradi, P.; et al. The emerging role of probiotics as a mitigation strategy against coronavirus disease 2019 (COVID-19). Arch. Virol. 2021, 8, 05036. [Google Scholar] [CrossRef]

| Study Ref. Clinical Trial Identifier | Study Title Study Focus | Probiotic Intervention | Procedure Synopsis | Country & Start/Completion of Study [Reference] |
|---|---|---|---|---|
| NCT05080244 No results | Probiotics: reduce occurrence of long COVID-19 | ProB—2 strains (ProB strains not disclosed) | 618 patients, RCT. 2 capsules/d for 10d, then 1/d to day 25. | Quebec, Canada Oct 2021/Dec 2022. [193] |
| NCT04621071 No results | Probiotics: reduce duration & symptoms of COVID-19 | ProB—2 strains (ProB strains not disclosed) | 17 patients, RCT. 2 capsules/d for 10d, 1 capsule/d to day 25. | Quebec, Canada Jan 2021/Oct 2021. [194] |
| NCT05474144 No results | Efficacy of ProB in patients with severe COVID-19 infection | SmartProbio C 19 strains + inulin + maltodextrin | 83 patients, RCT. Triple masking. Twice/d, 2 wks. | Brno, Czech Republic. Nov 2021/April 2022. [195] |
| NCT04390477 No results | Evaluate effect of ProB in COVID-19 | ProB strains (not disclosed) & maltodextrin | 41 participants No masking. 1 capsule per day for 30 d. | Alicante, Spain May 2020/March 2021. [196] |
| NCT04458519 No results | i/n ProB reductn of symptom severity in COVID-19 | Probiorinse L.lactis W136 | 23 participants, single blinded, for 14d | Montreal, Canada July 2020/May 2021 [197] |
| NCT04937556 No results | ProB supplementn in IR of COVID-19 participants | L. salivarius, Vit D + Zinc | 41 participants randomised triple masking, 28 d | Madrid, Spain Oct 2021/March 2022. [198] |
| NCT04734886 No results | ProB supplementn on SARS-CoV-2 Ab IR after COVID19 | L.reuteri DSM17938 + Vit D | 161 participants Quadruple masking, daily for 6 months | Orebro, Sweden Nov 2020/Sept 2021. [199] |
| NCT05043376 No results | ProB S. salivarius K12 for hospitalised (non-ICU) patients with COVID-19 | BLIS K12 Streptococcus salivarius K12. | 50 participants randomised, open label. Daily, to day 14. | Lahore, Pakistan Sept 2021/Nov 2021. [200] |
| NCT05175833 No results | Oral ProB and secondary bacterial pneumonia in severe COVID-19 | Oral gel ProB Streptococcus salivarius K12 & L.brevis CD2 | 70 participants Randomised, quadruple masking, 7 d course | Passo Fundo, Brazil Sept 2020/Jan 2021. [201] |
| NCT04847349 No results | Live microbials to boost SARS-CoV-2 immunity | Dietary supplement OL-1 (ProB consortium, strains not disclosed) | 54 participants Randomised Quadruple masking, daily 21 d | New Jersey USA April 2021/Jan 2022. [202] |
| NCT04462627 No results | Reduction of COVID-19 transmission to healthcare professionals | Dietary supplement probiotic (Probactiol Plus—Metagenics) B.lactis Bi-07; L.acidophilus NCFM | 566 participants, open-label, no masking. | Brussels, Belgium April 2020/April 2022. [203] |
| NCT04798677 No results | Efficacy & tolerability of ABBC1 in volunteers receiving influenza or COVID-19 vaccine. | ABBC1: beta-1,3/1,6-glucan + inactivated Saccharomyces cerevisiae (with Se, Zn) | 72 participants RCT, triple masking. 30 d supplementn | Barcelona, Spain Oct 2020/Sept 2021. [204] |
| NCT04517422 No results | Efficacy of L.plantarum & P.acidilactici in adults with SARS-CoV-2 & COVID-19 | L.plantarum CECT30292, CECT7484, CECT7485y & P.acidilactici CECT7483 with maltodextrin. | 300 participants. RCT, quadruple masking. 1 dose per day over 30 d dietary supplementn | Mexico City, Mexico Aug 2020/Feb 2021 [205] |
| NCT04399252 No results | Effect of Lactobacillus on the microbiome of household contacts exposed to COVID-19 | L.rhamnosus GG | 182 participants. RCT, triple masking. 2 capsules per day over 28 d. | North Carolina, USA June 2020/July 2021. [206] |
| NCT04507867 Results available. | Effcect of a NSS to reduce complications in patients with COVID-19 and comorbidities in stage III. | Nutritional Support System: i/m Vit B1,B6,B12. Saccharomyces boulardii CNCM I-745 “Floratil” | 80 participants. RCT, triple masking. 1 capsule, twice a day over 6 d. | Mexico State, Mexico. Sept 2020/April 2021. [207] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tomkinson, S.; Triscott, C.; Schenk, E.; Foey, A. The Potential of Probiotics as Ingestible Adjuvants and Immune Modulators for Antiviral Immunity and Management of SARS-CoV-2 Infection and COVID-19. Pathogens 2023, 12, 928. https://doi.org/10.3390/pathogens12070928
Tomkinson S, Triscott C, Schenk E, Foey A. The Potential of Probiotics as Ingestible Adjuvants and Immune Modulators for Antiviral Immunity and Management of SARS-CoV-2 Infection and COVID-19. Pathogens. 2023; 12(7):928. https://doi.org/10.3390/pathogens12070928
Chicago/Turabian StyleTomkinson, Sophie, Cloe Triscott, Emily Schenk, and Andrew Foey. 2023. "The Potential of Probiotics as Ingestible Adjuvants and Immune Modulators for Antiviral Immunity and Management of SARS-CoV-2 Infection and COVID-19" Pathogens 12, no. 7: 928. https://doi.org/10.3390/pathogens12070928