Identification and In Silico Characterization of a Genetically Distinct Avian Rotavirus D Capsid Gene, VP7



 ,
,  and
and 
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling and Amplification of RV-D Capsid Gene
2.2. Cloning, Sequencing and Sequence Analysis
2.3. Phyloanalysis and In Silico Motifs/Domain Predictions of Capsid Gene VP7
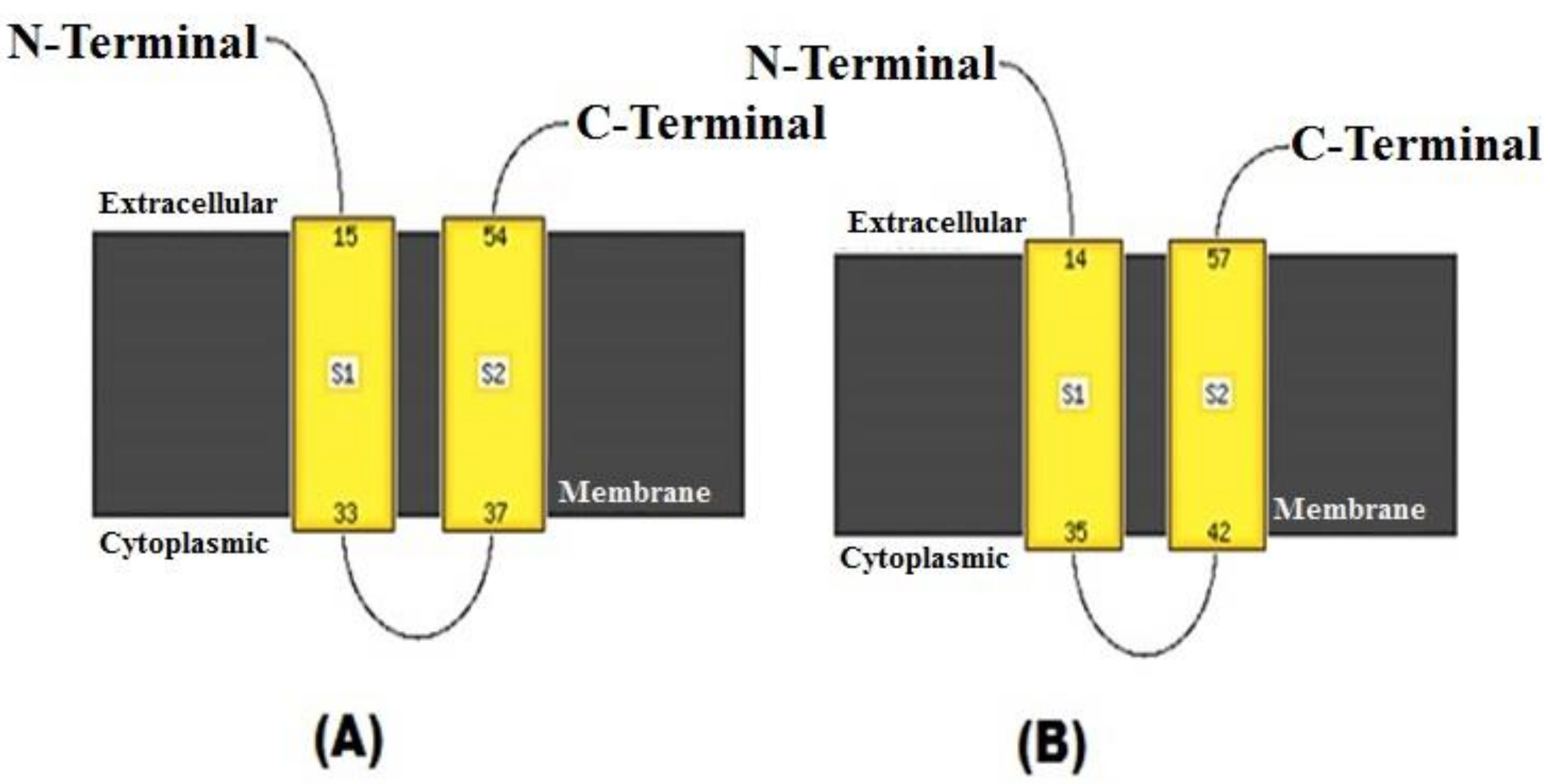
3. Results and Discussion
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Villarreal, L.Y.B.; Uliana, G.; Valenzuela, C.; Chácon, J.L.V.; Saindenberg, A.B.S.; Sanches, A.A.; Brandão, P.E.; Jerez, J.A.; Ferreira, A.J.P. Rotavirus detection and isolation from chickens with or without symptoms. Braz. J. Poult. Sci. 2006, 3, 187–191. [Google Scholar] [CrossRef]
- Dhama, K.; Saminathan, M.; Karthik, K.; Tiwari, R.; Shabbir, M.Z.; Kumar, N.; Malik, Y.S.; Singh, R.K. Avian rotavirus enteritis—An updated review. Vet. Q. 2015, 3, 142–158. [Google Scholar] [CrossRef] [PubMed]
- Otto, P.H.; Liebler-Tenorio, E.M.; Elschner, M.; Reetz, J.; Lohren, U.; Dille, R. Detection of rotaviruses and intestinal lesions in broiler chicks from flocks with runting and stunting syndrome (RSS). Avian Dis. 2006, 50, 411–418. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.; Crawford, S.E.; Czako, R.; Cortes-Penfield, N.W.; Smith, D.F.; LePendu, J.; Estes, M.K.; Prasad, B.V. Cell attachment protein VP8* of a human rotavirus specifically interacts with A-type histo-blood group antigen. Nature 2012, 7397, 256–259. [Google Scholar] [CrossRef] [PubMed]
- Estes, M.; Kapikian, A. Rotaviruses. In Fields Virology, 5th ed.; Knipe, D.M., Howley, P.M., Griffin, D.E., Lamb, R.A., Martin, M.A., Roizman, B., Straus, S.E., Eds.; Kluwer/Lippincott Williams and Wilkins: Philadelphia, PA, USA, 2007; pp. 1917–1974. [Google Scholar]
- Kattoor, J.J.; Malik, Y.S.; Sharma, K.; Kumar, N.; Batra, M.; Jindal, N.; Yadav, A.S. Molecular evidence of group D rotavirus in commercial broiler chicks in India. Avian Biol. Res. 2013, 6, 313–316. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed]
- TMHMM Server v.2.0. In DTU Bioinformatics. Department of Bio and Health Informatics. 2018. Available online: http://www.cbs.dtu.dk/services/TMHMM/ (accessed on 6 February 2018).
- Phyre2. In Protein Homology/Analogy Recognition Engine. 2018. Available online: http://www.sbg.bio.ic.ac.uk/phyre2/ (accessed on 6 February 2018).
- Trojnar, E.; Otto, P.; Roth, B.; Reetz, J.; Johne, R. The genome segments of a group D rotavirus possess group A-like conserved termini but encode group-specific proteins. J. Virol. 2010, 84, 10254–10265. [Google Scholar] [CrossRef] [PubMed]
- Mirazimi, A.; Svensson, L. Carbohydrates facilitate correct disulfide bond formation and folding of rotavirus VP7. J. Virol. 1998, 72, 3887–3892. [Google Scholar] [PubMed]
- Ciarlet, M.; Hidalgo, M.; Gorziglia, M.; Liprandi, F. Characterization of neutralization epitopes on the VP7 surface protein of serotype G11 porcine rotaviruses. J. Gen. Virol. 1994, 75, 1867–1873. [Google Scholar] [CrossRef] [PubMed]
- Kattoor, J.J.; Malik, Y.S.; Sasidharan, A.; Rajan, V.M.; Dhama, K.; Ghosh, S.; Bányai, K.; Kobayashi, N.; Singh, R.K. Analysis of codon usage pattern evolution in avian rotaviruses and their preferred host. Infect. Genet. Evol. 2015, 34, 17–25. [Google Scholar] [CrossRef] [PubMed]
- Maass, D.R.; Atkinson, P.H. Retention by the endoplasmic reticulum of rotavirus VP7 is controlled by three adjacent amino-terminal residues. J. Virol. 1994, 68, 366–378. [Google Scholar] [PubMed]



© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Deol, P.; Kattoor, J.J.; Sircar, S.; Batra, M.; Dhama, K.; Malik, Y.S. Identification and In Silico Characterization of a Genetically Distinct Avian Rotavirus D Capsid Gene, VP7. Pathogens 2018, 7, 37. https://doi.org/10.3390/pathogens7020037
Deol P, Kattoor JJ, Sircar S, Batra M, Dhama K, Malik YS. Identification and In Silico Characterization of a Genetically Distinct Avian Rotavirus D Capsid Gene, VP7. Pathogens. 2018; 7(2):37. https://doi.org/10.3390/pathogens7020037
Chicago/Turabian StyleDeol, Pallavi, Jobin Jose Kattoor, Shubhankar Sircar, Munish Batra, Kuldeep Dhama, and Yashpal Singh Malik. 2018. "Identification and In Silico Characterization of a Genetically Distinct Avian Rotavirus D Capsid Gene, VP7" Pathogens 7, no. 2: 37. https://doi.org/10.3390/pathogens7020037
APA StyleDeol, P., Kattoor, J. J., Sircar, S., Batra, M., Dhama, K., & Malik, Y. S. (2018). Identification and In Silico Characterization of a Genetically Distinct Avian Rotavirus D Capsid Gene, VP7. Pathogens, 7(2), 37. https://doi.org/10.3390/pathogens7020037