Effect of Early-Life Treatment of Piglets with Long-Acting Ceftiofur on Colonization of Streptococcus suis Serotype 7 and Elicitation of Specific Humoral Immunity in a Farm Dealing with Streptococcal Diseases

 , and
, and
Abstract
:1. Introduction
2. Results
2.1. Streptococcal Diseases on Farm Prior to the Beginning of the Study
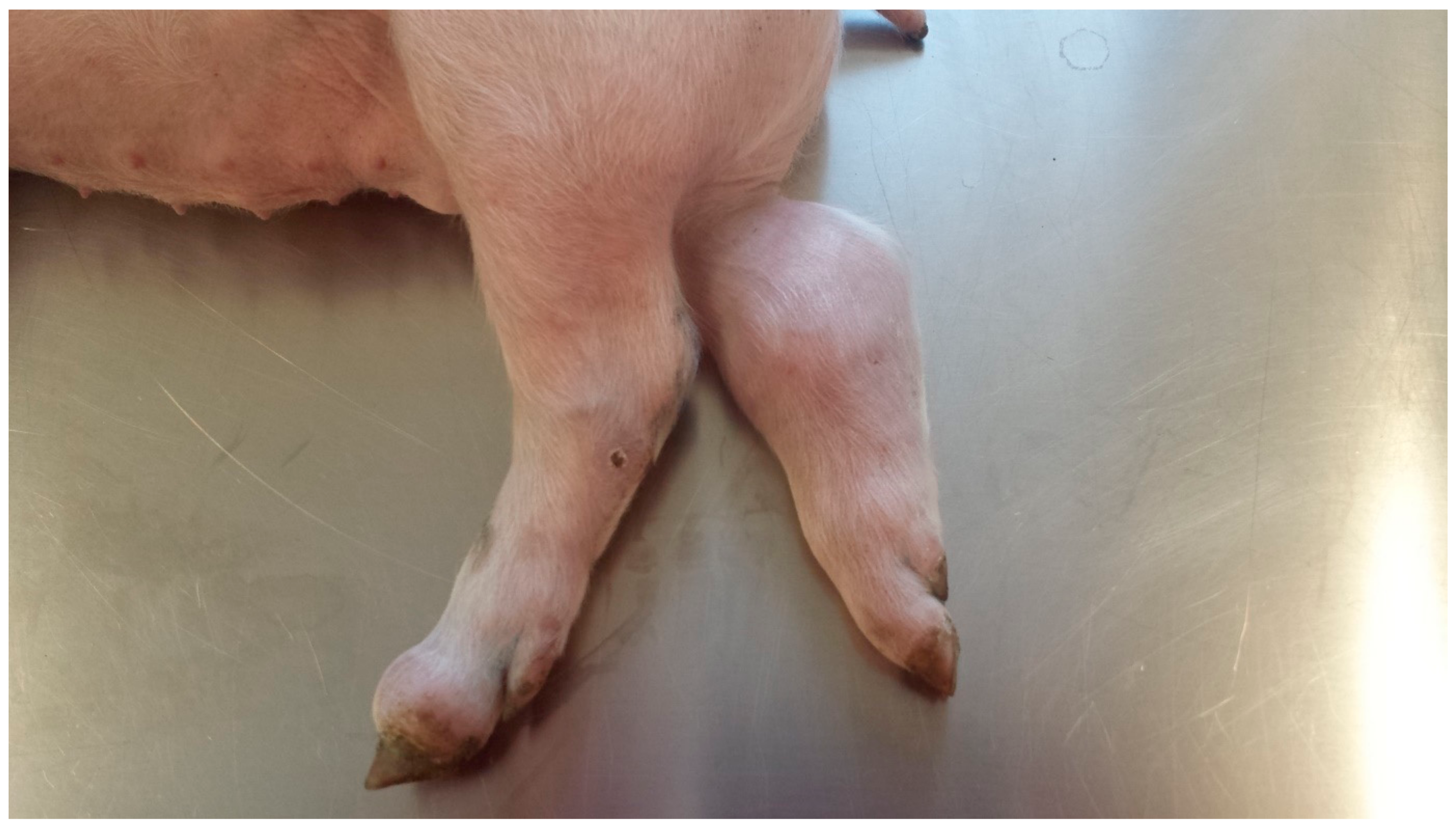
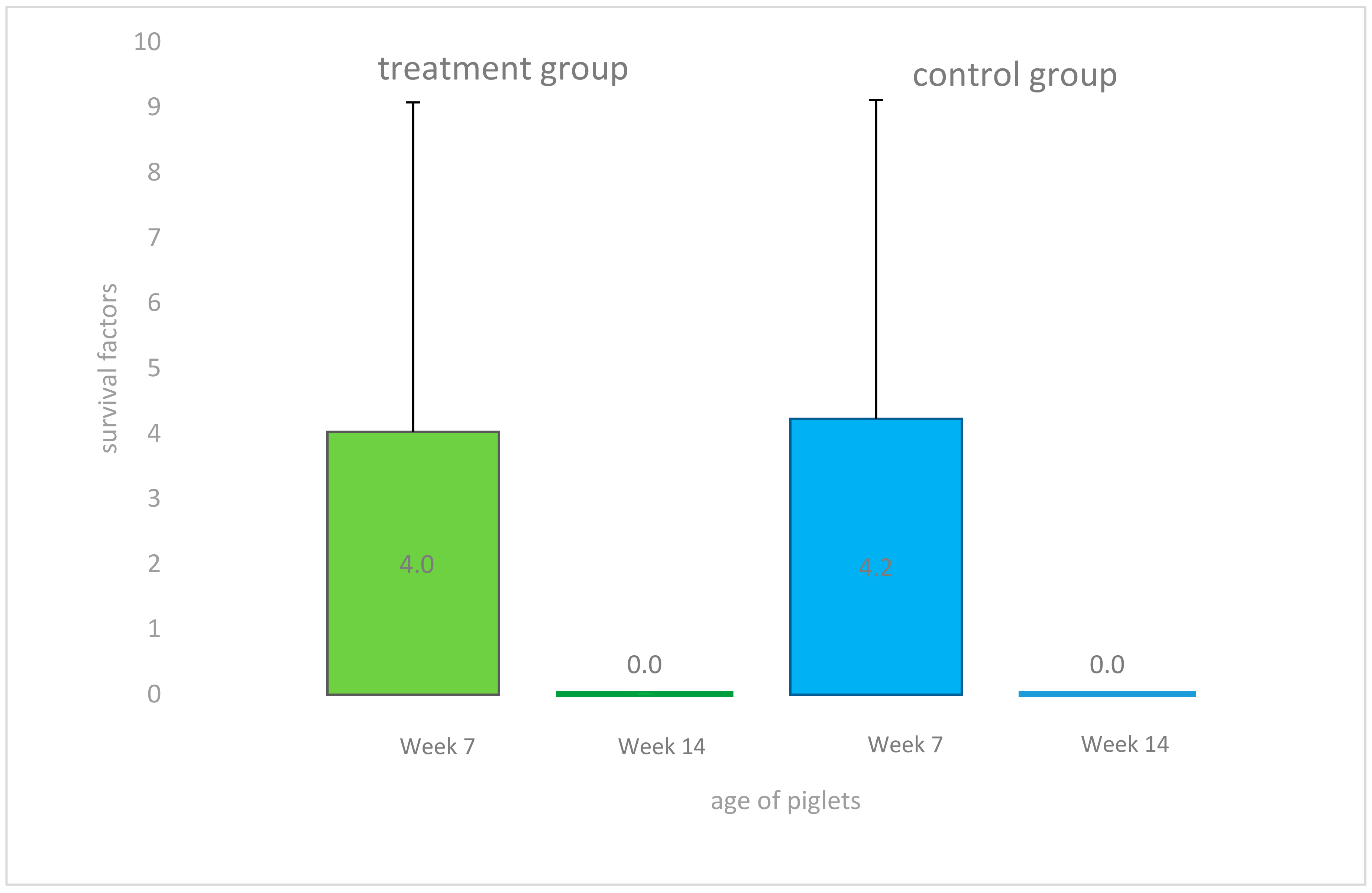
2.2. Clinical Findings, Body Weight Gain, Body Mass Index, and Ponderal Index during the Study Period
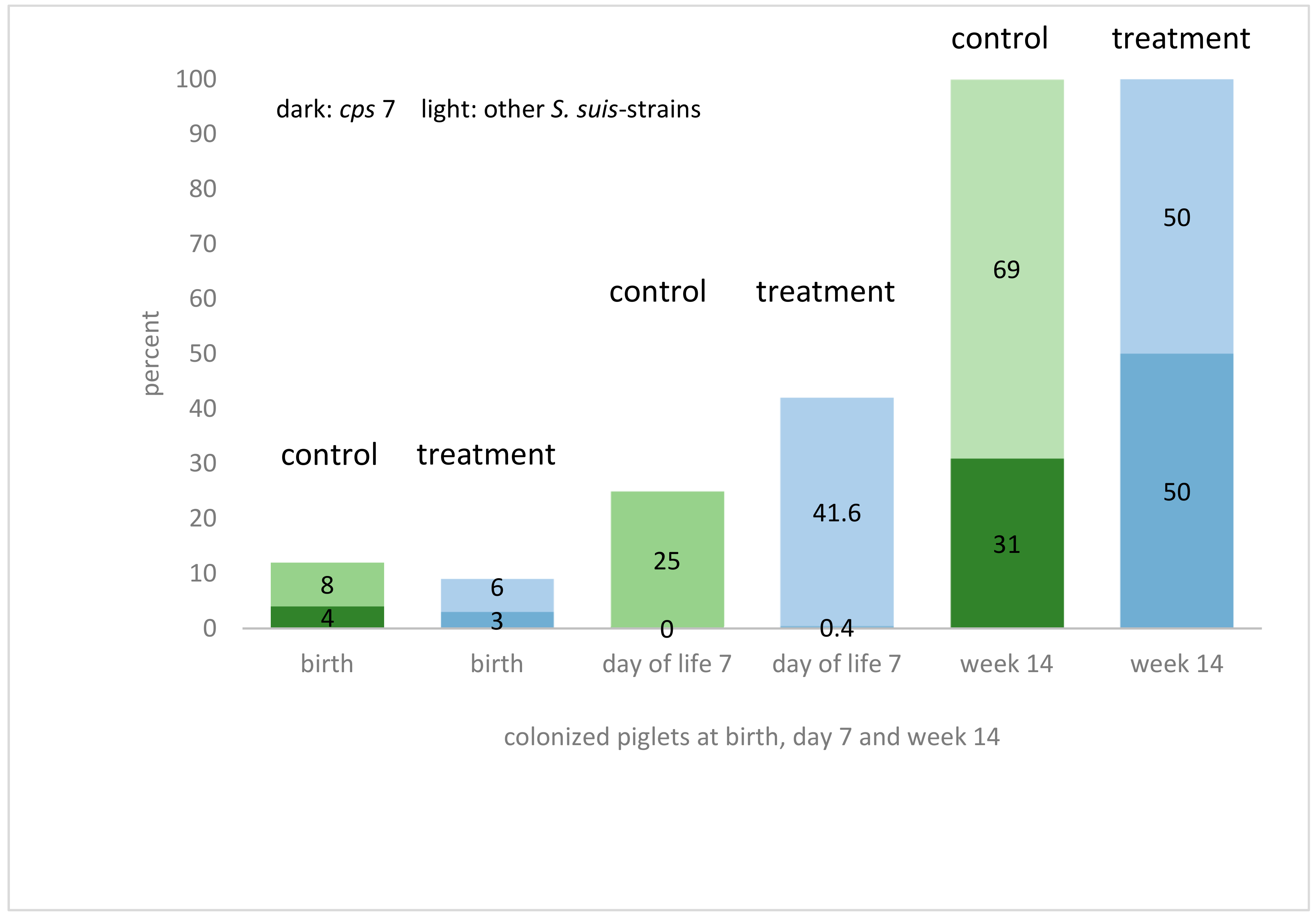
2.3. Prevalence of S. suis Colonization
2.4. Bactericidal Humoral Immune Response and Cytologic Findings
2.5. Antimicrobial Susceptibility Testing
3. Discussion
4. Material and Method
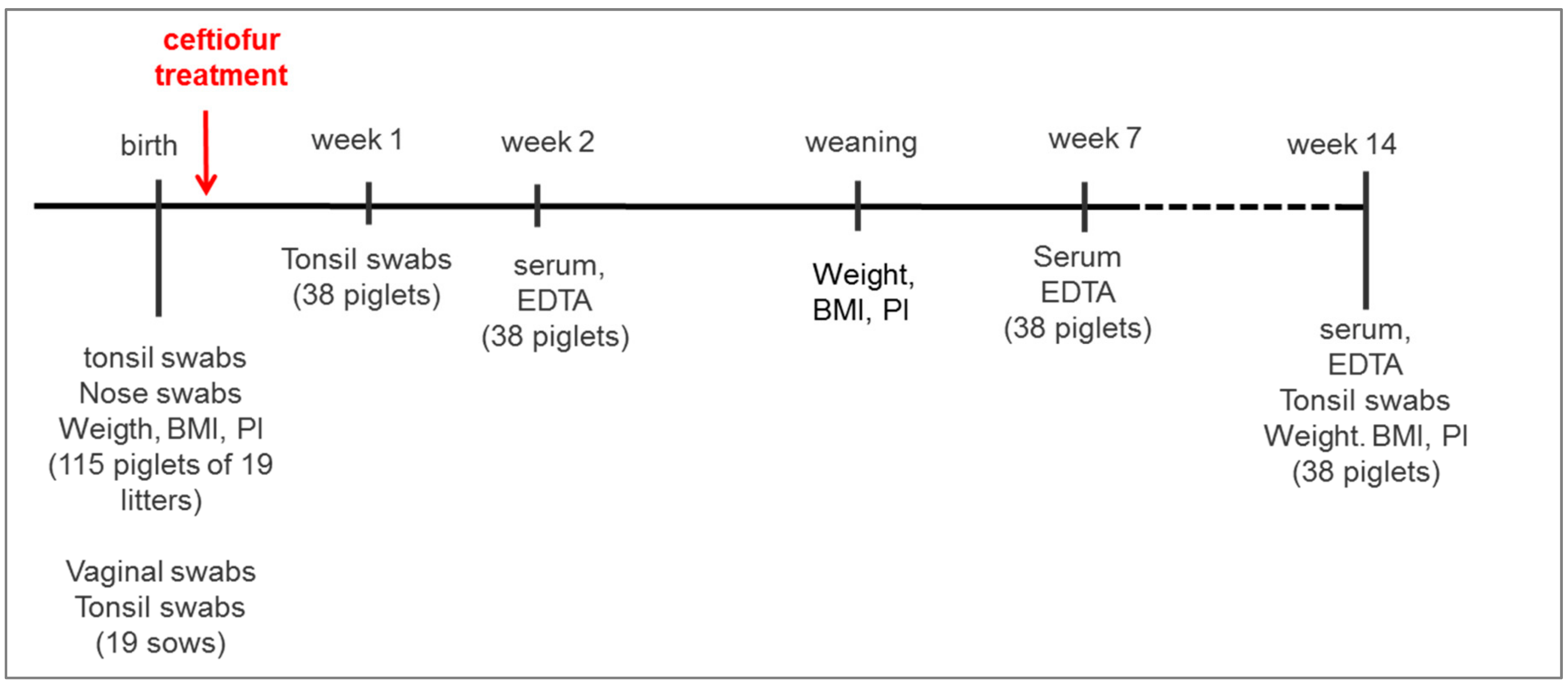
4.1. Study Farm, Animals, and the Design of the Study
4.2. Bacteriological Examination
4.3. Bactericidal Killing Assay
4.4. Statistical Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Jansen, E.J.; Van Dorssen, C.A. Meningoencephalitis bij varkens door streptococcen. Tijdschr. Diergeneeskd. 1951, 76, 815–832. [Google Scholar]
- Field, H.I.; Buntain, D.; Done, J.T. Studies on piglet mortality. I. Streptococcal meningitis and arthritis. Vet. Rec. 1954, 66, 453–455. [Google Scholar]
- Gottschalk, M. Streptococcosis. In Diseases of Swine, 10th ed.; Zimmerman, J., Karriker, L., Ramirez, A., Schwartz, K., Stevenson, G., Eds.; Wiley-Blackwell: West Sussex, UK, 2012; pp. 841–855. [Google Scholar]
- Staats, J.J.; Feder, I.; Okwumabua, O.; Chengappa, M.M. Streptococcus suis: Past and Present. Vet. Res. Commun. 1997, 21, 381–407. [Google Scholar] [CrossRef] [PubMed]
- Devriese, L.A.; Hommez, J.; Pot, B.; Haesebrouck, F. Identification and composition of the streptococcal and enterococcal flora of tonsils, intestines and faeces of pigs. J. Appl. Bacteriol. 1994, 77, 31–36. [Google Scholar] [CrossRef] [PubMed]
- MacInnes, J.I.; Gottschalk, M.; Lone, A.G.; Metcalf, D.S.; Ojha, S.; Rosendal, T.; Friendship, R.M. Prevalence of Actinobacillus pleuropneumoniae, Actinobacillus suis, Haemophilus parasuis, Pasteurella multocida, and Streptococcus suis in representative Ontario swine herds. Can. J. Vet. Res. 2008, 72, 242–248. [Google Scholar] [PubMed]
- Clifton-Hadley, F.A. Streptococcus suis type 2 infections. Br. Vet. J. 1983, 139, 1–5. [Google Scholar] [CrossRef]
- Sihyonen, L.; Kurl, D.N.; Salmela, J. Infection with Streptococcus suis serotypes 1 and 2 in the same diseased pig. Acta Vet. Scand. 1986, 27, 626–628. [Google Scholar]
- Brisebois, L.M.; Charlebois, R.; Higgins, R.; Nadeau, M. Prevalence of Streptococcus suis in four to eight week old clinically healthy piglets. Can. J. Vet. Res. 1990, 54, 174–177. [Google Scholar] [PubMed]
- Robertson, I.D.; Blackmore, D.K. Occupational exposure to Streptococcus suis type 2. Epidemiol. Infect. 1989, 103, 157–164. [Google Scholar] [CrossRef] [PubMed]
- Amass, S.F.; Clark, L.K.; Knox, K.E.; Wu, C.C.; Hill, M.A. Streptococcus suis colonization of piglets during parturition. Swine Health Prod. 1996, 4, 269–272. [Google Scholar]
- Torremorell, M.; Calsamiglia, M.; Pijoan, C. Colonization of suckling pigs by Streptococcus suis with particular reference to pathogenic serotype 2 strains. Can. J. Vet. Res. 1989, 62, 21–26. [Google Scholar]
- Cloutier, G.; D’Allaire, S.; Martinez, G.; Surprenant, C.; Lacouture, S.; Gottschalk, M. Epidemiology of Streptococcus suis serotype 5 infection in a pig herd with and without clinical disease. Vet. Microbiol. 2003, 97, 135–151. [Google Scholar] [CrossRef] [PubMed]
- Marois, C.; Le Devendec, L.; Gottschalk, M.; Kobisch, M. Detection and molecular typing of Streptococcus suis in tonsils from live pigs in France. Can. J. Vet. Res. 2007, 71, 14–22. [Google Scholar] [PubMed]
- Clifton-Hadley, F.A.; Alexander, T.J.; Enright, M.R.; Guise, J. Monitoring herds for Streptococcus suis type 2 by sampling tonsils of slaughter pigs. Vet. Rec. 1984, 115, 562–564. [Google Scholar] [CrossRef] [PubMed]
- Clifton-Hadley, F.A.; Alexander, T.J.; Upton, I.; Duffus, W.P. Further studies on the subclinical carrier state of Streptococcus suis type 2 in pigs. Vet. Rec. 1984, 114, 513–518. [Google Scholar] [CrossRef] [PubMed]
- Lamont, M.H.; Edwards, P.T.; Windsor, R.S. Streptococcal meningitis in pigs: Results of a five-year survey. Vet. Rec. 1980, 107, 467–469. [Google Scholar] [CrossRef] [PubMed]
- Chengappa, M.M.; Maddux, R.L.; Kadel, W.L.; Greer, S.C.; Herren, C.E. Streptococcus suis Infection in Pigs: Incidence and Experimental Reproduction of the Syndrome; American Association of Veterinary Laboratory Diagnosticians: Visalia, CA, USA, 1986; pp. 25–38. [Google Scholar]
- Gottschalk, M.; Segura, M. The pathogenesis of the meningitis caused by Streptococcus suis: The unresolved question. Vet. Microbiol. 2000, 76, 259–272. [Google Scholar] [CrossRef]
- Madsen, L.W.; Bak, H.; Nielsen, B.; Jensen, H.E.; Aalbaek, B.; Riising, H.J. Bacterial colonization and invasion in pigs experimentally exposed to Streptococcus suis serotype 2 in aerosol. Zoonoses Public Health 2002, 49, 211–215. [Google Scholar] [CrossRef]
- Beineke, A.; Bennecke, K.; Neis, C.; Schröder, C.; Waldmann, K.H.; Baumgärtner, W.; Valentin-Weigand, P.; Baums, C.G. Comparative evaluation of virulence and pathology of Streptococcus suis serotypes 2 and 9 in experimentally infected growers. Vet. Microbiol. 2008, 128, 423–430. [Google Scholar] [CrossRef] [PubMed]
- Unterweger, C.; Baums, C.G.; Höcher, M.; Fischer, L.; Weiss, A.; Hennig-Pauka, I. Clinics, diagnosis and prophylaxis of a Streptococcus suis serotype 7 farm problem. Berl. Münchener Tierarztliche Wochenschr. 2014, 127, 194–201. [Google Scholar]
- Wisselink, H.J.; Smith, H.E.; Stockhofe-Zurwieden, N.; Peperkamp, K.; Vecht, U. Distribution of capsular types and production of muramidase-released protein (MRP) and extracellular factor (EF) of Streptococcus suis strains isolated from diseased pigs in seven European countries. Vet. Microbiol. 2000, 74, 237–248. [Google Scholar] [CrossRef]
- Silva, L.M.; Baums, C.G.; Rehm, T.; Wisselink, H.J.; Goethe, R.; Valentin-Weigand, P. Virulence-associated gene profiling of Streptococcus suis isolates by PCR. Vet. Microbiol. 2006, 115, 117–127. [Google Scholar] [CrossRef] [PubMed]
- Mogollon, J.D.; Pijoan, C.; Murtaugh, M.P.; Collins, J.E.; Cleary, P.P. Identification of epidemic strains of Streptococcus suis by genomic fingerprinting. J. Clin. Microbiol. 1991, 29, 782–787. [Google Scholar] [PubMed]
- Aarestrup, F.M.; Rasmussen, S.R.; Artursson, K.; Jensen, N.E. Trends in the resistance to antimicrobial agents of Streptococcus suis isolates from Denmark and Sweden. Vet. Microbiol. 1998, 63, 71–80. [Google Scholar] [CrossRef]
- Tian, Y.; Aarestrup, F.M.; Lu, C.P. Characterization of Streptococcus suis serotype 7 isolates from diseased pigs in Denmark. Vet. Microbiol. 2004, 103, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Torremorell, M.; Pijoan, C.; Trigo, E. Vaccination against Streptococcus suis: Effect on nursery mortality. J. Swine Health Prod. 1997, 4, 139–143. [Google Scholar]
- Callens, B.; Persoons, D.; Maes, D.; Laanen, M.; Postma, M.; Boyen, F.; Haesebrouck, F.; Butaye, P.; Catry, B.; Dewulf, J. Prophylactic and metaphylactic antimicrobial use in Belgian fattening pig herds. Prev. Vet. Med. 2012, 106, 53–62. [Google Scholar] [CrossRef] [PubMed]
- Tantituvanont, A.; Yimprasert, W.; Werawatganone, P.; Nilubo, D. Pharmacokinetics of ceftiofur hydrochloride in pigs infected with porcine reproductive and respiratory syndrome virus. J. Antimicrob. Chemother. 2008, 63, 369–373. [Google Scholar] [CrossRef] [PubMed]
- Hornish, R.E.; Kotarski, S.F. Cephalosporins in veterinary medicine—Ceftiofur use in food animals. Curr. Top. Med. Chem. 2002, 2, 717–731. [Google Scholar] [CrossRef] [PubMed]
- Collignon, P.; Powers, J.H.; Chiller, T.M.; Aidara-Kane, A.; Aarestrup, F.M. World Health Organization ranking of antimicrobials according to their importance in human medicine: A critical step for developing risk management strategies for the use of antimicrobials in food production animals. Clin. Infect. Dis. 2009, 49, 132–141. [Google Scholar] [CrossRef] [PubMed]
- Gascho, D.; Long, D.; Holman, L.; Wolff, T. Effect of pre-farrow ceftiofur sodium (Naxcel®) administration on Streptococcus suis colonization of peri-parturient females and their litters. In Proceedings of the AASV Annual Meeting, New Orleans, LA, USA, 27 February–1 March 2016; pp. 247–248. [Google Scholar]
- Seele, J.; Hillermann, L.M.; Beineke, A.; Seitz, M.; von Pawel-Rammingen, U.; Valentin-Weigand, P.; Baums, C.G. The immunoglobulin M-degrading enzyme of Streptococcus suis, IdeSsuis, is a highly protective antigen against serotype 2. Vaccine 2015, 33, 2207–2212. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Fu, Y.; Ma, C.; He, Y.; Yu, Y.; Du, D.; Yao, H.; Lu, C.; Zhang, W. The non-conserved region of MRP is involved in the virulence of Streptococcus suis serotype 2. Virulence 2017, 8, 1274–1289. [Google Scholar] [CrossRef] [PubMed]
- Clinical and Laboratory Standard Institute (CLSI). Performance Standards for Antimicrobial Disk and Dilution Susceptibility Tests for Bacteria isolated from Animals; Approved Standard; 3rd CLSI Supplement VET01S Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2015; Available online: http://vet01s.edaptivedocs.info/ Login.aspx (accessed on 30 June 2015).
- Flores, J.L.M.; Higgins, R.; D’Allaire, S.; Charette, R.; Boudreau, M.; Gottschalk, M. Distribution of the different capsular types of Streptococcus suis in nineteen swine nurseries. Can. Vet. J. 1993, 34, 170–171. [Google Scholar] [PubMed]
- Pijoan, C. Diseases of high health pigs: Some ideas on pathogenesis. In Proceedings of the Allen D. Leman Swine Conference, St. Paul, MN, USA, 23–24 February 1995; Volume 22, pp. 16–17. [Google Scholar]
- Van der Velden, E.W.; Raemakers, S.G.; Vecht, U.; Cromwijk, W.A. Streptococcus suis type 2 bij varkens. Eeen glimporteerd problem? Tijdschr. Diergeneeskd. 1987, 112, 660–664. [Google Scholar] [PubMed]
- Dee, S.A.; Carlson, A.R.; Winkelman, N.L.; Corey, M.M. Effect of management practices on the Streptococcus suis carrier rate in nursery swine. J. Am. Vet. Med. Assoc. 1993, 203, 295–299. [Google Scholar] [PubMed]
- Van Leengoed, L.A.M.; Vecht, U.; Verheyen, E.R.M. Streptococcus type 2 infections in pigs in the Netherlands (part two). Vet. Q. 1987, 9, 111–117. [Google Scholar] [CrossRef] [PubMed]
- Clifton-Hadley, F.A.; Alexander, T.; Enright, M.R. The epidemiology, diagnosis, treatment and control of Streptococcus suis Type-2 infection. In Proceedings of the AASV Annual Meeting, Minneapolis, MN, USA, 16–18 March 1986; pp. 473–491. [Google Scholar]
- Clifton-Hadley, F.A.; Alexander, T.J.L. The carrier site and carrier rate of Streptococcus suis type II in pigs. Vet. Rec. 1980, 107, 40–41. [Google Scholar] [CrossRef] [PubMed]
- Baums, C.G.; (Faculty of Veterinary Medicine, Leipzig, Germany). Personal communication, 2018.
- Lapointe, L.; D´Allaire, S.; Lebrun, A.; Lacouture, S.; Gottschalk, M. Antibody response to an autogenous vaccine and serologic profile for Streptococcus suis capsular type 1/2. Can. J. Vet. Res. 2002, 66, 8–14. [Google Scholar] [PubMed]
- Baums, C.G.; Kock, C.; Beineke, A.; Bennecke, K.; Goethe, R.; Schroder, C.; Waldmann, K.H.; Valentin-Weigand, P. Streptococcus suis bacterin and subunit vaccine immunogenicities and protective efficacies against serotypes 2 and 9. Clin. Vaccine Immunol. 2009, 16, 200–208. [Google Scholar] [CrossRef] [PubMed]
- Van Rennings, L.; von Münchhausen, C.; Ottilie, H.; Hartmann, M.; Merle, R.; Honscha, W.; Käsbohrer, A.; Kreienbrock, L. Cross sectional study on antibiotic usage in pigs in Germany. PLoS ONE 2015, 10, e0119114. [Google Scholar] [CrossRef]
- Trauffler, M.; Griesbacher, A.; Fuchs, K.; Köfer, J. Antimicrobial drug use in Austrian pig farms: Plausibility check of electronic on-farm records and estimation of consumption. Vet. Rec. 2014, 175, 402–409. [Google Scholar] [CrossRef] [PubMed]
- Felman, R.H.; Sutherland, D.B.; Conklin, J.L.; Mitros, F.A. Eosinophilic cholecystitis, appendiceal inflammation, pericarditis and cephalosporin-associated eosinophilia. Dig. Dis. Sci. 1994, 39, 418–422. [Google Scholar] [CrossRef] [PubMed]
- Reams, R.Y.; Glickman, L.T.; Harrington, D.D.; Bowersock, T.L.; Thacker, H.L. Streptococcus suis infection in swine: A retrospective study of 256 cases. Part I. Epidemiologic factors and antibiotic susceptibility patterns. J. Vet. Diagn. Investig. 1993, 5, 363–367. [Google Scholar] [CrossRef] [PubMed]
- Gottschalk, M.; Higgins, R.; Jacques, M.; Mittal, K.R.; Henrichsen, J. Description of 14 new capsular types of Streptococcus suis. J. Clin. Microbiol. 1989, 27, 2633–2636. [Google Scholar] [PubMed]
- Varela, N.P.; Gadbois, P.; Thibault, C.; Gottschalk, M.; Dick, P.; Wilson, J. Antimicrobial Resistance and Prudent Drug Use for Streptococcus Suis. Anim. Health Res. Rev. 2013, 14, 68–77. [Google Scholar] [CrossRef] [PubMed]
- Vela, A.I.; Moreno, M.A.; Cebolla, J.A.; Gonzales, S.; Latre, M.V.; Dominguez, L.; Fernandez-Garayzabal, J.F. Antimicrobial susceptibility of clinical strains of Streptococcus suis isolated in Spain. Vet. Microbiol. 2005, 105, 143–147. [Google Scholar] [CrossRef] [PubMed]
- Princivalli, M.S.; Palmieri, C.; Magi, G.; Vignaroli, C.; Manzin, A.; Camporese, A.; Barocci, S.; Magistrali, C.; Facinelli, B. Genetic diversity of Streptococcus suis clinical isolates from pigs and humans in Italy (2003–2007). Euro Surveill. 2009, 14, 1–7. [Google Scholar] [CrossRef]
- Van Hout, J.; Heuvelink, A.; Gonggrijp, M. Monitoring of antimicrobial susceptibility of Streptococcus suis in The Netherlands, 2013–2015. Vet. Microbiol. 2016, 194, 5–10. [Google Scholar] [CrossRef] [PubMed]
- Baxter, E.M.; Jarvis, S.; D’Eath, R.B.; Ross, D.W.; Robson, S.K.; Farish, M.; Nevison, I.M.; Lawrence, A.B.; Edwards, S.A. Investigating the behavioural and physiological indicators of neonatal survival in pigs. Theriogenology 2008, 69, 773–783. [Google Scholar] [CrossRef] [PubMed]
- Clinical and Laboratory Standard Institute (CLSI). Performance Standards for Antimicrobial Disk and Dilution Susceptibility Tests for Bacteria Isolated from Animals; Approved Standard; CLSI Document M31-A3 Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2008. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Numbers of Isolated Strains | S. suis Serotype | epf | mrp | arcA | sly | Location of Isolation |
|---|---|---|---|---|---|---|---|
| 1 | 3 | 7 | − | + | + | − | meninges, cerebrospinal fluids |
| 2 | 2 | ? (not 1,2,7,9) | − | + | + | + | meninges, cerebrospinal fluids |
| 3 | 2 | ? (not 1,2,7,9) | − | + | + | − | meninges, joints |
| Group | Weaning | Week 14 | p | ||
|---|---|---|---|---|---|
| BMI | PI | BMI | PI | ||
| Control | 30.1 | 62.9 | 49.1 | 84.2 | NS |
| treatment | 30.4 | 64.7 | 49.3 | 84.4 | NS |
| Group | Time of Measure | ery | Hct | hb | leu | neu | eos | eos | bas | mono | lym |
|---|---|---|---|---|---|---|---|---|---|---|---|
| ×1012/L | % | g/dL | ×109/L | ×109/L | ×109/L | % | ×109/L | ×109/L | ×109/L | ||
| T | A | 6.132 | 34.279 | 10.229 | 13.495 | 4.986 | 0.163 | 1.217 | 0.008 | 0.580 | 7.757 |
| B | 6.988 | 37.738 | 11.557 | 30.643 | 9.543 | 0.125 * | 0.662 ** | 0.015 | 0.579 | 10.066 | |
| C | A | 6.172 | 33.300 | 9.963 | 14.583 | 5.727 | 0.138 | 1.041 | 0.008 | 0.598 | 8.111 |
| B | 6.962 | 38.400 | 11.711 | 24.075 | 9.263 | 0.147 * | 0.447 ** | 0.043 | 0.875 | 13.279 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Unterweger, C.; Ruczizka, U.; Spergser, J.; Baums, C.G.; Hennig-Pauka, I. Effect of Early-Life Treatment of Piglets with Long-Acting Ceftiofur on Colonization of Streptococcus suis Serotype 7 and Elicitation of Specific Humoral Immunity in a Farm Dealing with Streptococcal Diseases. Pathogens 2018, 7, 34. https://doi.org/10.3390/pathogens7020034
Unterweger C, Ruczizka U, Spergser J, Baums CG, Hennig-Pauka I. Effect of Early-Life Treatment of Piglets with Long-Acting Ceftiofur on Colonization of Streptococcus suis Serotype 7 and Elicitation of Specific Humoral Immunity in a Farm Dealing with Streptococcal Diseases. Pathogens. 2018; 7(2):34. https://doi.org/10.3390/pathogens7020034
Chicago/Turabian StyleUnterweger, Christine, Ursula Ruczizka, Joachim Spergser, Christoph Georg Baums, and Isabel Hennig-Pauka. 2018. "Effect of Early-Life Treatment of Piglets with Long-Acting Ceftiofur on Colonization of Streptococcus suis Serotype 7 and Elicitation of Specific Humoral Immunity in a Farm Dealing with Streptococcal Diseases" Pathogens 7, no. 2: 34. https://doi.org/10.3390/pathogens7020034