
The Host’s Reply to Candida Biofilm

University of Wisconsin-Madison, Departments of Medicine, Medical Microbiology and Immunology, 5203 Microbial Sciences Building, 1550 Linden Drive, Madison, WI 53706, USA
Pathogens 2016, 5(1), 33; https://doi.org/10.3390/pathogens5010033
Submission received: 15 January 2016
/
Revised: 9 March 2016
/
Accepted: 10 March 2016
/
Published: 18 March 2016
(This article belongs to the Special Issue Human Fungal Pathogens)
{kind=link}
{kind=link}
Abstract
:Candida spp. are among the most common nosocomial fungal pathogens and are notorious for their propensity toward biofilm formation. When growing on a medical device or mucosal surface, these organisms reside as communities embedded in a protective matrix, resisting host defenses. The host responds to Candida biofilm by depositing a variety of proteins that become incorporated into the biofilm matrix. Compared to free-floating Candida, leukocytes are less effective against Candida within a biofilm. This review highlights recent advances describing the host’s response to Candida biofilms using ex vivo and in vivo models of mucosal and device-associated biofilm infections.
1. Introduction
The vast majority of infections caused by Candida spp. involve proliferation of a biofilm on an artificial or biotic surface, such as the mucosa [1,2] (Figure 1). These adherent communities exhibit characteristics distinct from free-floating or planktonic cells, including the ability to tolerate high antifungal concentrations and to evade host immune detection [3,4,5,6]. Candidiasis is prominent in the hospital setting, with Candida spp. accounting for the 4th most common cause of bloodstream infection and the 3rd most common cause of urinary tract infection [7,8,9,10]. As medical care advances and device use increases, biofilm-associated infections have increased in parallel [11]. The most frequently used and infected devices include vascular catheters, urinary catheters, and dentures. However, pacemakers, artificial heart valves, voice prostheses, and central nervous system shunts are also at risk for infection [11,12]. The mortality due to invasive candidiasis, such as central venous catheter-associated infection, is astonishingly high, estimated at 26%–38% [11]. In addition, increased mortality has been observed when infected vascular catheters are retained, presumably due to the resilient nature of the biofilm communities [13].
Similar to device-associated infections, Candida spp. exhibit biofilm characteristics while adherent to biotic surfaces, such as the mucosa or the endothelium [14,15,16]. Mucosal biofilms are prevalent in the community. One of the most common mucosal biofilm infections, vaginal candidiasis, affects 30%–50% of women, with a subset of close to 6%–9% developing recurrent disease [17]. Oral candidiasis is similarly prevalent, particularly among patients who are elderly, immunosuppressed, or receiving antibiotics [18].
Like biofilms on abiotic surfaces, mucosal biofilms are also composed of collections of yeast and hyphal cells encased in an extracellular matrix [14,15,16,19]. However, their substrate for biofilm formation is a living structure, the mucosa, which responds to fungal adherence, secreted microbial products, and tissue invasion. Although many aspects of device-associated and mucosal biofilms are similar, the mucosal biofilms lack the adjacent abiotic surface and instead, are under the influence of immune factors induced by the Candida-epithelium interface [20,21]. Clinical studies demonstrating a difference in host susceptibility to Candida biofilm-associated infections suggest distinct immune responses to mucosal and device-associated biofilms [22]. For example, oral candidiasis is common in patients with dampened cell-mediated immunity, such as those with HIV and AIDS [18]. In contrast, denture stomatitis, a device-associated infection involving the same niche, is common in immunocompetent patients [23]. Clinical biofilms may be both mucosal and device-associated, such as dental stomatitis with oral candidiasis or vaginitis with intrauterine device infection [24,25].
Studies suggest that the biofilm lifestyle protects fungi from host recognition [26,27,28,29]. This is consistent with clinical studies showing that device-associated Candida biofilms are extraordinary difficult to cure, even for patients without immunocompromise [13,30,31]. This review will focus on how the host responds to Candida biofilm formation for various niches important for clinical infection (Figure 2). Although there are common themes to how the host responds to biofilms at these infection sites, variation exists. This is likely related to the differences in nutrients, host proteins, immunity, and physiological flow conditions. Most investigations have been undertaken with C. albicans. However, the majority of other pathogenic Candida spp., including C. dubliniensis, C. glabrata, C. krusei, C. tropicalis and C. parapsilosis, have also been shown to form biofilms of clinical significance [32].
2. Ex Vivo Models
2.1. Mononuclear cells
Investigations examining the leukocyte response to Candida biofilms have consistently shown a diminished response to these adherent communities [26,27,28,29]. Chandra et al. first used a co-culture system to analyze the interaction of peripheral blood mononuclear cells with C. albicans biofilms and identified several differences in leukocyte response [33]. When compared to co-culture with planktonic cells, mononuclear cells exposed to biofilms produced an altered cytokine profile with higher levels of IL-1β, IL-10, and MCP-1 and lower levels of IL-6 and MIP1β. As this pattern involves alteration of both pro- and anti-inflammatory pathways, it was hypothesized that mononuclear cell recognition of biofilms involves multiple interactions. Differences in phagocytic response were also observed. While the mononuclear cells elicited a phagocytic response to the planktonic cells, they migrated throughout biofilm structures without induction of phagocytosis or killing of the biofilm. Furthermore, the mononuclear cells augmented biofilm proliferation, increasing the biofilm thickness over two-fold [33]. The responsible factor has not been identified but was shown to be a soluble factor secreted into the supernatant during biofilm and mononuclear cell co-culture.
Additional investigation of the monocyte response to C. albicans biofilms by Katragkou et al. confirmed distinct differences in reaction to biofilm and planktonic cells [28]. For all conditions tested, monocytes had significantly less anti-biofilm activity. Monocyte activity (measured by damage to Candida in a tetrazolium salt XTT assay) against C. albicans biofilms was approximately half that observed for planktonic cultures. Consistent with prior investigation, the monocytes did not surround or engulf the biofilms and appeared inactive within the biofilm. Using a monocyte cell line, the authors showed a decreased pro-inflammatory cytokine response to biofilm, with reduced TNF-α release. Taken together, these ex vivo studies demonstrate altered recognition of Candida in the biofilm state. The mechanism of this is unclear but may involve masking of glucan as echinocandin treatment augmented the anti-biofilm activity of monocytes [28,34]. Surprisingly, studies examining that response to C. parapsilosis did not reveal significant differences in the activity of a monocyte cell line against biofilm and planktonic cells [26]. Possible etiologies underlying this phenomenon include the differences in biofilm architecture, filamentation, or extracellular matrix composition between C. albicans and C. parapsilosis biofilms.
2.2. Polymorphonuclear (PMN) Cells
PMNs cells exhibit diminished activity against C. albicans biofilms when compared to their impact on planktonic cultures [28]. In co-culture experiments, PMNs were approximately 50% less effective against biofilms, as measured by XTT. Interestingly, this difference was tightly linked to the biofilm architecture. When biofilms were physically disrupted by scraping, PMN activity increased to levels observed for planktonic cells. Further investigation questioned if cytokine-priming of PMNs or opsonization would augment the anti-biofilm activity [27]. However, pre-treatment of PMNs with interferon-γ (INF-γ) or granulocyte colony-stimulating factor (G-CSF) did not significantly enhance their activity against opsonized or unopsonized C. albicans biofilms. One possibility is that PMNs do not recognize C. albicans within a biofilm; hence, enhancement of ligand interactions by cytokine priming has minimal impact of PMN activity. This phenotype of neutrophil evasion is less pronounced for C. parapsilosis biofilms [26]. PMNs display similar activities against biofilm and planktonic C. parapsilosis.
Xie et al. further explored the neutrophil response to C. albicans biofilms to uncover the mechanism underlying the resistance to killing [29]. Several key observations were found. Compared to early biofilms (3 h), mature Candida biofilms (24 h) did not trigger production of reactive oxygen species (ROS) in neutrophils. The biofilm-exposed neutrophils remained viable for several hours and were able to be activated by alternative stimuli to induce fungal damage. Similar to a prior investigation, disruption of biofilm architecture promoted neutrophil activity and this was linked to increased ROS production [28]. The authors further correlated the hindered neutrophil response to the extracellular biofilm matrix, specifically, the presence of β-glucans.
3. Mucosal Biofilms Models
3.1. Oral Biofilms
Using a murine model of oral candidiasis, Dongari-Bagtzoglou et al. characterized C. albicans biofilm growth and examined the host response [16]. Candida biofilm induced a hyperkeratotic response and epithelial cell desquamation. Immuno-fluorescent imaging demonstrated incorporation of keratin and desquamated cells into the extracellular material surrounding the biofilm. In addition, the oral biofilm elicited neutrophil migration. Aggregates of neutrophils aligned adjacent to the biofilm with a subset migrating deeper in the biofilm. Although the neutrophils were present in the biofilm, they were not effective in clearing the infection. In part, this resistance of C. albicans oral biofilm to neutrophil killing appears to be due to the glycosylphosphatidylinositol (GPI)-anchored cell wall protein Hyr1 [35]. Expression of this hyphal-specific protein was found to promote resistance to neutrophil killing in vivo and in an ex vivo oral candidiasis model. Numerous investigations have examined the immune response to oral candidiasis [36,37]. Although these studies have not specifically described oral biofilms, the immune pathways involved in oral candidiasis likely apply to mucosal biofilms as well. Excellent reviews describe this response, which includes epithelial cell activation, priming of Th17 cells, and induction of cytokines, including IL-17 and IL-23 [36,37,38].
3.2. Vaginal Biofilms
Recent investigations have identified mucosal biofilm formation in a murine model of vaginal candidiasis [14,39,40]. Few studies have described the host response in the context of vaginal biofilms. However, considering the similarity of models used, much of current understanding of the immune response to vaginal candidiasis likely applies to vaginal Candida biofilms and the studies have been reviewed in more detail elsewhere [36,41,42,43]. Clinical studies demonstrate an inflammatory response with neutrophil infiltration in patients with symptomatic vaginal candidiasis [39]. However, the neutrophils appear to promote inflammation and tissue damage more than assist with fungal eradication. Using a murine vaginal candidiasis biofilm model, Yano et al. showed a similar pattern of neutrophil infiltration into vaginal lavage fluid [44]. In part, this response involves epithelial cell release of chemotactic factors, including alarmins S100-A8 and S100-A9 [44,45]. Although it is interesting to propose a role for IL-17 in vaginal candidiasis, a clear link has not been established [36,41]. Animal models of vaginal candidiasis demonstrate the importance of adaptive immunity [42,46]. Studies show the induction of protective immunity through vaccination and clinical trials are currently investigating the potential for use of vaccines.
4. Device-Associated Biofilms
4.1. Vascular Catheter Biofilms
Several animal models have been developed to mimic vascular catheter-associated Candida biofilm infection [47,48,49]. In these rabbit, rat, and mouse models, biofilms form on the surface of jugular venous catheters following luminal inoculation. Similar to mucosal biofilms, imaging of these device-associated biofilms revealed the incorporation of host cells within the biofilm [47]. While the majority of the cells were of fungal origin, few were larger and had the appearance of leukocytes. Further examination of these cells confirmed the presence of biofilm-associated neutrophils [50]. However, consistent with electron microscopy imaging, the neutrophils were relatively scarce, approximately one per 75 C. albicans cells. This is contrast to studies of mucosal biofilms, where neutrophils were observed to migrate throughout the biofilm [16].
A striking finding from both the rabbit and rat venous catheter biofilms was the presence of a robust extracellular matrix, more extensive than that observed for many in vitro conditions [47,48,50]. This observation suggested the incorporation of host proteins in the extracellular matrix and prompted proteomic analysis of the material [50]. Surprisingly, nearly all (98%) of the extracellular matrix proteins were of host origin. Over 100 host proteins of a variety of types were identified. The most abundant included hemoglobin, albumin, and alpha globulins. Several represented categories included matricellular proteins, inflammatory or leukocyte-associated proteins, and erythrocyte or heme-associated proteins. Although the role of many of these proteins is unclear, they may be involved in the immune response, fungal acquisition of iron, or scaffolding of the matrix [45,51,52,53,54,55,56]. Others may involve non-specific interactions.
4.2. Denture Biofilms
Several animal models have been utilized to study the host response to denture stomatitis biofilms [22,57,58,59]. For the most part, these studies have examined the inflammatory response of the adjacent mucosal tissue. Johnson et al. developed a rat model of chronic denture dermatitis, which utilizes custom fitted intraoral devices that can be removed and sampled over time [22,59]. Over the course of eight weeks, animals progressively developed palatal inflammation, erythema, and edema in response to the device-associated C. albicans biofilm [22]. Histopathology revealed prominent inflammatory infiltrates by 6–8 weeks. The palatal inflammation and lesions mimicked clinical denture stomatitis. The findings are also in line with prior investigations of chronic denture stomatitis in animal models that did not specifically examine biofilm formation [60,61].
A rat model of more acute denture stomatitis has also been utilized to examine the host response to C. albicans biofilm infection [50,58]. In this model, palatal devices are constructed in situ and device-associated biofilms form over two days. In contrast to the chronic stomatitis models, animals are immunosuppressed with corticosteroids. Similar to models of chronic infection, mucosal inflammation with leukocyte infiltration was observed [58]. However, hyphal invasion was noted as well, so it is unclear if the leukocyte infiltration was prompted by denture-associated biofilm, oral candidiasis, or the combination. Further examination of the host cells associating directly with the device revealed a combination of leukocytes and epithelial cells [50]. Host proteins were prominent in the extracellular matrix, with 132 identified. The most prevalent included amylase, hemoglobin, and antimicrobial peptides (bactericidal permeability-increasing or BPI-fold containing proteins). These findings show a variety of host cells and proteins are in direct contact with denture biofilms. Further investigations are needed to determine the role of these host components.
4.3. Urinary Catheter Biofilms
Wang et al. developed a murine model of urinary catheter-associated candidiasis and characterized the host response to biofilm [62]. In this model, catheter segments are surgically placed in the bladder and inoculated by intravesicular injection. C. albicans biofilm was found to illicit an inflammatory response marked by pyuria and submucosal bladder inflammation. Furthermore, their studies support a role for lysozyme, an innate immunity effector present on the mucosa and expressed by neutrophils, in the clearance of infection. Compared to the parent strains, lysozyme M-deficient mice (lysM-/-) developed higher fungal burdens and more pronounced pyuria. The inflammatory response is likely a result of not only the biofilm but also the concurrent cystitis.
The host response to Candida urinary catheter biofilm was further explored using a rat model [63]. In contrast to the murine model, this involves luminal inoculation of a urethral catheter. Over the course of 48 h, biofilms formed on both the luminal surface and the bladder epithelial surface. Histopathology demonstrated findings of acute cystitis, including fungal invasion and neutrophil infiltration. Using this infection model, a proteomic analysis of the extracellular material associating with the C. albicans urinary catheter biofilm was undertaken [50]. Analysis revealed numerous (>200) host proteins within the Candida biofilm. The most abundant proteins were fibrinogen, keratin, and hemoglobin. Similar to other models of infection, leukocyte-associated and inflammatory proteins were identified as well. Imaging of the biofilm individual cells confirmed the presence of few neutrophils associating with the biofilm.
4.4. Subcutaneous Biofilms
Several subcutaneous implant animal models have shed light on the host response to Candida biofilm [64,65,66]. Riciova et al. developed a rat model of Candida biofilm infection which involves implantation of vascular catheter segments in the subcutaneous tissue. Mature biofilms formed over the course of six days [64]. The animals rapidly cleared the C. albicans infection unless they were immunosuppressed with glucocorticoid. The robust host inflammatory response may have been related to a foreign body reaction or the adherent C. albicans, which had not yet formed a mature biofilm. Similarly, Nieminen et al. observed an inflammatory response to C. albicans biofilms using a murine model involving subcutaneous implantation of a chamber [66]. In addition to the wound healing response that was observed for the chamber-only (uninfected) controls, an infiltrate of neutrophils was also seen in animals with Candida biofilm infections. The authors further described an anti-inflammatory compound, leucine derivative DL-2-hydroxyisocaproic acid (HICA), which modulated this activity. Treatment with HICA, which is active against C. albicans biofilms, decreased infiltration of neutrophils and formation of granulation tissue.
5. Other Models
Galleria mellonella. Although mammalian models of biofilm-associated infection closely mimic clinical disease, there is interest in alternative invertebrate models to provide higher throughput screening, limit cost, and utilize less sentient animals. G. mellonella (Lepidoptera: the greater wax moth) offers an alternative model for biofilm study [67]. Although insect models lack factors for acquired immunity, they do have a well-developed innate immune system comprised of phagocytes (hemocytes of the hemolymph) and humoral components. In addition, the G. mellonella larvae model allows growth of Candida at 37 °C and collection of tissues for histology. Using this model, Borghi, et al. identified a correlation between in vitro C. albicans biofilm formation and pathogenesis [67]. On histology, invasive fungal mats resembling biofilms were visualized. Regardless of the burden, the host response included melanization and fat body cell sequestration, processes that contain infecting pathogens to the hemolymph. At the higher burden associated with more robust biofilm production, necrosis of the fat body was observed. Further studies have used optimized larval processing and histology to quantitate these associating hemocytes as well as measure the expression and activity of the host antifungal peptides and enzymes [68,69]. The unique model is a tool to examine the innate immune response to biofilm infection, although findings may be limited by differences between mammalian and invertebrate immunity.
6. Conclusions
The majority of Candida infections involve the production of surface-associated biofilm communities. Recent studies show that these structures evade host responses, including killing by both mononuclear and polymorphonuclear leukocytes. This appears to be a multifactorial process and likely varies by clinical niche of infection. Studies suggest a role for the extracellular matrix in this immune evasion phenomenon. However, much remains unknown regarding how the immune system recognizes the extracellular matrix and how many biofilms go virtually unrecognized. Novel strategies to treat fungal biofilm infections are of great interest. Further research in this area may identify biofilm-specific drug targets, including agents designed to disrupt extracellular matrix, augment current antifungal therapies, or disarm biofilm immune evasion.
Acknowledgments
Jeniel E. Nett is supported by the National Institutes of Health (K08 AI108727) and the Burroughs Wellcome Fund (1012299).
Conflicts of Interest
The author declares no conflicts of interest.
References
- Douglas, L.J. Candida biofilms and their role in infection. Trends Microbiol. 2003, 11, 30–36. [Google Scholar] [CrossRef]
- Kumamoto, C.A.; Vinces, M.D. Alternative Candida albicans lifestyles: Growth on surfaces. Annu. Rev. Microbiol. 2005, 59, 113–133. [Google Scholar] [CrossRef] [PubMed]
- Al-Fattani, M.A.; Douglas, L.J. Biofilm matrix of Candida albicans and Candida tropicalis: Chemical composition and role in drug resistance. J. Med. Microbiol. 2006, 55, 999–1008. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, P.K.; Chandra, J.; Kuhn, D.M.; Ghannoum, M.A. Mechanism of fluconazole resistance in Candida albicans biofilms: Phase-specific role of efflux pumps and membrane sterols. Infect. Immun. 2003, 71, 4333–4340. [Google Scholar] [CrossRef] [PubMed]
- Nett, J.; Lincoln, L.; Marchillo, K.; Massey, R.; Holoyda, K.; Hoff, B.; VanHandel, M.; Andes, D. Putative role of beta-1,3 glucans in Candida albicans biofilm resistance. Antimicrob. Agents Chemother. 2007, 51, 510–520. [Google Scholar] [CrossRef] [PubMed]
- Ramage, G.; Vandewalle, K.; Wickes, B.L.; Lopez-Ribot, J.L. Characteristics of biofilm formation by Candida albicans. Rev. Iberoam. Micol. 2001, 18, 163–170. [Google Scholar] [PubMed]
- Groeger, J.S.; Lucas, A.B.; Thaler, H.T.; Friedlander-Klar, H.; Brown, A.E.; Kiehn, T.E.; Armstrong, D. Infectious morbidity associated with long-term use of venous access devices in patients with cancer. Ann. Intern. Med. 1993, 119, 1168–1174. [Google Scholar] [CrossRef] [PubMed]
- Richards, M.J.; Edwards, J.R.; Culver, D.H.; Gaynes, R.P. Nosocomial infections in medical intensive care units in the United States. National Nosocomial Infections Surveillance System. Crit. Care Med. 1999, 27, 887–892. [Google Scholar] [CrossRef] [PubMed]
- Edmond, M.B.; Wallace, S.E.; McClish, D.K.; Pfaller, M.A.; Jones, R.N.; Wenzel, R.P. Nosocomial bloodstream infections in United States hospitals: A three-year analysis. Clin. Infect. Dis. 1999, 29, 239–244. [Google Scholar] [CrossRef] [PubMed]
- Pfaller, M.A.; Diekema, D.J. Epidemiology of invasive candidiasis: A persistent public health problem. Clin. Microbiol. Rev. 2007, 20, 133–163. [Google Scholar] [CrossRef] [PubMed]
- Kojic, E.M.; Darouiche, R.O. Candida infections of medical devices. Clin. Microbiol. Rev. 2004, 17, 255–267. [Google Scholar] [CrossRef] [PubMed]
- Donlan, R.M. Biofilms and device-associated infections. Emerg. Infect. Dis. 2001, 7, 277–281. [Google Scholar] [CrossRef] [PubMed]
- Andes, D.R.; Safdar, N.; Baddley, J.W.; Playford, G.; Reboli, A.C.; Rex, J.H.; Sobel, J.D.; Pappas, P.G.; Kullberg, B.J. Impact of treatment strategy on outcomes in patients with candidemia and other forms of invasive candidiasis: A patient-level quantitative review of randomized trials. Clin. Infect. Dis. 2012, 54, 1110–1122. [Google Scholar] [CrossRef] [PubMed]
- Harriott, M.M.; Lilly, E.A.; Rodriguez, T.E.; Fidel, P.L., Jr.; Noverr, M.C. Candida albicans forms biofilms on the vaginal mucosa. Microbiology 2010, 156, 3635–3644. [Google Scholar] [CrossRef] [PubMed]
- Ganguly, S.; Mitchell, A.P. Mucosal biofilms of Candida albicans. Curr. Opin. Microbiol. 2011, 14, 380–385. [Google Scholar] [CrossRef] [PubMed]
- Dongari-Bagtzoglou, A.; Kashleva, H.; Dwivedi, P.; Diaz, P.; Vasilakos, J. Characterization of mucosal Candida albicans biofilms. PLoS ONE 2009, 4, e7967. [Google Scholar] [CrossRef] [PubMed]
- Foxman, B.; Muraglia, R.; Dietz, J.P.; Sobel, J.D.; Wagner, J. Prevalence of recurrent vulvovaginal candidiasis in 5 European countries and the United States: Results from an internet panel survey. J. Low Genit. Tract Dis. 2013, 17, 340–345. [Google Scholar] [CrossRef] [PubMed]
- Vazquez, J.A.; Sobel, J.D. Mucosal candidiasis. Infect. Dis. Clin. N. Am. 2002, 16, 793–820. [Google Scholar] [CrossRef]
- Hawser, S.P.; Baillie, G.S.; Douglas, L.J. Production of extracellular matrix by Candida albicans biofilms. J. Med. Microbiol. 1998, 47, 253–256. [Google Scholar] [CrossRef] [PubMed]
- Naglik, J.R.; Richardson, J.P.; Moyes, D.L. Candida albicans pathogenicity and epithelial immunity. PLoS Pathog. 2014, 10, e1004257. [Google Scholar] [CrossRef] [PubMed]
- Williams, D.W.; Jordan, R.P.; Wei, X.Q.; Alves, C.T.; Wise, M.P.; Wilson, M.J.; Lewis, M.A. Interactions of Candida albicans with host epithelial surfaces. J. Oral Microbiol. 2013, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, C.C.; Yu, A.; Lee, H.; Fidel, P.L., Jr.; Noverr, M.C. Development of a contemporary animal model of Candida albicans-associated denture stomatitis using a novel intraoral denture system. Infect. Immun. 2012, 80, 1736–1743. [Google Scholar] [CrossRef] [PubMed]
- Webb, B.C.; Thomas, C.J.; Willcox, M.D.; Harty, D.W.; Knox, K.W. Candida-associated denture stomatitis. Aetiology and management: A review. Part 2. Oral diseases caused by Candida species. Aust. Dent. J. 1998, 43, 160–166. [Google Scholar] [CrossRef] [PubMed]
- Auler, M.E.; Morreira, D.; Rodrigues, F.F.; Abr Ao, M.S.; Margarido, P.F.; Matsumoto, F.E.; Silva, E.G.; Silva, B.C.; Schneider, R.P.; Paula, C.R. Biofilm formation on intrauterine devices in patients with recurrent vulvovaginal candidiasis. Med. Mycol. 2010, 48, 211–216. [Google Scholar] [CrossRef] [PubMed]
- Ramage, G.; Tomsett, K.; Wickes, B.L.; Lopez-Ribot, J.L.; Redding, S.W. Denture stomatitis: A role for Candida biofilms. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. Endod. 2004, 98, 53–59. [Google Scholar] [CrossRef] [PubMed]
- Katragkou, A.; Chatzimoschou, A.; Simitsopoulou, M.; Georgiadou, E.; Roilides, E. Additive antifungal activity of anidulafungin and human neutrophils against Candida parapsilosis biofilms. J. Antimicrob. Chemother. 2011, 66, 588–591. [Google Scholar] [CrossRef] [PubMed]
- Katragkou, A.; Simitsopoulou, M.; Chatzimoschou, A.; Georgiadou, E.; Walsh, T.J.; Roilides, E. Effects of interferon-gamma and granulocyte colony-stimulating factor on antifungal activity of human polymorphonuclear neutrophils against Candida albicans grown as biofilms or planktonic cells. Cytokine 2011, 55, 330–334. [Google Scholar] [CrossRef] [PubMed]
- Katragkou, A.; Kruhlak, M.J.; Simitsopoulou, M.; Chatzimoschou, A.; Taparkou, A.; Cotten, C.J.; Paliogianni, F.; Diza-Mataftsi, E.; Tsantali, C.; Walsh, T.J.; et al. Interactions between human phagocytes and Candida albicans biofilms alone and in combination with antifungal agents. J. Infect. Dis. 2010, 201, 1941–1949. [Google Scholar] [CrossRef] [PubMed]
- Xie, Z.; Thompson, A.; Sobue, T.; Kashleva, H.; Xu, H.; Vasilakos, J.; Dongari-Bagtzoglou, A. Candida albicans biofilms do not trigger reactive oxygen species and evade neutrophil killing. J. Infect. Dis. 2012, 206, 1936–1945. [Google Scholar] [CrossRef] [PubMed]
- Pappas, P.G.; Kauffman, C.A.; Andes, D.; Benjamin, D.K., Jr.; Calandra, T.F.; Edwards, J.E., Jr.; Filler, S.G.; Fisher, J.F.; Kullberg, B.J.; Ostrosky-Zeichner, L.; et al. Clinical practice guidelines for the management of candidiasis: 2009 update by the Infectious Diseases Society of America. Clin. Infect. Dis. 2009, 48, 503–535. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, M.H.; Nguyen, M.L.; Yu, V.L.; McMahon, D.; Keys, T.F.; Amidi, M. Candida prosthetic valve endocarditis: Prospective study of six cases and review of the literature. Clin. Infect. Dis. 1996, 22, 262–267. [Google Scholar] [CrossRef] [PubMed]
- Shin, J.H.; Kee, S.J.; Shin, M.G.; Kim, S.H.; Shin, D.H.; Lee, S.K.; Suh, S.P.; Ryang, D.W. Biofilm production by isolates of Candida species recovered from nonneutropenic patients: Comparison of bloodstream isolates with isolates from other sources. J. Clin. Microbiol. 2002, 40, 1244–1248. [Google Scholar] [CrossRef] [PubMed]
- Chandra, J.; McCormick, T.S.; Imamura, Y.; Mukherjee, P.K.; Ghannoum, M.A. Interaction of Candida albicans with adherent human peripheral blood mononuclear cells increases C. albicans biofilm formation and results in differential expression of pro- and anti-inflammatory cytokines. Infect. Immun. 2007, 75, 2612–2620. [Google Scholar] [CrossRef] [PubMed]
- Wheeler, R.T.; Fink, G.R. A drug-sensitive genetic network masks fungi from the immune system. PLoS Pathog. 2006, 2, e35. [Google Scholar] [CrossRef] [PubMed]
- Dwivedi, P.; Thompson, A.; Xie, Z.; Kashleva, H.; Ganguly, S.; Mitchell, A.P.; Dongari-Bagtzoglou, A. Role of Bcr1-activated genes Hwp1 and Hyr1 in Candida albicans oral mucosal biofilms and neutrophil evasion. PLoS ONE 2011, 6, e16218. [Google Scholar] [CrossRef] [PubMed]
- Conti, H.R.; Gaffen, S.L. IL-17-mediated immunity to the opportunistic fungal pathogen Candida albicans. J. Immunol. 2015, 195, 780–788. [Google Scholar] [CrossRef] [PubMed]
- Hebecker, B.; Naglik, J.R.; Hube, B.; Jacobsen, I.D. Pathogenicity mechanisms and host response during oral Candida albicans infections. Expert Rev. Anti-infect Ther. 2014, 12, 867–879. [Google Scholar] [CrossRef] [PubMed]
- Filler, S.G. Insights from human studies into the host defense against candidiasis. Cytokine 2012, 58, 129–132. [Google Scholar] [CrossRef] [PubMed]
- Yano, J.; Palmer, G.E.; Eberle, K.E.; Peters, B.M.; Vogl, T.; McKenzie, A.N.; Fidel, P.L., Jr. Vaginal epithelial cell-derived S100 alarmins induced by Candida albicans via pattern recognition receptor interactions are sufficient but not necessary for the acute neutrophil response during experimental vaginal candidiasis. Infect. Immun. 2014, 82, 783–792. [Google Scholar] [CrossRef] [PubMed]
- Yano, J.; Kolls, J.K.; Happel, K.I.; Wormley, F.; Wozniak, K.L.; Fidel, P.L., Jr. The acute neutrophil response mediated by S100 alarmins during vaginal Candida infections is independent of the Th17-pathway. PLoS ONE 2012, 7, e46311. [Google Scholar] [CrossRef] [PubMed]
- Peters, B.M.; Yano, J.; Noverr, M.C.; Fidel, P.L., Jr. Candida vaginitis: When opportunism knocks, the host responds. PLoS Pathog. 2014, 10, e1003965. [Google Scholar] [CrossRef] [PubMed]
- Cassone, A. Vulvovaginal Candida albicans infections: Pathogenesis, immunity and vaccine prospects. BJOG 2015, 122, 785–794. [Google Scholar] [CrossRef] [PubMed]
- Fidel, P.L., Jr. Immunity to Candida. Oral Dis. 2002, 8 (Suppl. S2), 69–75. [Google Scholar] [CrossRef] [PubMed]
- Yano, J.; Lilly, E.; Barousse, M.; Fidel, P.L., Jr. Epithelial cell-derived S100 calcium-binding proteins as key mediators in the hallmark acute neutrophil response during Candida vaginitis. Infect. Immun. 2010, 78, 5126–5137. [Google Scholar] [CrossRef] [PubMed]
- Yano, J.; Noverr, M.C.; Fidel, P.L., Jr. Cytokines in the host response to Candida vaginitis: Identifying a role for non-classical immune mediators, S100 alarmins. Cytokine 2012, 58, 118–128. [Google Scholar] [CrossRef] [PubMed]
- Sobel, J.D. Recurrent vulvovaginal candidiasis. Am. J. Obstet. Gynecol. 2016, 214, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Andes, D.; Nett, J.; Oschel, P.; Albrecht, R.; Marchillo, K.; Pitula, A. Development and characterization of an in vivo central venous catheter Candida albicans biofilm model. Infect. Immun. 2004, 72, 6023–6031. [Google Scholar] [CrossRef] [PubMed]
- Schinabeck, M.K.; Long, L.A.; Hossain, M.A.; Chandra, J.; Mukherjee, P.K.; Mohamed, S.; Ghannoum, M.A. Rabbit model of Candida albicans biofilm infection: Liposomal amphotericin B antifungal lock therapy. Antimicrob. Agents Chemother. 2004, 48, 1727–1732. [Google Scholar] [CrossRef] [PubMed]
- Lazzell, A.L.; Chaturvedi, A.K.; Pierce, C.G.; Prasad, D.; Uppuluri, P.; Lopez-Ribot, J.L. Treatment and prevention of Candida albicans biofilms with caspofungin in a novel central venous catheter murine model of candidiasis. J. Antimicrob. Chemother. 2009, 64, 567–570. [Google Scholar] [CrossRef] [PubMed]
- Nett, J.E.; Zarnowski, R.; Cabezas-Olcoz, J.; Brooks, E.G.; Bernhardt, J.; Marchillo, K.; Mosher, D.F.; Andes, D.R. Host contributions to construction of three device-associated Candida albicans biofilms. Infect. Immun. 2015, 83, 4630–4638. [Google Scholar] [PubMed]
- Kuznets, G.; Vigonsky, E.; Weissman, Z.; Lalli, D.; Gildor, T.; Kauffman, S.J.; Turano, P.; Becker, J.; Lewinson, O.; Kornitzer, D. A relay network of extracellular heme-binding proteins drives C. albicans iron acquisition from hemoglobin. PLoS Pathog. 2014, 10, e1004407. [Google Scholar] [CrossRef] [PubMed]
- Francois, P.; Schrenzel, J.; Stoerman-Chopard, C.; Favre, H.; Herrmann, M.; Foster, T.J.; Lew, D.P.; Vaudaux, P. Identification of plasma proteins adsorbed on hemodialysis tubing that promote Staphylococcus aureus adhesion. J. Lab. Clin. Med. 2000, 135, 32–42. [Google Scholar] [CrossRef]
- Proctor, R.A. Toward an understanding of biomaterial infections: A complex interplay between the host and bacteria. J. Lab. Clin. Med. 2000, 135, 14–15. [Google Scholar] [CrossRef]
- Jenney, C.R.; Anderson, J.M. Adsorbed serum proteins responsible for surface dependent human macrophage behavior. J. Biomed. Mater. Res. 2000, 49, 435–447. [Google Scholar] [CrossRef]
- Brash, J.L.; Ten Hove, P. Protein adsorption studies on “standard” polymeric materials. J. Biomater. Sci. Polym. Ed. 1993, 4, 591–599. [Google Scholar] [CrossRef] [PubMed]
- Yanagisawa, N.; Li, D.Q.; Ljungh, A. Protein adsorption on ex vivo catheters and polymers exposed to peritoneal dialysis effluent. Perit. Dial. Int. 2004, 24, 264–273. [Google Scholar] [PubMed]
- Samaranayake, Y.H.; Samaranayake, L.P. Experimental oral candidiasis in animal models. Clin. Microbiol. Rev. 2001, 14, 398–429. [Google Scholar] [CrossRef] [PubMed]
- Nett, J.E.; Marchillo, K.; Spiegel, C.A.; Andes, D.R. Development and validation of an in vivo Candida albicans biofilm denture model. Infect. Immun. 2010, 78, 3650–3659. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Yu, A.; Johnson, C.C.; Lilly, E.A.; Noverr, M.C.; Fidel, P.L., Jr. Fabrication of a multi-applicable removable intraoral denture system for rodent research. J. Oral Rehabil. 2011, 38, 686–690. [Google Scholar] [CrossRef] [PubMed]
- Samaranayake, Y.H.; Ye, J.; Yau, J.Y.; Cheung, B.P.; Samaranayake, L.P. In vitro method to study antifungal perfusion in Candida biofilms. J. Clin. Microbiol. 2005, 43, 818–825. [Google Scholar] [CrossRef] [PubMed]
- Olsen, I.; Bondevik, O. Experimental Candida-induced denture stomatitis in the Wistar rat. Scand. J. Dent. Res. 1978, 86, 392–398. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Fries, B.C. A murine model for catheter-associated candiduria. J. Med. Microbiol. 2011, 60, 1523–1529. [Google Scholar]
- Nett, J.E.; Brooks, E.G.; Cabezas-Olcoz, J.; Sanchez, H.; Zarnowski, R.; Marchillo, K.; Andes, D.R. Rat indwelling urinary catheter model of Candida albicans biofilm infection. Infect. Immun. 2014, 82, 4931–4940. [Google Scholar] [CrossRef] [PubMed]
- Ricicova, M.; Kucharikova, S.; Tournu, H.; Hendrix, J.; Bujdakova, H.; Van Eldere, J.; Lagrou, K.; Van Dijck, P. Candida albicans biofilm formation in a new in vivo rat model. Microbiology 2010, 156, 909–919. [Google Scholar] [CrossRef] [PubMed]
- Rajendran, R.; Mowat, E.; McCulloch, E.; Lappin, D.F.; Jones, B.; Lang, S.; Majithiya, J.B.; Warn, P.; Williams, C.; Ramage, G. Azole resistance of Aspergillus fumigatus biofilms is partly associated with efflux pump activity. Antimicrob. Agents Chemother. 2011, 55, 2092–2097. [Google Scholar] [CrossRef] [PubMed]
- Nieminen, M.T.; Hernandez, M.; Novak-Frazer, L.; Kuula, H.; Ramage, G.; Bowyer, P.; Warn, P.; Sorsa, T.; Rautemaa, R. DL-2-hydroxyisocaproic acid attenuates inflammatory responses in a murine Candida albicans biofilm model. Clin. Vaccine Immunol. 2014, 21, 1240–1245. [Google Scholar] [CrossRef] [PubMed]
- Borghi, E.; Romagnoli, S.; Fuchs, B.B.; Cirasola, D.; Perdoni, F.; Tosi, D.; Braidotti, P.; Bulfamante, G.; Morace, G.; Mylonakis, E. Correlation between Candida albicans biofilm formation and invasion of the invertebrate host Galleria mellonella. Future Microbiol. 2014, 9, 163–173. [Google Scholar] [CrossRef] [PubMed]
- Perdoni, F.; Falleni, M.; Tosi, D.; Cirasola, D.; Romagnoli, S.; Braidotti, P.; Clementi, E.; Bulfamante, G.; Borghi, E. A histological procedure to study fungal infection in the wax moth Galleria mellonella. Eur. J. Histochem. 2014, 58. [Google Scholar] [CrossRef] [PubMed]
- Rajendran, R.; Borghi, E.; Falleni, M.; Perdoni, F.; Tosi, D.; Lappin, D.F.; O’Donnell, L.; Greetham, D.; Ramage, G.; Nile, C. Acetylcholine protects against Candida albicans infection by inhibiting biofilm formation and promoting hemocyte function in a Galleria mellonella infection model. Eukaryot. Cell 2015, 14, 834–844. [Google Scholar] [CrossRef] [PubMed]
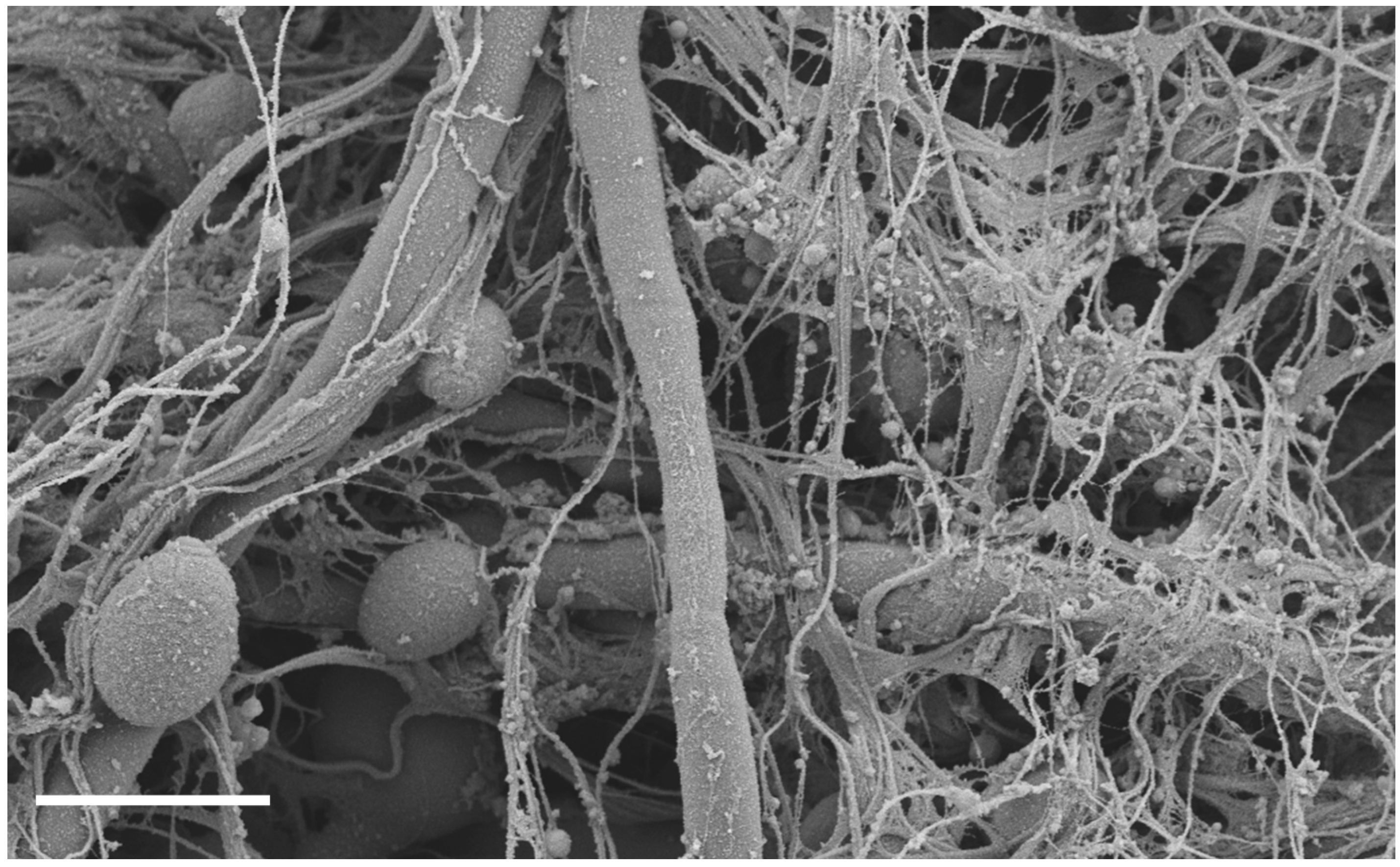
Figure 1.
C. albicans biofilm infection of rat venous catheter. Following C. albicans instillation and a two-day growth period, catheters were processed and imaged on a JEOL 1530. Measurement bar represents 5 μm. The biofilm is composed of both yeast and hyphae embedded in an extracellular matrix of host and fungal components.
Figure 1.
C. albicans biofilm infection of rat venous catheter. Following C. albicans instillation and a two-day growth period, catheters were processed and imaged on a JEOL 1530. Measurement bar represents 5 μm. The biofilm is composed of both yeast and hyphae embedded in an extracellular matrix of host and fungal components.

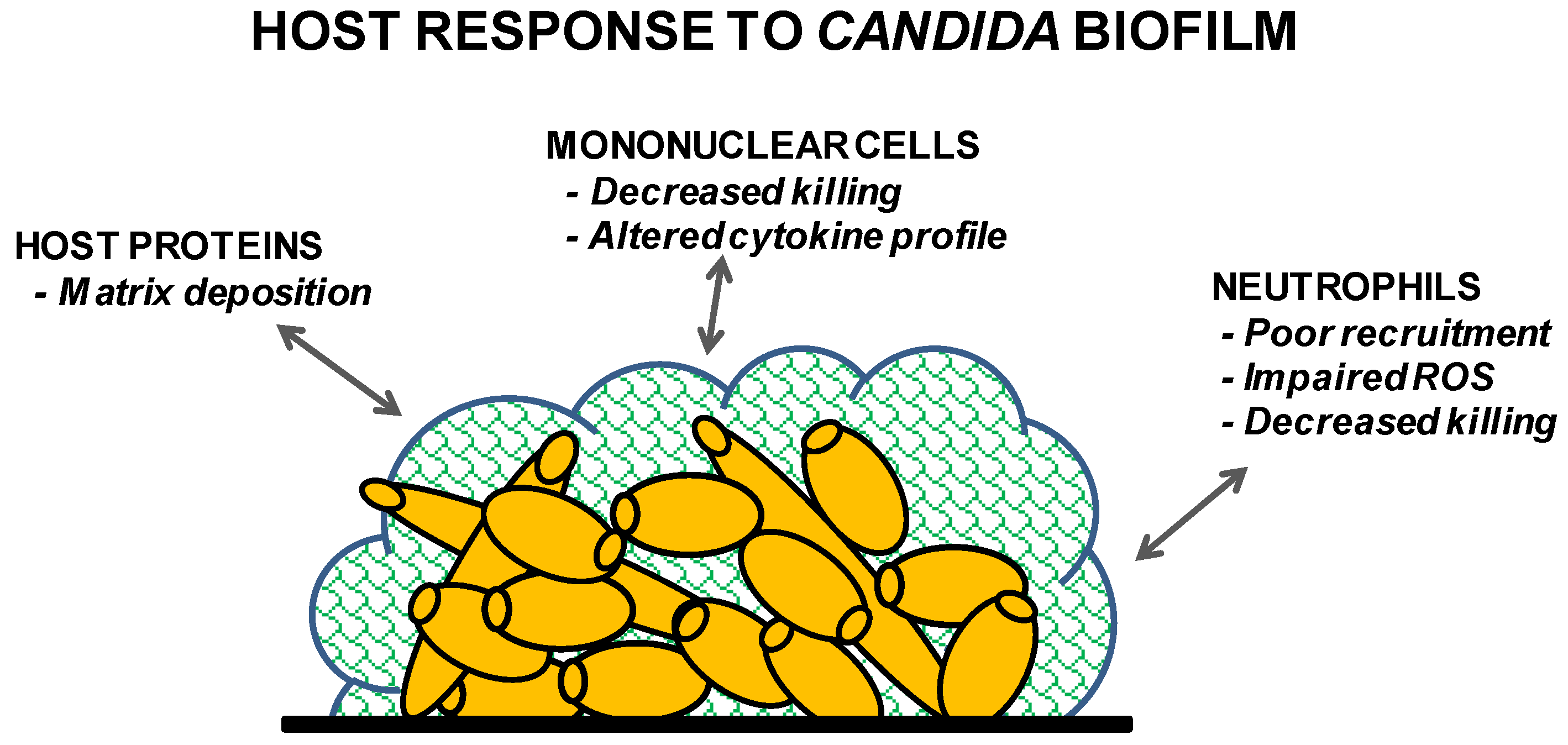
Figure 2.
Summary of the host’s response to Candida biofilm.

© 2016 by the author; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Nett, J.E. The Host’s Reply to Candida Biofilm. Pathogens 2016, 5, 33. https://doi.org/10.3390/pathogens5010033
AMA Style
Nett JE. The Host’s Reply to Candida Biofilm. Pathogens. 2016; 5(1):33. https://doi.org/10.3390/pathogens5010033
Chicago/Turabian StyleNett, Jeniel E. 2016. "The Host’s Reply to Candida Biofilm" Pathogens 5, no. 1: 33. https://doi.org/10.3390/pathogens5010033
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.