Fetal Hepatic Response to Bovine Viral Diarrhea Virus Infection in Utero

 ,
, 
Abstract
1. Introduction
2. Results
2.1. Detection of Bovine Viral Diarrhea Virus
2.2. Kupffer Cell Characterization
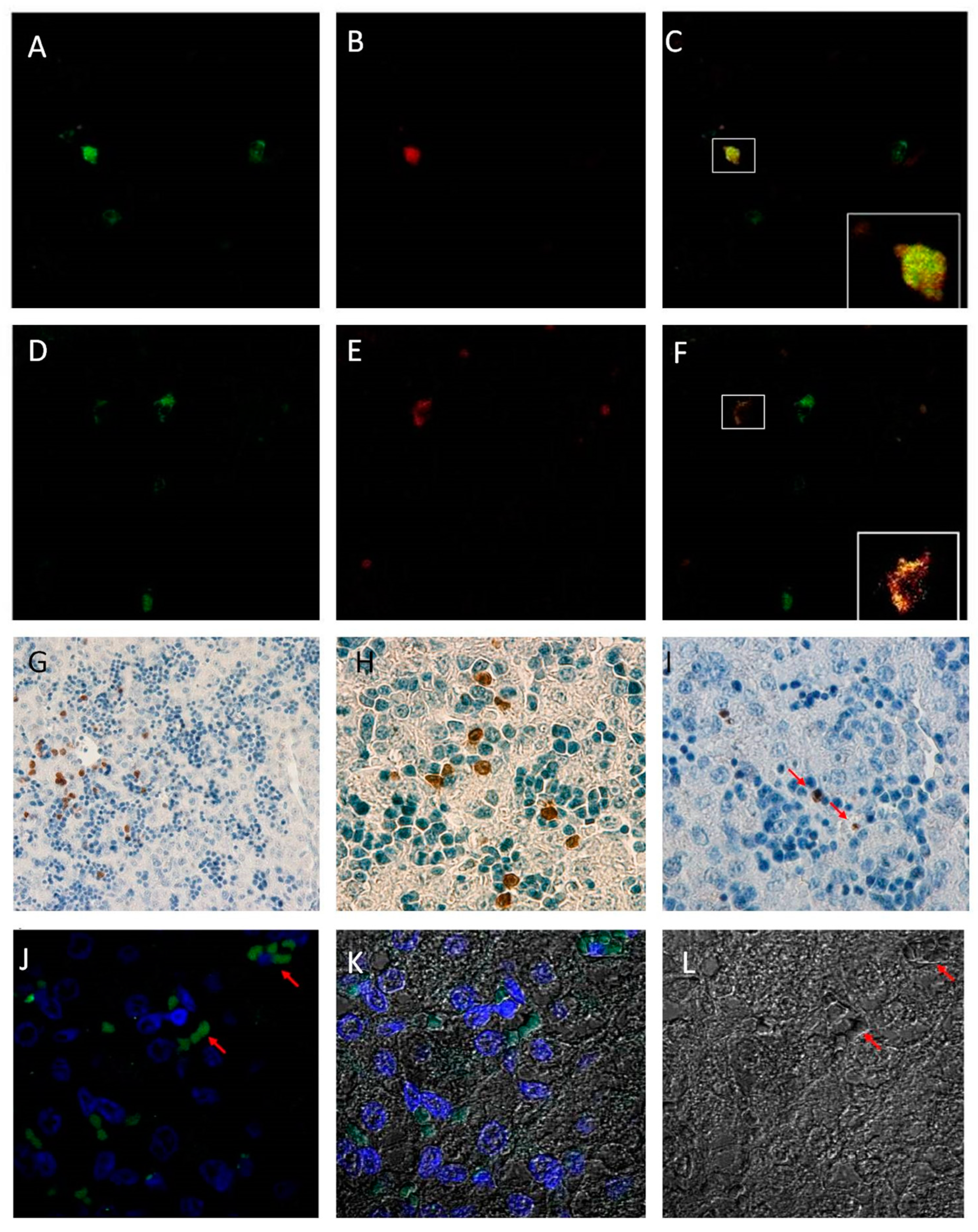
2.3. Immunohistochemistry and Immunofluorescence
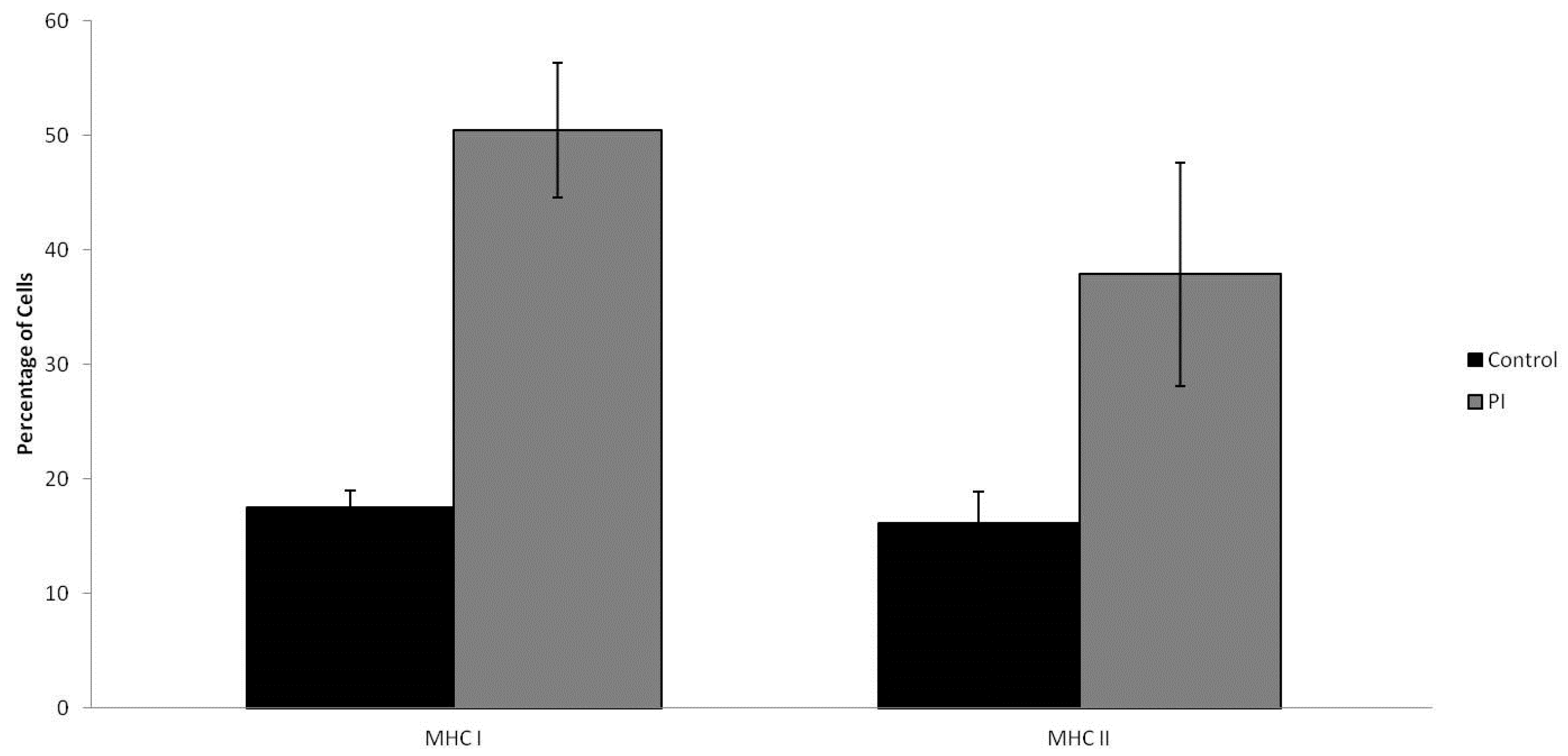
2.4. MHC I/MHC II Expression
3. Discussion
4. Methods
4.1. Animals and Sample Collection
4.2. Kupffer Cell Isolation
4.3. Phagocytic Activity Evaluation
4.4. Flow Cytometry
4.5. Immunohistochemistry
4.6. Immunofluorescence
4.7. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Houe, H. Epidemiological features and economical importance of bovine virus diarrhoea virus (BVDV) infections. Vet. Microbiol. 1999, 64, 89–107. [Google Scholar] [CrossRef]
- Gunn, G.J.; Stott, A.W.; Humphry, R.W. Modelling and costing BVD outbreaks in beef herds. Vet. J. 2004, 167, 143–149. [Google Scholar] [CrossRef]
- Baker, J.C. The clinical manifestations of bovine viral diarrhea infection. Vet. Clin. N. Am. Food Anim. Pract. 1995, 11, 425–445. [Google Scholar] [CrossRef]
- Neill, J.D.; Ridpath, J.F. Increase in proto-oncogene mRNA transcript levels in bovine lymphoid cells infected with a cytopathic type 2 bovine viral diarrhea virus. Virus Res. 2008, 135, 326–331. [Google Scholar] [CrossRef] [PubMed]
- Ohmann, H.B.; Jensen, M.H.; Sørensen, K.J.; Dalsgaard, K. Experimental fetal infection with bovine viral diarrhea virus, I. Virological and serological studies. Can. J. Comp. Med. 1982, 46, 357. [Google Scholar] [PubMed]
- Grooms, D.L. Reproductive consequences of infection with bovine viral diarrhea virus. Vet. Clin. N. Am. Food Anim. Pract. 2004, 20, 5–19. [Google Scholar] [CrossRef] [PubMed]
- Chase, C.C.L.; Elmowalid, G.; Yousif, A.A.A. The immune response to bovine viral diarrhea virus: A constantly changing picture. Vet. Clin. N. Am. Food Anim. Pract. 2004, 20, 95–114. [Google Scholar] [CrossRef] [PubMed]
- Peterhans, E.; Jungi, T.W.; Schweizer, M. How the bovine viral diarrhea virus outwits the immune system. DTW Deutsch. Tierarztliche Wochenschr. 2006, 113, 124–129. [Google Scholar]
- Fredriksen, B.; Press, C.M.; Sandvik, T.; Ødegaard, S.A.; Løken, T. Detection of viral antigen in placenta and fetus of cattle acutely infected with bovine viral diarrhea virus. Vet. Pathol. 1999, 36, 267–275. [Google Scholar] [CrossRef] [PubMed]
- Scherer, C.F.; Flores, E.F.; Weiblen, R.; Caron, L.; Irigoyen, L.F.; Neves, J.P.; Maciel, M.N. Experimental infection of pregnant ewes with bovine viral diarrhea virus type-2 (BVDV-2): Effects on the pregnancy and fetus. Vet. Pathol. 2001, 79, 285–299. [Google Scholar] [CrossRef]
- Swasdipan, S.; Bielefeldt-Ohmann, H.; Phillips, N.; Kirkland, P.D.; McGowan, M.R. Rapid transplacental infection with bovine pestivirus following intranasal inoculation of ewes in early pregnancy. Vet. Pathol. 2001, 38, 275–280. [Google Scholar] [CrossRef] [PubMed]
- Swasdipan, S.; McGowan, M.; Phillips, N.; Bielefeldt-Ohmann, H. Pathogenesis of transplacental virus infection: Pestivirus replication in the placenta and fetus following respiratory infection. Microb. Pathog. 2002, 32, 49–60. [Google Scholar] [CrossRef] [PubMed]
- Holt, P.G.; Jones, C.A. The development of the immune system during pregnancy and early life. Allergy 2000, 55, 688–697. [Google Scholar] [CrossRef] [PubMed]
- Osburn, B.I. Ontogeny of immune responses in cattle. In The Ruminant Immune System in Health and Disease; Morrison, W.I., Ed.; Cambridge University Press: Cambridge, UK, 1986; Chapter 14; pp. 252–260. [Google Scholar]
- Robinson, M.W.; Harmon, C.; O’Farrelly, C. Liver immunology and its role in inflammation and homeostasis. Cell. Mol. Immunol. 2016, 13, 267–276. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhivaki, D.; Lo-Man, R. Unique aspects of the perinatal immune system. NRI 2017, 17, 495–507. [Google Scholar] [CrossRef] [PubMed]
- Bilzer, M.; Roggel, F.; Gerbes, A.L. Role of Kupffer cells in host defense and liver disease. Liver Int. 2006, 26, 1175–1186. [Google Scholar] [CrossRef] [PubMed]
- Ginhoux, F.; Guilliams, M. Tissue-resident macrophage ontogeny and homeostasis. Immunity 2016, 44, 439–449. [Google Scholar] [CrossRef] [PubMed]
- Fox, E.S.; Thomas, P.E.; Broitman, S.A. Comparative studies of endotoxin uptake by isolated rat Kupffer and peritoneal cells. Infect. Immun. 1987, 55, 2962–2966. Available online: http://iai.asm.org/content/55/12/2962.full.pdf+html (accessed on 30 May 2018). [PubMed]
- Risalde, M.A.; Gómez-Villamandos, J.C.; Pedrera, M.; Molina, V.; Cerón, J.J.; Martinez-Subiela, S.; Sánchez-Cordón, P.J. Hepatic immune response in calves during acute subclinical infection with bovine viral diarrhoea virus type 1. Vet. J. 2011, 190, e110–e116. [Google Scholar] [CrossRef] [PubMed]
- Knoll, P.; Schlaak, J.; Uhrig, A.; Kempf, P.; zum Büschenfelde, K.H.; Gerken, G. Human Kupffer cells secrete IL-10 in response to lipopolysaccharide (LPS) challenge. J. Hepatol. 1995, 22, 226–229. [Google Scholar] [CrossRef]
- Knolle, P.A.; Gerken, G. Local control of the immune response in the liver. Immunol. Rev. 2000, 174, 21–34. [Google Scholar] [CrossRef] [PubMed]
- Cantor, H.M.; Dumont, A.E. Hepatic suppression of sensitization to antigen absorbed into the portal system. Nature 1967, 215, 744–745. [Google Scholar] [CrossRef] [PubMed]
- Triger, D.R.; Cynamon, M.H.; Wright, R. Studies on hepatic uptake of antigen: I. Comparison of inferior vena cava and portal vein routes of immunization. Immunology 1973, 25, 92. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC1422991/pdf/immunology00335-0026.pdf (accessed on 30 May 2018).
- Smirnova, N.P.; Webb, B.T.; Bielefeldt-Ohmann, H.; Van Campen, H.; Antoniazzi, A.Q.; Morarie, S.E.; Hansen, T.R. Development of fetal and placental innate immune responses during establishment of persistent infection with bovine viral diarrhea virus. Virus Res. 2012, 167, 329–336. [Google Scholar] [CrossRef] [PubMed]
- Bielefeldt-Ohmann, H.; Smirnova, N.P.; Tolnay, A.E.; Webb, B.T.; Antoniazzi, A.Q.; van Campen, H.; Hansen, T.R. Neuro-invasion by a ‘Trojan Horse’ strategy and vasculopathy during intrauterine flavivirus infection. Int. J. Exp. Pathol. 2012, 93, 24–33. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3311019/ (accessed on 30 May 2018). [CrossRef] [PubMed]
- Webb, B.T.; Norrdin, R.W.; Smirnova, N.P.; Campen, H.V.; Weiner, C.M.; Antoniazzi, A.Q.; Bielefeldt-Ohmann, H.; Hansen, T.R. Bovine viral diarrhea virus cyclically impairs long bone trabecular modeling in experimental persistently infected fetuses. Vet. Pathol. 2012, 49, 930–940. [Google Scholar] [CrossRef] [PubMed]
- Chase, C.C.; Thakur, N.; Darweesh, M.F.; Morarie-Kane, S.E.; Rajput, M.K. Immune response to bovine viral diarrhea virus—Looking at newly defined targets. Anim. Health Res. Rev. 2015, 16, 4–14. [Google Scholar] [CrossRef] [PubMed]
- Glew, E.J.; Howard, C.J. Antigen-presenting cells from calves persistently infected with bovine viral diarrhoea virus, a member of the Flaviviridae, are not compromised in their ability to present viral antigen. J. Gen. Virol. 2001, 82, 1677–1685. [Google Scholar] [CrossRef] [PubMed]
- Racanelli, V.; Rehermann, B. The liver as an immunological organ. Hepatology 2006, 43, S54–S62. [Google Scholar] [CrossRef] [PubMed]
- Crispe, I.N. The liver as a lymphoid organ. Annu. Rev. Immunol. 2009, 27, 147–163. [Google Scholar] [CrossRef] [PubMed]
- Schultz, R.D.; Dunne, H.W.; Heist, C.E. Ontogeny of the bovine immune response. Infect. Immun. 1973, 7, 981–991. Available online: http://iai.asm.org/content/7/6/981.full.pdf (accessed on 30 May 2018). [PubMed]
- Bertolino, P.; McCaughan, G.W.; Bowen, D.G. Role of primary intrahepatic T-cell activation in the ‘liver tolerance effect’. Immunol. Cell Biol. 2002, 80, 84–92. [Google Scholar] [CrossRef] [PubMed]
- Bowen, D.G.; Zen, M.; Holz, L.; Davis, T.; McCaughan, G.W.; Bertolino, P. The site of primary T cell activation is a determinant of the balance between intrahepatic tolerance and immunity. J. Clin. Investig. 2004, 114, 701–712. [Google Scholar] [CrossRef] [PubMed]
- Groux, H.; Bigler, M.; De Vries, J.E.; Roncarolo, M.G. Interleukin-10 induces a long-term antigen-specific anergic state in human CD4+ T cells. J. Exp. Med. 1996, 184, 19–29. [Google Scholar] [CrossRef] [PubMed]
- Limmer, A.; Ohl, J.; Kurts, C.; Ljunggren, H.G.; Reiss, Y.; Groettrup, M.; Momburg, F.; Arnold, B.; Knolle, P.A. Efficient presentation of exogenous antigen by liver endothelial cells to CD8+ T cells results in antigen-specific T-cell tolerance. Nat. Med. 2000, 6, 1348. [Google Scholar] [CrossRef] [PubMed]
- Guidotti, L.G.; Chisari, F.V. To kill or to cure: Options in host defense against viral infection. Curr. Opin. Immunol. 1996, 8, 478–483. [Google Scholar] [CrossRef]
- Huang, J.F.; Yang, Y.; Sepulveda, H.; Shi, W.; Hwang, I.; Peterson, P.A.; Jackson, M.R.; Sprent, J.; Cai, Z. TCR-mediated internalization of peptide-MHC complexes acquired by T cells. Science 1999, 286, 952–954. [Google Scholar] [CrossRef] [PubMed]
- Latchman, Y.; Wood, C.R.; Chernova, T.; Chaudhary, D.; Borde, M.; Chernova, I.; Iwai, Y.; Long, A.J.; Brown, J.A.; Nunes, R.; et al. PD-L2 is a second ligand for PD-1 and inhibits T cell activation. Nat. Immunol. 2001, 2, 261. [Google Scholar] [CrossRef] [PubMed]
- Okazaki, T.; Honjo, T. The PD-1–PD-L pathway in immunological tolerance. Trends Immunol. 2006, 27, 195–201. [Google Scholar] [CrossRef] [PubMed]
- Smirnova, N.P.; Bielefeldt-Ohmann, H.; Van Campen, H.; Austin, K.J.; Han, H.; Montgomery, D.L.; Shoemaker, M.L.; van Olphen, A.L.; Hansen, T.R. Acute non-cytopathic bovine viral diarrhea virus infection induces pronounced type I interferon response in pregnant cows and fetuses. Virus Res. 2008, 132, 49–58. [Google Scholar] [CrossRef] [PubMed]
- Davis, W.C.; Marusic, S.; Lewin, H.A.; Splitter, G.A.; Perryman, L.E.; McGuire, T.C.; Gorham, J.R. The development and analysis of species specific and cross reactive monoclonal antibodies to leukocyte differentiation antigens and antigens of the major histocompatibility complex for use in the study of the immune system in cattle and other species. Vet. Immunol. Immunopathol. 1987, 15, 337–376. [Google Scholar] [CrossRef]
- Cope, E.M.; Dilly, S.A. Kupffer cell numbers during human development. Clin. Exp. Immunol. 1990, 81, 485–488. [Google Scholar] [CrossRef] [PubMed]
- MacHugh, N.D.; Mburu, J.K.; Hamilton, M.J.; Davis, W.C. Characterisation of a monoclonal antibody recognising the CD3ε chain of the bovine T cell receptor complex. Vet. Immunol. Immunopathol. 1998, 61, 25–35. [Google Scholar] [CrossRef]
- Baszler, T.V.; Evermann, J.F.; Kaylor, P.S.; Byington, T.C.; Dilbeck, P.M. Diagnosis of naturally occurring bovine viral diarrhea virus infections in ruminants using monoclonal antibody-based immunohistochemistry. Vet. Pathol. 1995, 32, 609–618. [Google Scholar] [CrossRef] [PubMed]
- Cornish, T.E.; van Olphen, A.L.; Cavender, J.L.; Edwards, J.M.; Jaeger, P.T.; Vieyra, L.L.; Woodard, L.F.; Miller, D.R.; O’Toole, D. Comparison of ear notch immunohistochemistry, ear notch antigen-capture ELISA, and buffy coat virus isolation for detection of calves persistently infected with bovine viral diarrhea virus. J. Vet. Diagn. Investig. 2005, 17, 110–117. [Google Scholar] [CrossRef] [PubMed]
- Montgomery, D.L.; Van Olphen, A.; Van Campen, H.; Hansen, T.R. The fetal brain in bovine viral diarrhea virus-infected calves: Lesions, distribution, and cellular heterogeneity of viral antigen at 190 days gestation. Vet. Pathol. 2008, 45, 288–296. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Target | Cells Identified | Antibody Clone | Source | Isotype | Application | Dilution | Reference |
|---|---|---|---|---|---|---|---|
| MHC I | PBMC | H58A | VMRD | IgG2a | FC | (1:200) | Davis 1987 [42] |
| MHC II | APC, PBMC | H42A | VMRD | IgG2a | FC | (1:200) | Davis 1987 [42] |
| L-1 | Macrophage, Monocyte, Kupffer Cells | MAC387 | AbCam | IgG1 | IHC | (1:200) | Cope 1990 [43]; Risalde 2011 [20] |
| CD3 | T cell Lymphocytes | MM1A | VMRD | IgG1 | IFA | (1:200) | MacHugh 1998 [44] |
| BVDV | BVDV Infected Cells (viral antigen) | 15C5 | IDEX | IgG2 | IHC, IFA | (1:200) | Bazler 1995 [45]; Cornish 2005 [46]; Montgomery 2008 [47] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morarie-Kane, S.E.; Smirnova, N.P.; Hansen, T.R.; Mediger, J.; Braun, L.; Chase, C. Fetal Hepatic Response to Bovine Viral Diarrhea Virus Infection in Utero. Pathogens 2018, 7, 54. https://doi.org/10.3390/pathogens7020054
Morarie-Kane SE, Smirnova NP, Hansen TR, Mediger J, Braun L, Chase C. Fetal Hepatic Response to Bovine Viral Diarrhea Virus Infection in Utero. Pathogens. 2018; 7(2):54. https://doi.org/10.3390/pathogens7020054
Chicago/Turabian StyleMorarie-Kane, Susan E., Natalia P. Smirnova, Thomas R. Hansen, Jessica Mediger, Lyle Braun, and Christopher Chase. 2018. "Fetal Hepatic Response to Bovine Viral Diarrhea Virus Infection in Utero" Pathogens 7, no. 2: 54. https://doi.org/10.3390/pathogens7020054
APA StyleMorarie-Kane, S. E., Smirnova, N. P., Hansen, T. R., Mediger, J., Braun, L., & Chase, C. (2018). Fetal Hepatic Response to Bovine Viral Diarrhea Virus Infection in Utero. Pathogens, 7(2), 54. https://doi.org/10.3390/pathogens7020054