
Recruitment of Factor H to the Streptococcus suis Cell Surface is Multifactorial


Abstract
:1. Introduction
2. Results
2.1. Mutant Characterization
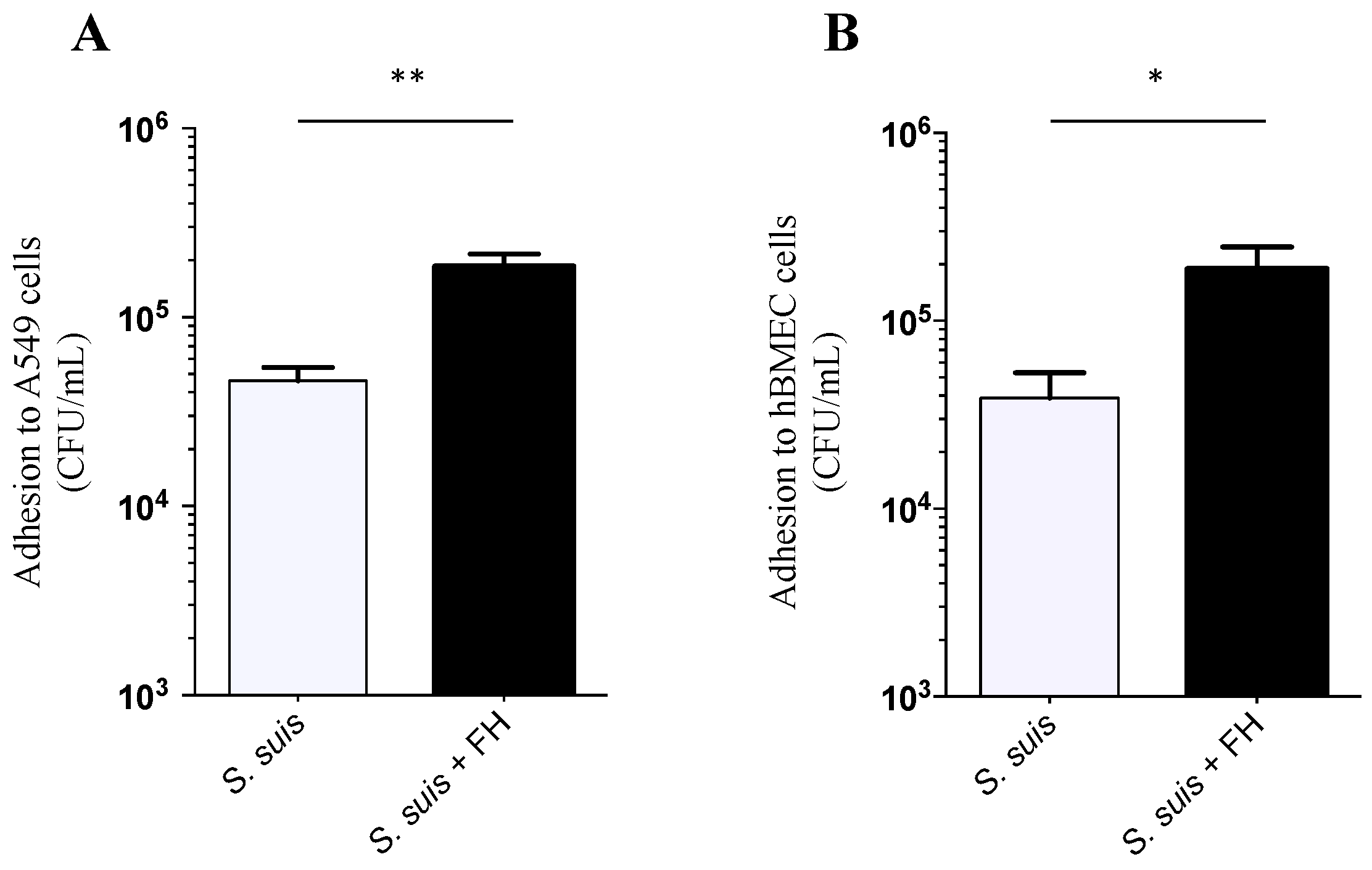
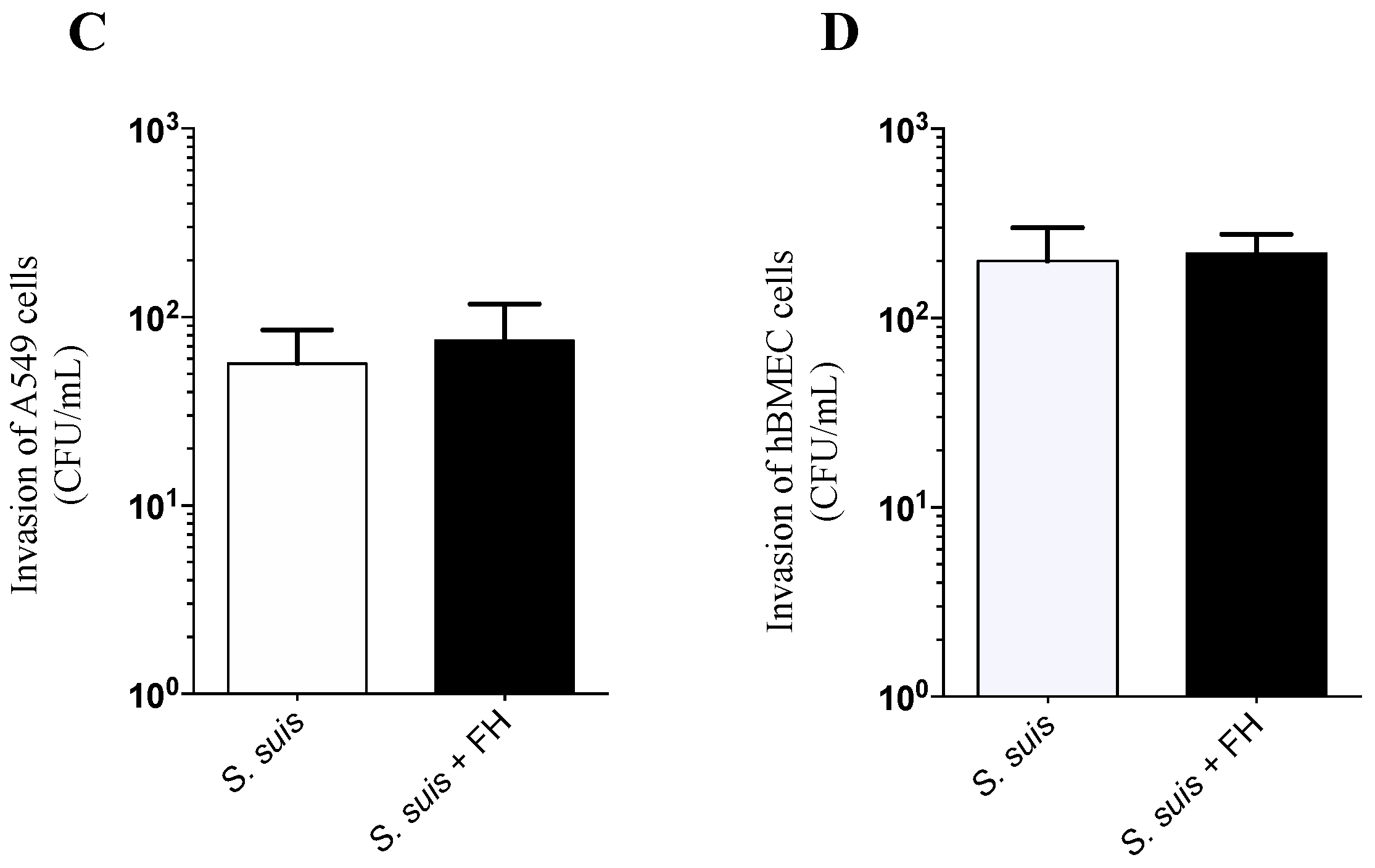
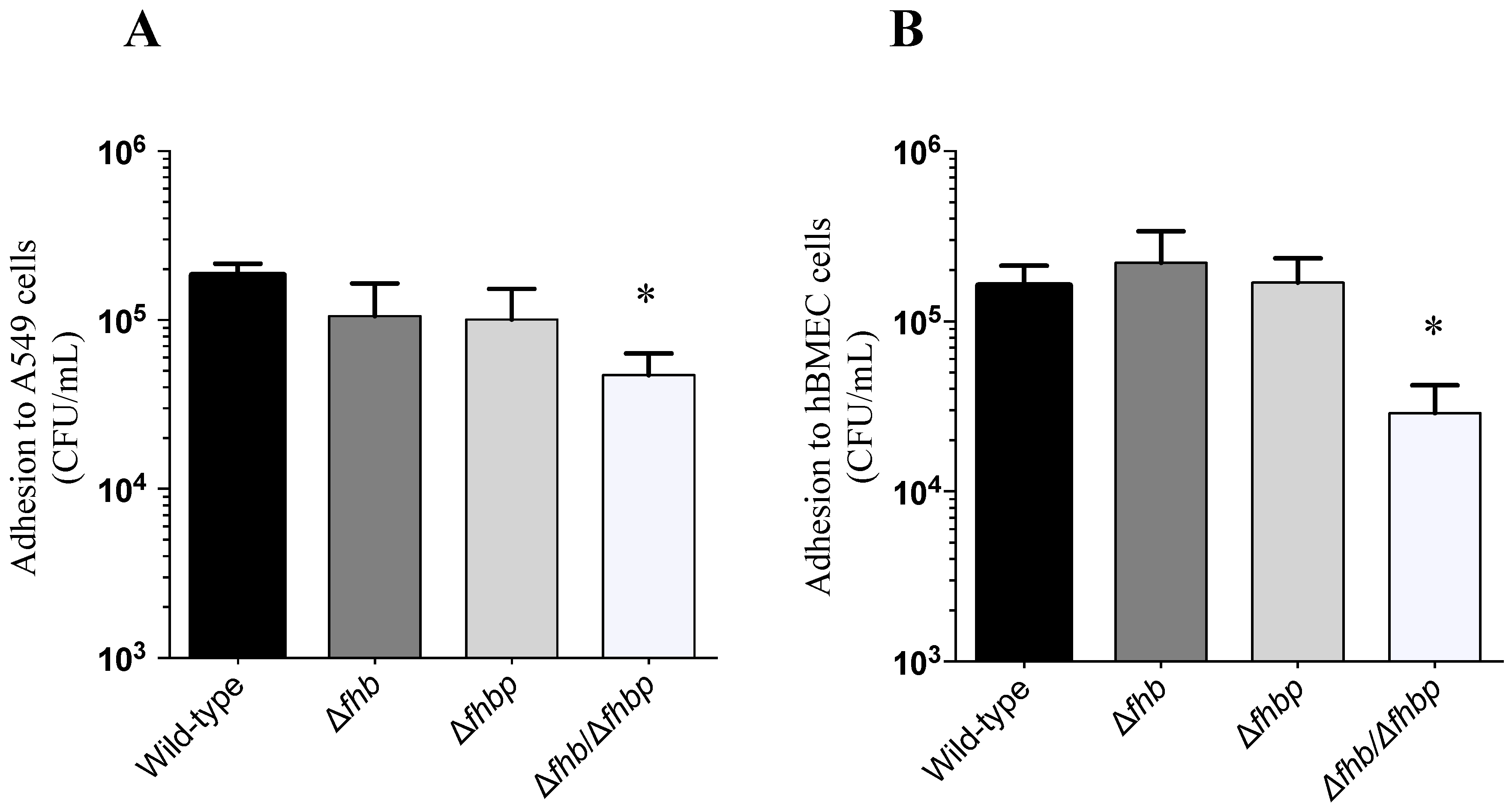
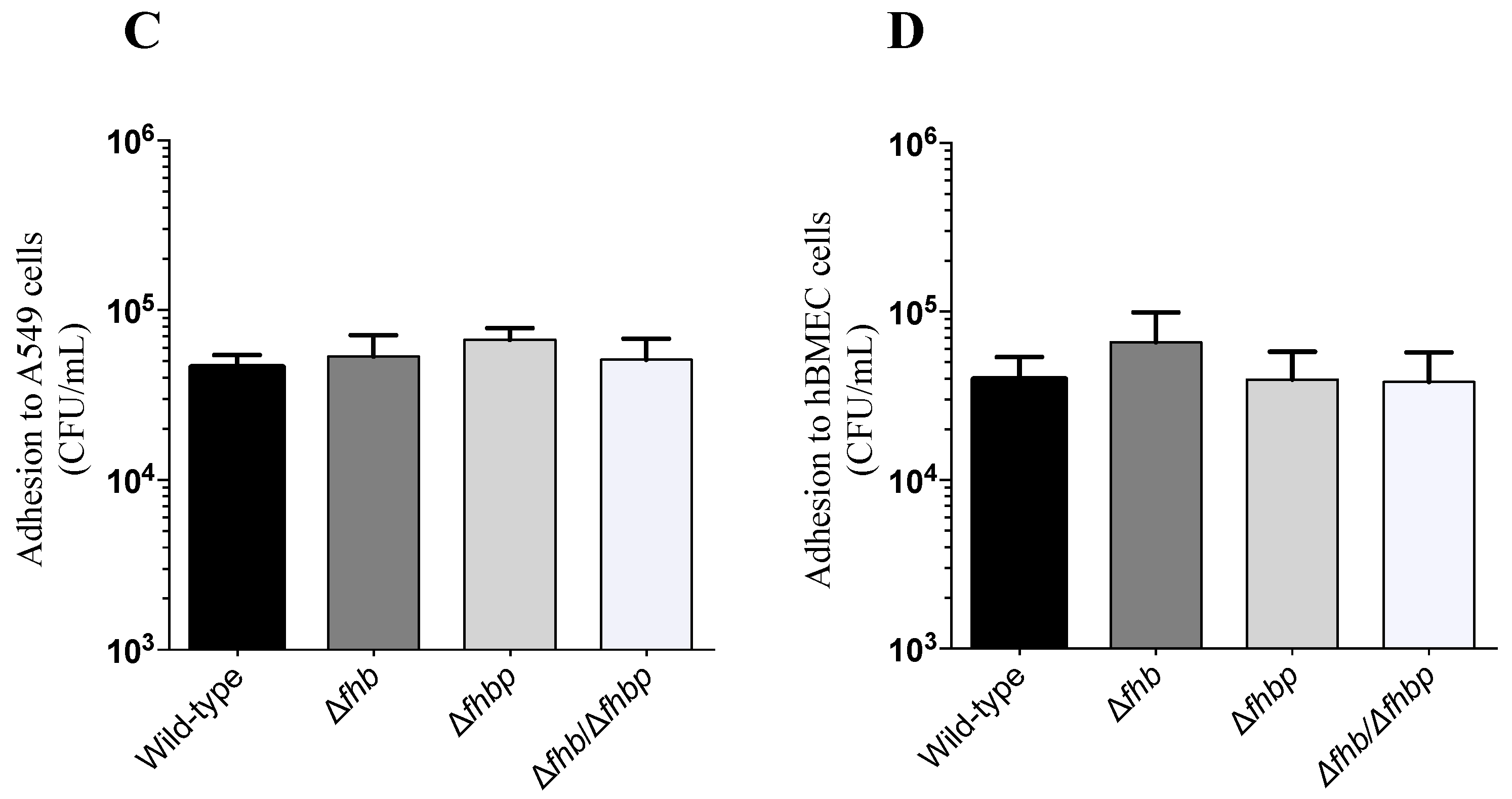
2.2. Adhesion and Invasion Assays
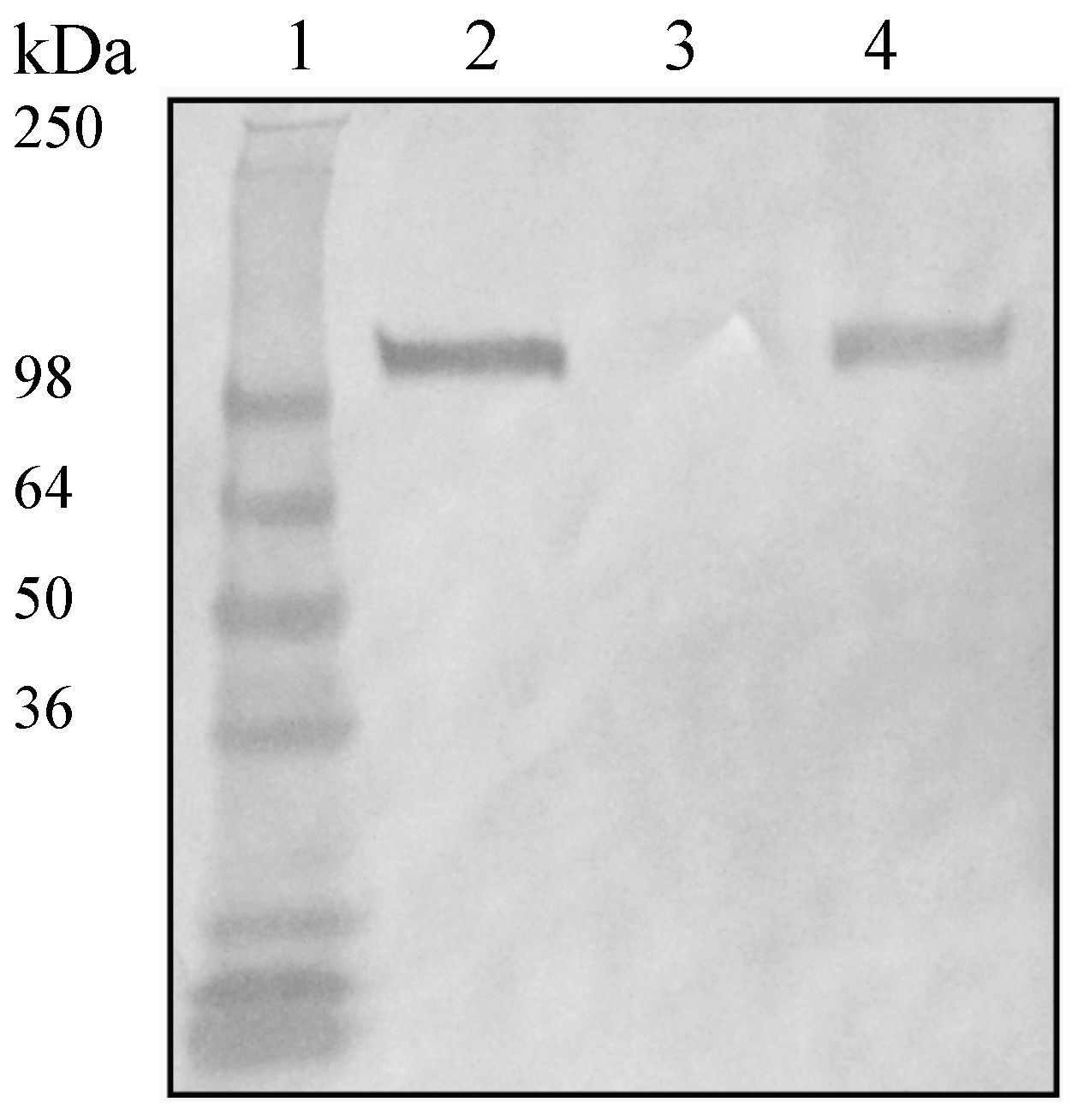
2.3. Evaluation of the Role of Factor H Binding Proteins on S. suis Factor H Deposition
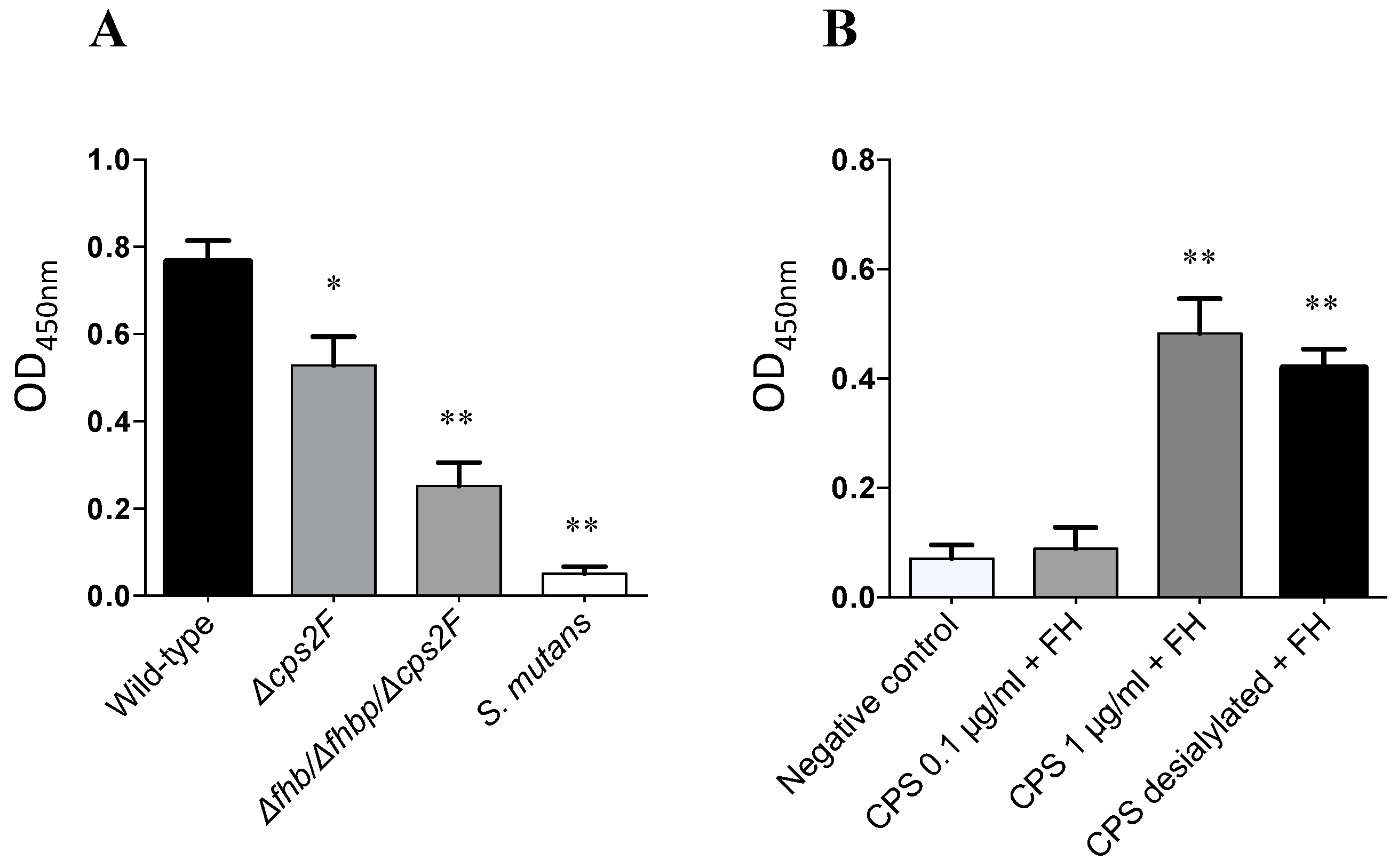
2.4. Evaluation of the Role of CPS for S. suis Factor H Deposition
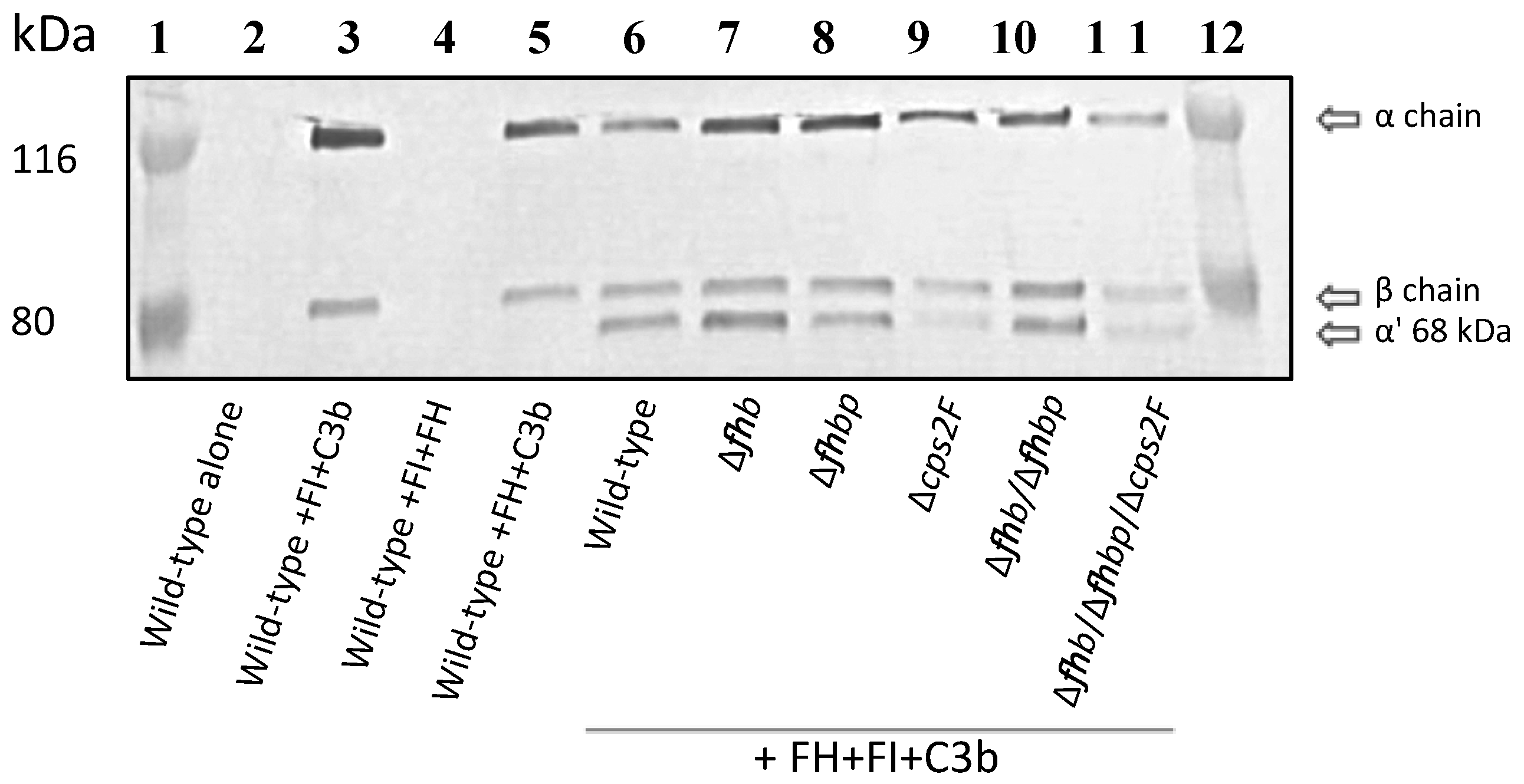
2.5. C3b Cleavage Assay
2.6. Phagocytosis Assay
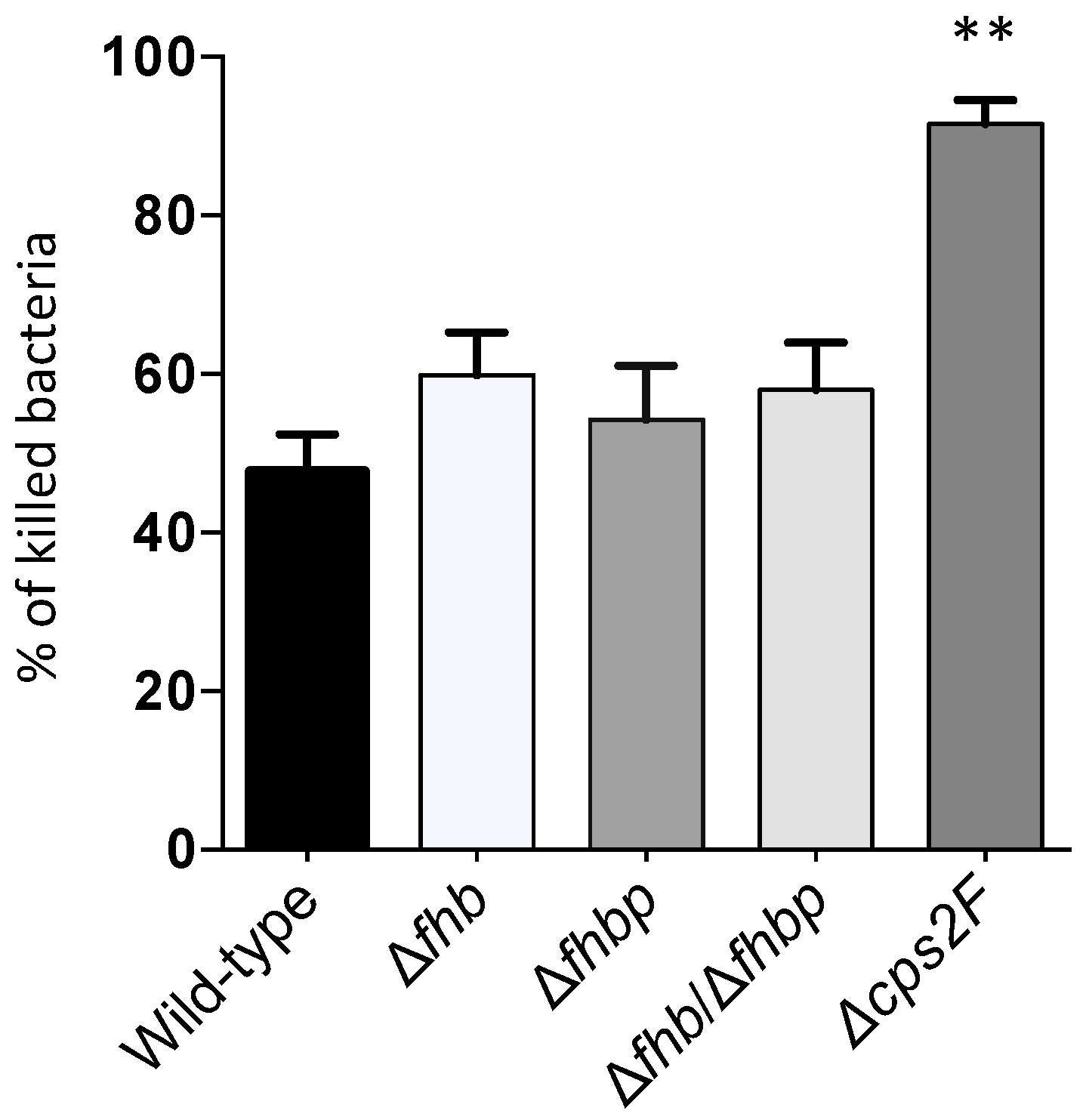
2.7. Whole Blood Bacterial Killing Assay
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains and Culture Conditions
4.2. Cell Lines and Cell Culture
4.3. DNA Manipulations
4.4. Construction of Allelic Deletion Mutants
4.5. Construction of Complemented Δfhbp Mutant
4.6. Adhesion and Invasion Assays
4.7. Evaluation of Factor H Deposition to S. suis Strains
4.8. Factor H Recruitment by the Capsular Polysaccharide
4.9. C3b Cleavage Assay
4.10. Phagocytosis Assays
4.11. Whole Blood Bacterial Killing Assay
4.12. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| CFU | colony-forming unit |
| CPS | capsular polysaccharide |
| Km | kanamycin |
| hBMEC | human brain microvascular endothelial cells |
| MOI | multiplicity of infection |
| PBS | phosphate-buffered saline |
| PBS-T | PBS-tween |
| SEM | standard error of the mean |
| Sp | spectinomycin |
| THA | Todd-Hewitt agar |
| THB | Todd-Hewitt broth |
References
- Gottschalk, M. Streptococcocis, 10th ed.; Wiley-Blackwell: Ames, IA, USA, 2012. [Google Scholar]
- Gottschalk, M.; Xu, J.; Calzas, C.; Segura, M. Streptococcus suis: A new emerging or an old neglected zoonotic pathogen? Future Microbiol. 2010, 5, 371–391. [Google Scholar] [CrossRef] [PubMed]
- Wertheim, H.F.; Nghia, H.D.; Taylor, W.; Schultsz, C. Streptococcus suis: An emerging human pathogen. Clin. Infect. Dis. 2009, 48, 617–625. [Google Scholar] [CrossRef] [PubMed]
- Goyette-Desjardins, G.; Auger, J.P.; Xu, J.; Segura, M.; Gottschalk, M. Streptococcus suis, an important pig pathogen and emerging zoonotic agent-an update on the worldwide distribution based on serotyping and sequence typing. Emergy Microb. Infect. 2014, 3, e45. [Google Scholar] [CrossRef] [PubMed]
- Fittipaldi, N.; Segura, M.; Grenier, D.; Gottschalk, M. Virulence factors involved in the pathogenesis of the infection caused by the swine pathogen and zoonotic agent Streptococcus suis. Future Microbiol. 2012, 7, 259–279. [Google Scholar] [CrossRef] [PubMed]
- Zipfel, P.F. Complement factor h: Physiology and pathophysiology. Semin. Thromb. Hemost. 2001, 27, 191–199. [Google Scholar] [CrossRef] [PubMed]
- de Cordoba, S.R.; de Jorge, E.G. Translational mini-review series on complement factor h: Genetics and disease associations of human complement factor h. Clin. Exp. Immunol. 2008, 151, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Welsch, J.A.; Ram, S. Factor h and neisserial pathogenesis. Vaccine 2008, 26 (Suppl. 8), I40–I45. [Google Scholar] [CrossRef] [PubMed]
- Keller, L.E.; Jones, C.V.; Thornton, J.A.; Sanders, M.E.; Swiatlo, E.; Nahm, M.H.; Park, I.H.; McDaniel, L.S. Pspk of Streptococcus pneumoniae increases adherence to epithelial cells and enhances nasopharyngeal colonization. Infect. Immun. 2013, 81, 173–181. [Google Scholar] [CrossRef] [PubMed]
- Vaillancourt, K.; Bonifait, L.; Grignon, L.; Frenette, M.; Gottschalk, M.; Grenier, D. Identification and characterization of a new cell surface protein possessing factor h-binding activity in the swine pathogen and zoonotic agent Streptococcus suis. J. Med. Microbiol. 2013, 62, 1073–1080. [Google Scholar] [CrossRef] [PubMed]
- Pian, Y.; Gan, S.; Wang, S.; Guo, J.; Wang, P.; Zheng, Y.; Cai, X.; Jiang, Y.; Yuan, Y. Fhb, a novel factor h-binding surface protein, contributes to the antiphagocytic ability and virulence of Streptococcus suis. Infect. Immun. 2012, 80, 2402–2413. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Liu, L.; Qiu, D.; Chen, H.; Zhou, R. Identification of Streptococcus suis serotype 2 genes preferentially expressed in the natural host. Int. J. Med. Microbiol. 2010, 300, 482–488. [Google Scholar] [CrossRef] [PubMed]
- Lecours, M.P.; Gottschalk, M.; Houde, M.; Lemire, P.; Fittipaldi, N.; Segura, M. Critical role for Streptococcus suis cell wall modifications and suilysin in resistance to complement-dependent killing by dendritic cells. J. Infect. Dis. 2011, 204, 919–929. [Google Scholar] [CrossRef] [PubMed]
- Brazeau, C.; Gottschalk, M.; Vincelette, S.; Martineau-Doize, B. In vitro phagocytosis and survival of Streptococcus suis capsular type 2 inside murine macrophages. Microbiology 1996, 142, 1231–1237. [Google Scholar] [CrossRef] [PubMed]
- Seitz, M.; Beineke, A.; Singpiel, A.; Willenborg, J.; Dutow, P.; Goethe, R.; Valentin-Weigand, P.; Klos, A.; Baums, C.G. Role of capsule and suilysin in mucosal infection of complement-deficient mice with Streptococcus suis. Infect. Immun. 2014, 82, 2460–2471. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez de Cordoba, S.; Esparza-Gordillo, J.; Goicoechea de Jorge, E.; Lopez-Trascasa, M.; Sanchez-Corral, P. The human complement factor h: Functional roles, genetic variations and disease associations. Mol. Immunol. 2004, 41, 355–367. [Google Scholar] [CrossRef] [PubMed]
- Horstmann, R.D.; Sievertsen, H.J.; Knobloch, J.; Fischetti, V.A. Antiphagocytic activity of streptococcal m protein: Selective binding of complement control protein factor h. Proc. Natl. Acad. Sci. USA 1988, 85, 1657–1661. [Google Scholar] [CrossRef] [PubMed]
- Biedzka-Sarek, M.; Jarva, H.; Hyytiainen, H.; Meri, S.; Skurnik, M. Characterization of complement factor h binding to Yersinia enterocolitica serotype o:3. Infect. Immun. 2008, 76, 4100–4109. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, V.; Asmat, T.M.; Luo, S.; Jensch, I.; Zipfel, P.F.; Hammerschmidt, S. Complement regulator factor h mediates a two-step uptake of Streptococcus pneumoniae by human cells. J. Biol. Chem. 2010, 285, 23486–23495. [Google Scholar] [CrossRef] [PubMed]
- Meri, T.; Amdahl, H.; Lehtinen, M.J.; Hyvarinen, S.; McDowell, J.V.; Bhattacharjee, A.; Meri, S.; Marconi, R.; Goldman, A.; Jokiranta, T.S. Microbes bind complement inhibitor factor h via a common site. PLoS Pathog. 2013, 9, e1003308. [Google Scholar] [CrossRef]
- Quin, L.R.; Onwubiko, C.; Moore, Q.C.; Mills, M.F.; McDaniel, L.S.; Carmicle, S. Factor h binding to pspc of Streptococcus pneumoniae increases adherence to human cell lines in vitro and enhances invasion of mouse lungs in vivo. Infect. Immun. 2007, 75, 4082–4087. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.M.; Hao, H.J.; Yu, Y.; Kong, D.C.; Chen, S.L.; Jiang, H.; Yuan, Y.; Zheng, Y.L.; Yang, M.J.; Jiang, Y.Q. Structural basis of the interaction between the meningitis pathogen Streptococcus suis adhesin fhb and its human receptor. FEBS Lett. 2016. [Google Scholar] [CrossRef] [PubMed]
- Kouki, A.; Haataja, S.; Loimaranta, V.; Pulliainen, A.T.; Nilsson, U.J.; Finne, J. Identification of a novel streptococcal adhesin p (sadp) protein recognizing galactosyl-alpha1–4-galactose-containing glycoconjugates: Convergent evolution of bacterial pathogens to binding of the same host receptor. J. Biol. Chem. 2011, 286, 38854–38864. [Google Scholar] [CrossRef] [PubMed]
- Schweppe, C.H.; Bielaszewska, M.; Pohlentz, G.; Friedrich, A.W.; Buntemeyer, H.; Schmidt, M.A.; Kim, K.S.; Peter-Katalinic, J.; Karch, H.; Muthing, J. Glycosphingolipids in vascular endothelial cells: Relationship of heterogeneity in gb3cer/cd77 receptor expression with differential shiga toxin 1 cytotoxicity. Glycoconj. J. 2008, 25, 291–304. [Google Scholar] [CrossRef] [PubMed]
- Hammerschmidt, S.; Agarwal, V.; Kunert, A.; Haelbich, S.; Skerka, C.; Zipfel, P.F. The host immune regulator factor h interacts via two contact sites with the pspc protein of Streptococcus pneumoniae and mediates adhesion to host epithelial cells. J. Immunol. 2007, 178, 5848–5858. [Google Scholar] [CrossRef] [PubMed]
- Ram, S.; Sharma, A.K.; Simpson, S.D.; Gulati, S.; McQuillen, D.P.; Pangburn, M.K.; Rice, P.A. A novel sialic acid binding site on factor h mediates serum resistance of sialylated Neisseria gonorrhoeae. J. Exp. Med. 1998, 187, 743–752. [Google Scholar] [CrossRef] [PubMed]
- Langford-Smith, A.; Day, A.J.; Bishop, P.N.; Clark, S.J. Complementing the sugar code: Role of gags and sialic acid in complement regulation. Front. Immunol. 2015, 6, 25. [Google Scholar] [CrossRef] [PubMed]
- Van Calsteren, M.R.; Gagnon, F.; Lacouture, S.; Fittipaldi, N.; Gottschalk, M. Structure determination of Streptococcus suis serotype 2 capsular polysaccharide. Biochem. Cell. Biol. 2010, 88, 513–525. [Google Scholar] [CrossRef] [PubMed]
- Meijerink, M.; Ferrando, M.L.; Lammers, G.; Taverne, N.; Smith, H.E.; Wells, J.M. Immunomodulatory effects of Streptococcus suis capsule type on human dendritic cell responses, phagocytosis and intracellular survival. PLoS ONE 2012, 7, e35849. [Google Scholar] [CrossRef] [PubMed]
- Hyams, C.; Trzcinski, K.; Camberlein, E.; Weinberger, D.M.; Chimalapati, S.; Noursadeghi, M.; Lipsitch, M.; Brown, J.S. Streptococcus pneumoniae capsular serotype invasiveness correlates with the degree of factor h binding and opsonization with c3b/ic3b. Infect. Immun. 2013, 81, 354–363. [Google Scholar] [CrossRef] [PubMed]
- McNeil, L.K.; Murphy, E.; Zhao, X.J.; Guttmann, S.; Harris, S.L.; Scott, A.A.; Tan, C.; Mack, M.; DaSilva, I.; Alexander, K.; et al. Detection of lp2086 on the cell surface of Neisseria meningitidis and its accessibility in the presence of serogroup b capsular polysaccharide. Vaccine 2009, 27, 3417–3421. [Google Scholar] [CrossRef] [PubMed]
- Marques, M.B.; Kasper, D.L.; Pangburn, M.K.; Wessels, M.R. Prevention of c3 deposition by capsular polysaccharide is a virulence mechanism of type iii group b streptococci. Infect. Immun. 1992, 60, 3986–3993. [Google Scholar] [PubMed]
- Lecours, M.P.; Fittipaldi, N.; Takamatsu, D.; Okura, M.; Segura, M.; Goyette-Desjardins, G.; Van Calsteren, M.R.; Gottschalk, M. Sialylation of Streptococcus suis serotype 2 is essential for capsule expression but is not responsible for the main capsular epitope. Microb. Infect. 2012, 14, 941–950. [Google Scholar] [CrossRef] [PubMed]
- Roy, D.; Auger, J.P.; Segura, M.; Fittipaldi, N.; Takamatsu, D.; Okura, M.; Gottschalk, M. Role of the capsular polysaccharide as a virulence factor for Streptococcus suis serotype 14. Can. J. Vet. Res. 2015, 79, 141–146. [Google Scholar] [PubMed]
- Lakkitjaroen, N.; Takamatsu, D.; Okura, M.; Sato, M.; Osaki, M.; Sekizaki, T. Capsule loss or death: The position of mutations among capsule genes sways the destiny of Streptococcus suis. FEMS Microbiol. Lett 2014, 354, 46–54. [Google Scholar] [CrossRef] [PubMed]
- Roy, D.; Fittipaldi, N.; Dumesnil, A.; Lacouture, S.; Gottschalk, M. The protective protein sao (surface antigen one) is not a critical virulence factor for Streptococcus suis serotype 2. Microb. Pathog. 2014, 67–68, 31–35. [Google Scholar] [CrossRef] [PubMed]
- Shao, Z.; Pan, X.; Li, X.; Liu, W.; Han, M.; Wang, C.; Wang, J.; Zheng, F.; Cao, M.; Tang, J. Htps, a novel immunogenic cell surface-exposed protein of Streptococcus suis, confers protection in mice. FEMS Microbiol. Lett. 2011, 314, 174–182. [Google Scholar] [CrossRef] [PubMed]
- Casadaban, M.J.; Cohen, S.N. Analysis of gene control signals by DNA fusion and cloning in Escherichia coli. J. Mol. Biol. 1980, 138, 179–207. [Google Scholar] [CrossRef]
- Slater, J.D.; Allen, A.G.; May, J.P.; Bolitho, S.; Lindsay, H.; Maskell, D.J. Mutagenesis of Streptococcus equi and Streptococcus suis by transposon tn917. Vet. Microbiol. 2003, 93, 197–206. [Google Scholar] [CrossRef]
- Takamatsu, D.; Osaki, M.; Sekizaki, T. Thermosensitive suicide vectors for gene replacement in Streptococcus suis. Plasmid 2001, 46, 140–148. [Google Scholar] [CrossRef] [PubMed]
- Okura, M.; Osaki, M.; Fittipaldi, N.; Gottschalk, M.; Sekizaki, T.; Takamatsu, D. The minor pilin subunit sgp2 is necessary for assembly of the pilus encoded by the srtg cluster of Streptococcus suis. J. Bacteriol. 2011, 193, 822–831. [Google Scholar] [CrossRef] [PubMed]
- Charland, N.; Nizet, V.; Rubens, C.E.; Kim, K.S.; Lacouture, S.; Gottschalk, M. Streptococcus suis serotype 2 interactions with human brain microvascular endothelial cells. Infect. Immun. 2000, 68, 637–643. [Google Scholar] [CrossRef] [PubMed]
- Segura, M.; Vadeboncoeur, N.; Gottschalk, M. Cd14-dependent and-independent cytokine and chemokine production by human thp-1 monocytes stimulated by Streptococcus suis capsular type 2. Clin. Exp. Immunol. 2002, 127, 243–254. [Google Scholar] [CrossRef] [PubMed]
- Warrens, A.N.; Jones, M.D.; Lechler, R.I. Splicing by overlap extension by pcr using asymmetric amplification: An improved technique for the generation of hybrid proteins of immunological interest. Gene 1997, 186, 29–35. [Google Scholar] [CrossRef]
- Burnette, W.N. "Western blotting": Electrophoretic transfer of proteins from sodium dodecyl sulfate—Polyacrylamide gels to unmodified nitrocellulose and radiographic detection with antibody and radioiodinated protein a. Anal. Biochem. 1981, 112, 195–203. [Google Scholar] [CrossRef]
- Gottschalk, M.; Higgins, R.; Jacques, M.; Mittal, K.R.; Henrichsen, J. Description of 14 new capsular types of Streptococcus suis. J. Clin. Microbiol. 1989, 27, 2633–2636. [Google Scholar] [PubMed]
- Charland, N.; Jacques, M.; Lacouture, S.; Gottschalk, M. Characterization and protective activity of a monoclonal antibody against a capsular epitope shared by Streptococcus suis serotypes 1, 2 and 1/2. Microbiology 1997, 143, 3607–3614. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Gagnon, C.A.; Savard, C.; Music, N.; Srednik, M.; Segura, M.; Lachance, C.; Bellehumeur, C.; Gottschalk, M. Capsular sialic acid of Streptococcus suis serotype 2 binds to swine influenza virus and enhances bacterial interactions with virus-infected tracheal epithelial cells. Infect. Immun. 2013, 81, 4498–4508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lalonde, M.; Segura, M.; Lacouture, S.; Gottschalk, M. Interactions between Streptococcus suis serotype 2 and different epithelial cell lines. Microbiology 2000, 146, 1913–1921. [Google Scholar] [CrossRef] [PubMed]
- Calzas, C.; Lemire, P.; Auray, G.; Gerdts, V.; Gottschalk, M.; Segura, M. Antibody response specific to the capsular polysaccharide is impaired in Streptococcus suis serotype 2-infected animals. Infect. Immun. 2015, 83, 441–453. [Google Scholar] [CrossRef] [PubMed]
- Segura, M.; Vanier, G.; Al-Numani, D.; Lacouture, S.; Olivier, M.; Gottschalk, M. Proinflammatory cytokine and chemokine modulation by Streptococcus suis in a whole-blood culture system. FEMS Immunol. Med. Microbiol. 2006, 47, 92–106. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
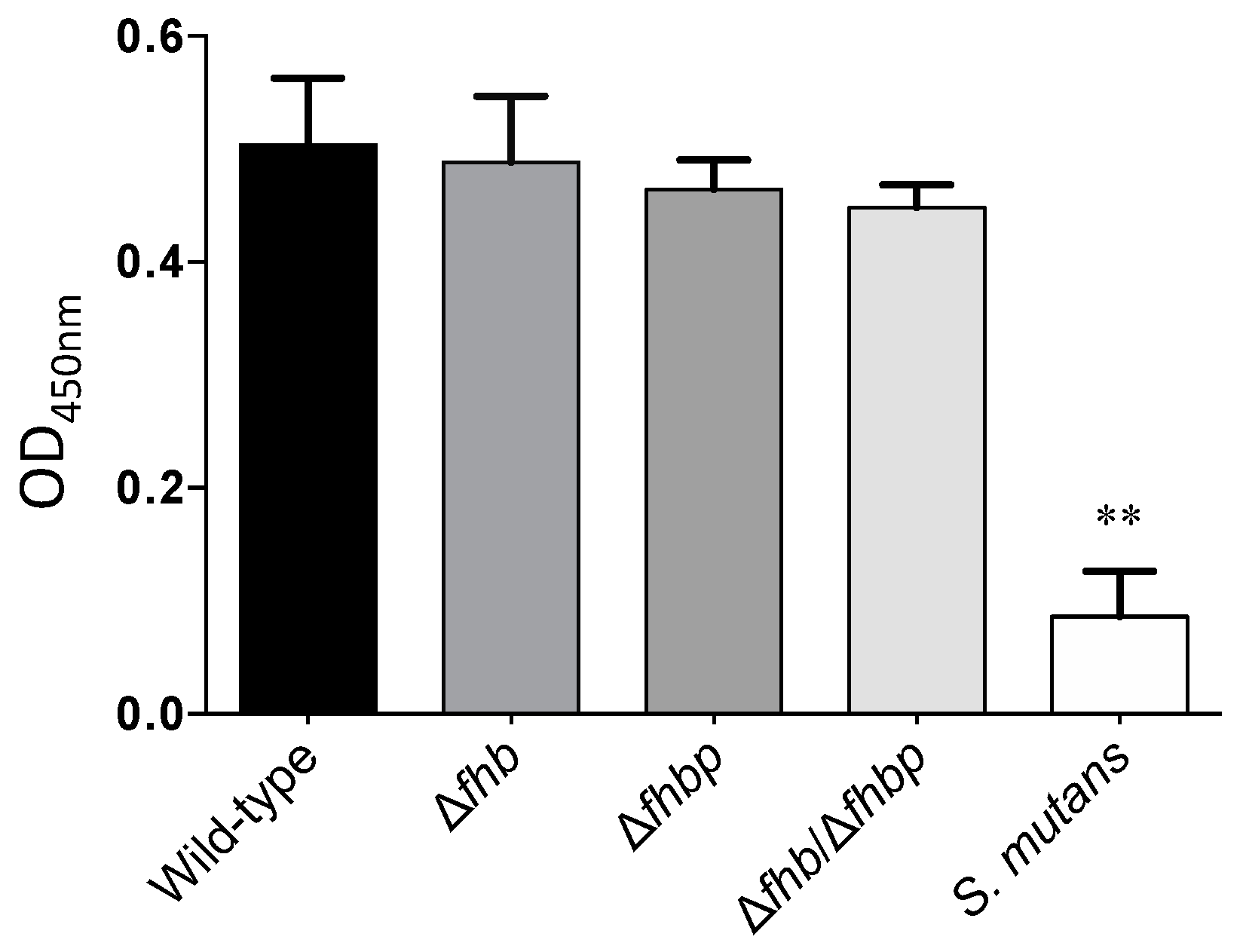{kind=link}
{kind=link}
{kind=link}
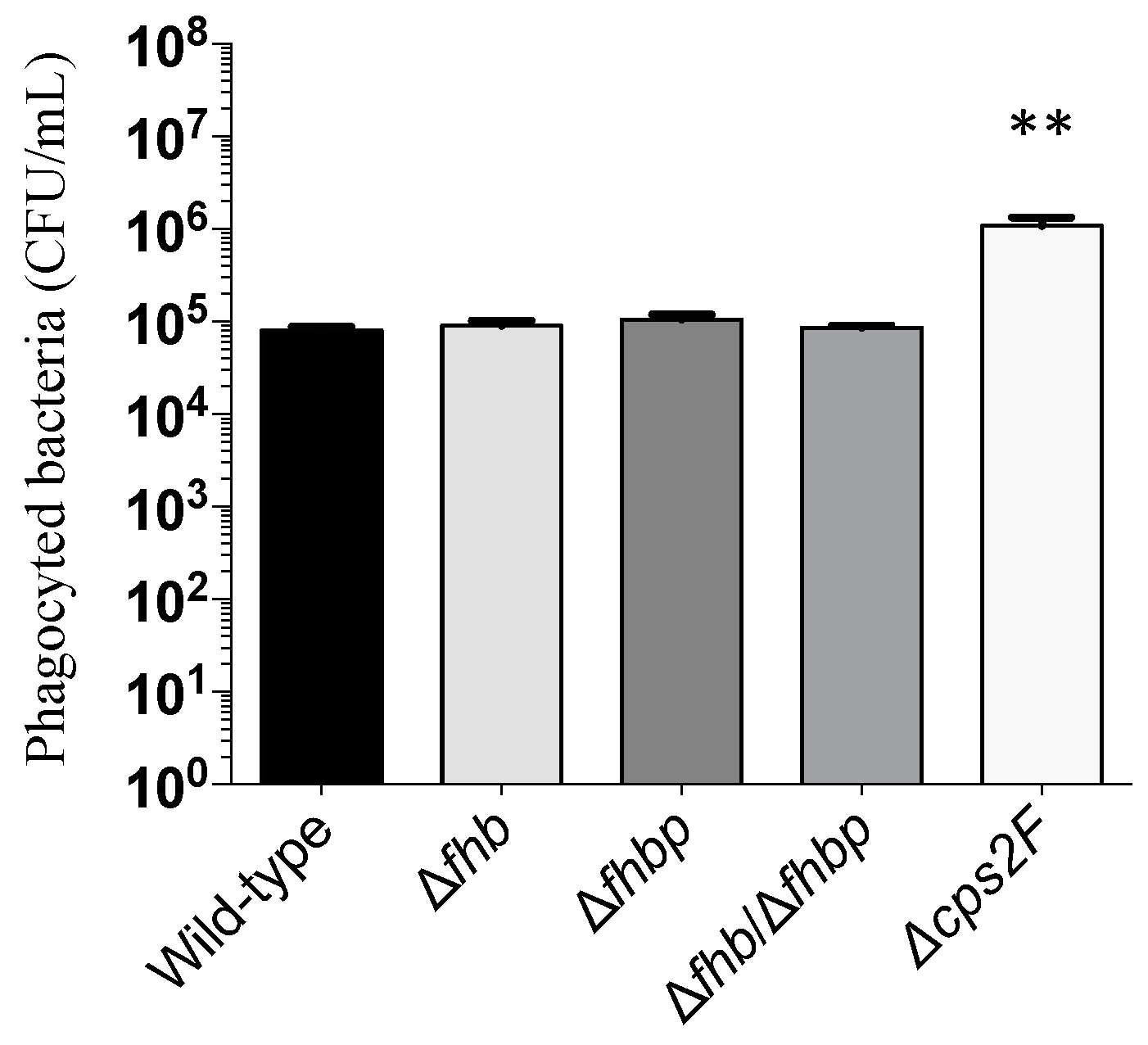{kind=link}
{kind=link}
| Strains/Plasmid | General Characteristics | Source/Reference |
|---|---|---|
| Escherichia coli | ||
| TOP 10 | F-mrcA Δ(mrr-hsdRMS-mcrBC)ϕ80 lacZΔM5 ΔlacX74 recA1 araD139 Δ(ara-leu) 7697 galU galK rpsL (StrR) endA1 nupG | Invitrogen |
| MC1061 | araD139 Δ(ara-leu)7697 ΔlacX74 galU galK hsdR2(rK-mK+) mcrB1 rpsL | [38] |
| Streptococcus suis | ||
| P1/7 | Wild-type strain, highly encapsulated serotype 2 strain isolated from a clinical swine case of infection in the United Kingdom | [39] |
| Δcps2F | Non-encapsulated isogenic mutant strain derived from strain P1/7. Deletion of the cps2F gene | [13] |
| Δfhb | Fhb expression-deficient strain derived from strain P1/7. Deletion of the fhb gene (SSU0253) | This work |
| Δfhbp | Fhbp expression-deficient strain derived from strain P1/7. Deletion of the fhbp gene (SSU0186) | This work |
| Δfhb/Δfhbp | Fhb and Fhbp expression-deficient strain derived from strain P1/7. Deletion of the fhb and fhbp genes | This work |
| Δfhb/Δfhbp/Δcps2F | Non-encapsulated mutant derived from strain Δfhb/Δfhbp. Deletion of the cps2F gene | This work |
| compΔfhbp | Mutant Δfhbp complemented with pMXfhbp complementation vector | This work |
| Streptococcus mutans 25175 | ||
| Wild-type strain, isolated from a carious dentine case | ATCC 25175 | |
| Plasmids | ||
| pCR2.1 | Apr, Kmr, oriR(f1) MCS oriR (ColE1) | Invitrogen |
| pSET-4s | Thermosensitive vector for allelic replacement. Replication functions of pG+host3, MCS oriR pUC19 lacZ SpR | [40] |
| pMX1 | Replication functions of pSSU1, MCS pUC19 lacZ SpR, malX promoter of S. suis, derivative of pSET2 | [40,41] |
| p4Δfhb | pSET-4s carrying the construct for fhb allelic replacement | This work |
| p4Δfhbp | pSET-4s carrying the construct for fhbp allelic replacement | This work |
| p4Δcps2F | pSET-4s carrying the construct for cps2F allelic replacement | This work |
| pMXfhbp | pMX1 complementation vector carrying intact fhbp gene | This work |
| Oligonucleotide Primers, Sequence (5’–3’) a | Constructs | |
|---|---|---|
| fhbp-ID1 | ACTGACAACATGACCGACCTCC | p4Δfhbp |
| fhbp-ID2 | TGTTGAAGTCTCTGTCGTCGC | p4Δfhbp |
| fhbp-ID3 | AAGTCATAAGGGCGCACCTTC | p4Δfhbp |
| fhbp-ID4 | TGTAGCCAGCGATAAGGCTCTG | p4Δfhbp |
| fhbp-ID5 | AACAGCCAGGCTTATGGAAGG | p4Δfhbp |
| fhbp-ID6 | TATAGCTGTAGCGACACGAATACTATATCT | p4Δfhbp |
| fhbp-ID7 | AGATATAGTATTCGTGTCGCTACAGCTATA | p4Δfhbp |
| fhbp-ID8 | TGTCAAGCCAATCCATGTCTGG | p4Δfhbp |
| fhb-ID1 | TCGGTGCTATCTTGCGTAGTC | p4Δfhb |
| fhb-ID2 | CATCTGGTTCTAGCGATTCTGC | p4Δfhb |
| fhb-ID3 | TGATGCCAAAAGCAGAGGCAC | p4Δfhb |
| fhb-ID4 | TGGAACTTTCGAGGTCGGTG | p4Δfhb |
| fhb-ID5_EcoRI b | GGCGCGAATTCCAAAGTTCTTGCCAGATGCCAC | p4Δfhb |
| fhb-ID6 | CCAGCCTATTGCGCTCCCTAATACGACTGT | p4Δfhb |
| fhb-ID7 | ACAGTCGTATTAGGGAGCGCAATAGGCTGG | p4Δfhb |
| fhb-ID8_PstI b | GGCGCCTGCAGAAATTTCCGCCCCTGACACAC | p4Δfhb |
| pFHBP_F_PsI b | GCGCCTGCAGCACATCCGACCACCTGAATATC | pMXfhbp |
| pFHBP_R_PstI b | GGCGCCTGCAGGTTCTAAAAAGAGGCTGGGCG | pMXfhbp |
| cps-ID1 | CCAGCAAAGTATGGTGGTTTCG | p4Δcps2F |
| cps-ID2 | GCGCACCAACTTCTCTTAATGC | p4Δcps2F |
| cps-ID3 | CTTAGTCACTCCGAACTCACCG | p4Δcps2F |
| cps-ID4 | CCACGCCAGATTCAATGAGC | p4Δcps2F |
| cps-ID5 | AGACGGTCATGAATGGCTACG | p4Δcps2F |
| cps-ID6 | GAGGGAGGTGTAGACTTCTGCTCCAGCATG | p4Δcps2F |
| cps-ID7 | CATGCTGGAGCAGAAGTCTACACCTCCCTC | p4Δcps2F |
| cps-ID8 | CATCAGAATGATGCCAAACAGG | p4Δcps2F |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Roy, D.; Grenier, D.; Segura, M.; Mathieu-Denoncourt, A.; Gottschalk, M. Recruitment of Factor H to the Streptococcus suis Cell Surface is Multifactorial. Pathogens 2016, 5, 47. https://doi.org/10.3390/pathogens5030047
Roy D, Grenier D, Segura M, Mathieu-Denoncourt A, Gottschalk M. Recruitment of Factor H to the Streptococcus suis Cell Surface is Multifactorial. Pathogens. 2016; 5(3):47. https://doi.org/10.3390/pathogens5030047
Chicago/Turabian StyleRoy, David, Daniel Grenier, Mariela Segura, Annabelle Mathieu-Denoncourt, and Marcelo Gottschalk. 2016. "Recruitment of Factor H to the Streptococcus suis Cell Surface is Multifactorial" Pathogens 5, no. 3: 47. https://doi.org/10.3390/pathogens5030047