
Fasciola hepatica: Histology of the Reproductive Organs and Differential Effects of Triclabendazole on Drug-Sensitive and Drug-Resistant Fluke Isolates and on Flukes from Selected Field Cases

Abstract
:1. Introduction
2. Methods and Materials
2.1. Sources of Flukes
2.1.1. Experimental Isolates

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| F. hepatica Infection | Number of Sheep Treated with 10 mg/kg TCBZ | Number of Sheep Receiving no Treatment | Time Post-Treatment at Which Flukes were Collected: | |||
|---|---|---|---|---|---|---|
| 24 h | 48 h | 72 h | 96 h | |||
| TCBZ-S Cullompton 250 mc (6 sheep) | 4 | 2 | 1 | 1 | 1 | 1 |
| 2 | ||||||
| TCBZ-R Sligo 250 mc (4 sheep) | 2 | 2 | 2 | |||
| 2 | ||||||
2.1.2. Field Cases with Suspected TCBZ- Resistance
2.1.3. Rats Infected with Metacercariae from Bovine Field Case
2.2. Preparation of Flukes for Histology
2.3. In Situ Hybridization Method to Demonstrate Apoptosis
2.4. Preparation and Examination of Stained Whole-Mounts
3. Results
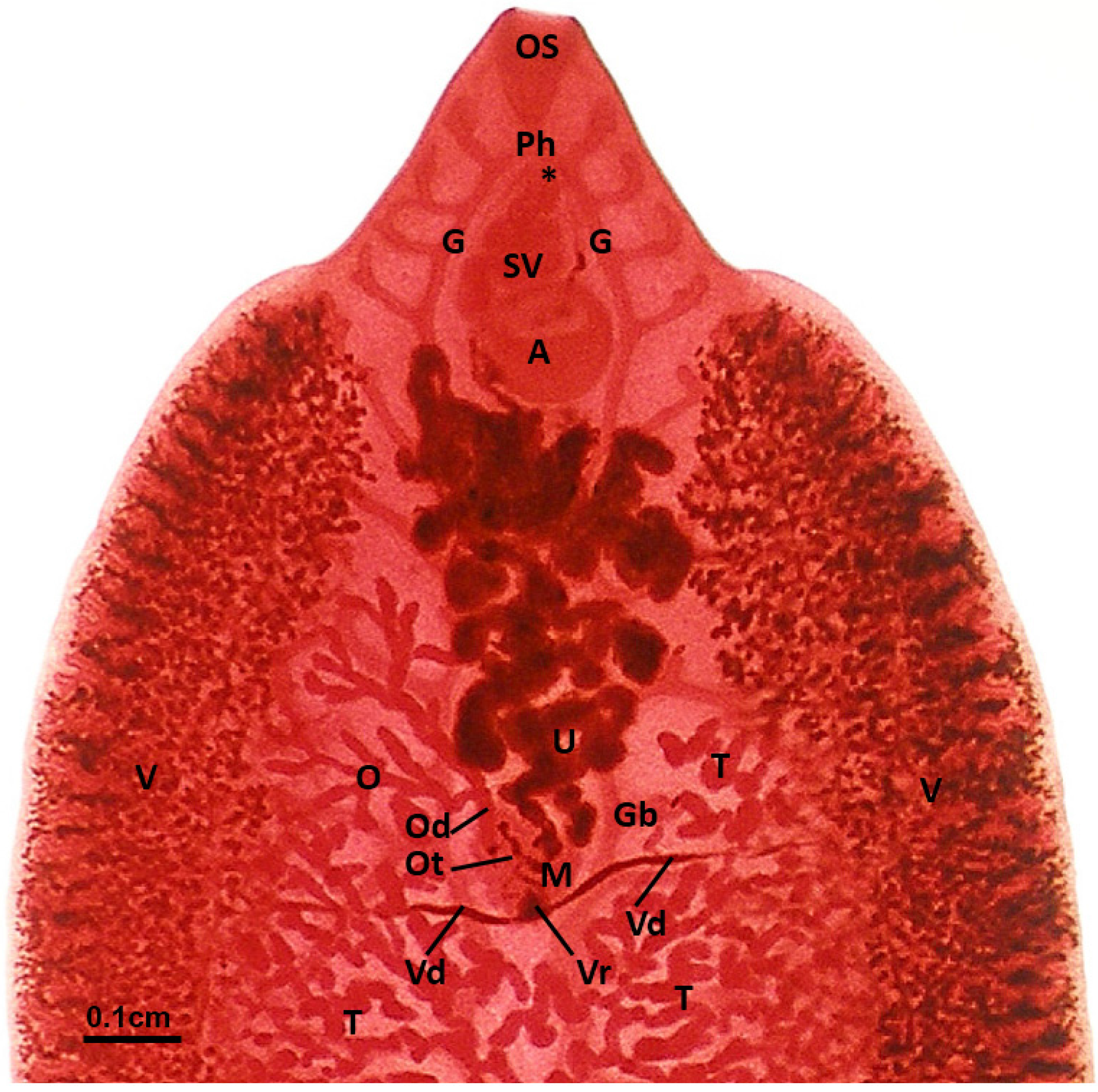
3.1. Arrangement of the Gonads, Accessory Organs and Ducts

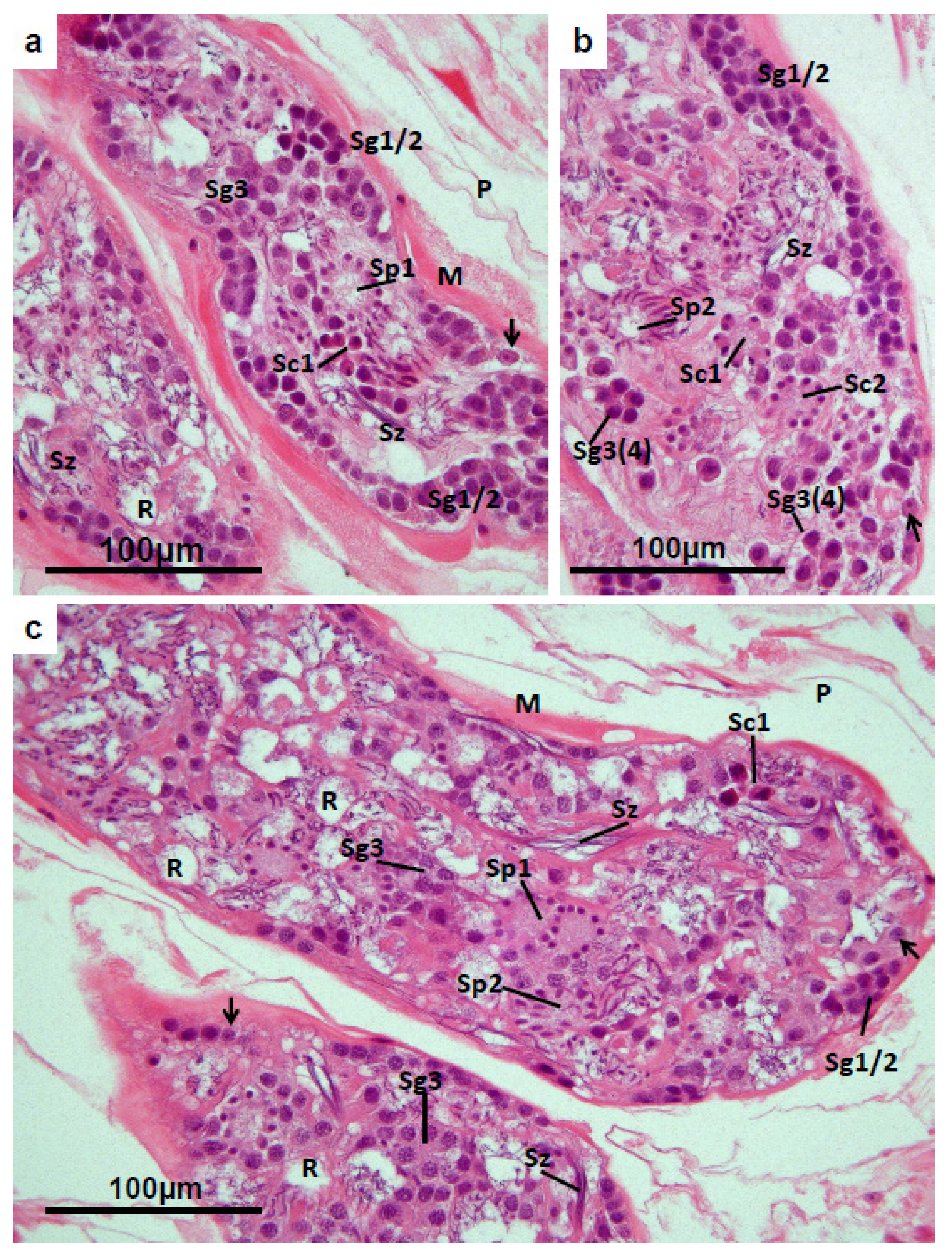
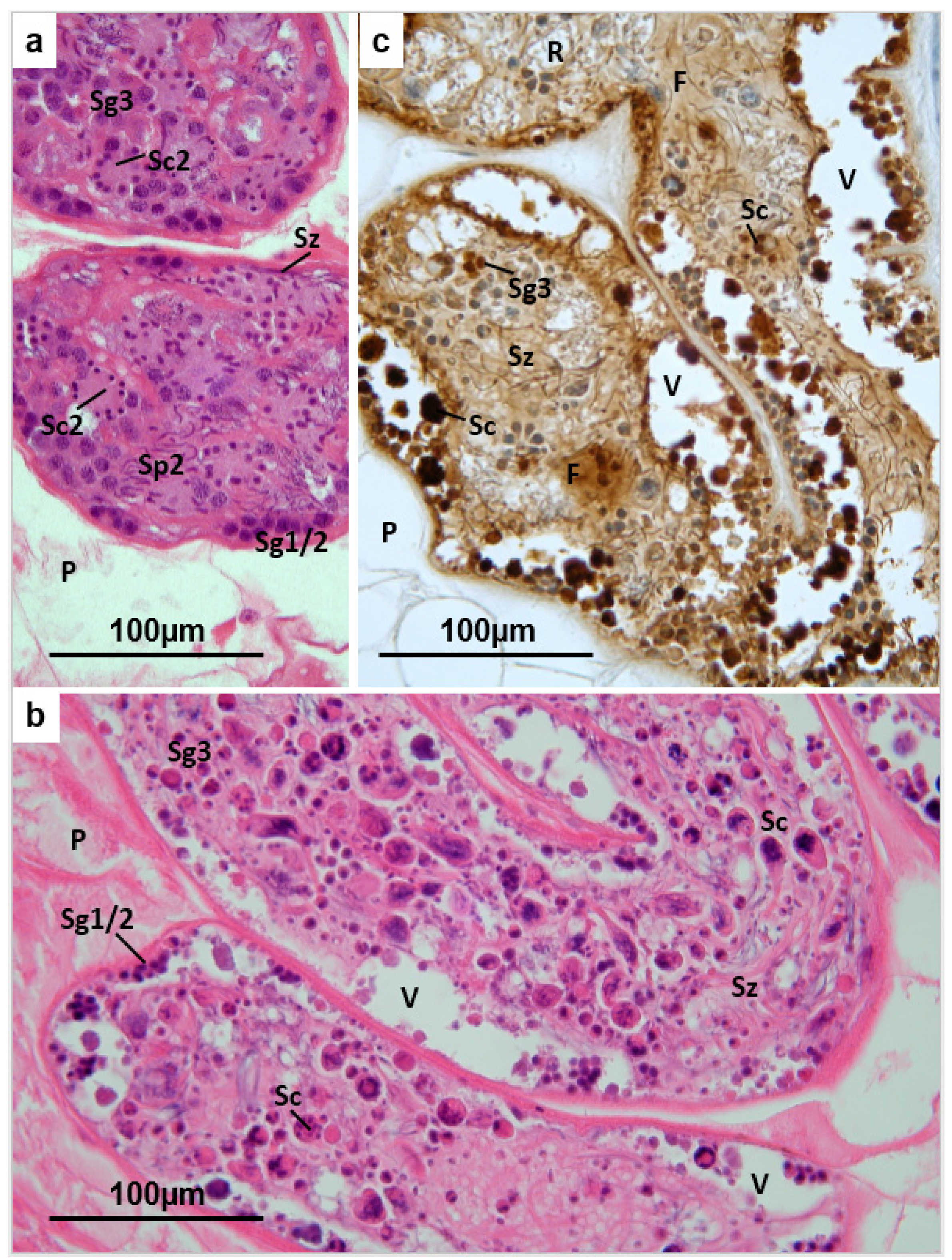
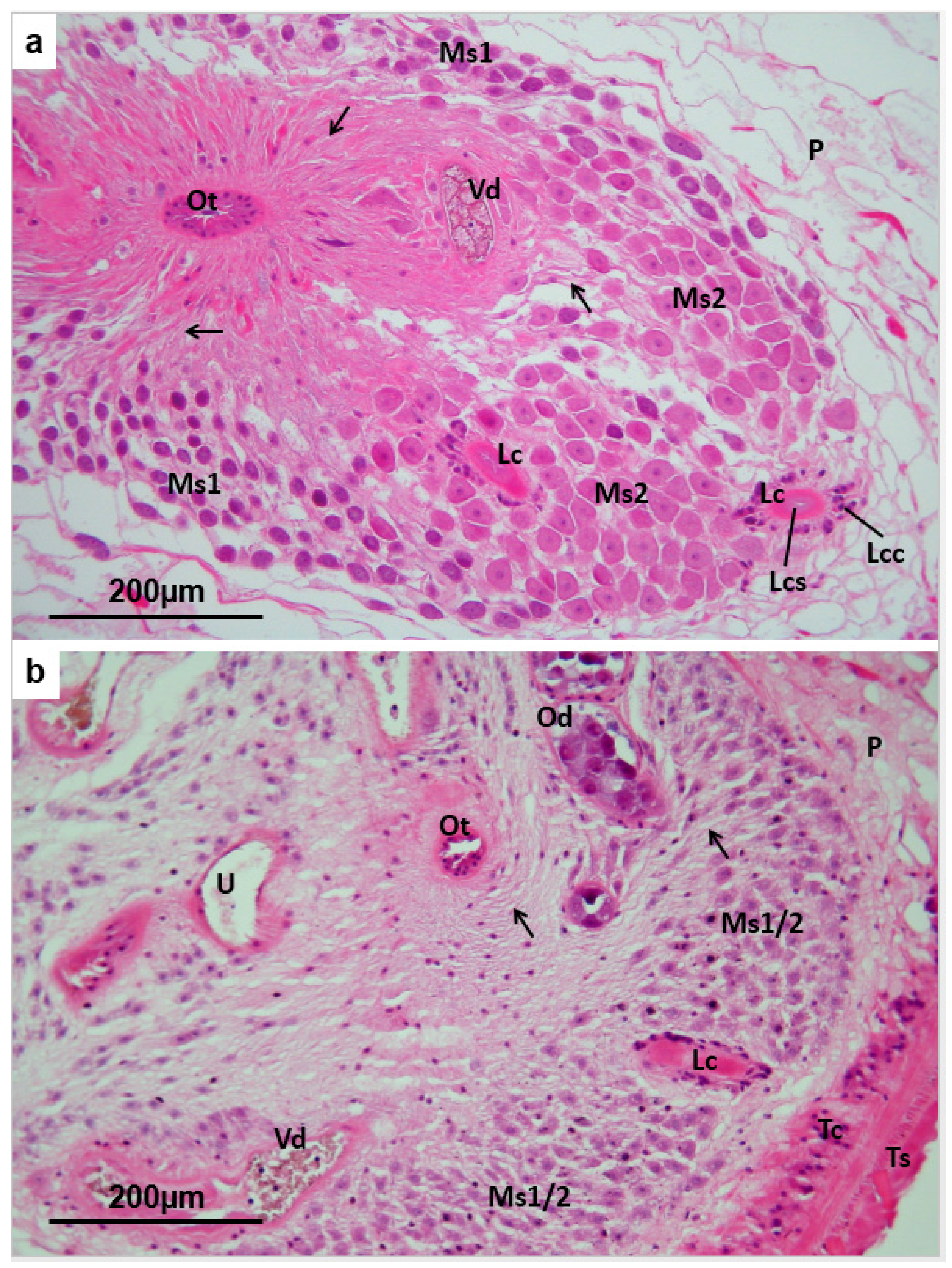
3.2. Testis


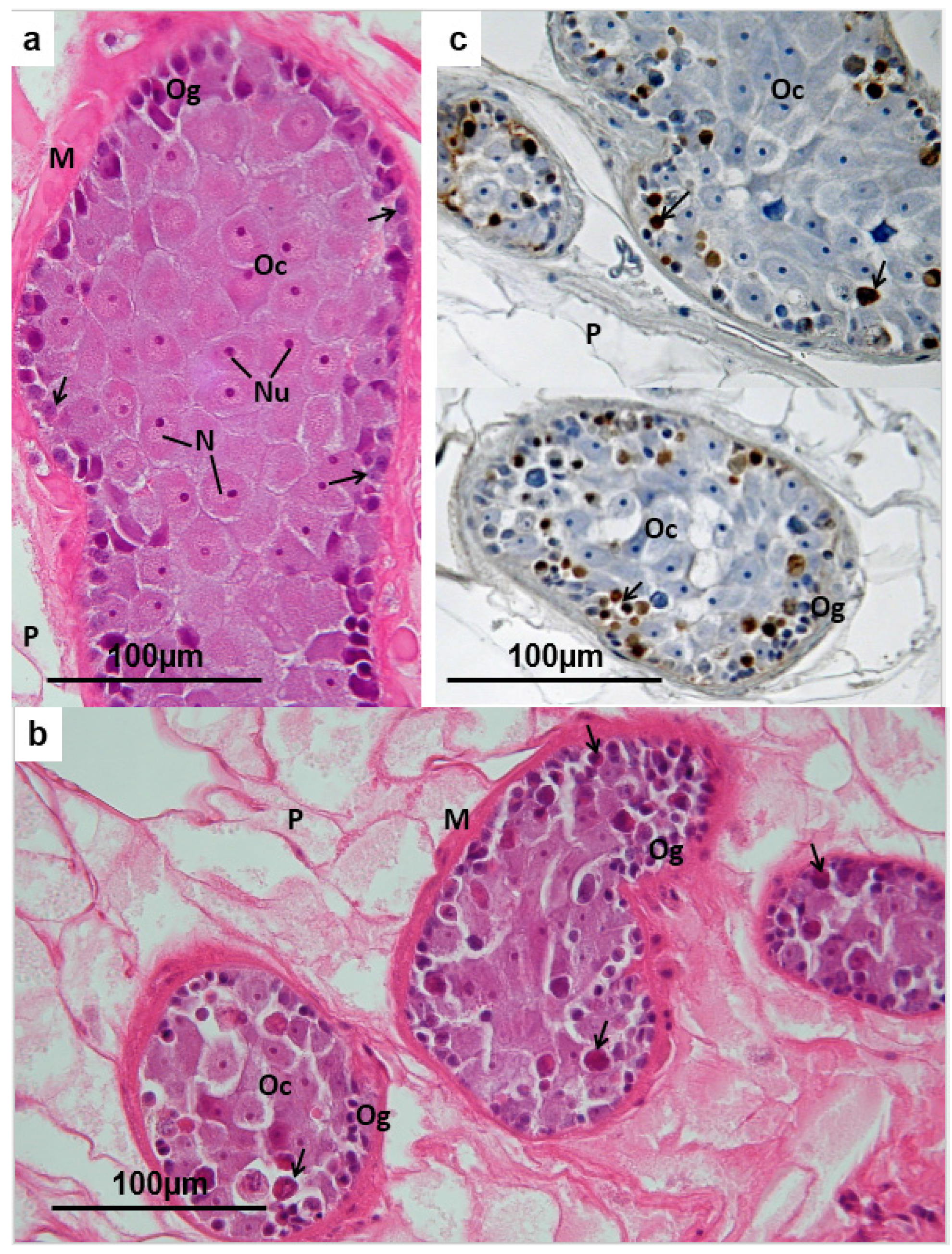
3.3. Ovary
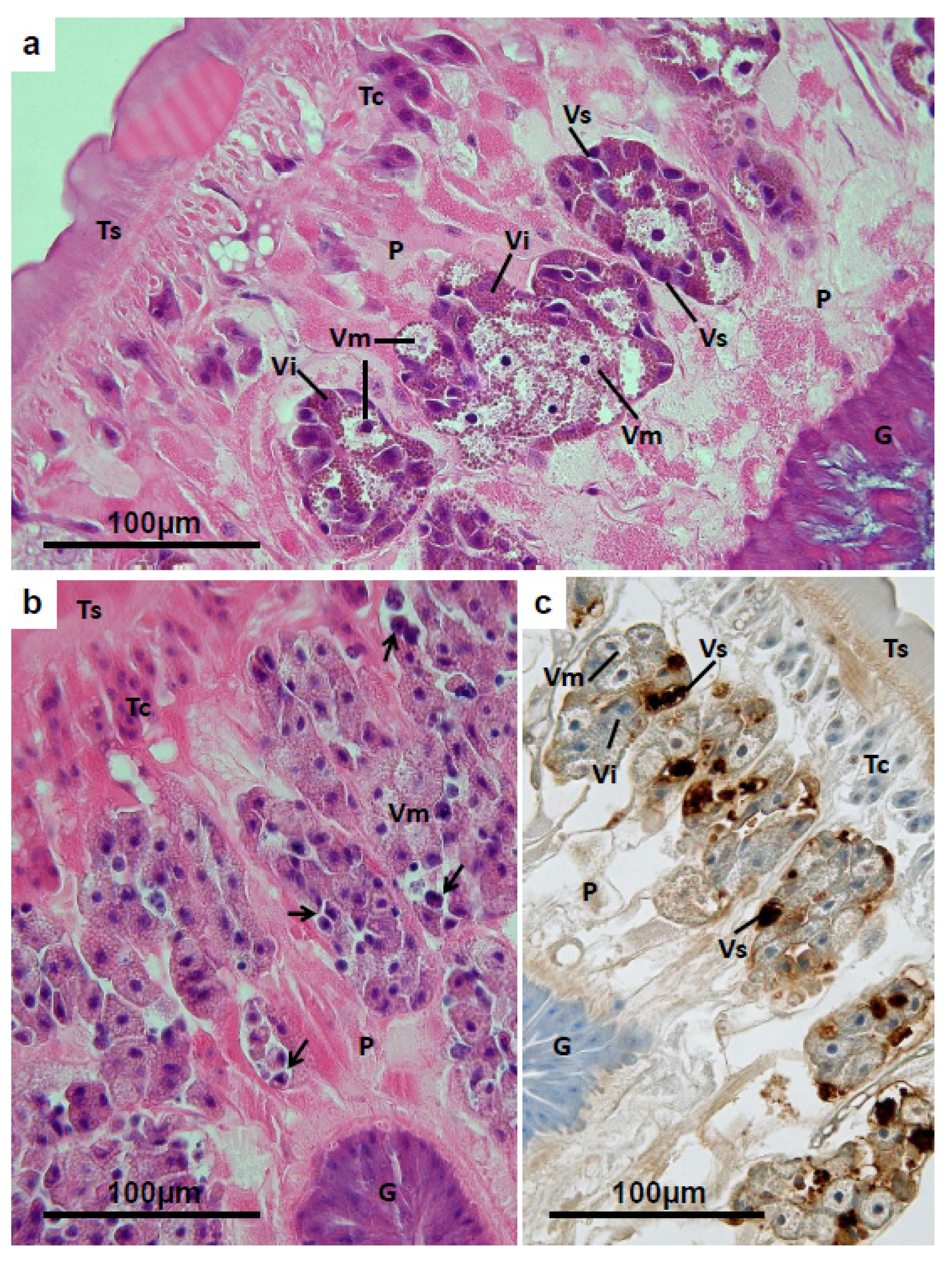
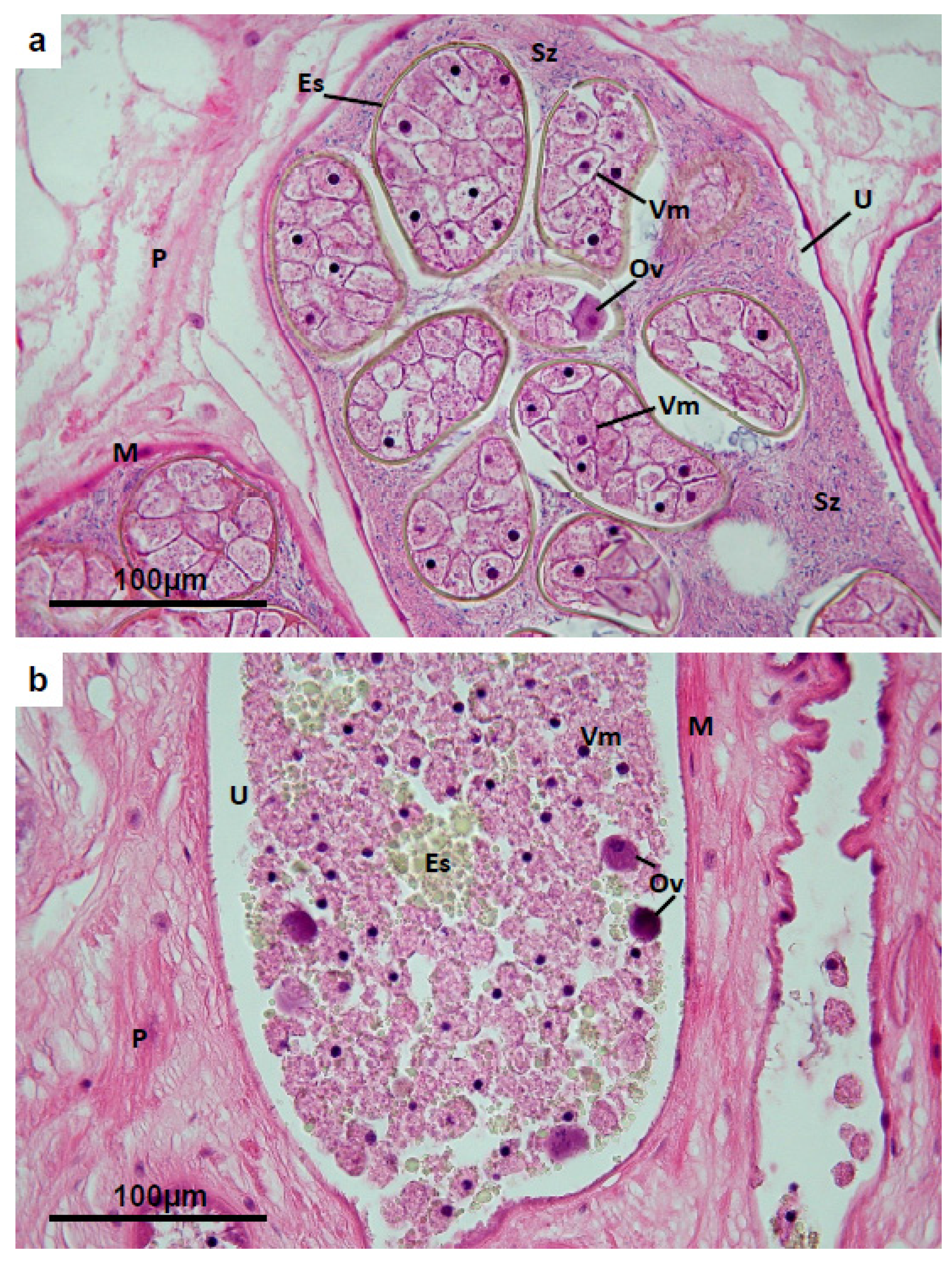
3.4. Vitelline Follicles


3.5. Mehlis’ Gland
3.6. Uterus


4. Discussion
Acknowledgments
Conflicts of Interest
References
- Leuckart, K.G.F.R. Zur Entwickelungsgeschichte des Leberegels. Zweite Mitt. Zool. Anz. 1882, 5, 524–528. (In German) [Google Scholar]
- Lutz, A. Zur Lebensgeschichte des Distoma hepaticum. Zentralbl. Bakteriol. Parasitenkd. Infektionskr. Abteilung 1892, 11, 783–796. (In German) [Google Scholar]
- Lutz, A. Weiteres zur Lebensgeschichte des Distoma hepaticum. Zentralbl. Bakteriol. Parasitenkd. Infektionskr. Abteilung 1893, 13, 320–328. (In German) [Google Scholar]
- Andrews, S.J. The Life Cycle of Fasciola hepatica. In Fasciolosis; Dalton, J.P., Ed.; CABI Publishing: Wallingford, Oxon, UK, 1999; Chapter 1; pp. 1–29. [Google Scholar]
- Hanna, R.E.B.; Cromie, L.; Taylor, S.M.; Couper, A. The effect of a parenteral ivermectin/closantel injection on the growth and reproductive development of Fasciola hepatica in cattle. Vet. Parasitol. 2006, 142, 78–90. [Google Scholar] [CrossRef] [PubMed]
- McConville, M.; Brennan, G.P.; Flanagan, A.; Edgar, H.W.J.; Hanna, R.E.B.; McCoy, M.; Gordon, A.W.; Castillo, R.; Hernández-Campos, A.; Fairweather, I. An evaluation of the efficacy of compound alpha and triclabendazole against two isolates of Fasciola hepatica. Vet. Parasitol. 2009, 162, 75–88. [Google Scholar] [CrossRef] [PubMed]
- McConville, M.; Hanna, R.E.B.; Brennan, G.P.; Edgar, H.W.J.; McConnell, S.; McCoy, M.; Castillo, R.; Hernandez-Campos, A.; Fairweather, I. Impact of compound alpha treatment in vivo on egg production by the liver fluke, Fasciola hepatica. Vet. Parasitol. 2012, 187, 183–195. [Google Scholar] [CrossRef] [PubMed]
- Happich, F.A.; Boray, J.C. Quantitative diagnosis of Chronic fascioliosis. 2. The estimation of daily total egg production of Fasciola hepatica and the number of adult flukes in sheep by faecal egg counts. Aust. Vet. J. 1969, 45, 329–331. [Google Scholar] [CrossRef] [PubMed]
- Lagrange, E.; Gutmann, A. Sur l’infestation experimentale de la souris par Fasciola hepatica. Riv. Parassitol. 1961, 22, 93–101. (In French) [Google Scholar]
- Hanna, R.E.B.; Gordon, A.; Moffett, D.; Edgar, H.; McConnell, S.; Brennan, G.P.; Fairweather, I. Fasciola hepatica: An analysis of the relative influence of the presence of co-specifics and the activity of the host’s defense mechanisms on the growth and reproductive development of flukes in experimental infections in rats. Vet. Parasitol. 2011, 178, 251–263. [Google Scholar] [CrossRef] [PubMed]
- Hanna, R.E.B.; Edgar, H.; Moffett, D.; McConnell, S.; Fairweather, I.; Brennan, G.P.; Trudgett, A.; Hoey, E.M.; Cromie, L.; Taylor, S.M.; et al. Fasciola hepatica: Histology of the testis in egg-producing adults of several laboratory-maintained isolates of flukes grown to maturity in cattle and sheep and in flukes from naturally-infected hosts. Vet. Parasitol. 2008, 157, 222–234. [Google Scholar] [CrossRef] [PubMed]
- Fairweather, I.; Boray, J.C. Mechanisms of fasciolicide action and drug resistance in Fasciola hepatica. In Fasciolosis; Dalton, J.P., Ed.; CABI Publishing: Wallingford, UK, 1999; Chapter 7; pp. 225–276. [Google Scholar]
- Fairweather, I. Triclabendazole: New skills to unravel an old(ish) enigma. J. Helminthol. 2005, 79, 227–234. [Google Scholar] [CrossRef] [PubMed]
- Boray, J.C.; Crowfoot, P.D.; Strong, M.B.; Allison, J.R.; Schellenbaum, M.; von Orelli, M.; Sarasin, G. Treatment of immature and mature Fasciola hepatica infections in sheep with triclabendazole. Vet. Rec. 1983, 113, 315–317. [Google Scholar] [CrossRef] [PubMed]
- Overend, D.J.; Bowen, F.L. Resistance of Fasciola hepatica to triclabendazole. Aust. Vet. J. 1995, 72, 275–276. [Google Scholar] [CrossRef] [PubMed]
- Fairweather, I. Triclabendazole progress report, 2005–2009: An advancement of learning? J. Helminthol. 2009, 83, 139–150. [Google Scholar] [CrossRef] [PubMed]
- Stitt, A.W.; Fairweather, I. Fasciola hepatica: Tegumental surface changes in adult and juvenile flukes following treatment in vitro with the sulphoxide metabolite of triclabendazole (“Fasinex”). Parasitol. Res. 1993, 79, 529–536. [Google Scholar] [CrossRef] [PubMed]
- Meaney, M.; Fairweather, I.; Brennan, G.P.; Ramasamy, P.; Subramanian, P.B. Fasciola gigantica: Tegumental surface alterations following treatment in vitro with the sulphoxide metabolite of triclabendazole. Parasitol. Res. 2002, 88, 315–325. [Google Scholar] [CrossRef] [PubMed]
- Walker, S.M.; McKinstry, B.; Boray, J.C.; Brennan, G.P.; Trudgett, A.; Hoey, E.M.; Fletcher, H.; Fairweather, I. Response of two isolates of Fasciola hepatica to treatment with triclabendazole in vivo and in vitro. Parasitol. Res. 2004, 94, 427–438. [Google Scholar] [CrossRef] [PubMed]
- McConville, M.; Brennan, G.P.; McCoy, M.; Castillo, R.; Hernández -Campos, A.; Ibarra, F.; Fairweather, I. Adult triclabendazole-resistant Fasciola hepatica: Surface and subsurface tegumental responses to in vitro treatment with the sulphoxide metabolite of the experimental fasciolicide compound alpha. Parasitology 2006, 133, 195–208. [Google Scholar] [CrossRef] [PubMed]
- McConville, M.; Brennan, G.P.; McCoy, M.; Castillo, R.; Hernández -Campos, A.; Ibarra, F.; Fairweather, I. Immature triclabendazole-resistant Fasciola hepatica: Tegumental responses to in vitro treatment with the sulphoxide metabolite of the experimental fasciolicide compound alpha. Parasitol. Res. 2007, 100, 365–377. [Google Scholar] [CrossRef] [PubMed]
- Halferty, L.; Brennan, G.P.; Hanna, R.E.B.; Edgar, H.W.; Meaney, M.M.; McConville, M.; Trudgett, A.; Hoey, L.; Fairweather, I. Tegumental surface changes in juvenile Fasciola hepatica in response to treatment in vivo with triclabendazole. Vet. Parasitol. 2008, 115, 49–58. [Google Scholar] [CrossRef] [PubMed]
- Halferty, L.; Brennan, G.P.; Trudgett, A.; Hoey, L.; Fairweather, I. Relative activity of triclabendazole metabolites against the liver fluke, Fasciola hepatica. Vet. Parasitol. 2009, 159, 126–138. [Google Scholar] [CrossRef] [PubMed]
- McConville, M.; Brennan, G.P.; Flanagan, A.; Edgar, H.W.J.; McCoy, M.; Castillo, R.; Herna’ndez-Campos, A.; Fairweather, I. Surface and internal tegumental changes in juvenile Fasciola hepatica following treatment in vivo with the experimental fasciolicide, compound alpha. Vet. Parasitol. 2008, 153, 52–64. [Google Scholar] [CrossRef] [PubMed]
- McConville, M.; Brennan, G.P.; Edgar, H.W.J.; Castillo, R.; Hernández—Campos, A.; Fairweather, I. Ultrastructural changes to the tegumental system and the gastrodermal cells in adult Fasciola hepatica following in vivo treatment with the experimental fasciolicide, compound alpha. Parasitology 2009, 136, 655–680. [Google Scholar] [CrossRef] [PubMed]
- McConville, M.; Brennan, G.P.; Flanagan, A.; Hanna, R.E.B.; Edgar, H.W.J.; Castillo, R.; Hernández-Campos, A.; Fairweather, I. Surface changes in adult Fasciola hepatica following treatment in vivo with the experimental fasciolicide, compound alpha. Parasitol. Res. 2009, 105, 757–767. [Google Scholar] [CrossRef] [PubMed]
- McConville, M.; Hanna, R.E.B.; Brennan, G.P.; McCoy, M.; Edgar, H.W.J.; McConnell, S.; Castillo, R.; Hernández-Campos, A.; Fairweather, I. Fasciola hepatica: Disruption of spermatogenesis by the fasciolicide, compound alpha. Parasitol. Res. 2010, 106, 311–323. [Google Scholar] [CrossRef] [PubMed]
- Flanagan, A.; Edgar, H.W.J.; Gordon, A.; Hanna, R.E.B.; Brennan, G.P.; Fairweather, I. Comparison of two assays, a faecal egg count reduction test (FECRT) and a coproantigen reduction test (CRT), for the diagnosis of resistance to triclabendazole in Fasciola hepatica in sheep. Vet. Parasitol. 2011, 176, 170–176. [Google Scholar] [CrossRef] [PubMed]
- Flanagan, A.M.; Edgar, H.W.J.; Forster, F.; Gordon, A.; Hanna, R.E.B.; McCoy, M.; Brennan, G.P.; Fairweather, I. Standardisation of a coproantigen reduction test (CRT) protocol for the diagnosis of resistance to triclabendazole in Fasciola hepatica. Vet. Parasitol. 2011, 176, 34–42. [Google Scholar] [CrossRef] [PubMed]
- Hanna, R.E.; McMahon, C.; Ellison, S.; Edgar, H.W.; Kajugu, P.-E.; Gordon, A.; Irwin, D.; Barley, J.P.; Malone, F.E.; Brennan, G.P.; et al. Fasciola hepatica: A comparative survey of adult fluke resistance to triclabendazole, nitroxynil and closantel on selected upland and lowland sheep farms in Northern Ireland using faecal egg counting, coproantigen ELISA testing and fluke histology. Vet. Parasitol. 2015, 207, 34–43. [Google Scholar] [CrossRef] [PubMed]
- Hanna, R.E.B.; Edgar, H.W.J.; McConnell, S.; Toner, E.; McConville, M.; Brennan, G.P.; Devine, C.; Flanagan, A.; Halferty, L.; Meaney, M.; et al. Fasciola hepatica: Histological changes in the reproductive structures of triclabendazole (TCBZ)-sensitive and TCBZ-resistant flukes collected from experimentally-infected sheep one to four days after treatment with TCBZ and the related benzimidazole derivative, Compound Alpha. Vet. Parasitol. 2010, 168, 240–254. [Google Scholar] [PubMed]
- Fairweather, I. Liver fluke isolates: A question of provenance. Vet. Parasitol. 2011, 176, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Hennessy, D.R.; Lacey, E.; Steel, J.W.; Prichard, R.K. The kinetics of triclabendazole disposition in sheep. J. Vet. Pharmacol. Ther. 1987, 10, 64–72. [Google Scholar] [CrossRef] [PubMed]
- Hanna, R.E.B.; Scarcella, S.; Solana, H.; McConnell, S.; Fairweather, I. Early onset of changes to the reproductive system of Fasciola hepatica following in vivo treatment with triclabendazole. Vet. Parasitol. 2012, 184, 341–347. [Google Scholar] [CrossRef] [PubMed]
- Toner, E.; Brennan, G.P.; Hanna, R.E.B.; Edgar, H.W.J.; Fairweather, I. Fasciola hepatica: Time-dependent disruption of spermatogenesis following in vivo treatment with triclabendazole. Parasitol. Res. 2011, 109, 1035–1043. [Google Scholar] [CrossRef] [PubMed]
- Toner, E.; Brennan, G.P.; Hanna, R.E.B.; Edgar, H.W.J.; Fairweather, I. Disruption of egg formation by Fasciola. hepatica following treatment in vivo with triclabendazole in the sheep host. Vet. Parasitol. 2011, 177, 79–89. [Google Scholar] [CrossRef] [PubMed]
- Stitt, A.W.; Fairweather, I. Spermatogenesis and the fine structure of the mature spermatozoon of the liver fluke, Fasciola hepatica (Trematoda: Digenea). Parasitology 1990, 101, 395–407. [Google Scholar] [CrossRef] [PubMed]
- Hanna, R.E.B.; Moffett, D.; Brennan, G.P.; Fairweather, I. Fasciola hepatica: A light and electron microscope study of sustentacular tissue and heterophagy in the testis. Vet. Parasitol. 2012, 187, 168–182. [Google Scholar] [CrossRef] [PubMed]
- Hanna, R.E.B.; Forster, F.I.; Brennan, G.P.; Fairweather, I. Fasciola hepatica: Histological demonstration of apoptosis in the reproductive organs of flukes of triclabendazole-sensitive and triclabendazole-resistant isolates, and in field-derived flukes from triclabendazole-treated hosts, using in situ hybridization to visualise endonuclease-generated DNA strand breaks. Vet. Parasitol. 2013, 191, 240–251. [Google Scholar] [PubMed]
- Bjorkman, N.; Thorsell, W. On the ultrastructure of the ovary of the liver fluke (Fasciola hepatica L.). Z. Zellforsch. Mikrosk. Anat. 1964, 63, 538–549. [Google Scholar] [CrossRef]
- Gresson, R.A.R. Electron microscopy of the ovary of Fasciola hepatica. Quart. J. Microsc. Sci. 1964, 105, 213–218. [Google Scholar]
- Irwin, S.W.B.; Threadgold, L.T. Electron microscope studies on Fasciola hepatica. VIII. The development of the vitelline cells. Exp. Parasitol. 1970, 28, 399–411. [Google Scholar] [CrossRef]
- Threadgold, L.T.; Irwin, S.W.B. Electron microscope studies of Fasciola hepatica IX. The fine structure of Mehlis’ gland. Z. Parasitkde 1970, 35, 16–30. [Google Scholar]
- Stitt, A.W.; Fairweather, I.; Mackender, R.O. The effect of triclabendazole (“Fasinex”) on protein synthesis by the liver fluke, Fasciola hepatica. Int. J. Parasitol. 1995, 25, 421–429. [Google Scholar] [CrossRef]
- Kumar, V.; Abbas, A.K.; Fausto, N. Robbins and Cotran Pathologic Basis of Disease, 7th ed.; Elsevier Saunders: Philadelphia, PA, USA, 2005; pp. 26–32. [Google Scholar]
- Lacey, E. The role of the cytoskeletal protein, tubulin, in the mode of action and mechanism of drug resistance to benzimidazoles. Int. J. Parasitol. 1988, 8, 399–404. [Google Scholar] [CrossRef]
- Smyth, J.D.; Halton, D.W. The Physiology of Trematodes, 2nd ed.; Cambridge University Press: Cambridge, UK, 1983; p. 446. [Google Scholar]
- Fairweather, I.; Threadgold, L.T.; Hanna, R.E.B. Development of Fasciola hepatica in the Mammalian Host. In Fasciolosis; Dalton, J.P., Ed.; CABI Publishing: Wallingford, UK, 1999; Chapter 3; pp. 47–111. [Google Scholar]
- Fairweather, I.; Maule, A.G.; Mitchell, S.H.; Johnston, C.F.; Halton, D.W. Immunocytochemical demonstration of 5-hydroxytryptamine (serotonin) in the nervous system of the liver fluke. Fasciola hepatica (Trematoda: Digenea). Parasitol. Res. 1987, 73, 255–258. [Google Scholar] [PubMed]
- Magee, R.M.; Fairweather, I.; Johnston, C.F.; Halton, D.W.; Shaw, C. Immunocytochemical demonstration of neuropeptides in the nervous system of the liver fluke. Fasciola hepatica (Trematoda: Digenea). Parasitology 1989, 98, 227–238. [Google Scholar] [PubMed]
- Threadgold, L.T. Fasciola hepatica: Stereological analysis of vitelline cell development. Exp. Parasitol. 1982, 54, 352–365. [Google Scholar] [CrossRef]
- Fairweather, I. Raising the bar on reporting “triclabendazole resistance”. Vet. Rec. 2011, 168, 514–515. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hanna, R. Fasciola hepatica: Histology of the Reproductive Organs and Differential Effects of Triclabendazole on Drug-Sensitive and Drug-Resistant Fluke Isolates and on Flukes from Selected Field Cases. Pathogens 2015, 4, 431-456. https://doi.org/10.3390/pathogens4030431
Hanna R. Fasciola hepatica: Histology of the Reproductive Organs and Differential Effects of Triclabendazole on Drug-Sensitive and Drug-Resistant Fluke Isolates and on Flukes from Selected Field Cases. Pathogens. 2015; 4(3):431-456. https://doi.org/10.3390/pathogens4030431
Chicago/Turabian StyleHanna, Robert. 2015. "Fasciola hepatica: Histology of the Reproductive Organs and Differential Effects of Triclabendazole on Drug-Sensitive and Drug-Resistant Fluke Isolates and on Flukes from Selected Field Cases" Pathogens 4, no. 3: 431-456. https://doi.org/10.3390/pathogens4030431