
Astrocytes Enhance Streptococcus suis-Glial Cell Interaction in Primary Astrocyte-Microglial Cell Co-Cultures

 , ,
, , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
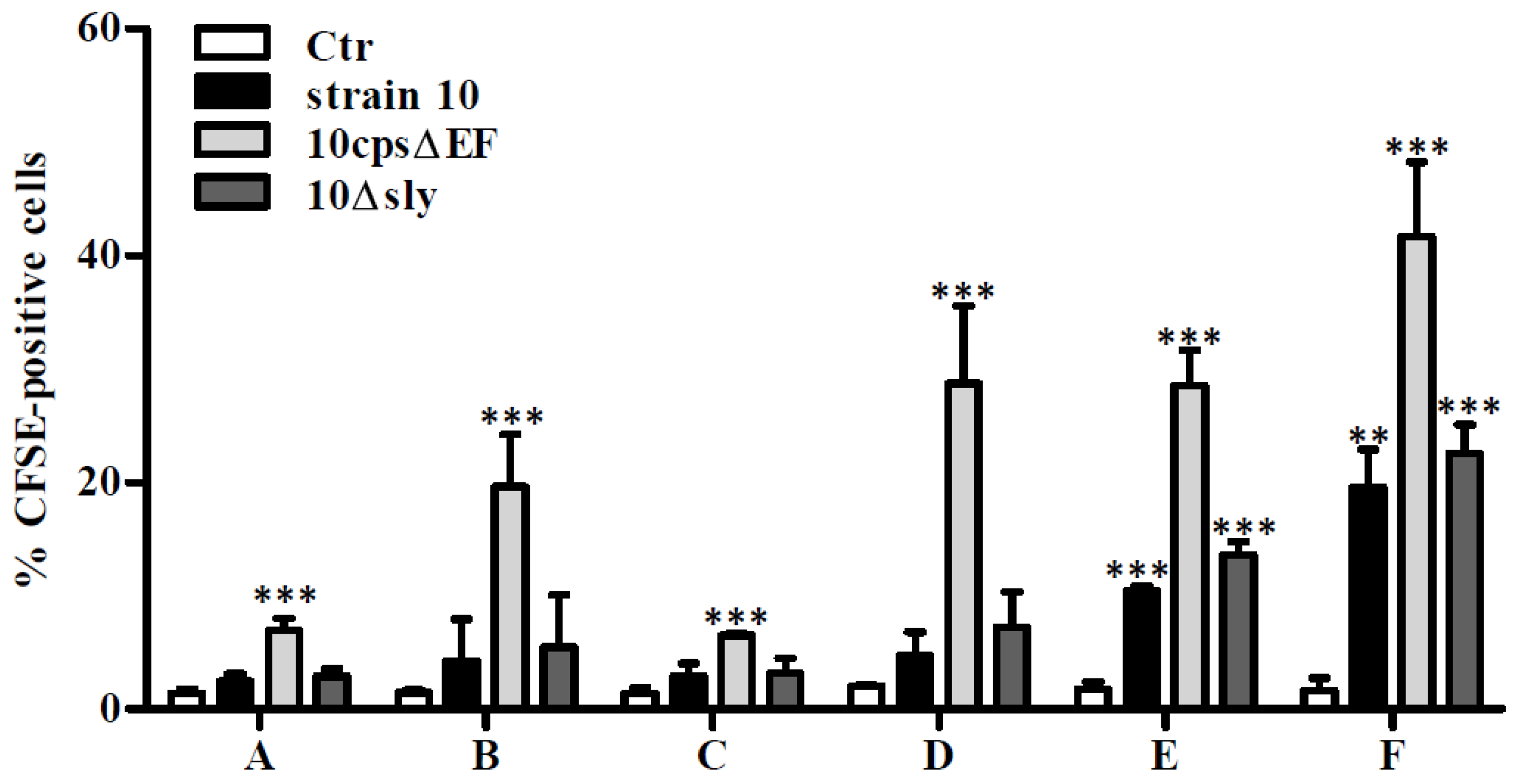
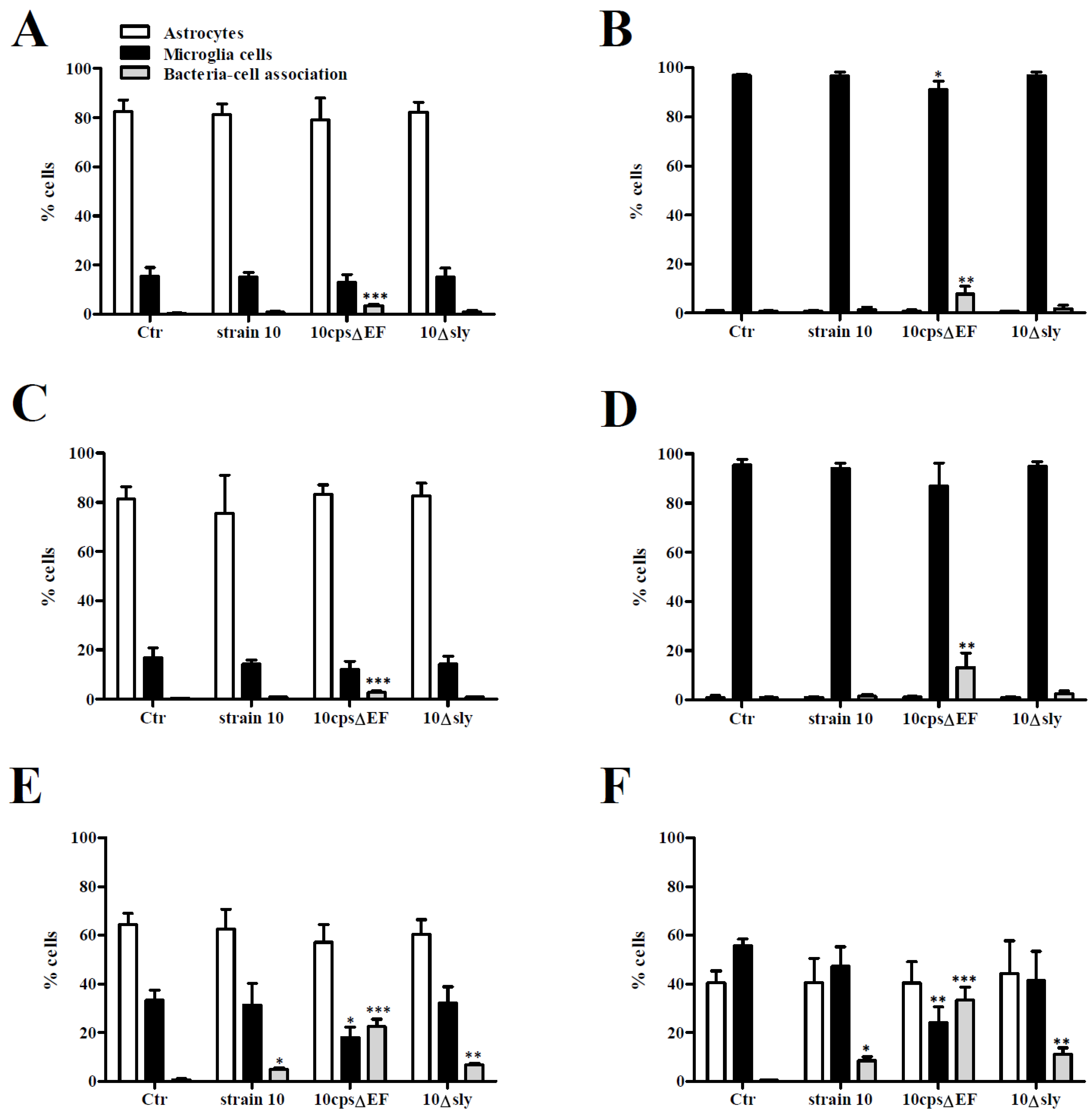
2.1. Association of S. suis with Primary Astrocytes and Microglial Cells
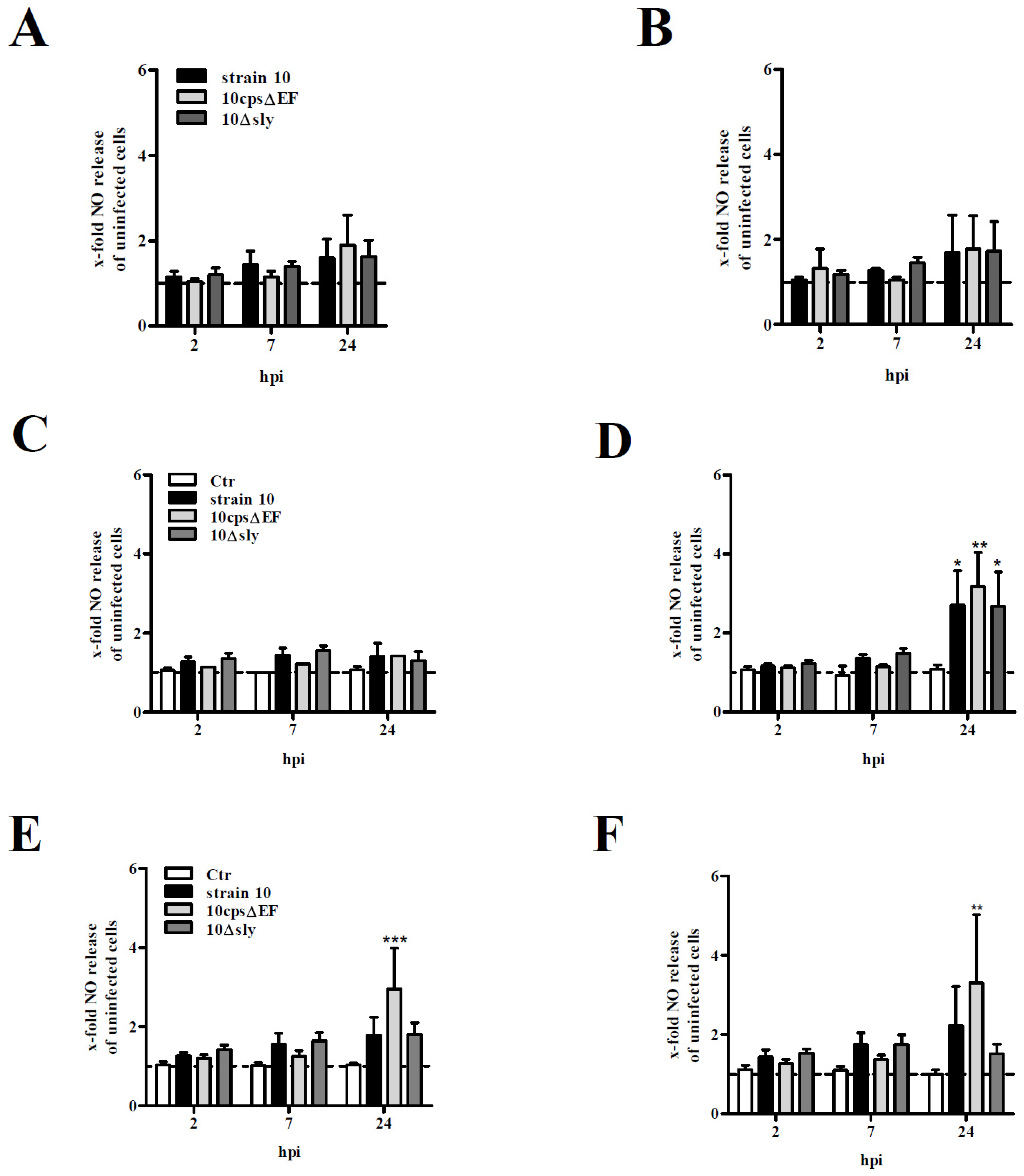
2.2. NO Release by Primary Mouse Glial Cells after Infection with S. suis
3. Experimental Section
3.1. Bacterial Strains and Growth Conditions
3.2. Isolation and Cultivation of Mixed Glial Cell Cultures
3.3. Cultivation and Separation of Astrocytes and Microglial Cells
3.4. Pre-incubation of Astrocytes and Microglial Cells with Cell Supernatants
3.5. Flow Cytometry Characterization of Astrocytes and Microglial Cells
3.6. Flow Cytometry Analysis of Bacteria-Cell-Association
3.7. NO Measurement
3.8. Evaluation of Cytotoxic Effects
3.9. Statistical Analysis
4. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Reams, R.Y.; Glickman, L.T.; Harrington, D.D.; Thacker, H.L.; Bowersock, T.L. Streptococcus suis infection in swine: A retrospective study of 256 cases. Part ii. Clinical signs, gross and microscopic lesions, and coexisting microorganisms. J. Vet. Diagn. Investig. 1994, 6, 326–334. [Google Scholar] [CrossRef] [PubMed]
- Straw, B.E. Diseases of Swine, 9th ed.; Blackwell Publisher: Ames, IA, USA, 2006. [Google Scholar]
- Huong, V.T.; Ha, N.; Huy, N.T.; Horby, P.; Nghia, H.D.; Thiem, V.D.; Zhu, X.; Hoa, N.T.; Hien, T.T.; Zamora, J.; et al. Epidemiology, clinical manifestations, and outcomes of Streptococcus suis infection in humans. Emerg. Infect. Dis. 2014, 20, 1105–1114. [Google Scholar] [CrossRef] [PubMed]
- Baums, C.G.; Valentin-Weigand, P. Surface-associated and secreted factors of Streptococcus suis in epidemiology, pathogenesis and vaccine development. Anim. Health Res. Rev. 2009, 10, 65–83. [Google Scholar] [CrossRef] [PubMed]
- Charland, N.; Harel, J.; Kobisch, M.; Lacasse, S.; Gottschalk, M. Streptococcus suis serotype 2 mutants deficient in capsular expression. Microbiology 1998, 144, 325–332. [Google Scholar] [CrossRef] [PubMed]
- Smith, H.E.; Damman, M.; van der Velde, J.; Wagenaar, F.; Wisselink, H.J.; Stockhofe-Zurwieden, N.; Smits, M.A. Identification and characterization of the cps locus of Streptococcus suis serotype 2: The capsule protects against phagocytosis and is an important virulence factor. Infect. Immun. 1999, 67, 1750–1756. [Google Scholar] [PubMed]
- Chabot-Roy, G.; Willson, P.; Segura, M.; Lacouture, S.; Gottschalk, M. Phagocytosis and killing of Streptococcus suis by porcine neutrophils. Microb. Pathog. 2006, 41, 21–32. [Google Scholar] [CrossRef] [PubMed]
- Seitz, M.; Beineke, A.; Singpiel, A.; Willenborg, J.; Dutow, P.; Goethe, R.; Valentin-Weigand, P.; Klos, A.; Baums, C.G. Role of capsule and suilysin in mucosal infection of complement-deficient mice with Streptococcus suis. Infect. Immun. 2014, 82, 2460–2471. [Google Scholar] [CrossRef] [PubMed]
- Fittipaldi, N.; Segura, M.; Grenier, D.; Gottschalk, M. Virulence factors involved in the pathogenesis of the infection caused by the swine pathogen and zoonotic agent Streptococcus suis. Future Microbiol. 2012, 7, 259–279. [Google Scholar] [CrossRef] [PubMed]
- Gottschalk, M.G.; Lacouture, S.; Dubreuil, J.D. Characterization of Streptococcus suis capsular type 2 haemolysin. Microbiology 1995, 141, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Benga, L.; Friedl, P.; Valentin-Weigand, P. Adherence of Streptococcus suis to porcine endothelial cells. J. Vet. Med. B Infect. Dis. Vet. Public Health 2005, 52, 392–395. [Google Scholar] [CrossRef] [PubMed]
- Charland, N.; Nizet, V.; Rubens, C.E.; Kim, K.S.; Lacouture, S.; Gottschalk, M. Streptococcus suis serotype 2 interactions with human brain microvascular endothelial cells. Infect. Immun. 2000, 68, 637–643. [Google Scholar] [CrossRef] [PubMed]
- Vanier, G.; Segura, M.; Friedl, P.; Lacouture, S.; Gottschalk, M. Invasion of porcine brain microvascular endothelial cells by Streptococcus suis serotype 2. Infect. Immun. 2004, 72, 1441–1449. [Google Scholar] [CrossRef] [PubMed]
- Vanier, G.; Segura, M.; Gottschalk, M. Characterization of the invasion of porcine endothelial cells by Streptococcus suis serotype 2. Can. J. Vet. Res. 2007, 71, 81–89. [Google Scholar] [PubMed]
- Tenenbaum, T.; Papandreou, T.; Gellrich, D.; Friedrichs, U.; Seibt, A.; Adam, R.; Wewer, C.; Galla, H.J.; Schwerk, C.; Schroten, H. Polar bacterial invasion and translocation of Streptococcus suis across the blood-cerebrospinal fluid barrier in vitro. Cell. Microbiol. 2009, 11, 323–336. [Google Scholar] [CrossRef] [PubMed]
- Sofroniew, M.V. Astrocyte barriers to neurotoxic inflammation. Nat. Rev. Neurosci. 2015, 16, 249–263. [Google Scholar] [CrossRef] [PubMed]
- Kimelberg, H.K.; Nedergaard, M. Functions of astrocytes and their potential as therapeutic targets. Neurotherapeutics 2010, 7, 338–353. [Google Scholar] [PubMed]
- Stoner, T.D.; Weston, T.A.; Trejo, J.; Doran, K.S. Group B streptococcal infection and activation of human astrocytes. PLoS ONE 2015, 10, e0128431. [Google Scholar] [CrossRef] [PubMed]
- Olson, J.K.; Miller, S.D. Microglia initiate central nervous system innate and adaptive immune responses through multiple tlrs. J. Immunol. 2004, 173, 3916–3924. [Google Scholar] [CrossRef] [PubMed]
- Min, K.J.; Yang, M.S.; Kim, S.U.; Jou, I.; Joe, E.H. Astrocytes induce hemeoxygenase-1 expression in microglia: A feasible mechanism for preventing excessive brain inflammation. J. Neurosci. 2006, 26, 1880–1887. [Google Scholar] [CrossRef] [PubMed]
- Ovanesov, M.V.; Ayhan, Y.; Wolbert, C.; Moldovan, K.; Sauder, C.; Pletnikov, M.V. Astrocytes play a key role in activation of microglia by persistent borna disease virus infection. J. Neuroinflammation 2008, 5, 50. [Google Scholar] [CrossRef] [PubMed]
- Carson, M.J.; Thrash, J.C.; Walter, B. The cellular response in neuroinflammation: The role of leukocytes, microglia and astrocytes in neuronal death and survival. Clin. Neurosci. Res. 2006, 6, 237–245. [Google Scholar] [CrossRef] [PubMed]
- Cooley, I.D.; Chauhan, V.S.; Donneyz, M.A.; Marriott, I. Astrocytes produce il-19 in response to bacterial challenge and are sensitive to the immunosuppressive effects of this il-10 family member. Glia 2014, 62, 818–828. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Chauhan, V.S.; Young, A.B.; Marriott, I. Nod2 mediates inflammatory responses of primary murine glia to Streptococcus pneumoniae. Glia 2010, 58, 839–847. [Google Scholar] [PubMed]
- Hansson, E.; Ronnback, L. Glial neuronal signaling in the central nervous system. FASEB J. 2003, 17, 341–348. [Google Scholar] [CrossRef] [PubMed]
- Bezzi, P.; Domercq, M.; Brambilla, L.; Galli, R.; Schols, D.; de Clercq, E.; Vescovi, A.; Bagetta, G.; Kollias, G.; Meldolesi, J.; et al. CXCR4-activated astrocyte glutamate release via tnfalpha: Amplification by microglia triggers neurotoxicity. Nat. Neurosci. 2001, 4, 702–710. [Google Scholar] [CrossRef] [PubMed]
- Dominguez-Punaro Mde, L.; Segura, M.; Contreras, I.; Lachance, C.; Houde, M.; Lecours, M.P.; Olivier, M.; Gottschalk, M. In vitro characterization of the microglial inflammatory response to Streptococcus suis, an important emerging zoonotic agent of meningitis. Infect Immun. 2010, 78, 5074–5085. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.; Punaro, M.C.; Segura, M.; Lachance, C.; Rivest, S.; Xu, J.; Houde, M.; Gottschalk, M. Toll-like receptor 2 is partially involved in the activation of murine astrocytes by Streptococcus suis, an important zoonotic agent of meningitis. J. Neuroimmunol. 2011, 234, 71–83. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.; Sun, H.; Dominguez-Punaro Mde, L.; Bai, X.; Ji, S.; Segura, M.; Xu, J. Evaluation of the pathogenesis of meningitis caused by Streptococcus suis sequence type 7 using the infection of bv2 microglial cells. J. Med. Microbiol. 2013, 62, 360–368. [Google Scholar] [CrossRef] [PubMed]
- Auger, J.P.; Christodoulides, M.; Segura, M.; Xu, J.; Gottschalk, M. Interactions of Streptococcus suis serotype 2 with human meningeal cells and astrocytes. BMC Res. Notes 2015, 8, 607. [Google Scholar] [CrossRef] [PubMed]
- Dowell, J.A.; Johnson, J.A.; Li, L. Identification of astrocyte secreted proteins with a combination of shotgun proteomics and bioinformatics. J. Proteome Res. 2009, 8, 4135–4143. [Google Scholar] [CrossRef] [PubMed]
- Seitz, M.; Baums, C.G.; Neis, C.; Benga, L.; Fulde, M.; Rohde, M.; Goethe, R.; Valentin-Weigand, P. Subcytolytic effects of suilysin on interaction of Streptococcus suis with epithelial cells. Vet. Microbiol. 2013, 167, 584–591. [Google Scholar] [CrossRef] [PubMed]
- Benga, L.; Fulde, M.; Neis, C.; Goethe, R.; Valentin-Weigand, P. Polysaccharide capsule and suilysin contribute to extracellular survival of Streptococcus suis co-cultivated with primary porcine phagocytes. Vet. Microbiol. 2008, 132, 211–219. [Google Scholar] [CrossRef] [PubMed]
- Segura, M.; Gottschalk, M. Streptococcus suis interactions with the murine macrophage cell line j774: Adhesion and cytotoxicity. Infect. Immun. 2002, 70, 4312–4322. [Google Scholar] [CrossRef] [PubMed]
- Norton, P.M.; Rolph, C.; Ward, P.N.; Bentley, R.W.; Leigh, J.A. Epithelial invasion and cell lysis by virulent strains of Streptococcus suis is enhanced by the presence of suilysin. FEMS Immunol Med. Microbiol. 1999, 26, 25–35. [Google Scholar] [CrossRef] [PubMed]
- Tenenbaum, T.; Adam, R.; Eggelnpohler, I.; Matalon, D.; Seibt, A.; Novotny, G.E.K.; Galla, H.J.; Schroten, H. Strain-dependent disruption of blood-cerebrospinal fluid barrier by Streptoccocus suis in vitro. FEMS Immunol. Med. Microbiol. 2005, 44, 25–34. [Google Scholar] [CrossRef] [PubMed]
- Sola, C.; Casal, C.; Tusell, J.M.; Serratosa, J. Astrocytes enhance lipopolysaccharide-induced nitric oxide production by microglial cells. Eur. J. Neurosci. 2002, 16, 1275–1283. [Google Scholar] [CrossRef] [PubMed]
- Chien, H.F.; Yeh, K.Y.; Jiang-Shieh, Y.F.; Wei, I.H.; Chang, C.Y.; Chang, M.L.; Wu, C.H. Signal transduction pathways of nitric oxide release in primary microglial culture challenged with gram-positive bacterial constituent, lipoteichoic acid. Neuroscience 2005, 133, 423–436. [Google Scholar] [CrossRef] [PubMed]
- Wiley, J. The many faces of nitric oxide: Cytotoxic, cytoprotective or both. Neurogastroenterol. Motil. 2007, 19, 541–544. [Google Scholar] [CrossRef] [PubMed]
- Baums, C.G.; Kaim, U.; Fulde, M.; Ramachandran, G.; Goethe, R.; Valentin-Weigand, P. Identification of a novel virulence determinant with serum opacification activity in Streptococcus suis. Infect. Immun. 2006, 74, 6154–6162. [Google Scholar] [CrossRef] [PubMed]
- Baums, C.G.; Kock, C.; Beineke, A.; Bennecke, K.; Goethe, R.; Schroder, C.; Waldmann, K.H.; Valentin-Weigand, P. Streptococcus suis bacterin and subunit vaccine immunogenicities and protective efficacies against serotypes 2 and 9. Clin. Vaccine Immunol. 2009, 16, 200–208. [Google Scholar] [CrossRef] [PubMed]
- De Greeff, A.; Buys, H.; Verhaar, R.; Dijkstra, J.; van Alphen, L.; Smith, H.E. Contribution of fibronectin-binding protein to pathogenesis of Streptococcus suis serotype 2. Infect. Immun. 2002, 70, 1319–1325. [Google Scholar] [CrossRef] [PubMed]
- Smith, H.E.; Vecht, U.; Wisselink, H.J.; Stockhofe-Zurwieden, N.; Biermann, Y.; Smits, M.A. Mutants of Streptococcus suis types 1 and 2 impaired in expression of muramidase-released protein and extracellular protein induce disease in newborn germfree pigs. Infect. Immun. 1996, 64, 4409–4412. [Google Scholar] [PubMed]
- Council of Europe. Details of Treaty No.123. Available online: http://conventions.coe.int/treaty/en/treaties/html/123.htm (accessed on 7 June 2016).
- Redlich, S.; Ribes, S.; Schutze, S.; Eiffert, H.; Nau, R. Toll-like receptor stimulation increases phagocytosis of Cryptococcus neoformans by microglial cells. J. Neuroinflammation 2013, 10, 71. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Benardais, K.; Stanslowsky, N.; Thau-Habermann, N.; Hensel, N.; Huang, D.; Claus, P.; Dengler, R.; Stangel, M.; Petri, S. Therapeutic potential of mesenchymal stromal cells and msc conditioned medium in amyotrophic lateral sclerosis (als)--in vitro evidence from primary motor neuron cultures, nsc-34 cells, astrocytes and microglia. PLoS ONE 2013, 8, e72926. [Google Scholar] [CrossRef] [PubMed]
- Benga, L.; Goethe, R.; Rohde, M.; Valentin-Weigand, P. Non-encapsulated strains reveal novel insights in invasion and survival of Streptococcus suis in epithelial cells. Cell. Microbiol. 2004, 6, 867–881. [Google Scholar] [CrossRef] [PubMed]





© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Seele, J.; Nau, R.; Prajeeth, C.K.; Stangel, M.; Valentin-Weigand, P.; Seitz, M. Astrocytes Enhance Streptococcus suis-Glial Cell Interaction in Primary Astrocyte-Microglial Cell Co-Cultures. Pathogens 2016, 5, 43. https://doi.org/10.3390/pathogens5020043
Seele J, Nau R, Prajeeth CK, Stangel M, Valentin-Weigand P, Seitz M. Astrocytes Enhance Streptococcus suis-Glial Cell Interaction in Primary Astrocyte-Microglial Cell Co-Cultures. Pathogens. 2016; 5(2):43. https://doi.org/10.3390/pathogens5020043
Chicago/Turabian StyleSeele, Jana, Roland Nau, Chittappen K. Prajeeth, Martin Stangel, Peter Valentin-Weigand, and Maren Seitz. 2016. "Astrocytes Enhance Streptococcus suis-Glial Cell Interaction in Primary Astrocyte-Microglial Cell Co-Cultures" Pathogens 5, no. 2: 43. https://doi.org/10.3390/pathogens5020043
APA StyleSeele, J., Nau, R., Prajeeth, C. K., Stangel, M., Valentin-Weigand, P., & Seitz, M. (2016). Astrocytes Enhance Streptococcus suis-Glial Cell Interaction in Primary Astrocyte-Microglial Cell Co-Cultures. Pathogens, 5(2), 43. https://doi.org/10.3390/pathogens5020043