
Helminth Prevalence in European Deer with a Focus on Abomasal Nematodes and the Influence of Livestock Pasture Contact: A Meta-Analysis


School of Biological Sciences, Queen’s University Belfast, Belfast BT9 5DL, UK
*
Author to whom correspondence should be addressed.
Pathogens 2024, 13(5), 378; https://doi.org/10.3390/pathogens13050378
Submission received: 13 March 2024
/
Revised: 21 April 2024
/
Accepted: 24 April 2024
/
Published: 1 May 2024
(This article belongs to the Special Issue Emerging and Endemic Infections in Wildlife: Epidemiology, Ecology, & Management in a Changing World)
Abstract
:Deer are susceptible to infection with parasitic helminths, including species which are of increasing economic concern to the livestock industry due to anthelmintic drug resistance. This paper systematically collates helminth prevalence data from deer across Europe and explores patterns in relation to host and parasite species, as well as landscape factors. A livestock pasture contact index (LPCI) is developed to predict epidemiological overlap between deer and livestock, and hence to examine deer helminth fauna in the context of their surrounding environment. Fifty-eight studies comprising fallow (Dama dama), red (Cervus elaphus), roe (Capreolus capreolus) and sika (Cervus nippon) deer were identified. Deer populations in “likely” contact with livestock pasture had a higher mean prevalence of the abomasal nematodes Haemonchus contortus, Ostertagia ostertagi, Teladorsagia circumcincta and Trichostrongylus axei (p = 0.01), which are common in livestock and not primarily associated with deer. Roe deer populations had a higher prevalence of T. circumcincta (p = 0.02) and T. axei (p = 0.01) than fallow deer and a higher prevalence of H. contortus than both red (p = 0.01) and fallow deer (p = 0.02). Liver fluke and lungworm species were present sporadically at low prevalence, while the abomasal nematode Ashworthius sidemi occurred locally at high prevalence. Insights from this research suggest that deer helminth fauna is reflective of their surrounding environment, including the livestock species which inhabit areas of shared grazing. This is explored from an epidemiological perspective, and the prospect of helminth transmission between wild and domestic hosts is discussed, including drug-resistant strains, alongside the role of helminths as indicators relevant to the transmission of other pathogens at the wildlife–livestock interface.
1. Introduction
Helminth infection is an increasing economic burden on the livestock industry and is becoming harder to manage due to anthelmintic resistance [1], and infection can also affect host fitness and population viability in wild ruminants [2]. With rising cases of anthelmintic resistance in livestock in Europe [3], it is important to consider avenues in which helminths can spread across landscapes and between livestock farms. The helminth fauna of deer includes species with high host specificity, as well as more generalist species, including those which are typically associated with livestock [4,5]. Transmission is indirect, occurring when hosts ingest helminths during the infective period of their lifecycle, from grass and other forage. The infective period following the deposition of helminth eggs typically has seasonal trends, influenced by interconnected environmental parameters such as rainfall, temperature, and, for trematodes such as the liver fluke Fasciola hepatica and some nematode species, the availability of intermediate hosts.
Roe (Capreolus capreolus Linnaeus, 1758) and red deer (Cervus elaphus Linnaeus, 1758) are the most abundant cervid species in Europe and are often sympatric [6], while fallow (Dama dama Linnaeus, 1758) and sika deer (Cervus nippon Temminck, 1838) are also widely distributed [7,8]. Substantiated helminth studies have been conducted for these deer species [9,10,11], with helminths typically being identified morphologically. Species associated with sheep and goats such as the haematophagous abomasal nematode Haemonchus contortus have been recorded in multiple deer species [10,12,13]. Further, drug-resistant genotypes of H. contortus have been identified in deer in the United Kingdom and Hungary, indicating transmission from livestock pasture [14,15,16]. For trichostrongylid nematodes such as H. contortus, eggs develop into larvae and migrate from faeces to pasture at a rate dependent on local weather conditions [17,18]. Due to the indirect nature of helminth transmission, wild hosts can accumulate infections by grazing areas previously used by livestock, and vice versa, without being present on the pasture at the same time [19]. In addition to evidence of drug-resistant nematodes in wild hosts, there is also confirmation that they can transmit these back to domestic livestock [14]. Further, there is evidence that common nematode species regularly circulate between wild and domestic alpine ruminant hosts [20]. The probability and rate of transmission at the wildlife–livestock interface, however, is generally not well understood and is likely to differ due to various host, climate and landscape factors.
Helminth transmission is a dynamic process, and understanding the conditions under which wild hosts become infected with different species could provide important epidemiological insights about their role in transmitting livestock-related nematodes between farms. Deer helminth studies in Europe have typically been at a regional or local scale [12,21,22] and record the prevalence of helminths within a sampled population or the abundance of helminths within each deer species. Despite numerous studies, few have directly explored the relationship between livestock pasture contact and the prevalence of livestock-associated helminths in deer [11,14,15].
This paper reviews helminth prevalence data from previous cervid studies in Europe, with a focus on abomasal nematodes, and through a meta-analysis examines the susceptibility of red, fallow, roe and sika deer to these nematodes. Further, contextual information from the existing studies is used to create a livestock pasture contact index (LPCI), which, in turn, is utilized to explore how cervid abomasal nematode fauna is influenced by their contact with surrounding livestock pastures. Prevalence data of other helminths along the gastrointestinal tract and from the liver and lungs are also collated and examined. This study aims to provide a platform for understanding deer helminth prevalence in Europe, and for understanding helminth transmission between livestock and cervids.
2. Materials and Methods
2.1. Review Protocol and Data Collection
This research followed guidance from the Preferred Reporting Items for Systematic review and Meta-Analysis (PRISMA) [23]. A combination of organ, nematode, deer species and location-related terms were searched consistently across Google Scholar and Science Direct by a single reviewer. The Global Mammal Parasite Database was also searched (GMPD) [24], filtered by host, parasite type and continent. A search using Web of Science and PubMed using the same search terms yielded no additional papers. All returned papers were assessed for relevance and data availability.
The prevalence of helminths in red, fallow, roe and sika deer was explored (prevalence = number of hosts infected ÷ number of hosts sampled). Firstly, studies with abomasal nematodes were identified, and then additional helminth prevalence data from other organs within these studies was also extracted. A subsequent search adding ‘intestine’, ‘liver’ and ‘lung’ found no additional papers. Only studies from continental Europe were included. Where taxonomic revision has been subsequently published, specifically for minor morphs, obsolete helminth species names were synchronized with their equivalent current taxonomic classification. For instance, the abomasal nematode Teladorsagia circumcincta is considered the same species as Teladorsagia trifurcata and Teladorsagia davtiani [25], and prevalence data from studies with any of these species were grouped together. Similarly, the abomasal nematode Spiculopteragia spiculoptera, also known as Spiculopteragia boehmi, is considered the major morph of Spiculopteragia mathevossian [26], and prevalence data recorded for all of these classifications were also combined.
Studies from wild, enclosed and farmed deer were included. Data were collated onto a spreadsheet and comprised study location, abomasal nematode prevalence, study date, sample number, species and sex ratio of deer sampled. Further, the prevalence of helminth species from the large and small intestine, liver and lungs was included if also present in studies which contained abomasal nematode data. No date restrictions were imposed, and each publication was screened for the availability of suitable data. Publications were divided into multiple studies where appropriate (for example, if multiple deer species were sampled in the same study) and given unique study numbers. A “study” was defined as providing helminth data for one deer species, in one location, over a given time period. Studies which included data on multiple organs were assigned the same study number. Publications included studies which primarily surveyed deer helminths or studies which had helminth prevalence data available for another reason.
2.2. Determining Livestock Contact
An index of livestock pasture contact was added to the dataset as determined by contextual information within each publication. The livestock pasture contact index (LPCI) was arranged into three categories: “likely”, “unlikely” and “unknown”. Livestock pasture contact was determined as “likely” if deer were wild and if the surrounding landscape was stated as being used for livestock farming. Alternatively, if farmed deer utilized land which was stated as previously grazed by sheep or cattle, livestock contact was also placed under the “likely” classification. If studies occurred in an enclosed space with no livestock, contact was classified as “unlikely”, and this was also the case if farmed deer were grazing on pasture not previously used by sheep or cattle. Studies were also classified as having “unlikely” livestock pasture contact if a sampled wild deer population was recorded to occur in an area not used for livestock farming. If studies did not determine livestock presence in the surrounding area, livestock contact was classified as “unknown”. If a study included data compiled from both wild and enclosed deer, with different levels of livestock contact, an “unknown” classification was also provided.
2.3. Data Analysis and Visualization
Data analysis was undertaken in R 4.2.0. Figures were prepared using the R packages pheatmap 1.0.12 [27] and ggplot2 3.3.6 [28]. The distribution of helminth prevalence data was examined using the Shapiro–Wilk test and visually using histograms. As the data required for analysis were nonparametric, pairwise Wilcoxon tests were used to explore differences in the prevalence of common abomasal nematodes between cervid species. Bonferroni correction was applied to subsequent statistical outputs to account for multiple tests. When exploring helminth prevalence between different cervid species, only fallow, red and roe deer were included, as there were too few studies of other species to support meaningful statistical analysis.
3. Results
3.1. Information on Studies
In total, 58 studies which included abomasal nematode prevalence data were identified (Figure 1). These were derived from 34 publications across 16 countries (Figure 2). Red deer had the most prevalence studies with 23, followed by fallow deer with 16, roe deer with 15 and sika deer with only 4. All selected studies provided data on abomasal nematodes, while most also included data on helminths of the large and small intestines (Figure 3). Only 20 lungworm prevalence studies were deemed suitable in total, including 7 from fallow deer, 8 from red deer, 4 from roe deer and 1 from sika deer. Even fewer studies provided liver helminth prevalence data, with only six, seven, three and two studies from fallow, red, roe and sika deer, respectively (Figure 3). As such there was insufficient power for statistical analysis of liver and lung helminths.
3.2. Abomasal Nematodes
Nematode species already known to be associated with cervids were the most commonly found across the studies included. Thus, fallow deer were most commonly infected with Spiculopteragia asymmetrica, while red and roe deer were most commonly infected with Spiculopteragia spiculoptera and Ostertagia leptospicularis. Additionally, nematode species that are common in livestock in Europe also occurred in deer, especially in roe deer. H. contortus and Trichostrongylus axei were found in roe deer in 11 of 15 studies, with a mean overall prevalence of 14.7% and 15.8%, respectively. Trichostrongylus axei is a nematode which regularly infects sheep, cattle and other ruminants [55,56]. Further, Te. circumcincta, a common nematode of sheep, was present in 9 of 15 roe deer studies with a mean prevalence of 8.4%. There was a significantly higher prevalence of Te. circumcincta and Tr. axei in roe deer compared to fallow deer, and a significantly higher prevalence of H. contortus in roe deer compared to red and fallow deer (Table 1). Despite substantial infection of livestock-associated nematodes in roe deer, the prevalence of the cervid-related species O. leptospicularis was higher, having a mean overall prevalence of 60.7% and being present in 11 of 15 studies.
Spiculopteragia asymmetrica was the only deer-specific abomasal nematode to show significant prevalence differences amongst cervid species (Table 1), occurring more in fallow than in roe deer populations. Livestock-associated nematodes in fallow deer were rare with Ostertagia ostertagi, a common cattle nematode which causes ostertagiosis [57], having the highest mean prevalence of only 5.4%, and being present in only 3 of 16 studies. In red deer, Tr. axei was the most common livestock-associated nematode with a mean prevalence of 8.9% (12/23 studies), however, O. leptospicularis, a nematode that regularly infects cervids [58], was present most frequently, occurring in 19 of 23 studies, with a mean prevalence of 41.5%. Few conclusions can be drawn regarding sika deer infection, as only four studies were suitable for analysis (Figure 3), however, all of them included livestock-associated abomasal nematodes.
3.3. Intestinal Nematodes
In the small intestine of fallow and red deer, Capillaria bovis was the most commonly reported species, being present in 9 of 12 fallow deer studies which included intestinal nematode data, and in 4 of 12 red deer studies. The species, which is common in cervids despite first being identified in livestock [59], had a mean overall prevalence of 11.6% and 6.1% in fallow and red deer, respectively. Further, there was a significantly higher prevalence of C. bovis in fallow deer compared to roe deer (p = 0.037) which had a mean prevalence of only 1%. In roe deer, Nematodirus filicollis, a species which regularly infects sheep [60], was the most common nematode in the small intestine, having a mean prevalence of 17.3% and being present in 6 of 12 studies. In the small intestine of sika deer, Nematodirus roscidus, a nematode associated with cervids [4,10], and Cooperia pectinata, a nematode primarily associated with cattle [61], were present in one study at a prevalence of 16% and 42%, respectively.
In the large intestine, Oesophagostomum venulosum was the most common nematode in red and fallow deer. This nematode also commonly infects sheep and goats [62] but is rarely considered pathogenic [63]. Oesophagostomum venulosum was present in 9 of 12 fallow deer studies at a mean prevalence of 32.4%, and in 9 of 11 red deer studies with a mean prevalence of 32.9%. In roe deer, Chabertia ovina, a parasite which can cause anaemia and weight loss in sheep [64,65], was the most commonly identified large intestinal species, being present in 8 of 12 studies, and having the highest mean prevalence of 17.5%. In sika deer, O. venulosum was found in 2 of the 3 studies which examined the large intestines. The nematode occurred with a prevalence of 51% and 9%, but Oesophagostomum sikae, a species associated with cervids [4,10], had a higher prevalence of 88% in sika deer despite only being identified in one study.
3.4. Liver Fluke and Lungworm
Fasciola hepatica, a multi-host species found globally [66], was present in 4 of 18 studies which reported liver helminths, including in 2 fallow deer studies and in single red and sika deer studies (Figure 3). The maximum prevalence of F. hepatica occurred in a fallow deer study at 44%. Dicrocoelium dendriticum, a liver fluke typical of grazing ruminants [67], was also present in 4 of 18 studies, including in 1 fallow, 2 red and 1 roe deer studies, with a maximum prevalence of 25% in red deer (Figure 3). Fascioloides magna, a large liver fluke of ruminants originating from North America [68,69], was present in one red deer study from Poland with a prevalence of around 1%. Further, prevalence data for Dicrocoelium chinensis, a small liver fluke originally isolated from musk deer [70], was available from one sika deer study in Austria with a prevalence of 28%.
Dictyocaulus eckerti, an important lungworm of farmed red deer [71], was present in 8 of 20 studies which provided lungworm prevalence data, including in 2 studies with wild fallow, 3 with wild red deer, 2 with wild roe deer and 1 with wild sika deer (Figure 3). The maximum prevalence of D. eckerti was 81% in red deer which occurred in a wild population in Germany. Dictyocaulus noerni prevalence data were also available for red and fallow deer, with a 100% prevalence in a farmed red deer population and 44% in a farmed fallow deer population (Figure 3), but it remains unknown if this is a separate species to D. eckerti [72,73]. Dictyocaulus capreolus, a nematode with high host specificity to roe deer [74], was recorded in 3 of 4 suitable roe deer studies, ranging from 2% to 36% prevalence. Finally, prevalence data of Varestrongylus sagittatus, a lung nematode which settles in the alveolar septum [75], was present in four red deer studies from northern and southern Poland, and from western Germany, and in two fallow deer studies from Austria and north-western Poland. The maximum prevalence occurred at a maximum of 46% prevalence in fallow deer.
3.5. Impact of Livestock Contact on Abomasal Nematode Prevalence
Livestock contact was determined as “likely” in 28 studies, (6 fallow, 11 red, 9 roe, 2 sika) “unlikely” in 12, (5 fallow, 4 red, 1 roe) and “unknown” in 18 (4 fallow, 7 red, 2 roe, 1 sika). There was a significantly higher frequency of livestock-related nematodes including H. contortus, Tr. axei, O. ostertagi and Te. circumcincta in studies with “likely” contact than in those with either “unlikely” or “unknown” levels of contact with livestock pasture (p = 0.01; Figure 4). For studies with “likely” contact, 25 of 28 had at least one abomasal nematode which is typically associated with domestic ruminants, and on average there were 1.8 of these nematode species present. Further, studies with “likely” livestock contact had a combined mean prevalence of such livestock-related nematodes of 9.3%. Roe deer had “likely” contact with livestock in 60% of the studies, compared to only 48% and 38% for red and fallow deer studies, respectively. Of studies with “unlikely” livestock contact, 6 of 12 had at least one livestock-related abomasal nematode, but the average number of livestock-associated species was 0.9. Further, the mean combined prevalence of livestock-associated nematode species was only 2.6%, much lower than in studies with “likely” contact. Fallow deer were proportionally most unlikely to be in contact with livestock pasture, with this being the case in 31% of studies, compared to 17% and 20% for red and roe deer studies, respectively. The frequency and prevalence of nematode species usually associated with deer were not affected by the assessed likelihood of contact with livestock pasture (Figure 4).
4. Discussion
In this study, the prevalence of helminths in common European deer species is examined in the context of surrounding livestock pasture contact. This heuristic analysis was possible by creating a basic livestock pasture contact index (LPCI), which aimed to assess how livestock proximity, and deer access to livestock pasture, influence their helminth fauna. The approach provided epidemiological insights relevant to the spread of livestock and deer-related helminth species and also indicated how deer helminth fauna could be used to infer their grazing patterns. The LPCI classifications added epidemiological value to the existing studies, providing insights into potential transmission at the wildlife–livestock interface. The associated database summarizes the state of knowledge regarding helminths of deer in Europe, especially in relation to the occurrence of deer specialists and livestock-associated species, and can provide a stimulus for further investigating helminth epidemiology among host species at the wildlife–livestock interface.
4.1. Host Range and Ecology
Teladorsagia circumcincta and Tr. axei were more prevalent in roe deer than in fallow deer, and H. contortus was more prevalent in roe deer than in both red and fallow deer (Table 1). Determining whether roe deer are more susceptible to these nematodes is difficult, however, as they could also be more likely to encounter infective larvae due to their habitat preferences, for example by utilizing edge habitats in fragmented farmed landscapes in order to meet their minimum woodland requirement [76]. This, in turn, could bring them in closer contact with livestock pasture and increase the chance of their ingesting larvae of livestock-related helminths. On the other hand, roe deer are considered to browse more for food than red, fallow or sika deer [77], which implies that they might not encounter as many infective larvae from grass on livestock pasture, compared to deer species which have feeding patterns closer resembling those of grazing livestock. Roe deer show significant digestive plasticity, however, and have been recorded eating higher proportions of grass in fragmented landscapes [78], which could bring them in closer proximity to nematode species such as H. contortus. Indeed, in France, geopositioning system (GPS)-tagged roe deer had higher faecal egg counts in areas with greater livestock density, which could indicate that they acquired more livestock-related gastrointestinal nematode infections in these areas than deer in less livestock-dense areas [79]. To date, roe deer have been the only species identified with drug-resistant nematodes originating from livestock [14,15,16] adding further evidence of their regular presence on livestock pasture. Despite commonly being infected with nematodes such as H. contortus, however, ex situ research has shown that infection intensity in roe deer is unlikely to match that of sheep, with roe deer only producing a maximum of 150 eggs per gram of faeces after being infected with 8000 drug-resistant H. contortus larvae [80]. In the same study, European mouflon, the ancestor of domestic sheep, had a maximum of over 25,000 eggs per gram of faeces after infection with the same number of larvae. Hypothetically, therefore, roe deer might act to reduce livestock-related helminths in farmed landscapes by removing infective larvae from pasture as wild ungulates appear to do in mixed-use grazing systems in Africa [81]. Regardless of their capacity to diminish or amplify pasture contamination, however, roe deer might also be capable of transmitting drug-resistant nematodes to livestock [14]. The extent to which this occurs in the wild is unknown and could not be inferred from papers in the present analysis as the anthelmintic-resistance status of the nematodes recovered was rarely determined.
In the present research, roe deer studies had “likely” contact with livestock in 60% of the studies, compared to only 48% and 38% for red and fallow deer studies, respectively. This is perhaps reflective of roe deer ecology, and their capacity to utilize farmed and fragmented landscapes. In such landscapes, roe deer have been recorded with higher levels of faecal nitrogen content [82], and juvenile roe deer have been over 3 kg heavier than their forest-dwelling equivalents [83]. This again highlights that roe deer change their diet in open landscapes due to the availability of higher-quality food. Although this could increase their chance of encountering livestock-related helminth fauna, it is also possible that a higher quality diet could improve their immunocompetence [84], and thus reduce their ability to maintain infections and propagate helminths between farms. It is also likely that the home range of a deer population will influence how they spread helminths across a landscape. Roe deer typically have a smaller home range size than fallow, red or sika deer [85,86,87], with a male roe deer range being recorded as less than 20 ha in a study in Italy [88]. It might be unlikely, therefore, that individual roe deer contribute heavily to the movement of helminths, including drug-resistant genotypes, long distances across a landscape or between multiple farms. Further, due to the territorial nature of roe deer [89,90], they likely concentrate their feeding in more recurrent areas compared to other deer. If this territory is within a farmed landscape, this could explain why they harbour so many livestock-related nematodes compared to other deer which have fewer local scale territorial restrictions and larger ranges.
4.2. Seasonal Patterns
Studies used in this analysis include roe deer hunted in Ukraine in winter [29], Croatia in spring/summer [13] and from Czech Republic [30] in autumn. There were also seasonal differences for the other species surveyed, with fallow deer studies, for instance, occurring in Poland during spring/summer [31], and during autumn/winter in Romania [12]. In Europe, large discrepancies exist between countries regarding the open hunting seasons of wild ruminants [91], and this inevitably increases bias in surveys of helminths in deer. Different species of helminths, including abomasal nematodes, develop to their infectious stage under different environmental conditions [18], and, therefore, sampling hosts during different seasons of the year will influence the prevalence and abundance of helminths discovered. Indeed, seasonal bias has been suggested as an explanation for discovering drug-resistant H. contortus in roe deer in Hungary, which were hunted in spring and summer, compared to red deer with no drug-resistant nematodes, which were hunted in autumn and winter [16] when H. contortus is typically not as prominent [92]. Further, the role of hypobioisis in wild hosts is poorly understood; arrested development of nematodes during unfavourable conditions, such as over winter, could further decrease apparent prevalence at these times of the year [93,94,95]. Increasingly, research using non-invasive techniques such as metabarcoding is allowing the collection of longitudinal species-specific helminth data in wild ruminants [32] from faecal samples and hence not limited by hunting seasons, although the limitation of hypobiosis remains. Further longitudinal research using these techniques could help reduce seasonal bias in wildlife helminth research.
4.3. Abomasal Nematodes as Epidemiological Indicators
The present study indicates that the deer grazing environment influences the prevalence of livestock-related, but not deer-specific nematodes (Figure 4). As such, the helminth fauna in a deer population might provide an indication of where they have grazed, and what types of domestic livestock pasture they grazed on. For instance, a fallow deer population in Romania had a 54% prevalence of H. contortus, a nematode associated with sheep and goats, when in contact with pasture containing small domestic ruminants [12]. Further, a roe deer population in England, sampled from an area of intensive cattle pasture, showed a 70% prevalence of O. ostertagi [14], which is a common parasite in cattle. This, in turn, might provide insights into the proximity of wild hosts to other multi-host pathogens which can persist in the environment, such as Mycobacterium bovis [96], the pathogen which causes bovine tuberculosis in cattle, but which also infects deer [97,98,99]. Mycobacterium bovis has been found to survive for up to 6 months on pasture during winter periods [99], 16 weeks on animal foodstuffs and 58 days in water [100]. Indirect transmission of M. bovis has increasingly been explored with respect to badger–cattle and direct “nose-to-nose” interactions are considered rare [101]. It is also considered a rarity for wild deer and livestock to have direct contact [102] and consequently, access to an indicator of indirect livestock pasture contact could be a useful epidemiological tool. In Wicklow, Ireland, sika deer are considered a maintenance host of M. bovis [97,99] and understanding the helminth fauna of deer in such areas, might provide insights into the possible transmission routes of the bacteria via indirect sources such as pasture.
Other livestock-related pathogens such as the bovine viral diarrhoea (BVD) virus have also been recorded to persist in the environment, including in slurry for 3 weeks [103], and the virus can infect fallow, red, roe and sika deer [104,105,106,107]. Perhaps then, in landscapes with high densities of livestock and wild ruminants, regular helminth surveys using increasingly convenient molecular tools [32] could act as an epidemiological indicator and provide a guide for subsequent and more targeted viral or bacterial research. Deer are also one of the main drivers of tick expansion in Europe [108], and tick-borne diseases such as anaplasmosis are of increasing veterinary and public health importance [109,110]. For instance, the bacterium Anaplasma phagocytophilum, which can infect deer [111] and is often spread by the tick Ixodes ricinus in Europe, can cause human granulocytic anaplasmosis [110], and also pasture fever in domestic ruminants [112]. Indeed, one strain of the bacterium is potentially capable of infecting red deer, roe deer, cattle and humans [113]. Again, understanding the prevalence of different abomasal nematodes in deer might indicate their proximity to livestock pasture and, therefore, their risk of becoming infested with questing ticks, or alternatively the likelihood that they could transmit ticks and/or tick-borne diseases to livestock pasture. Following attempts for helminths [114] might involve mechanistic models of pathogen transmission that account for spatial contact through shared pasture use [19].
5. Conclusions
Deer in Europe can be infected with a wide range of helminths which can often be described as deer-specific, livestock-associated or generalist species. This is particularly noticeable in the abomasum, with deer-specific species being present in almost every study reviewed, whilst livestock or generalist species were more likely to be present if deer had probable access to the pasture of domestic ruminants. Using a basic livestock pasture contact index, the epidemiological value was added to existing studies and highlighted that the abomasal nematode fauna in deer has the potential to act as a wider epidemiological indicator, particularly for multi-host and environmentally persistent pathogens. Roe deer had a higher prevalence of livestock-related nematodes compared to the other deer species examined. It is unknown, however, if this is because they are more susceptible, or if they simply spend more time on livestock pasture, and thus ingest larvae of livestock-related nematodes in greater numbers. Advances in molecular techniques such as metabarcoding are allowing for more fine-scale and longitudinal data collection on deer helminths, including using ante mortem faecal samples, and with more refined epidemiological tools such as mechanistic models of parasite transmission can sustain advances in understanding the role of contact patterns in driving helminth transmission at the deer–livestock interface.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/pathogens13050378/s1, Table S1: Raw helminth prevalence data. References [4,5,9,10,11,12,13,14,21,29,30,31,32,33,34,35,36,37,38,39,40,41,42,43,44,45,46,47,48,49,50,51,52,53,54] are cited in the Supplementary Materials.
Author Contributions
Conceptualization, T.L.B. and E.R.M.; methodology, T.L.B.; formal analysis, T.L.B.; resources, E.R.M.; data curation, T.L.B.; writing—original draft preparation, T.L.B.; writing—review and editing, T.L.B. and E.R.M.; visualization, T.L.B.; supervision, E.R.M.; project administration, E.R.M.; funding acquisition, T.L.B. and E.R.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Department of Agriculture, Environment and Rural Affairs of Northern Ireland through a PhD scholarship to T.L.B.
Institutional Review Board Statement
This research used secondary data analysis only and did not require ethical approval.
Data Availability Statement
Data used for the analyses presented are provided as Supplementary Materials.
Acknowledgments
We thank Andrew Byrne and Siobhán Porter for initiating the collaboration that led to this work.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Charlier, J.; Rinaldi, L.; Musella, V.; Ploeger, H.W.; Chartier, C.; Vineer, H.R.; Chartier, C.; Hinney, B.; von Samson-Himmelstjerna, G.; Bacescu, B. Initial assessment of the economic burden of major parasitic helminth infections to the ruminant livestock industry in Europe. Prev. Vet. Med. 2020, 182, 105103. [Google Scholar] [CrossRef] [PubMed]
- Kołodziej-Sobocińska, M.; Demiaszkiewicz, A.W.; Pyziel, A.M.; Marczuk, B.; Kowalczyk, R. Does the blood-sucking nematode Ashworthius sidemi (Trichostrongylidae) cause deterioration of blood parameters in European bison (Bison bonasus)? Eur. J. Wildl. Res. 2016, 62, 781–785. [Google Scholar] [CrossRef]
- Vineer, H.R.; Morgan, E.R.; Hertzberg, H.; Bartley, D.J.; Bosco, A.; Charlier, J.; Chartier, C.; Claerebout, E.; de Waal, T.; Hendrickx, G. Increasing importance of anthelmintic resistance in European livestock : Creation and meta-analysis of an open database. Parasite 2020, 27, 2020062. [Google Scholar] [CrossRef] [PubMed]
- Rehbein, S.; Visser, M.; Jekel, I.; Silaghi, C. Endoparasites of the fallow deer (Dama dama) of the Antheringer Au in Salzburg, Austria. Wien. Klin. Wochenschr. 2014, 126, 37–41. [Google Scholar] [CrossRef] [PubMed]
- Santín-Durán, M.; Alunda, J.M.; Hoberg, E.P.; de la Fuente, C. Age Distribution and Seasonal Dynamics of Abomasal Helminths in Wild Red Deer from Central Spain. J. Parasitol. 2008, 94, 1031–1037. [Google Scholar] [CrossRef] [PubMed]
- Borkowski, J.; Banul, R.; Jurkiewicz-Azab, J.; Hołdyński, C.; Święczkowska, J.; Nasiadko, M.; Załuski, D. There is only one winner: The negative impact of red deer density on roe deer numbers and distribution in the Słowiński National Park and its vicinity. Ecol. Evol. 2021, 11, 6889–6899. [Google Scholar] [CrossRef] [PubMed]
- Harris, R.B. Cervus nippon, Sika Deer. The IUCN Red List of Threatened Species. Mammal Rev. 2015, 41, 313–325. [Google Scholar]
- Masseti, M.; Mertzanidou, D. Dama dama, Fallow Deer. The IUCN Red List of Threatened Species. Mamm. Species 2008, 317, 1–8. [Google Scholar] [CrossRef]
- Pato, F.J.; Vázquez, L.; Díez-Baños, N.; López, C.; Sánchez-Andrade, R.; Fernández, G.; Díez-Baños, P.; Panadero, R.; Díaz, P.; Morrondo, P. Gastrointestinal nematode infections in roe deer (Capreolus capreolus) from the NW of the Iberian Peninsula: Assessment of some risk factors. Vet. Parasitol. 2013, 196, 136–142. [Google Scholar] [CrossRef]
- Rehbein, S.; Visser, M. The endoparasites of Sika deer (Cervus nippon) in Austria. Wien. Klin. Wochenschr. 2007, 119, 96–101. [Google Scholar] [CrossRef]
- Santín-Durán, M.; Alunda, J.M.; Hoberg, E.P.; de la Fuente, C. Abomasal parasites in wild sympatric cervids, Red Deer, Cervus elaphus and Fallow Deer, Dama dama, from three localities across Central and Western Spain: Relationship to host density and park management. J. Parasitol. 2004, 90, 1378–1386. [Google Scholar] [CrossRef] [PubMed]
- Hora, F.S.; Genchi, C.; Ferrari, N.; Morariu, S.; Mederle, N.; Dărăbuș, G. Frequency of gastrointestinal and pulmonary helminth infections in wild deer from western Romania. Vet. Parasitol. Region. Stud. Rep. 2017, 8, 75–77. [Google Scholar] [CrossRef] [PubMed]
- Kusak, J.; Špičić, S.; Slijepčević, V.; Bosnić, S.; Rajković Janje, R.; Duvnjak, S.; Sindičić, M.; Majnarić, D.; Cvetnić, Ž.; Huber, Đ. Health status of red deer and roe deer in Gorski Kotar, Croatia. Vet. Arh. 2012, 82, 59–73. [Google Scholar]
- Chintoan-Uta, C.; Morgan, E.R.; Skuce, P.J.; Coles, G.C. Wild deer as potential vectors of anthelmintic-resistant abomasal nematodes between cattle and sheep farms. Proc. R. Soc. B Biol. Sci. 2014, 281, 20132985. [Google Scholar] [CrossRef] [PubMed]
- Csivincsik, Á.; Nagy, G.; Halász, T.; Zsolnai, A. Shared pastures and anthelmintic resistance in wildlife and livestock. Agric. Conspec. Sci. 2017, 82, 189–191. [Google Scholar]
- Nagy, G.; Csivincsik, Á.; Sugár, L.; Zsolnai, A. Benzimidazole resistance within red deer, roe deer and sheep populations within a joint habitat in Hungary. Small Rumin. Res. 2017, 149, 172–175. [Google Scholar] [CrossRef]
- Grenfell, B.T. Maximum-likelihood estimates of the mortality and migration rates of the infective larvae of Ostertagia ostertagi and Cooperia oncophora. Parasitology 1986, 92, 643–652. [Google Scholar] [CrossRef]
- O’Connor, L.J.; Walkden-Brown, S.W.; Kahn, L.P. Ecology of the free-living stages of major trichostrongylid parasites of sheep. Vet. Parasitol. 2006, 142, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Morgan, E.R.; Milner-Gulland, E.J.; Torgerson, P.R.; Medley, G.F. Ruminating on complexity: Macroparasites of wildlife and livestock. Trends Ecol. Evol. 2004, 19, 181–188. [Google Scholar] [CrossRef]
- Cerutti, M.C.; Citterio, C.V.; Bazzocchi, C.; Epis, S.; D'Amelio, S.; Ferrari, N.; Lanfranchi, P. Genetic variability of Haemonchus contortus (Nematoda: Trichostrongyloidea) in alpine ruminant host species. J. Helminthol. 2010, 84, 276–283. [Google Scholar] [CrossRef]
- Davidson, R.K.; Kutz, S.J.; Madslien, K.; Hoberg, E.; Handeland, K. Gastrointestinal parasites in an isolated Norwegian population of wild red deer (Cervus elaphus). Acta Vet. Scand. 2014, 56, 59. [Google Scholar] [CrossRef] [PubMed]
- Demiaszkiewicz, A.W.; Merta, D.; Kobielski, J.; Filip, K.J.; Pyziel, A.M. Expansion of Ashworthius sidemi in red deer and roe deer from the Lower Silesian Wilderness and its impact on infection with other gastrointestinal nematodes. Acta Parasitol. 2017, 62, 853–857. [Google Scholar] [CrossRef] [PubMed]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E. The PRISMA 2020 statement: An updated guideline for reporting systematic reviews. Br. Med. J. 2021, 372, n71. [Google Scholar] [CrossRef] [PubMed]
- Stephens, P.R.; Pappalardo, P.; Huang, S.; Byers, J.E.; Farrell, M.J.; Gehman, A.; Ghai, R.R.; Haas, S.E.; Han, B.; Park, A.W.; et al. Global mammal parasite database version 2.0. Ecology 2017, 98, 1476. [Google Scholar] [CrossRef] [PubMed]
- Stevenson, L.A.; Gasser, R.B.; Chilton, N.B. The ITS-2 rDNA of Teladorsagia circumcincta, T. trifurcata and T. davtiani (Nematoda: Trichostrongylidae) indicates that these taxa are one species. Int. J. Parasitol. 1996, 26, 1123–1126. [Google Scholar] [CrossRef]
- Lienard, E.; Depaquit, J.; Ferté, H. Spiculopteragia mathevossiani Ruchliadev, 1948 is the minor morph of Spiculopteragia spiculoptera (Gushanskaya, 1931): Molecular evidence. Vet. Res. 2006, 37, 683–694. [Google Scholar] [CrossRef]
- Kolde, R. Package “pheatmap”: Pretty heatmaps. Version 1012 1–8. 2019. Available online: https://cran.ms.unimelb.edu.au/web/packages/pheatmap/pheatmap.pdf (accessed on 20 June 2023).
- Wickham, H. Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar] [CrossRef]
- Kuzmina, T.; Kharchenko, V.; Malega, A. Helminth fauna of roe deer (Capreolus capreolus) in Ukraine: Biodiversity and parasite community. Vestn. Zool. 2010, 44, 12–19. [Google Scholar] [CrossRef]
- Vetýška, V. Endoparasites of Roe Deer in the Strakonice region. Acta Vet. Brno 1980, 49, 91–103. [Google Scholar] [CrossRef]
- Cisek, A.; Balicka-Ramisz, A.; Ramisz, A.; Pilarczyk, B. Occurrence of gastro-intestinal nematodes in cervids (Cervidae) of north-western Poland. Electr. J. Pol. Agric. Univ. 2003, 6, 1–7. [Google Scholar]
- Beaumelle, C.; Redman, E.M.; de Rijke, J.; Wit, J.; Benabed, S.; Debias, F.; Duhayer, J.; Pardonnet, S.; Poirel, M.T.; Capron, G.; et al. Metabarcoding in two isolated populations of wild roe deer (Capreolus capreolus) reveals variation in gastrointestinal nematode community composition between regions and among age classes. Parasites Vectors 2021, 14, 594. [Google Scholar] [CrossRef]
- Kowal, J.; Nosal, P.; Bonczar, Z.; Wajdzik, M. Parasites of captive fallow deer (Dama dama L.) from southern Poland with special emphasis on Ashworthius sidemi. Ann. Parasitol. 2012, 58, 23–26. [Google Scholar] [PubMed]
- Ramajo Martín, V.; Pérez Sánchez, R.; Ramajo Hernández, A.; Oleaga, A. Preliminary data about the parasitism caused by protozoa, helminths and ticks in cervids and wild bovids from Salamanca (western Spain). Res. Rev. Parasitol. 2007, 67, 69–77. [Google Scholar]
- Sleeman, D.P. Parasites of Deer in Ireland. J. Life Sci. R. Dublin Soc. 1983, 4, 203–210. [Google Scholar]
- Drózdz, J.; Malczewski, A.; Demiaszkiewicz, A.W.; Lachowicz, J. The helminthofauna of farmed deer (Cervidae) in Poland. Acta Parasitol. 1997, 42, 225–229. [Google Scholar]
- Batty, A.F.; Chapman, D.I. Gastro-intestinal parasites of wild Fallow Deer (Dama dama L.). J. Helminthol. 1970, 44, 57–61. [Google Scholar] [CrossRef]
- Ambrosi, M.; Manfredi, M.T.; Lanfranchi, P. Pattern of abomasal helminths in fallow deer farming in Umbria (central Italy). Vet. Parasitol. 1993, 47, 81–86. [Google Scholar] [CrossRef] [PubMed]
- Vengušt, G.; Bidovec, A. Parasites of fallow deer (Dama dama) in Slovenia. Helminthologia 2003, 40, 161–164. [Google Scholar]
- Barth, D.; Matzke, P. Gastro-intestinal nematodes of fallow deer (Dama dama L.) in Germany. Vet. Parasitol. 1984, 16, 173–176. [Google Scholar] [CrossRef]
- Balicka-Ramisz, A.; Pilarczyk, B.; Ramisz, A.; Cisek, A. Occurrence of gastrointestinal and pulmonary nematodes of fallow deer (Dama dama L.) in North-West Poland. Acta Parasitol. 2005, 50, 94–96. [Google Scholar]
- Rehbein, S.; Lutz, W.; Visser, M.; Winter, R. Beiträge zur Kenntnis der Parasitenfauna des Wildes in Nordrhein-Westfalen. 2. Der Endoparasitenbefall des Damwildes. Eur. J. Wildl. Res. 2001, 47, 1. [Google Scholar] [CrossRef]
- Magdálek, J.; Bourgoin, G.; Vadlejch, J. Non-native nematode Ashworthius sidemi currently dominates the abomasal parasite community of cervid hosts in the Czech Republic. Front. Vet. Sci. 2022, 9, 862092. [Google Scholar] [CrossRef]
- Demiaszkiewicz, A.W.; Pyziel, A.; Lachowicz, J. Abomasum and lung nematodes of red deer in Strzałowo Forest District (Piska Primeval Forest). Sylwan 2009, 153, 57–61. [Google Scholar]
- Demiaszkiewicz, A.W.; Merta, D.; Kobielski, J. Infection of red deer by parasites in South-Western Poland (Lower Silesian Wilderness). Med. Weter. 2016, 72, 317–320. [Google Scholar]
- Drozdz, J.; Demiaszkiewicz, A.W.; Lachowicz, J. Exchange of gastrointestinal nematodes between roe and red deer [Cervidae] and European bison [Bovidae] in the Bieszczady Mountains [Carpathians, Poland]. Acta Parasitol. 2002, 47, 314–317. [Google Scholar]
- Manfredi, M.T.; Di Cerbo, A.R.; Tranquillo, V.; Nassuato, C.; Pedrotti, L.; Piccolo, G. Abomasal nematodes of the red deer Cervus elaphus in north-eastern Italy. J. Helminthol. 2007, 81, 247–253. [Google Scholar] [CrossRef]
- Zaffaroni, E.; Manfredi, M.T.; Citterio, C.; Sala, M.; Piccolo, G.; Lanfranchi, P. Host specificity of abomasal nematodes in free ranging alpine ruminants. Vet. Parasitol. 2000, 90, 221–230. [Google Scholar] [CrossRef] [PubMed]
- Rehbein, S.; Lutz, W.; Visser, M.; Winter, R. Contributions à la connaissance de la faune parasitaire des espèces-gibier en Rhénanie-Westphalie 3. Endoparasitoses chez le Cerf. Eur. J. Wildl. Res. 2002, 48, 69. [Google Scholar] [CrossRef]
- García-Romero, C.; Valcárcel, F.; Corchero, J.M.; Olmeda, A.S.; Pérez-Jiménez, J.M. A contribution to the study of parasites of red deer (Cervus elaphus) in the provinces of Toledo and Ciudad Real (Castille-La Mancha, Spain). Ecología 2000, 14, 235–249. [Google Scholar]
- Valcárcel, F.; Corchero, J.; Olmeda, A.S.; Rojo Vázquez, F.A.; García Romero, C. Gastrointestinal nematode infections of Cervus elaphus in Castilla-La Mancha (Central Spain). Rev. Ibérica De Parasitol. 2002, 62, 108–113. [Google Scholar]
- Kuznetsov, D.N.; Romashova, N.B.; Romashov, B.V. Gastrointestinal nematodes of European roe deer (Capreolus capreolus) in Russia. Russ. J. Theriol. 2020, 19, 85–93. [Google Scholar] [CrossRef]
- Rehbein, S.; Lutz, W.; Visser, M.; Winter, R. Investigation of the parasite fauna of wildlife in North Rhine-Westphalia. 1. Endoparasites of roe deer. Zeitschrift für Jagdwissenschaft 2000, 46, 248–269. [Google Scholar] [CrossRef]
- Šarkūnas, M.; Veličkaitė, S.; Bružinskaitė, R.; Malakauskas, A.; Petkevičius, S. Faecal egg output and herbage contamination with infective larvae of species of Ostertagia and Oesophagostomum from naturally infected farmed sika deer Cervus nippon in Lithuania. J. Helminthol. 2007, 81, 79–84. [Google Scholar] [CrossRef] [PubMed]
- Barone, C.D.; Wit, J.; Hoberg, E.P.; Gilleard, J.S.; Zarlenga, D.S. Wild ruminants as reservoirs of domestic livestock gastrointestinal nematodes. Vet. Parasitol. 2020, 279, 109041. [Google Scholar] [CrossRef]
- Borgsteede, F.H.M.; Tibben, J.; Cornelissen, J.B.W.; Agneessens, J.; Gaasenbeek, C.P. Nematode parasites of adult dairy cattle in the Netherlands. Vet. Parasitol. 2000, 89, 287–296. [Google Scholar] [CrossRef]
- Charlier, J.; Höglund, J.; Morgan, E.R.; Geldhof, P.; Vercruysse, J.; Claerebout, E. Biology and epidemiology of gastrointestinal nematodes in cattle. Vet. Clin. N. Am. Food Anim. Pract. 2020, 36, 1–15. [Google Scholar] [CrossRef]
- Wyrobisz-Papiewska, A.; Kowalm, J.; Nosal, P.; Chovancová, G.; Rehbein, S. Host specificity and species diversity of the Ostertagiinae Lopez-Neyra, 1947 in ruminants: A European perspective. Parasites Vectors 2018, 11, 369. [Google Scholar] [CrossRef]
- Justine, J.-L.; Ferté, H. Redescription de Capillaria bovis (Schnyder, 1906) (Nematoda, Capillariinae), parasite du mouflon, du daim et du chevreuil en France. Bull. Mus. Natn. Hist. Nat. Paris 1989, 4, 79–96. [Google Scholar]
- Van Dijk, J.; Morgan, E.R. Hatching behaviour of Nematodirus filicollis in a flock co-infected with Nematodirus battus. Parasitology 2009, 136, 805–811. [Google Scholar] [CrossRef] [PubMed]
- Herlich, H. The development of Cooperia pectinata, a nematode parasite of cattle. Am. J. Vet. Res. 1965, 26, 1026–1031. [Google Scholar]
- Sissay, M.M.; Uggla, A.; Waller, P.J. Prevalence and seasonal incidence of nematode parasites and fluke infections of sheep and goats in eastern Ethiopia. Trop. Anim. Health Prod. 2007, 39, 521–531. [Google Scholar] [CrossRef]
- Goldberg, A. Effects of the nematode Oesophagostomum venulosum on sheep and goats. J. Parasitol. 1952, 38, 35–47. [Google Scholar] [CrossRef] [PubMed]
- Herd, R.P. The Pathogenic Importance of Chaberia Ovina (Fabricius, 1788) in Experimentally Infected Sheep; Pergamon Press: Oxford, UK, 1971. [Google Scholar]
- Ross, J.G.; Dow, C.; Purcell, D.A. A study of Chabertia ovina infections in lambs. Br. Vet. J. 1969, 125, 136–140. [Google Scholar] [CrossRef] [PubMed]
- Vázquez, A.A.; Sabourin, E.; Alda, P.; Leroy, C.; Leray, C.; Carron, E.; Mulero, S.; Caty, C.; Hasfia, S.; Boisseau, M.; et al. Genetic diversity and relationships of the liver fluke Fasciola hepatica (Trematoda) with native and introduced definitive and intermediate hosts. Transbound. Emerg. Dis. 2021, 68, 2274–2286. [Google Scholar] [CrossRef] [PubMed]
- Majidi-Rad, M.; Meshgi, B.; Bokaie, S. The prevalence and intensity rate of Dicrocoelium dendriticum infection in ruminants of three provinces in coastal regions of the Caspian Sea. Iran. J. Vet. Med. 2018, 12, 27–33. [Google Scholar] [CrossRef]
- Leontovyč, R.; Košťáková, M.; Siegelová, V.; Melounová, K.; Pankrác, J.; Vrbová, K.; Horák, P.; Kašný, M. Highland cattle and Radix labiata, the hosts of Fascioloides magna. BMC Vet. Res. 2014, 10, 41. [Google Scholar] [CrossRef] [PubMed]
- Rehbein, S.; Visser, M.; Hamel, D.; Reindl, H. Occurrence of the giant liver fluke, Fascioloides magna, in sympatric wild ungulates in one area in the Upper Palatinate Forest (northeastern Bavaria, Germany). Parasitol. Res. 2021, 120, 553–561. [Google Scholar] [CrossRef] [PubMed]
- Otranto, D.; Rehbein, S.; Weigl, S.; Cantacessi, C.; Parisi, A.; Lia, R.P.; Olson, P.D. Morphological and molecular differentiation between Dicrocoelium dendriticum (Rudolphi, 1819) and Dicrocoelium chinensis (Sudarikov and Ryjikov, 1951) Tang and Tang, 1978 (Platyhelminthes: Digenea). Acta Trop. 2007, 104, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Johnson, M.; Mackintosh, C.G.; Labes, R.E.; Taylor, M.J. Dictyocaulus eckerti, lungworm infecting farmed red deer in new zealand. N. Z. Vet. J. 2001, 49, 34–35. [Google Scholar] [CrossRef]
- Bangoura, B.; Brinegar, B.; Creekmore, T.E. Dictyocaulus cervi-like lungworm infection in a rocky mountain elk (Cervus canadensis nelsoni) from Wyoming, USA. J. Wildl. Dis. 2021, 57, 71–81. [Google Scholar] [CrossRef]
- Divina, B.P.; Wilhelmsson, E.; Mattsson, J.G.; Waller, P.; Höglund, J. Identification of Dictyocaulus spp. in ruminants by morphological and molecular analyses. Parasitology 2000, 121, 193–201. [Google Scholar] [CrossRef]
- Bolukbas, C.S.; Gurler, A.T.; Beyhan, Y.E.; Acici, M.; Umur, S. Helminths of roe deer (Capreolus capreolus) in the Middle Black Sea Region of Turkey. Parasitol. Int. 2012, 61, 729–730. [Google Scholar] [CrossRef] [PubMed]
- Panayotova-Pencheva, M.S.; Alexandrov, M.T. Etiopathological aspects of Elaphostrongylus cervi and Varestrongylus sagittatus infections in red deer in Bulgaria. Acta Vet. Brno 2011, 80, 349–352. [Google Scholar] [CrossRef]
- Morellet, N.; van Moorter, B.; Cargnelutti, B.; Angibault, J.M.; Lourtet, B.; Merlet, J.; Ladet, S.; Hewison, A.M. Landscape composition influences roe deer habitat selection at both home range and landscape scales. Landsc. Ecol. 2011, 26, 999–1010. [Google Scholar] [CrossRef]
- Hofmann, R.R. Evolutionary steps of ecophysiological adaptation and diversification of ruminants: A comparative view of their digestive system. Oecologia 1989, 78, 443–457. [Google Scholar] [CrossRef]
- Ferron, E.S.; Verheyden, H.; Hummel, J.; Cargnelutti, B.; Lourtet, B.; Merlet, J.; Gonzalez-Candela, M.; Angibault, J.M.; Hewison, A.J.; Clauss, M. Digestive plasticity as a response to woodland fragmentation in roe deer. Ecol. Res. 2012, 27, 77–82. [Google Scholar] [CrossRef]
- Verheyden, H.; Richomme, C.; Sevila, J.; Merlet, J.; Lourtet, B.; Chaval, Y.; Hoste, H. Relationship between the excretion of eggs of parasitic helminths in roe deer and local livestock density. J. Helminthol. 2020, 94, 1–6. [Google Scholar] [CrossRef]
- Laca Megyesi, Š.; Königová, A.; Babják, M.; Molnár, L.; Rajský, M.; Szestáková, E.; Major, P.; Soroka, J.; Urda Dolinská, M.; Komáromyová, M.; et al. Wild ruminants as a potential risk factor for transmission of drug resistance in the abomasal nematode Haemonchus contortus. Eur. J. Wildl. Res. 2020, 66, 9. [Google Scholar] [CrossRef]
- Walker, J.G.; Evans, K.E.; Rose Vineer, H.; van Wyk, J.A.; Morgan, E.R. Prediction and attenuation of seasonal spillover of parasites between wild and domestic ungulates in an arid mixed-use system. J. Appl. Ecol. 2018, 55, 1976–1986. [Google Scholar] [CrossRef] [PubMed]
- Verheyden, H.; Aubry, L.; Merlet, J.; Petibon, P.; Chauveau-Duriot, B.; Guillon, N.; Duncan, P. Faecal nitrogen, an index of diet quality in roe deer Capreolus capreolus? Wildl. Biol. 2011, 17, 166–175. [Google Scholar] [CrossRef]
- Hewison, A.J.M.; Morellet, N.; Verheyden, H.; Daufresne, T.; Angibault, J.M.; Cargnelutti, B.; Merlet, J.; Picot, D.; Rames, J.L.; Joachim, J.; et al. Landscape fragmentation influences winter body mass of roe deer. Ecography 2009, 32, 1062–1070. [Google Scholar] [CrossRef]
- Navarro-Gonzalez, N.; Verheyden, H.; Hoste, H.; Cargnelutti, B.; Lourtet, B.; Merlet, J.; Daufresne, T.; Lavín, S.; Hewison, A.M.; Morand, S.; et al. Diet quality and immunocompetence influence parasite load of roe deer in a fragmented landscape. Eur. J. Wildl. Res. 2011, 57, 639–645. [Google Scholar] [CrossRef]
- Jerina, K. Roads and supplemental feeding affect home-range size of Slovenian red deer more than natural factors. J. Mammal. 2012, 93, 1139–1148. [Google Scholar] [CrossRef]
- Nugent, G. Home range size and its development for fallow deer in the Blue Mountains, New Zealand. Acta Theriol. 1994, 39, 159–175. [Google Scholar] [CrossRef]
- Stephens, P.A.; Zaumyslova, O.Y.; Miquelle, D.G.; Myslenkov, A.I.; Hayward, G.D. Estimating population density from indirect sign: Track counts and the Formozov-Malyshev-Pereleshin formula. Anim. Conserv. 2006, 9, 339–348. [Google Scholar] [CrossRef]
- Lovari, S.; Serrao, G.; Mori, E. Woodland features determining home range size of roe deer. Behav. Process. 2017, 140, 115–120. [Google Scholar] [CrossRef] [PubMed]
- Hoem, S.A.; Melis, C.; Linnell, J.D.C.; Andersen, R. Fighting behaviour in territorial male roe deer Capreolus capreolus: The effects of antler size and residence. Eur. J. Wildl. Res. 2007, 53, 1–8. [Google Scholar] [CrossRef]
- Maublanc, M.L.; Bideau, E.; Willemet, R.; Bardonnet, C.; Gonzalez, G.; Desneux, L.; Cèbe, N.; Gerard, J.F. Ranging behaviour of roe deer in an experimental high-density population: Are females territorial? Comptes Rendus–Biol. 2012, 335, 735–743. [Google Scholar] [CrossRef] [PubMed]
- Apollonio, M.; Putman, R.; Grignolio, S.; Bartoš, L. Hunting Seasons in Relation to Biological Breeding Seasons and the Implications for the Control or Regulation of Ungulate Populations; Cambridge University Press: Cambridge, UK, 2011. [Google Scholar]
- O’Connor, L.J.; Kahn, L.P.; Walkden-Brown, S.W. Moisture requirements for the free-living development of Haemonchus contortus: Quantitative and temporal effects under conditions of low evaporation. Vet. Parasitol. 2007, 150, 128–138. [Google Scholar] [CrossRef] [PubMed]
- Connan, R.M. Hypobiosis in the ostertagids of red deer and the efficacy of ivermectin and fenbendazole against them. Vet. Rec. 1997, 140, 203–205. [Google Scholar] [CrossRef]
- Gibbs, H.C. Hypobiosis in parasitic nematodes—An update. Adv. Parasitol. 1986, 25, 129–174. [Google Scholar] [CrossRef]
- Sargison, N.D.; Wilson, D.J.; Bartley, D.J.; Penny, C.D.; Jackson, F. Haemonchosis and teladorsagiosis in a Scottish sheep flock putatively associated with the overwintering of hypobiotic fourth stage larvae. Vet. Parasitol. 2007, 147, 326–331. [Google Scholar] [CrossRef]
- Allen, A.R.; Ford, T.; Skuce, R.A. Does Mycobacterium tuberculosis var. bovis survival in the environment confound bovine tuberculosis control and eradication? A literature review. Vet. Med. Int. 2021, 2021, 8812898. [Google Scholar] [CrossRef] [PubMed]
- Crispell, J.; Cassidy, S.; Kenny, K.; McGrath, G.; Warde, S.; Cameron, H.; Rossi, G.; MacWhite, T.; White, P.C.; Lycett, S.; et al. Mycobacterium bovis genomics reveals transmission of infection between cattle and deer in Ireland. Microb. Genom. 2020, 6, 7. [Google Scholar] [CrossRef] [PubMed]
- García-Bocanegra, I.; de Val, B.B.; Arenas-Montes, A.; Paniagua, J.; Boadella, M.; Gortázar, C.; Arenas, A. Seroprevalence and risk factors associated to mycobacterium bovis in wild artiodactyl species from southern spain, 2006–2010. PLoS ONE 2012, 7, 2006–2010. [Google Scholar] [CrossRef] [PubMed]
- Kelly, D.J.; Mullen, E.; Good, M. Bovine tuberculosis: The emergence of a new wildlife maintenance host in Ireland. Front. Vet. Sci. 2021, 8, 632525. [Google Scholar] [CrossRef] [PubMed]
- Palmer, M.V.; Waters, W.R.; Whipple, D.L. Investigation of the transmission of Mycobacterium bovis from deer to cattle through indirect contact. Am. J. Vet. Res. 2004, 65, 1483–1489. [Google Scholar] [CrossRef] [PubMed]
- Campbell, E.L.; Byrne, A.W.; Menzies, F.D.; McBride, K.R.; McCormick, C.M.; Scantlebury, M.; Reid, N. Interspecific visitation of cattle and badgers to fomites: A transmission risk for bovine tuberculosis? Ecol. Evol. 2019, 9, 8479–8489. [Google Scholar] [CrossRef] [PubMed]
- Cowie, C.E.; Hutchings, M.R.; Barasona, J.A.; Gortázar, C.; Vicente, J.; White, P.C. Interactions between four species in a complex wildlife: Livestock disease community: Implications for Mycobacterium bovis maintenance and transmission. Eur. J. Wildl. Res. 2016, 62, 51–64. [Google Scholar] [CrossRef]
- Houe, H. Epidemiology of bovine viral diarrhea virus. Vet. Clin. N. Am. Food Anim. Pract. 1995, 11, 521–547. [Google Scholar] [CrossRef]
- Casaubon, J.; Vogt, H.R.; Stalder, H.; Hug, C.; Ryser-Degiorgis, M.P. Bovine viral diarrhea virus in free-ranging wild ruminants in Switzerland: Low prevalence of infection despite regular interactions with domestic livestock. BMC Vet. Res. 2012, 8, 204. [Google Scholar] [CrossRef]
- Gao, Y.; Wang, S.; Du, R.; Wang, Q.; Sun, C.; Wang, N.; Zhang, P.; Zhang, L. Isolation and identification of a bovine viral diarrhea virus from sika deer in China. Virol. J. 2011, 8, 2–7. [Google Scholar] [CrossRef] [PubMed]
- Graham, D.A.; Gallagher, C.; Carden, R.F.; Lozano, J.M.; Moriarty, J.; O’Neill, R. A survey of free-ranging deer in Ireland for serological evidence of exposure to bovine viral diarrhoea virus, bovine herpes virus-1, bluetongue virus and Schmallenberg virus. Ir. Vet. J. 2017, 70, 13. [Google Scholar] [CrossRef]
- Rodríguez-Prieto, V.; Kukielka, D.; Rivera-Arroyo, B.; Martínez-López, B.; de las Heras, A.I.; Sánchez-Vizcaíno, J.M.; Vicente, J. Evidence of shared bovine viral diarrhea infections between red deer and extensively raised cattle in south-central Spain. BMC Vet. Res. 2016, 12, 1–11. [Google Scholar] [CrossRef]
- Medlock, J.M.; Hansford, K.M.; Bormane, A.; Derdakova, M.; Estrada-Peña, A.; George, J.C.; Golovljova, I.; Jaenson, T.G.; Jensen, J.K.; Jensen, P.M.; et al. Driving forces for changes in geographical distribution of Ixodes ricinus ticks in Europe. Parasites Vectors 2013, 6, 1. [Google Scholar] [CrossRef] [PubMed]
- Lacasta, D.; Lorenzo, M.; González, J.M.; Ruiz de Arcaute, M.; Benito, A.Á.; Baselga, C.; Milian, M.E.; Lorenzo, N.; Jiménez, C.; Villanueva-Saz, S.; et al. Epidemiological study related to the first outbreak of ovine anaplasmosis in spain. Animals 2021, 11, 2036. [Google Scholar] [CrossRef]
- Matei, I.A.; Estrada-Peña, A.; Cutler, S.J.; Vayssier-Taussat, M.; Varela-Castro, L.; Potkonjak, A.; Zeller, H.; Mihalca, A.D. A review on the eco-epidemiology and clinical management of human granulocytic anaplasmosis and its agent in Europe. Parasites Vectors 2019, 12, 599. [Google Scholar] [CrossRef]
- Robinson, M.T.; Shaw, S.E.; Morgan, E.R. Anaplasma phagocytophilum infection in a multi-species deer community in the New Forest, England. Eur. J. Wildl. Res. 2009, 55, 439–442. [Google Scholar] [CrossRef]
- Woldehiwet, Z. Anaplasma phagocytophilum in ruminants in Europe. Ann. N. Y. Acad. Sci. 2006, 1078, 446–460. [Google Scholar] [CrossRef]
- Grassi, L.; Franzo, G.; Martini, M.; Mondin, A.; Cassini, R.; Drigo, M.; Pasotto, D.; Vidorin, E.; Menandro, M.L. Ecotyping of Anaplasma Phagocytophilum from wild ungulates and ticks shows circulation of zoonotic strains in northeastern Italy. Animals 2021, 11, 310. [Google Scholar] [CrossRef]
- Brown, T.L.; Airs, P.M.; Porter, S.; Caplat, P.; Morgan, E.R. Understanding the role of wild ruminants in anthelmintic resistance in livestock. Biol. Lett. 2022, 18, 20220057. [Google Scholar] [CrossRef]
Figure 1.
Review process used to gather data for meta-analysis and the number of papers returned, including (a) structure of search terms used to identify relevant papers on Google Scholar and Science Direct. Equivalent searches were also carried out using The Global Mammal Parasite Database. (b) Filtering process to identify suitable papers which included abomasal nematode prevalence data. Scientific names for deer species were included alongside the common English names.
Figure 1.
Review process used to gather data for meta-analysis and the number of papers returned, including (a) structure of search terms used to identify relevant papers on Google Scholar and Science Direct. Equivalent searches were also carried out using The Global Mammal Parasite Database. (b) Filtering process to identify suitable papers which included abomasal nematode prevalence data. Scientific names for deer species were included alongside the common English names.
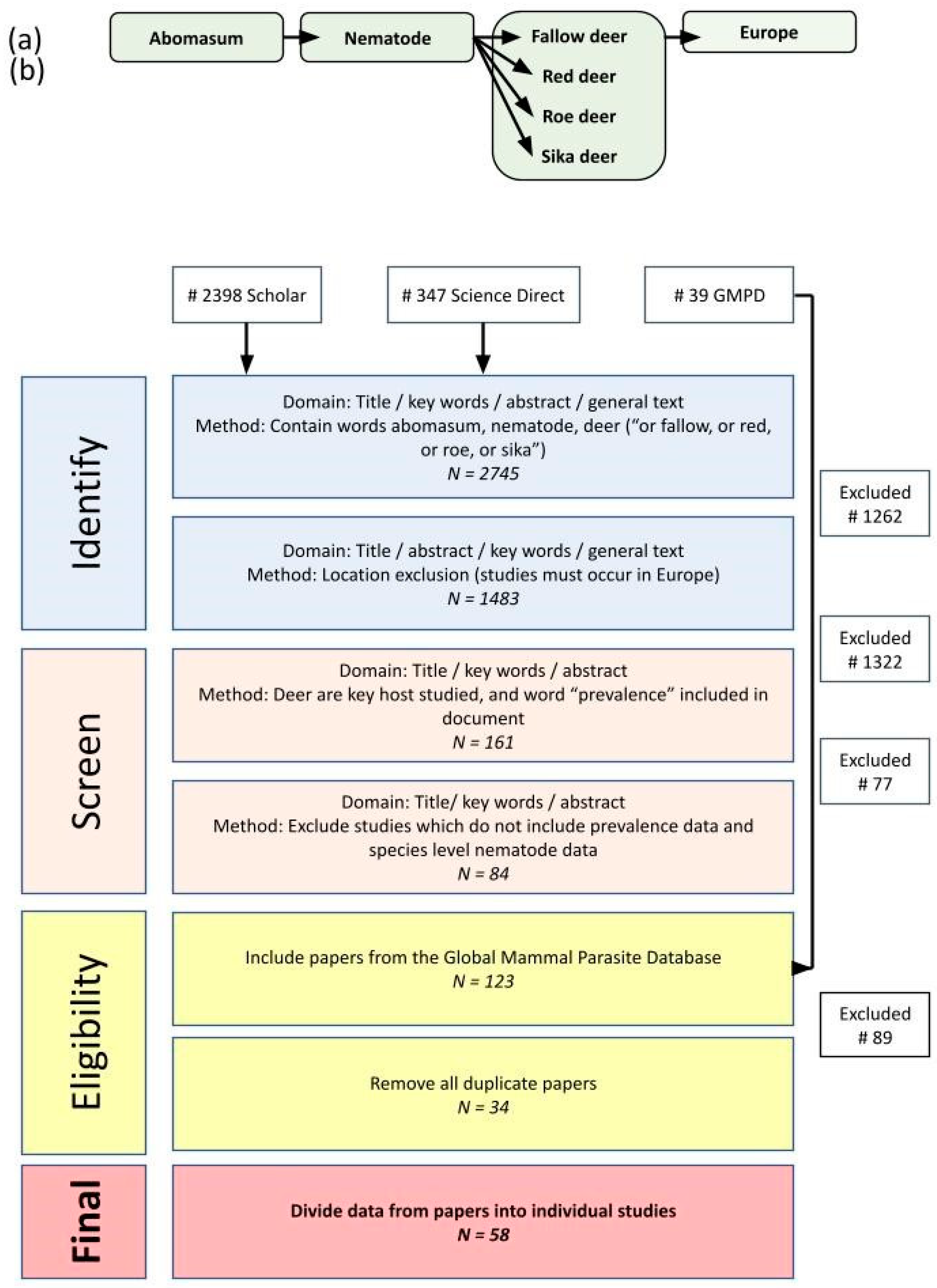
Figure 2.
Estimated locations of deer helminth studies used for this meta-analysis. Note that points may be slightly offset to improve the visibility of multiple studies in one area.
Figure 2.
Estimated locations of deer helminth studies used for this meta-analysis. Note that points may be slightly offset to improve the visibility of multiple studies in one area.

Figure 3.
Heatmap showing the prevalence of helminths in deer species across studies in Europe. White space indicates where no data were available. Small intestine—Nematodirus battus, Nematodirus filicollis, Nematodirus helvetianus, Nematodirus roscidus, Nematodirus europaeus, Capillaria bovis, Trichostrongylus colubriformis, Trichostrongylus vitrinus, Cooperia pectinata, Cooperia oncophora and Trichostrongylus capricola. Large intestine—Oesophagostornum venulosum, Oesophagostornum radiatum, Oesophagostornum sikae, Trichuris ovis, Chabertia ovina, Trichuris capreoli and Trichuris globulosa. Liver—Fasciola hepatica, Dicrocoelium chinensis, Dicrocoelium dendriticum and Fascioloides magna. Lungs—Varestrongylus sagittatus, Dictyocaulus capreolus, Dictyocaulus eckerti and Dictyocaulus noerneri. Abomasal species associated primarily with deer (Spiculopteragia spp. and O. leptospicularis) are grouped together. [4,5,9,10,11,12,13,14,21,29,30,31,32,33,34,35,36,37,38,39,40,41,42,43,44,45,46,47,48,49,50,51,52,53,54].
Figure 3.
Heatmap showing the prevalence of helminths in deer species across studies in Europe. White space indicates where no data were available. Small intestine—Nematodirus battus, Nematodirus filicollis, Nematodirus helvetianus, Nematodirus roscidus, Nematodirus europaeus, Capillaria bovis, Trichostrongylus colubriformis, Trichostrongylus vitrinus, Cooperia pectinata, Cooperia oncophora and Trichostrongylus capricola. Large intestine—Oesophagostornum venulosum, Oesophagostornum radiatum, Oesophagostornum sikae, Trichuris ovis, Chabertia ovina, Trichuris capreoli and Trichuris globulosa. Liver—Fasciola hepatica, Dicrocoelium chinensis, Dicrocoelium dendriticum and Fascioloides magna. Lungs—Varestrongylus sagittatus, Dictyocaulus capreolus, Dictyocaulus eckerti and Dictyocaulus noerneri. Abomasal species associated primarily with deer (Spiculopteragia spp. and O. leptospicularis) are grouped together. [4,5,9,10,11,12,13,14,21,29,30,31,32,33,34,35,36,37,38,39,40,41,42,43,44,45,46,47,48,49,50,51,52,53,54].
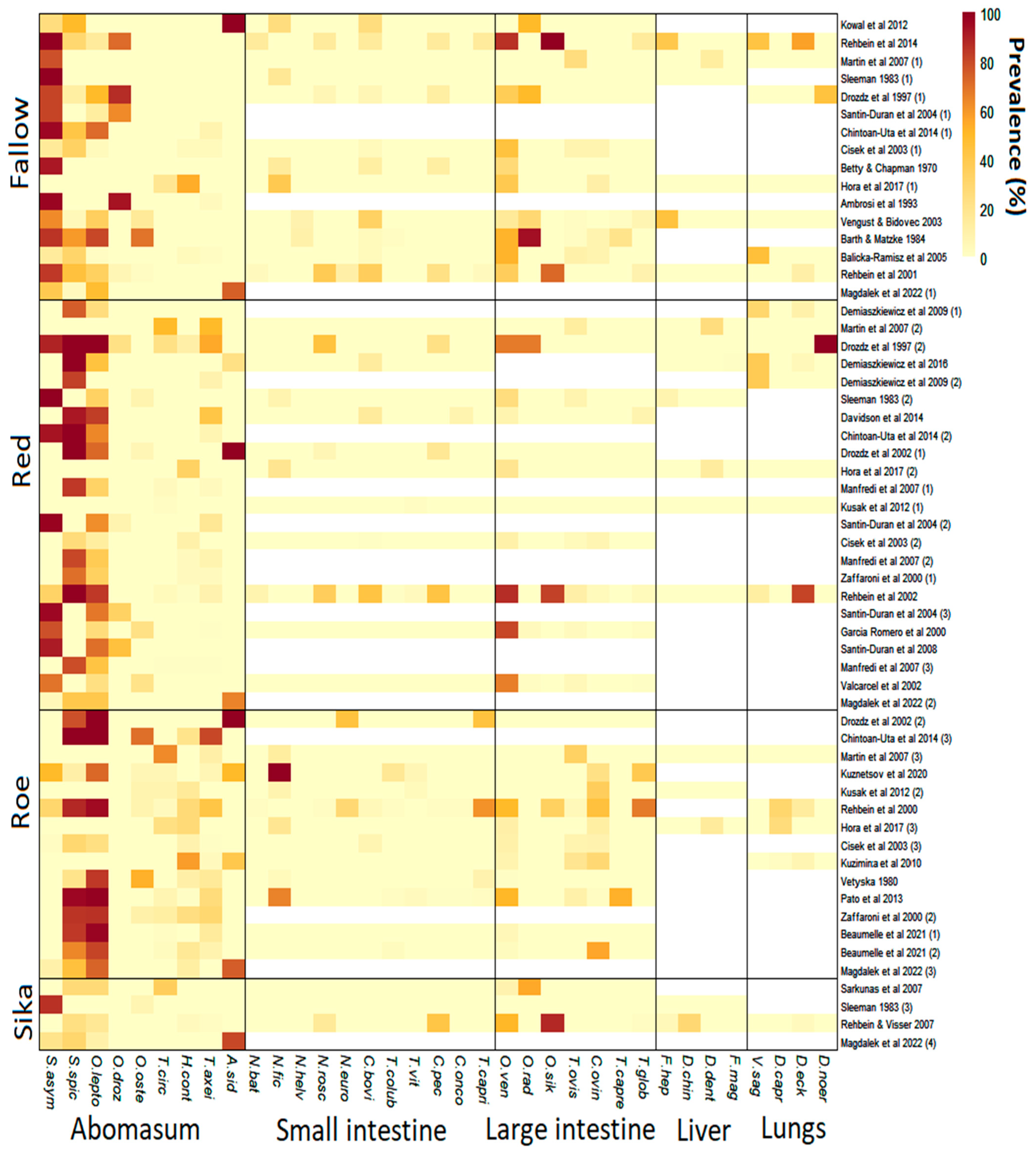
Figure 4.
Violin plot of the prevalence of abomasal nematodes in fallow, red, roe and sika deer from studies with different levels of livestock pasture contact. Pairwise Wilcoxon tests (with Bonferroni correction) show a higher probability (p = 0.01) of infection in “likely” contact scenario vs. “unlikely” scenarios in studies with O. ostertagi, T. circumcincta, H. contortus and T. axei infection. No difference (p = 1) was found in studies with S. asymmetrica, S. spiculoptera, O. leptospicularis and O. drozdi under “likely” or “unlikely” livestock pasture contact scenarios.
Figure 4.
Violin plot of the prevalence of abomasal nematodes in fallow, red, roe and sika deer from studies with different levels of livestock pasture contact. Pairwise Wilcoxon tests (with Bonferroni correction) show a higher probability (p = 0.01) of infection in “likely” contact scenario vs. “unlikely” scenarios in studies with O. ostertagi, T. circumcincta, H. contortus and T. axei infection. No difference (p = 1) was found in studies with S. asymmetrica, S. spiculoptera, O. leptospicularis and O. drozdi under “likely” or “unlikely” livestock pasture contact scenarios.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Differences in the prevalence of abomasal nematodes in fallow, red and roe deer from studies across Europe. Note that O. ostertagi, Te. Circumcincta, H. contortus and T. axei are commonly found in livestock. Bonferroni correction multiplied the p-value by 3 (= the number of univariate comparisons conducted), reported rounded to 2 decimal places.
Table 1.
Differences in the prevalence of abomasal nematodes in fallow, red and roe deer from studies across Europe. Note that O. ostertagi, Te. Circumcincta, H. contortus and T. axei are commonly found in livestock. Bonferroni correction multiplied the p-value by 3 (= the number of univariate comparisons conducted), reported rounded to 2 decimal places.
| p-Values with Bonferroni Correction | |||
|---|---|---|---|
| Nematodes | Fallow-Red (n = 39) | Fallow-Roe (n = 31) | Red-Roe (n = 38) |
| Spiculopteragia asymmetrica | 0.08 | <0.001 * | 0.74 |
| Spiculopteragia spiculoptera | 0.61 | 0.48 | 1.00 |
| Ostertagia leptospicularis | 0.40 | 0.10 | 0.61 |
| Ostertagia drozdi | 1.00 | 0.11 | 0.49 |
| Ostertagia ostertagi | 1.00 | 1.00 | 0.74 |
| Teladorsagia circumcincta | 0.81 | 0.02 ** | 0.19 |
| Haemonchus contortus | 1.00 | 0.02 ** | 0.01 ** |
| Trichostrongylus axei | 0.35 | 0.01 ** | 0.86 |
| Ashworthius sidemi | 1.00 | 1.00 | 1.00 |
* asterisk—fallow deer have a significantly higher prevalence. ** asterisks—roe deer have a significantly higher prevalence.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Brown, T.L.; Morgan, E.R. Helminth Prevalence in European Deer with a Focus on Abomasal Nematodes and the Influence of Livestock Pasture Contact: A Meta-Analysis. Pathogens 2024, 13, 378. https://doi.org/10.3390/pathogens13050378
AMA Style
Brown TL, Morgan ER. Helminth Prevalence in European Deer with a Focus on Abomasal Nematodes and the Influence of Livestock Pasture Contact: A Meta-Analysis. Pathogens. 2024; 13(5):378. https://doi.org/10.3390/pathogens13050378
Chicago/Turabian StyleBrown, Tony L., and Eric R. Morgan. 2024. "Helminth Prevalence in European Deer with a Focus on Abomasal Nematodes and the Influence of Livestock Pasture Contact: A Meta-Analysis" Pathogens 13, no. 5: 378. https://doi.org/10.3390/pathogens13050378
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.