Detection and Molecular Identification of Salmonella Virulence Genes in Livestock Production Systems in South Africa


1
Discipline of Genetics, School of Life Sciences, University of KwaZulu-Natal, Private Bag X54001, Durban 4000, South Africa
2
Virology and Microbiology Research Group, College of Pharmacy, City University College of Ajman, Al Tallah 2, Ajman, P.O. Box 18484, UAE
*
Author to whom correspondence should be addressed.
Pathogens 2019, 8(3), 124; https://doi.org/10.3390/pathogens8030124
Submission received: 18 May 2019
/
Revised: 26 July 2019
/
Accepted: 5 August 2019
/
Published: 9 August 2019
Abstract
:Livestock are an important source of protein and food for humans, however opportunistic pathogens such as Salmonella spp. turn livestock into vehicles of foodborne diseases. This study investigated the prevalence of virulence genes in Salmonella spp. isolated from livestock production systems in two provinces of South Africa. During the period from May to August, 2018, a total of 361 faecal (189), oral (100), environmental (soil (36) and water (27)) and feed (9) samples were randomly collected from different animals (cattle, sheep, goats, pigs, ducks and chickens) that were housed in small-scale livestock production systems from Eastern Cape and KwaZulu-Natal Provinces in South Africa. Salmonella spp. were isolated and identified using microbiological and DNA molecular methods. Salmonella spp. were present in 29.0% of the samples of which 30.2% belonged to the Salmonella enterica species as confirmed by the positive amplification of the species specific iroB gene. Virulence genes that were screened from livestock-associated Salmonella were invA, iroB, spiC, pipD and int1. Statistically significant associations (p < 0.05) were established between the virulence genes, sampling location, animal host as well as the season when samples were collected. Furthermore, statistically significant (p < 0.05) positive correlations were observed between most of the virulence genes investigated. This is one of the recent studies to detect and investigate livestock-associated Salmonella spp. in South Africa. This study highlights the importance of continuous monitoring and surveillance for pathogenic salmonellae. It also demonstrated the detection and prevalence of virulent Salmonella spp. harbored by livestock in South Africa. This study demonstrated the potential risks of pathogenic Salmonella enterica to cause foodborne diseases and zoonotic infections from farm-to-fork continuum using the global one-health approach.
Keywords:
Salmonella; zoonosis; pathogenicity; virulence; food-borne; livestock; Xylose-Lysine-Deoxycholate; pathogen; PCR; integron; infection; humans; one-health1. Introduction
Salmonellae are facultative intracellular Gram-negative bacteria that cause high morbidity and mortality in a wide range of hosts including humans, birds, mammals, and insects [1]. Salmonellae are one of the most problematic, foodborne, and zoonotic pathogens that cause health threats and challenges to general human well-being [2]. Salmonella spp. reside in the gastrointestinal tract of warm-blooded animals. The bacteria cause salmonellosis in humans, a disease that is presented mostly by mild diarrhea, also well-known as food poisoning [3,4]. Salmonellosis may be fatal, depending on the dose of infection and the immune status of the infected individual [5]. In the United States, Salmonella spp. are currently on the top of the list of pathogens that cause foodborne infections [6]. In South Africa, Salmonella spp. are regarded as one of the leading causes of foodborne outbreaks [7]. Foodborne outbreaks were reported in South Africa due to consumption of animal and poultry contaminated products [8,9,10,11]. Salmonella infection causes economic losses in the agriculture sector and it negatively impacts food animals which are reared for the generation of income [12].
Although Salmonella is a major cause of foodborne diseases in South Africa, there are limited data on the disease since it usually causes self-limiting gastroenteritis and cases are rarely reported [8,12]. It has been reported that six out of seven Salmonella enterica serovar Enteritidis outbreaks that occurred in South Africa from 2013 to 2015 were of food origin [13]. It was reported that 141 (43%) out of the 327 foodborne outbreaks reported in South Africa between 2013 to 2017 were reported in warmer months from KwaZulu-Natal [7]. An outbreak of food origin caused by S. enterica serotype Virchow was reported at a school in South Africa [14].
Surveillance of Salmonella is frequently conducted by different organizations worldwide in order to study its prevalence and epidemiology [8,15]. In South Africa surveillance is mainly the responsibility of governmental departments such as the Department of Health as well as the Department of Agriculture, Forestry and Fisheries. In order to bolster the surveillance, government departments have collaborations with local universities and other research institutions. Salmonella species play a role in metabolism when it is in a non-virulent state [16]. Factors such as stressful conditions, environmental changes and mutations can trigger virulence in a bacterium thus turning a previously non-virulent strain into a pathogenic strain [17].
The Salmonella genus includes more than 2500 serological variants (serovars) and is broadly categorized into S. bongori and S. enterica species [18]. According to the U.S. Centers for Disease Control (CDC) S. enterica is further subdivided into six subspecies that are designated by taxonomic names such as S. enterica subsp. enterica, S. enterica subsp. salamae, S. enterica subsp. arizonae, S. enterica subsp. diarizonae, S. enterica subsp. houtenae and S. enterica subsp. indica. S. enterica species is highly diverse consisting of more than 2600 serovars which are further divided into typhoidal Salmonella and non-typhoidal Salmonella (NTS), depending on the disease it causes [19,20]. Typhoidal Salmonella spp. are restricted to human hosts while non-typhoidal Salmonella spp. can infect a wide range of hosts [21]. Faecal shedding of NTS results in environmental contamination and transmission to humans, leading to disease outbreak [16]. NTS has a broad host range and is often associated with foodborne outbreaks in humans [21]. S. enterica serovar Typhimurium and S. enterica serovar Enteritidis are the most frequently reported pathogens in Salmonella outbreaks and the prominent cause of gastroenteritis in humans [22,23].
Salmonella pathogenicity is mediated by numerous genes such as invA, spiC and pipD, which code for effectors that induce successful host infection. Pathogenicity of Salmonella is expressed in three ways such as host cell invasion, intracellular survival and colonization [24]. Numerous virulence genes are essential for Salmonella pathogenesis and these genes are located on various elements of the genome including the chromosome, plasmids, integrated bacteriophage DNA, Salmonella pathogenicity islands (SPIs), and Salmonella genomic islands (SGIs) [19,25]. SPIs are large gene cassettes and only SPI-1 and SPI-2 (not all SPIs) encode a membrane-associated type III secretion system (T3SS) [26] which secretes a pool of 44 effector proteins [27], that alter the functioning of eukaryotic cells in order to facilitate bacterial pathogenicity inside the cell [28,29,30]. Previous studies reported that SPIs are acquired by horizontal transmission and vertically pass to new clones [31]. More than 20 SPIs have been characterized, with greater focus on SPI-1 and SPI-2 that function via encoded T3SS since they harbor host invasion and intracellular survival genes [29,32]. Inside the host cell, SPI-2 expresses genes that are important in intracellular survival, proliferation, and persistence in internal organs such as the spleen and liver [30,33]. Salmonella spp. use virulence genes and factors located in SPI-1 for cell invasion and to initiate its pathogenicity [29]. The invasion A (invA) is one of the most studied virulence factors that is also used as a biomarker for Salmonella spp. detection as it contains sequences that are unique to the genus Salmonella. [34] Invasion A is a factor in the outer membrane of Salmonella spp. that is responsible for entering the host epithelial cells in the intestines thus initiating infection [34]. The inv locus in S. enterica serovar Typhimurium was characterized and it was reported that invA is essential in the display of virulence in the intestine [35].
One of the most important genes is iroB, a Fur-regulated gene located in a large DNA region which is used in the detection of S. enterica subspecies enterica [36,37]. Previous studies which detected typhoid and non-typhoid Salmonella by PCR used invA and iroB together with flagellar genes [38,39]. Furthermore, iroB was used to detect Salmonella from blood in another study [40]. The IroB gene is a member of the iroA (iroBCDEN) gene cluster which is responsible for the synthesis and transport of enterobactin, a siderophore produced by Salmonella spp. and is essential for iron uptake inside the host [41]. Besides enabling bacterial iron uptake, expression of the iroA cluster also facilitates the host immune escape by interrupting macrophage homeostasis [42]. The specific role of iroB is to encode glucosyltransferase which glucosylates enterobactin [41]. Enterobactin glucosylation contributes to the virulence of the bacteria by preventing the host antimicrobial protein (lipocalin-2) from sequestering the siderophore [43,44].
spiC is another gene in the SPI-2 that is essential for intracellular survival and host defense escape [45]. Macrophages are important innate immune barriers which defend the host against infections and their function is activated by gamma interferon and facilitated by factors such as cytokines and eicosanoids [46]. Upon activation, macrophages kill pathogens that are capable of surviving inside them. In order to escape the host’s defense, spiC is involved in the signal transduction pathway which expresses the suppressor of cytokine signaling, leading to gamma interferon signaling inhibition [45]. It was reported that spiC is also involved in the translocation of effectors into the cytosol of macrophages [47].
The SPI-5 harbors six genes in which mutations in four of these genes were reported to radically lower enteropathogenicity [48,49]. pipD is one of the genes in the SPI-5 that is involved in inflammatory enteritis by coding a cysteine protease homolog that is essential in the long term systemic infection [48,49,50].
Gastroenteritis is the most common disease caused by non-typhoidal Salmonella. This disease usually resolves without treatment but it can be systemic in severe cases and require antimicrobial treatment. There is, however, an enormous challenge with using antibiotics as Salmonella is one of the ‘superbugs’ which are resistant to several classes of antibiotics [51]. The antimicrobial resistance phenotype is attributed to the possession of class 1 integron by some of the Salmonella serovars.
The class 1 integron is a mobilizable cluster of antimicrobial resistance genes found in Salmonella genomic island [52,53,54,55]. Class 1 integrons are made up of integrase gene, a primary recombination site and a promoter region [56]. The role of int1 is to recombine gene cassettes (associated with antibiotic resistance), which are only transcribed in an integron since they lack a promoter [57,58]. Class 1 integron carries gene cassettes for resistance to antibiotics such as those which were used as first line treatment for salmonellosis. The presence of class 1 integrons carrying gene cassettes in virulent Salmonella spp. increases the threat to humans as it limits the treatment options available [59,60]. Infections by non-tyhpoidal Salmonella spp. affect both developing and developed countries. Studies and incidences revealed that food animals are the carriers of NTS and are potential zoonotic sources of infection to humans [61,62,63]. Against this background, this study focused on the detection and determination of the prevalence of virulent Salmonella spp. in livestock production systems in the KwaZulu-Natal and Eastern Cape Provinces in South Africa.
2. Materials and Methods
2.1. Ethical Approval
The study was approved by the Animal Research Ethics Committee of the University of Kwa-Zulu Natal (Reference numbers AREC/051/017M, AREC 071/017 and AREC 014/018). The field sampling protocols, samples collected from animals, and the research were conducted in full compliance with Section 20 of the Animal Diseases Act of 1984 (Act No 35 of 1984) and were approved by the South African Department of Agriculture, Forestry and Fisheries DAFF (Section 20 approval reference number 12/11/1/5 granted to Prof. Dr. ME El Zowalaty).
2.2. Samples and Pre-Enrichment
During the autumn and winter months of the year 2018, a total of three hundred and sixty-one (361) oral, faecal, soil, water and feed samples were randomly collected from different animal hosts such as cattle, sheep, goats, pigs, ducks and chickens. The animals were housed in small-scale commercial farms in Flagstaff (O.R Tambo, Eastern Cape), Verulam (eThekwini, KwaZulu-Natal) and the South Coast (Amandawe and Mtwalume, UGU, KwaZulu-Natal) as depicted in Figure 1.
All samples were randomly collected between May and August 2018 from different farms in Eastern Cape and KwaZulu-Natal Provinces of South Africa. In the farms, livestock were bought from large-scale farms and were sold to the communities. ISO 6579-1 was used to collect and isolate Salmonella however, PCR was used for detection and confirmation of the presence of Salmonella spp. Fresh environmental faecal samples emanating from livestock, as well as samples from livestock environments including soil, water and feed were collected using sterile collection swabs. All swab samples were transferred into 10 mL of 0.1% buffered sterilized peptone water (Merck, Johannesburg, South Africa). Water samples were collected from the containers inside the livestock houses. Samples were transported on ice to the discipline of genetics laboratories where enrichment of the samples was conducted by incubation overnight at 37 °C.
2.3. Selective Enrichment
From each of the enriched samples, 0.1 mL was aseptically transferred into 10 mL of Rappaport Vassiliadis (RV) broth and incubated for 24 h at 42 °C (Sigma-Aldrich, Mumbai, India). RV is a selective medium that is enriched with malachite green which inhibits the growth of microorganisms other than Salmonella. A previously identified and confirmed S. enterica was used as a positive control for this experiment [63]. Microbiological isolation was performed on Xylose-Lysine-Deoxycholate (XLD) agar (Sigma-Aldrich, Buchs, Switzerland) by aseptically streaking a loopful of the culture from RV broth onto the XLD plates. S. enterica is differentiated from Escherichia coli and Shigella spp. by producing red colonies with black centers on XLD agar. After 24 h of incubation at 35 °C, the plates were observed for the growth of the expected colonies. Single colonies were picked from each plate and transferred into tubes containing 10 mL of tryptose soy broth (Merck, Johannesburg, South Africa) and incubated at 37 °C for 18–24 h. A 2 mL of the culture was used for DNA extraction. Equal amounts of 0.5 mL each of 60% glycerol and Salmonella pure culture were mixed in 1.5 mL cryotubes and stored at −80 °C for future use.
2.4. DNA Extraction
Total genomic DNA was extracted from Salmonella cultures using the conventional boiling method. One milliliter of the cultured sample was transferred into 1.5 mL Eppendorf tube and centrifuged at 14,000× g for 5 min. The supernatant was discarded and another 1 mL of culture was added to the pellet and centrifuged again to get a bigger pellet. Six hundred µL of sterile distilled water was added to the pellet and centrifuged for 5 min at 14,000× g. The supernatant was discarded, 200 µL of sterile distilled water was added again and incubated in a heating block at 100 °C (Labnet, FL, USA) for 10 min with immediate cooling on ice for 5 min. After cooling, the sample was centrifuged at 14,000× g for 5 min; the resulting supernatant was transferred into a fresh Eppendorf tube and stored at −20 °C until use in PCR.
2.5. Molecular Confirmation of Salmonella spp. Using PCR
Salmonella spp. isolates were confirmed by amplifying the invA gene, which is genus specific and the iroB gene for identification of S. enterica using specific primers as previously reported [36,64].
The PCR reaction volume was 25 µL which consisted of 12.5 µL Taq master mix (Thermo-Fischer Scientific, Johannesburg, South Africa), 0.5 µL each of forward and reverse primers, 6.5 µL sterile distilled water and 5 µL template DNA. The PCR reaction conditions consisted of initial denaturation cycle for 5 min, followed by 34 amplification cycles for invA gene using the following conditions: Denaturation for 30 s at 95 °C, annealing for 30 s at 61 °C, extension for 1 min at 72 °C and final extension for 5 min at 72 °C. The same amplification parameters were used for the iroB gene using a different annealing temperature of 55 °C. The PCR amplicons were stored at −20 °C till future use.
2.6. Gel Electrophoresis and Visualization of PCR Products
One and a half percent (1.5%) agarose gel was prepared by slightly boiling 1.5 g agarose powder (Cleaver scientific, Rugby, UK) in 100 mL of 1× TAE buffer (Bio Concept, Allschwil, Switzerland). A volume of two µL of ethidium bromide was added to the gel before pouring it into the casting tray. A one hundred bp molecular weight maker (New England biolabs, Beijing, China) was used to estimate the size of the products. A volume of two µL of the molecular weight marker was mixed with 2 µL of 6× purple dye and diluted with 8 µL of nuclease free water. A volume of ight µL of each of the PCR products was loaded in each well. Electrophoresis was carried out at 80 volts for 55 min using Enduro gel XL electrophoresis system (Labnet, FL, USA). DNA bands from the gel were visualized using ChemiDoc MP imaging system (Bio-Rad, CA, USA).
2.7. Determination of Virulence Profiles of Salmonella Isolates
3. Statistical Analyses
Descriptive statistics (IBM SPSS, version 25) and Microsoft excel 2016 were used to determine the frequencies of Salmonella spp. in each livestock species. The effects of location (whether the samples were collected from Flagstaff, Verulam or South Coast), animal host (chicken, pig, sheep, cow, duck, and goat) and season of sampling (autumn or winter) on the presence of virulent Salmonella spp. were investigated using the Fischer’s exact test. The Fischer’s exact test is a parametric test of significance that is used in the place of a Chi- Square test in two by two tables. The Pearson’s correlation test was implemented in order to establish the strength and direction of the relationship between the virulence genes. Furthermore, binary logistic regression was used to model the association between the binary outcomes (presence or absence of Salmonella spp. and virulence genes) and exposure variables (location, animal host, sampling season, sample material). The dependent variable was determined whether a virulence gene was present (1) or absent (0). The association between using XLD and PCR in detecting Salmonella spp. was measured via the Pearson’s correlation test. The null hypothesis tested was that there is no significant (p > 0.05) association between location, animal host and season of sampling in the prevalence of virulent Salmonella spp. The statistical results were regarded as significant only if the p < 0.05. All statistical tests were performed using IBM SPSS software (version 25).
4. Results
Out of the collected 361 samples (114 samples from chicken, 79 samples from goat, 58 samples from pig, 50 samples from sheep, 50 samples from cow and 10 samples from duck) as shown in Table 1 it was found that 195 (54%) samples showed positive growth on RV medium and XLD agar.
The positive control strain showed red colonies with black centers, as expected for S. enterica on XLD agar however, some of the XLD plates with tested samples showed yellow colonies as a result of lactose fermentation, a characteristic used to differentiate Salmonella spp. from E. coli spp. Single colonies were selected for DNA extraction and confirmation by PCR amplification.
The invA gene is a genus specific marker that is used for detection of Salmonella spp. [64]. Total genomic DNA was extracted from all the 195 isolates which showed positive growth on XLD agar. The extracted DNA was of pure quality, with the mean absorbance ratio (A260/A280) of 2.0, which is accepted for pure DNA. Positive amplification of the invA gene was regarded as an indication of the presence of Salmonella spp. Out of the 195 presumptive Salmonella isolates, 106 (29.4% of the collected samples) were confirmed to be Salmonella spp. by amplification of the invA gene (Figure 2A). S. enterica was confirmed by the detection of the iroB gene of band size of 606 bp (Figure 2B) in 32 out of the 106 (30.2%) samples [64].
The total number of samples collected from each animal species were shown in Table 1. The prevalence of Salmonella spp. in different hosts was determined and it was found that the highest prevalence was in ducks, followed by chickens, sheep, pigs, cows and goats respectively. In this study, 30.2% of the Salmonella isolates were confirmed to be S. enterica by iroB gene amplification. The 606 bp amplicon of the iroB gene was shown in Figure 2B.
Figure 3 show the amplification of virulence genes of Salmonella spp, (A) a 309 bp band for spiC amplicon, (B) a 350 band for bp pipD amplicon and (C) a 569 bp band for int1 amplicon respectively.
The distribution of the virulence genes in the current study was found to be more frequent in chicken, goat, sheep and cow (Figure 4), with eight chicken isolates, one cow isolate, one sheep isolate and two goat isolates portraying the capability to manifest infection as all isolates from all livestock production animals harbored the screened virulence genes. The frequency of the tested genes was highest in chicken while it was not significantly different in other livestock hosts (Figure 4). However, duck and pig possessed only three of the tested virulence genes respectively. Isolates from goat and sheep had a higher prevalence of virulence genes compared to cow hosts.
The overall results on the prevalence of the virulence genes are illustrated in Figure 5. Out of the 106 isolates with invA gene, 30% possessed iroB gene while 62.3% possessed pipD gene, 18.9% possessed spiC gene and the int1 gene was found in 34.9% of the isolates. PipD was the most prevalent virulence gene compared to the other tested genes besides invA. The difference in the prevalence of the tested virulence genes can be attributed to the location of the gene in Salmonella. SpiC and iroB are both located in the SPI-2 which is found only in S. enterica while invA and pipD are in SPIs 1 and 5. Int1 is found in the SGI-1 and plasmids.
The effects of location, animal host species and sample material on the presence of iroB. spiC, pipD, and int1 were evaluated by binary logistic regression. Animal host species and sample material did not significantly (p > 0.05) predict the presence of all the virulence genes. Table 2 showed that location significantly (p < 0.05) contributes to predicting the presence of iroB, pipD and int1. Verulam and Flagstaff were statistically significant (p < 0.05) for location. The odds ratio reveals that Verulam was more likely to have the presence of iroB [OR = 5.429 (1.577, 18.686)], pipD [OR = 19.991 (2.330, 171.530)] and int1 [OR = 8.053 (1.801, 59.968)] compared to Flagstaff.
The association between using XLD and amplifying the invA gene, a universal marker for Salmonella spp., used for Salmonella detection was assessed via the Pearson’s correlation following the difference in the results obtained from these methods (Figure 5). A significant (p < 0.05) positive 40.2% correlation between using XLD and invA amplification was obtained (Table 3). A significant positive correlation (p < 0.05) between the tested virulence genes was observed, except for spiC and int1. The highest correlation was between spiC and iroB which are both located in the SPI-2. Although the tested virulence genes have different locations; they are all responsible for virulence in Salmonella therefore the significant correlation means that one gene can be used to predict the presence of another gene.
The Fisher’s exact test was used to investigate the association between the presence of virulence genes and location, animal species, seasons of sampling as well as sample material. As shown in Table 4, there was a significant (p < 0.05) association between the prevalence of the virulence genes with location, animal host and season of sampling however, there was no significant (p > 0.05) association between sample material and virulence genes except for int1. It was, therefore, evident that the variables that were tested except the sample material significantly influenced the presence or absence of virulence genes.
5. Discussion
S. enterica is responsible for infections in humans and animals, with serovars Enteritidis and Typhimurium being the most reported [67]. The present study investigated the prevalence and genetic characteristics of Salmonella virulence genes in livestock production systems in South Africa using microbiological culturing and molecular methods.
It is recommended that both microbiological culture methods and DNA molecular techniques are concurrently applied for the detection of Salmonella spp. even though culturing is more laborious and time consuming while molecular techniques are quick and more sensitive [68,69]. Of interest to this study was that most of the XLD agar plates had yellow colonies instead of the expected red colonies with black centers. Yellow-pigmented colonies appear due to lactose fermentation by the microorganisms. Escherichia coli grow as yellow colonies on XLD while Salmonella spp. is known as non-lactose fermenters and appear as pink with black center colonies. There is some controversy however, with using XLD for Salmonella spp. detection as there are Salmonella serovars which have horizontally inherited the lactose fermentation gene from E. coli [70]. The acquisition of the lac operon by Salmonella spp. reduces the virulence potential of the pathogen [71]. It is worth mentioning that microbial culture methods are presumptive and have to be complemented by genomics and molecular methods for the accurate identification and subsequent characterization of microbial species. In the current study, we called both red colonies with black centers as well as yellow colonies because they are both presumptive. The final confirmation was based on molecular PCR methods and the positive amplification of the genus specific biomarker, i.e., the invA gene.
Several studies have reported that S. enterica serovar Typhimurium and other S. enterica serovars which grow as yellow colonies on XLD agar [72,73]. Out of the 195 colonies that grew on the presumptive XLD agar, only 35 were red with black centers while 160 colonies were yellow pigmented, and some with black centers. Screening of the 195 isolates for the invA gene showed that the prevalence of Salmonella spp. was 29% (106 out of 361 samples) as depicted in Figure 5. This raises a concern regarding the microbiological media which use lactose fermentation characteristic for differentiating Salmonella spp. from E. coli as some Salmonella isolates can be falsely reported as E. coli based on phenotypic characters and colony morphology. Our findings revealed that 32 samples (30.2%) of the Salmonella isolates were confirmed to be S. enterica using molecular PCR methods of iroB gene amplification. This could be explained that the 74 invA positive-iroB negative Salmonella isolates may belong to S. bongori species and suggested a co-infection which requires further investigation. In addition, as shown in the supplementary data, it may be indicated that the sensitivity of the iroB PCR to identify S. enterica was not perfect, since five iroB-negative samples were spiC positive. Since spiC is a SPI-2 gene and this island is specific to S. enterica (not present in S. bongori), this result indicated that at least these five samples also correspond to S. enterica.
In the present study, we only focused on the detection of S. enterica since this species is of national public health importance and is the main cause of salmonellosis in the area.
Several studies reported the isolation of Salmonella spp. from food animals [62,74,75]. Previous studies reported lower prevalence rates of Salmonella spp. than the results of the current study with rates of 2.81%, 8.3% and 10.4%, respectively [62,74,75]. However; recent studies reported higher Salmonella spp. prevalence rates of 51% and 48%, respectively [63,76]. The difference in the prevalence of Salmonella spp. isolated from livestock can be explained by factors such as environmental conditions, farm management, and biosecurity practices. Most of the previous studies focused on Salmonella spp. isolation from poultry and poultry products [63,77,78]. Our study is unique in that it determined the prevalence of Salmonella spp. in different animals and animal hosts including avian, swine, ovine, and bovine, some of which were housed together within the same epidemiological distance. As previously alluded, Salmonella spp. may be in a virulent or non-virulent state and asymptomatic food animals are possible potential sources of transmission of virulent Salmonella to humans [16].
Since Salmonella pathogenicity is determined by genes which work collaboratively for successful infection, invA, iroB, spiC and pipD were screened from the isolates. The possibility of the isolates to be resistant to antibiotics was determined by screening for the class 1 integron gene. Figure 4 shows the prevalence of the virulence genes in all the livestock. Prevalence rates of 47% for SpiC and 35% for pipD in Salmonella spp. isolated from chickens in South Africa was reported [63]. Another study reported prevalence rates of 78% for spiC and 95% for pipD [79] in clinical samples from human and livestock. The prevalence rates of spiC were higher in these studies [63,79] than the rate in the current study however, the prevalence rate of pipD in the present study was higher than that was previously reported [63].
IroB, spiC and pipD are located in the SPIs while int1 (class 1 integron) is located in the Salmonella genomic island-1 which can explain the difference in the prevalence of these genes with respect to sample material. SPIs are found in the chromosome of pathogenic strains of Salmonella spp. while an integron is a mobile element that can integrate into the chromosome of a bacterium [80,81,82]. The presence of 34.9% int1 (Figure 5) from isolates in the current study explains that there is a possibility of antibiotic resistance emergence in the isolated Salmonella spp. The class 1 integron is associated with resistances to antibiotics such as ampicillin, chloramphenicol, streptomycin, sulfonamides ant tetracycline [83], which were used as first line drugs for the treatment of salmonellosis. The detection of different virulence genes in the Salmonella isolates in this study represents a public health threats including zoonotic potential and development of antimicrobial resistance. This study demonstrated that the presence of the virulence genes was significantly (p < 0.05) predicted by location when location, animal host species, season of sampling and sample material were evaluated by binary logistic regression as shown in Table 2. Interestingly, the findings of the present study showed that the presence of int1 gene is more associated to faecal samples, and the presence of virulence genes is more associated to the samples obtained in winter compared to autumn. Environmental conditions in which the animals are housed and reared might have an impact on triggering virulence in Salmonella. It was previously reported that the risk of salmonellosis is related to an increase in temperature in coastal areas compared to non-coastal areas [84]. Similarly, in the present study, Verulam is an urban area where livestock feed mostly on preserved food, which have traces of chemicals and amino acids while livestock in rural areas feed on natural grass that could explain the high prevalence of virulent Salmonella spp. in Verulam.
6. Conclusions
The findings of the current study showed that food animals are a potential source of virulent Salmonella spp., exposing humans to zoonotic infections through potential exposure route via food or direct exposure. Control and biosecurity measures are not always implemented in small-scale farms, particularly in rural areas in South Africa. Small-scale chicken farms are abundant in South Africa, as they do not require large hectares of land and chicken are more feasible and economic to rear compared to the other livestock animals. Salmonella spp. carried by livestock in this study does not only pose infection risks to humans but also has the potential of being resistant to antibiotics once infection has initiated and manifested. Some people in rural areas lack knowledge about contamination risk that comes with livestock therefore action plans must be taken to educate people about the importance of hygiene, especially with animal and food handling. This study also demonstrated the importance of implementing one-health control measures in addressing the challenges of foodborne disease and virulence in zoonotic pathogens including Salmonella spp.
Supplementary Materials
Supplementary File 1Author Contributions
Conceptualization, O.T.Z. and M.E.E.Z.; methodology, T.P.M.; software, O.T.Z.; validation, O.T.Z. and M.E.E.Z.; formal analysis, O.T.Z.; investigation, M.E.E.Z. and O.T.Z.; resources, O.T.Z. and M.E.E.Z.; data curation, T.P.M., M.E.E.Z. and O.T.Z.; writing—original draft preparation, T.P.M.; writing—review and editing, M.E.E.Z. and O.T.Z.; visualization, M.E.E.Z. and O.T.Z.; supervision, O.T.Z. and M.E.E.Z.; project administration, O.T.Z. and M.E.E.Z.; funding acquisition, O.T.Z. and M.E.E.Z.; critical revisions and editing, M.E.E.Z.
Funding
This research was funded by SA-NRF South African National Research Foundation Thuthuka Funding Instrument: TTK170411226583.
Acknowledgments
We would like to acknowledge the four anonymous farmers who allowed us to collect samples for this research from their farms. The authors would like to thank the three anonymous reviewers for their valuable comments that significantly improved the manuscript. We would like to thank the South African National Research Foundation for supporting this research through the Thuthuka Funding Instrument (grant number TTK170411226583). We would also like to thank the College of Agriculture, Engineering and Science as well as the School of Life Sciences at University of KwaZulu-Natal (Westville campus) for their support during the execution of this research. Authors would like to thank Stephen M. Boyle from Virginia Tech for editorial comments on the manuscript.
Conflicts of Interest
The authors declare that they have no financial or personal relationships that may have inappropriately influenced them in writing this article.
References
- Bäumler, A.J.; Tsolis, R.M.; Ficht, T.A.; Adams, L.G. Evolution of Host Adaptation in Salmonella enterica. Infect. Immun. 1998, 66, 4579–4587. [Google Scholar] [PubMed]
- Balasubramanian, R.; Im, J.; Lee, J.-S.; Jeon, H.J.; Mogeni, O.D.; Kim, J.H.; Rakotozandrindrainy, R.; Baker, S.; Marks, F. The global burden and epidemiology of invasive non-typhoidal Salmonella infections. Hum. Vaccines Immunother. 2019, 15, 1421–1426. [Google Scholar] [CrossRef] [PubMed]
- Hennekinne, J.-A.; Herbin, S.; Firmesse, O.; Auvray, F. European food poisoning outbreaks involving meat and meat-based products. Procedia Food Sci. 2015, 5, 93–96. [Google Scholar] [CrossRef]
- Food and Drug Administration. Bad Bug Book, Foodborne Pathogenic Microorganisms and Natural Toxins; Center for Food Safety and Applied Nutrition of the Food and Drug Administration (FDA), US Department of Health and Human Services: Silver Spring, MD, USA, 2012.
- World Health Organization. Salmonella (Non-Typhodial). 2018. Available online: https://www.who.int/news-room/fact-sheets/detail/salmonella-(non-typhoidal) (accessed on 18 April 2018).
- U.S. Food & Drug Administration. Foodborne Illnesses: What You Need to Know. 2018. Available online: https://www.fda.gov/food/foodborneillnesscontaminants/foodborneillnessesneedtoknow/default.htm (accessed on 30 October 2018).
- Shonhiwa, A.; Ntshoe, G.; Essel, V.; Thomas, J.; McCarthy, K. A review of foodborne diseases outbreaks reported to the outbreak response unit, national institute for communicable diseases, South Africa, 2013–2017. Int. J. Infect. Dis. 2019, 79 (Suppl. 1), 73. [Google Scholar] [CrossRef]
- Marks, F.; Von Kalckreuth, V.; Aaby, P.; Adu-Sarkodie, Y.; El Tayeb, M.A.; Ali, M.; Aseffa, A.; Baker, S.; Biggs, H.M.; Bjerregaard-Andersen, M.; et al. Incidence of invasive Salmonella disease in sub-Saharan Africa: A multicentre population-based surveillance study. Lancet Glob. Health 2017, 5, e310–e323. [Google Scholar] [CrossRef]
- Olobatoke, R.Y.; Mulugeta, S.D. Incidence of non-typhoidal Salmonella in poultry products in the North West Province, South Africa. S. Afr. J. Sci. 2015, 111, 1–7. [Google Scholar] [CrossRef]
- Christison, C.; Lindsay, D.; Von Holy, A. Microbiological survey of ready-to-eat foods and associated preparation surfaces in retail delicatessens, Johannesburg, South Africa. Food Control 2008, 19, 727–733. [Google Scholar] [CrossRef]
- van Nierop, W.; Dusé, A.; Marais, E.; Aithma, N.; Thothobolo, N.; Kassel, M.; Stewart, R.; Potgieter, A.; Fernandes, B.; Galpin, J.; et al. Contamination of chicken carcasses in Gauteng, South. Africa, by Salmonella, Listeria monocytogenes and Campylobacter. Int. J. Food Microbiol. 2005, 99, 1–6. [Google Scholar] [CrossRef] [PubMed]
- International Federation for Animal Health. The Costs of Animal Disease—A Report Produced for the International Federation for Animal Health; IFAH; Oxford Analytica Ltd.: Oxford, UK, 2012; Available online: https://www.bft-online.de/fileadmin/bft/publikationen/IFAH_Oxford-Analytica_The-Costs-of-Animal-Disease_October2012.pdf (accessed on 12 June 2018).
- Muvhali, M.; Smith, A.M.; Rakgantso, A.M.; Keddy, K.H. Investigation of Salmonella Enteritidis outbreaks in South. Africa using multi-locus variable-number tandem-repeats analysis, 2013–2015. BMC Infect. Dis. 2017, 17, 661. [Google Scholar] [CrossRef] [PubMed]
- Smith, A.M.; Gouws, A.-M.; Hoyland, G.; Sooka, A.; Keddy, K.H. Outbreaks of food-borne disease: A common occurrence but rarely reported. S. Afr. Med. J. 2007, 97, 1272. [Google Scholar] [PubMed]
- Hendriksen, R.S.; Vieira, A.R.; Karlsmose, S.; Wong, D.M.L.F.; Jensen, A.B.; Wegener, H.C.; Aarestrup, F.M. Global monitoring of Salmonella serovar distribution from the World Health Organization Global Foodborne Infections Network Country Data Bank: Results of quality assured laboratories from 2001 to 2007. Foodborne Pathog. Dis. 2011, 8, 887–900. [Google Scholar] [CrossRef] [PubMed]
- Ahmer, B.M.; Gunn, J.S. Interaction of Salmonella spp. with the intestinal microbiota. Front. Microbiol. 2011, 2, 101. [Google Scholar] [PubMed]
- Anderson, C.J.; Kendall, M.M. Salmonella enterica serovar Typhimurium strategies for host adaptation. Front. Microbiol. 2017, 8, 1983. [Google Scholar] [CrossRef] [PubMed]
- Porwollik, S.; Boyd, E.F.; Choy, C.; Cheng, P.; Florea, L.; Proctor, E.; McClelland, M. Characterization of Salmonella enterica subspecies I genovars by use of microarrays. J. Bacteriol. 2004, 186, 5883–5898. [Google Scholar] [CrossRef] [PubMed]
- Card, R.; Vaughan, K.; Bagnall, M.; Spiropoulos, J.; Cooley, W.; Strickland, T.; Davies, R.; Anjum, M.F. Virulence characterisation of Salmonella enterica isolates of differing antimicrobial resistance recovered from UK livestock and imported meat samples. Front. Microbiol. 2016, 7, 640. [Google Scholar] [CrossRef]
- Grimont, P.A.; Weill, F.-X. Antigenic Formulae of the Salmonella Serovars; WHO Collaborating Centre for Reference and Research on Salmonella: Paris, France, 2007; Volume 9, pp. 1–166. [Google Scholar]
- Gal-Mor, O.; Boyle, E.C.; Grassl, G.A. Same species, different diseases: How and why typhoidal and non-typhoidal Salmonella enterica serovars differ. Front. Microbiol. 2014, 5, 391. [Google Scholar] [CrossRef]
- Kariuki, S.; Revathi, G.; Kariuki, N.; Kiiru, J.; Mwituria, J.; Muyodi, J.; Githinji, J.W.; Kagendo, D.; Munyalo, A.; Hart, C.A. Invasive multidrug-resistant non-typhoidal Salmonella infections in Africa: oonotic or anthroponotic transmission? J. Med. Microbiol. 2006, 55, 585–591. [Google Scholar] [CrossRef]
- Smith, A.M. Review of molecular subtyping methodologies used to investigate outbreaks due to multidrug-resistant enteric bacterial pathogens in sub-Saharan Africa. Afr. J. Lab. Med. 2019, 8, 760. [Google Scholar] [CrossRef]
- Foley, S.L.; Johnson, T.J.; Ricke, S.C.; Nayak, R.; Danzeisen, J. Salmonella pathogenicity and host adaptation in chicken-associated serovars. Microbiol. Mol. Biol. Rev. 2013, 77, 582–607. [Google Scholar] [CrossRef]
- Bayoumi, M.A.; Griffiths, M.W. Probiotics down-regulate genes in Salmonella enterica serovar Typhimurium pathogenicity islands 1 and 2. J. Food Prot. 2010, 73, 452–460. [Google Scholar] [CrossRef]
- Galán, J.E. Salmonella interactions with host cells: Type III secretion at work. Annu. Rev. Cell Dev. Biol. 2001, 17, 53–86. [Google Scholar] [CrossRef] [PubMed]
- Ramos-Morales, F. Impact of Salmonella enterica type III secretion system effectors on the eukaryotic host cell. ISRN Cell Biol. 2012, 2012, 787934. [Google Scholar] [CrossRef]
- Hallstrom, K.N.; McCormick, B.A. Pathogenicity Islands: Origins, Structure, and Roles in Bacterial Pathogenesis, in Molecular Medical Microbiology; Elsevier: Issy-les-Moulineaux, France, 2015; pp. 303–314. [Google Scholar]
- Coburn, B.; Li, Y.; Owen, D.; Vallance, B.A.; Finlay, B.B. Salmonella enterica serovar Typhimurium pathogenicity island 2 is necessary for complete virulence in a mouse model of infectious enterocolitis. Infect. Immun. 2005, 73, 3219–3227. [Google Scholar] [CrossRef] [PubMed]
- Santos, R.L.; Zhang, S.; Tsolis, R.M.; Kingsley, R.A.; Adams, L.G.; Baumler, A.J. Animal models of Salmonella infections: Enteritis versus typhoid fever. Microbes Infect. 2001, 3, 1335–1344. [Google Scholar] [CrossRef]
- Juhas, M.; Van Der Meer, J.R.; Gaillard, M.; Harding, R.M.; Hood, D.W.; Crook, D.W. Genomic islands: Tools of bacterial horizontal gene transfer and evolution. FEMS Microbiol. Rev. 2009, 33, 376–393. [Google Scholar] [CrossRef] [PubMed]
- Fookes, M.; Schroeder, G.N.; Langridge, G.C.; Blondel, C.J.; Mammina, C.; Connor, T.R.; Seth-Smith, H.; Vernikos, G.S.; Robinson, K.S.; Sanders, M. Salmonella bongori provides insights into the evolution of the Salmonellae. PLoS Pathog. 2011, 7, e1002191. [Google Scholar] [CrossRef]
- Gerlach, R.G.; Hensel, M. Salmonella pathogenicity islands in host specificity, host pathogen-interactions and antibiotics resistance of Salmonella enterica. Berl. Munch. Tierarztl. Wochenschr. 2007, 120, 317–327. [Google Scholar]
- El-Sebay, N.A.; Abu Shady, H.M.; El-Rashed El-Zeedy, S.A.; Samy, A.A. InvA gene sequencing of Salmonella Typhimurium isolated from Egyptian poultry. Asian J. Sci. Res. 2017, 10, 194–202. [Google Scholar] [CrossRef]
- Galán, J.E. Molecular genetic bases of Salmonella entry into host cells. Mol. Microbiol. 1996, 20, 263–271. [Google Scholar] [CrossRef]
- Bäumler, A.J.; Heffron, F.; Reissbrodt, R. Rapid detection of Salmonella enterica with primers specific for iroB. J. Clin. Microbiol. 1997, 35, 1224–1230. [Google Scholar]
- Baumler, A.J.; Tsolis, R.M.; Van Der Velden, A.W.; Stojiljkovic, I.; Anic, S.; Heffron, F. Identification of a new iron regulated locus of Salmonella Typhi. Gene 1996, 183, 207–213. [Google Scholar] [CrossRef]
- Li, X.; Yang, F.; Gao, W.; Song, H.; Tian, H.; Xu, B. Application of pyrosequencing for Salmonella enterica rapid identification. J. Microbiol. Methods 2012, 89, 49–52. [Google Scholar] [CrossRef]
- Shanmugasundaram, M.; Radhika, M.; Murali, H.S.; Batra, H.V. Detection of Salmonella enterica serovar Typhimurium by selective amplification of fliC, fljB, iroB, invA, rfbJ, STM2755, STM4497 genes by polymerase chain reaction in a monoplex and multiplex format. World J. Microbiol. Biotechnol. 2009, 25, 1385–1394. [Google Scholar] [CrossRef]
- Ganesan, V.; Harish, B.N.; Menezes, G.A.; Parija, S.C. Detection of Salmonella in blood by PCR using iroB gene. J. Clin. Diagn. Res. JCDR 2014, 8, DC01–DC03. [Google Scholar] [CrossRef]
- Hantke, K.; Nicholson, G.; Rabsch, W.; Winkelmann, G. Salmochelins, siderophores of Salmonella enterica and uropathogenic Escherichia coli strains, are recognized by the outer membrane receptor IroN. Proc. Natl. Acad. Sci. USA 2003, 100, 3677–3682. [Google Scholar] [CrossRef]
- Crouch, M.L.V.; Castor, M.; Karlinsey, J.E.; Kalhorn, T.; Fang, F.C. Biosynthesis and IroC-dependent export of the siderophore salmochelin are essential for virulence of Salmonella enterica serovar Typhimurium. Mol. Microbiol. 2008, 67, 971–983. [Google Scholar] [CrossRef]
- Fischbach, M.A.; Lin, H.; Zhou, L.; Yu, Y.; Abergel, R.J.; Liu, D.R.; Raymond, K.N.; Wanner, B.L.; Strong, R.K.; Walsh, C.T.; et al. The pathogen-associated iroA gene cluster mediates bacterial evasion of lipocalin 2. Proc. Natl. Acad. Sci. USA 2006, 103, 16502–16507. [Google Scholar] [CrossRef]
- Raffatellu, M.; George, M.D.; Akiyama, Y.; Hornsby, M.J.; Nuccio, S.P.; Paixao, T.A.; Butler, B.P.; Chu, H.; Santos, R.L.; Berger, T.; et al. Lipocalin-2 resistance confers an advantage to Salmonella enterica serotype Typhimurium for growth and survival in the inflamed intestine. Cell Host Microbe 2009, 5, 476–486. [Google Scholar] [CrossRef]
- Uchiya, K.-I.; Nikai, T. Salmonella virulence factor SpiC is involved in expression of flagellin protein and mediates activation of the signal transduction pathways in macrophages. Microbiology 2008, 154, 3491–3502. [Google Scholar] [CrossRef]
- Ingram, J.P.; Brodsky, I.E.; Balachandran, S. Interferon-γ in Salmonella pathogenesis: New tricks for an old dog. Cytokine 2017, 98, 27–32. [Google Scholar] [CrossRef]
- Figueira, R.; Holden, D.W. Functions of the Salmonella pathogenicity island 2 (SPI-2) type III secretion system effectors. Microbiology 2012, 158, 1147–1161. [Google Scholar] [CrossRef]
- Wood, M.W.; Jones, M.A.; Watson, P.R.; Hedges, S.; Wallis, T.S.; Galyov, E.E. Identification of a pathogenicity island required for Salmonella enteropathogenicity. Mol. Microbiol. 1998, 29, 883–891. [Google Scholar] [CrossRef]
- Khoo, C.H.; Sim, J.H.; Salleh, N.A.; Cheah, Y.K. Pathogenicity and phenotypic analysis of sopB, sopD and pipD virulence factors in Salmonella enterica serovar Typhimurium and Salmonella enterica serovar Agona. Antonie Leeuwenhoek 2015, 107, 23–37. [Google Scholar] [CrossRef]
- Lawley, T.D.; Chan, K.; Thompson, L.J.; Kim, C.C.; Govoni, G.R.; Monack, D.M. Genome-wide screen for Salmonella genes required for long-term systemic infection of the mouse. PLoS Pathog. 2006, 2, e11. [Google Scholar] [CrossRef]
- Ashbolt, N.J.; Amézquita, A.; Backhaus, T.; Borriello, P.; Brandt, K.K.; Collignon, P.; Coors, A.; Finley, R.; Gaze, W.H.; Heberer, T.; et al. Human health risk assessment (HHRA) for environmental development and transfer of antibiotic resistance. Environ. Health Perspect. 2013, 121, 993–1001. [Google Scholar] [CrossRef]
- Hall, R.M. Integrons and gene cassettes: Hotspots of diversity in bacterial genomes. Ann. N. Y. Acad. Sci. 2012, 1267, 71–78. [Google Scholar] [CrossRef]
- Khan, A.A.; Ponce, E.; Nawaz, M.S.; Cheng, C.M.; Khan, J.A.; West, C.S. Identification and characterization of class 1 integron resistance gene cassettes among Salmonella strains isolated from imported seafood. Appl. Environ. Microbiol. 2009, 75, 1192–1196. [Google Scholar] [CrossRef]
- Ribeiro, V.B.; Lincopan, N.; Landgraf, M.; Franco, B.D.; Destro, M.T. Characterization of class 1 integrons and antibiotic resistance genes in multidrug-resistant Salmonella enterica isolates from foodstuff and related sources. Braz. J. Microbiol. 2011, 42, 685–692. [Google Scholar] [CrossRef]
- Zhang, H.; Shi, L.; Li, L.; Guo, S.; Zhang, X.; Yamasaki, S.; Miyoshi, S.-I.; Shinoda, S. Identification and characterization of class 1 integron resistance gene cassettes among Salmonella strains isolated from healthy humans in China. Microbiol. Immunol. 2004, 48, 639–645. [Google Scholar] [CrossRef]
- Doublet, B.; Boyd, D.; Mulvey, M.R.; Cloeckaert, A. The Salmonella genomic island 1 is an integrative mobilizable element. Mol. Microbiol. 2005, 55, 1911–1924. [Google Scholar] [CrossRef]
- Boucher, Y.; Labbate, M.; Koenig, J.E.; Stokes, H. Integrons: Mobilizable platforms that promote genetic diversity in bacteria. Trends Microbiol. 2007, 15, 301–309. [Google Scholar] [CrossRef]
- Partridge, S.R.; Kwong, S.M.; Firth, N.; Jensen, S.O. Mobile genetic elements associated with antimicrobial resistance. Clin. Microbiol. Rev. 2018, 31, e00088-17. [Google Scholar] [CrossRef]
- Gillings, M.; Boucher, Y.; Labbate, M.; Holmes, A.; Krishnan, S.; Holley, M.; Stokes, H.W. The evolution of class 1 integrons and the rise of antibiotic resistance. J. Bacteriol. 2008, 190, 5095–5100. [Google Scholar] [CrossRef]
- Vo, A.T.T.; Van Duijkeren, E.; Gaastra, W.; Fluit, A.C. Antimicrobial resistance, class 1 integrons, and genomic island 1 in Salmonella isolates from Vietnam. PLoS ONE 2010, 5, e9440. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention. Reports of Selected Salmonella Outbreak Investigations. US Centers for Disease Control and Prevention: Atlanta, GA, USA, 2018. Available online: https://www.cdc.gov/Salmonella/outbreaks.html (accessed on 18 October 2018).
- Mathole, M.; Muchadeyi, F.; Mdladla, K.; Malatji, D.; Dzomba, E.; Madoroba, E. Presence, distribution, serotypes and antimicrobial resistance profiles of Salmonella among pigs, chickens and goats in South Africa. Food Control 2017, 72, 219–224. [Google Scholar] [CrossRef]
- Zishiri, O.T.; Mkhize, N.; Mukaratirwa, S. Prevalence of virulence and antimicrobial resistance genes in Salmonella spp. isolated from commercial chickens and human clinical isolates from South Africa and Brazil. Onderstepoort J. Vet. Res. 2016, 83, 1–11. [Google Scholar] [CrossRef]
- Li, Q.; Cheng, W.; Zhang, D.; Yu, T.; Yin, Y.; Ju, H.; Ding, S. Rapid and sensitive strategy for Salmonella detection using an InvA gene-based electrochemical DNA sensor. Int. J. Electrochem. Sci. 2012, 7, 844–856. [Google Scholar]
- Hughes, A.L.; Shopland, S.; Wigley, P.; Bradon, H.; Leatherbarrow, A.H.; Williams, N.J.; Bennett, M.; De Pinna, E.; Lawson, B.; Cunningham, A.A.; et al. Characterisation of Salmonella enterica serotype Typhimurium isolates from wild birds in northern England from 2005–2006. BMC Vet. Res. 2008, 4, 4. [Google Scholar] [CrossRef]
- Pan, J.-C.; Ye, R.; Meng, D.-M.; Zhang, W.; Wang, H.-Q.; Liu, K.-Z. Molecular characteristics of class 1 and class 2 integrons and their relationships to antibiotic resistance in clinical isolates of Shigella sonnei and Shigella flexneri. J. Antimicrob. Chemother. 2006, 58, 288–296. [Google Scholar] [CrossRef]
- Andino, A.; Hanning, I. Salmonella enterica: Survival, colonization, and virulence differences among serovars. Sci. World J. 2015, 2015, 520179. [Google Scholar] [CrossRef]
- Ahmed, O.B.; Asghar, A.; Abd El-Rahim, I.; Al, H. Detection of Salmonella in food samples by culture and polymerase chain reaction methods. J. Bacteriol. Parasitol. 2014, 5, 187. Available online: https://www.longdom.org/open-access/detection-of-salmonella-in-food-samples-by-culture-and-polymerase-chain-reaction-methods-2155-9597.1000187.pdf (accessed on 6 October 2018).
- Klose, A.; Irsigler, H.; Jaeger, D.; Hafez, H.M.; Langkabel, N.; Bräutigam, L.; Fries, R. Comparison of methods for the detection of Salmonella in poultry. J. Appl. Poult. Res. 2014, 23, 403–408. [Google Scholar]
- Leonard, S.R.; Lacher, D.W.; Lampel, K.A. Acquisition of the lac operon by Salmonella enterica. BMC Microbiol. 2015, 15, 173. [Google Scholar] [CrossRef]
- Eswarappa, S.M.; Karnam, G.; Nagarajan, A.G.; Chakraborty, S.; Chakravortty, D. lac repressor is an antivirulence factor of Salmonella enterica: Its role in the evolution of virulence in Salmonella. PLoS ONE 2009, 4, e5789. [Google Scholar] [CrossRef]
- Hurley, D.; Hoffmann, M.; Muruvanda, T.; Allard, M.W.; Brown, E.W.; Martins, M.; Fanning, S. Atypical Salmonella enterica serovars in murine and human infection models: Is it time to reassess our approach to the study of salmonellosis? BioRxiv 2016, 058610. Available online: https://www.biorxiv.org/content/biorxiv/early/2016/06/13/058610.1.full.pdf (accessed on 10 October 2018).
- McDonough, P.L.; Shin, S.J.; Lein, D.H. Diagnostic and public health dilemma of lactose-fermenting Salmonella enterica serotype Typhimurium in cattle in the northeastern United States. J. Clin. Microbiol. 2000, 38, 1221–1226. [Google Scholar]
- Jamshidi, A.; Kalidari, G.A.; Hedayati, M. Isolation and identification of Salmonella Enteritidis and Salmonella Typhimurium from the eggs of retail stores in Mashhad, Iran. using conventional culture method and multiplex PCR assay. J. Food Saf. 2010, 30, 558–568. [Google Scholar]
- Manning, J.; Gole, V.; Chousalkar, K. Screening for Salmonella in backyard chickens. Prev. Vet. Med. 2015, 120, 241–245. [Google Scholar] [CrossRef]
- Abatcha, M.G.; Effarizah, M.E.; Rusul, G. Prevalence, antimicrobial resistance, resistance genes and class 1 integrons of Salmonella serovars in leafy vegetables, chicken carcasses and related processing environments in Malaysian fresh food markets. Food Control 2018, 91, 170–180. [Google Scholar] [CrossRef]
- Ahmed, A.M.; Shimamoto, T. Genetic analysis of multiple antimicrobial resistance in Salmonella isolated from diseased broilers in Egypt. Microbiol. Immunol. 2012, 56, 254–261. [Google Scholar] [CrossRef]
- Magwedere, K.; Rauff, D.; De Klerk, G.; Dziva, F.; Keddy, K.H. Incidence of nontyphoidal Salmonella in food-producing animals, animal feed, and the associated environment in South. Africa, 2012–2014. Clin. Infect. Dis. 2015, 61, S283–S289. [Google Scholar] [CrossRef]
- Dione, M.M.; Ikumapayi, U.; Saha, D.; Mohammed, N.I.; Adegbola, R.A.; Geerts, S.; Ieven, M.; Antonio, M. Antimicrobial resistance and virulence genes of non-typhoidal Salmonella isolates in The Gambia and Senegal. J. Infect. Dev. Ctries. 2011, 5, 765–775. [Google Scholar] [CrossRef]
- Gyles, C.; Boerlin, P. Horizontally transferred genetic elements and their role in pathogenesis of bacterial disease. Vet. Pathol. 2014, 51, 328–340. [Google Scholar] [CrossRef]
- Gillings, M.R. Integrons: Past, present, and future. Microbiol. Mol. Biol. Rev. 2014, 78, 257–277. [Google Scholar] [CrossRef]
- Domingues, S.; da Silva, G.J.; Nielsen, K.M. Integrons: Vehicles and pathways for horizontal dissemination in bacteria. Mob. Genet. Elem. 2012, 2, 211–223. [Google Scholar] [CrossRef]
- Threlfall, E.J. Antimicrobial drug resistance in Salmonella: Problems and perspectives in food-and water-borne infections. FEMS Microbiol. Rev. 2002, 26, 141–148. [Google Scholar] [CrossRef]
- Jiang, C.; Shaw, K.S.; Romeo, C.; Blythe, D.; Mitchell, C.; Murtugudde, R.; Sapkota, A.R.; Sapkota, A. Climate change, extreme events and increased risk of salmonellosis in Maryland, USA: Evidence for coastal vulnerability. Environ. Int. 2015, 83, 58–62. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Map of South Africa showing the geographic locations of the farms where samples were collected for this study.
Figure 1.
Map of South Africa showing the geographic locations of the farms where samples were collected for this study.
Figure 2.
Molecular detection of Salmonella enterica using PCR methods. (A) Salmonella spp. invA gene amplicon (284 bp) visualized on 1.5% agarose gel. Lane MW represents the molecular weight marker is the 100 bp DNA ladder (New England Biolabs, Beijing, China). Lane PC represents the positive control and lanes one to eight represents test samples. Lane NC represents the negative control (NC). (B) The 606 bp amplicon of iroB gene on 1.5% agaroe gel. Lane PC represents the positive control and lanes one to five represent test samples. Lane NC represents the negative control (NC).
Figure 2.
Molecular detection of Salmonella enterica using PCR methods. (A) Salmonella spp. invA gene amplicon (284 bp) visualized on 1.5% agarose gel. Lane MW represents the molecular weight marker is the 100 bp DNA ladder (New England Biolabs, Beijing, China). Lane PC represents the positive control and lanes one to eight represents test samples. Lane NC represents the negative control (NC). (B) The 606 bp amplicon of iroB gene on 1.5% agaroe gel. Lane PC represents the positive control and lanes one to five represent test samples. Lane NC represents the negative control (NC).
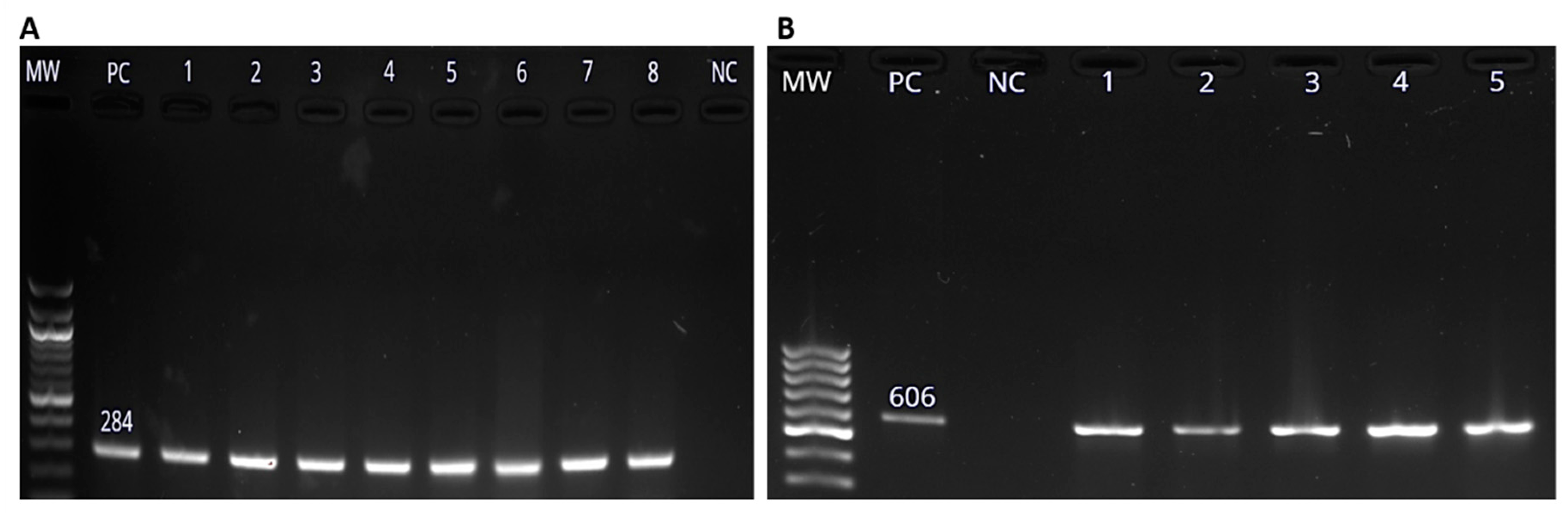
Figure 3.
Molecular detection of Salmonella enterica virulence genes using PCR methods. (A). The 309 bp spiC gene amplicon, (B) pipD gene amplicons (350 bp), and (C) The 569 bp amplicon of int1 gene as visualized on 1.5% agarose gel. Lanes labelled MW, PC and NC are the 100 bp molecular weight marker (New England Biolabs, Beijing, China), positive control (PC) and negative control (NC) respectively. Numbered lanes represent the tested isolates.
Figure 3.
Molecular detection of Salmonella enterica virulence genes using PCR methods. (A). The 309 bp spiC gene amplicon, (B) pipD gene amplicons (350 bp), and (C) The 569 bp amplicon of int1 gene as visualized on 1.5% agarose gel. Lanes labelled MW, PC and NC are the 100 bp molecular weight marker (New England Biolabs, Beijing, China), positive control (PC) and negative control (NC) respectively. Numbered lanes represent the tested isolates.
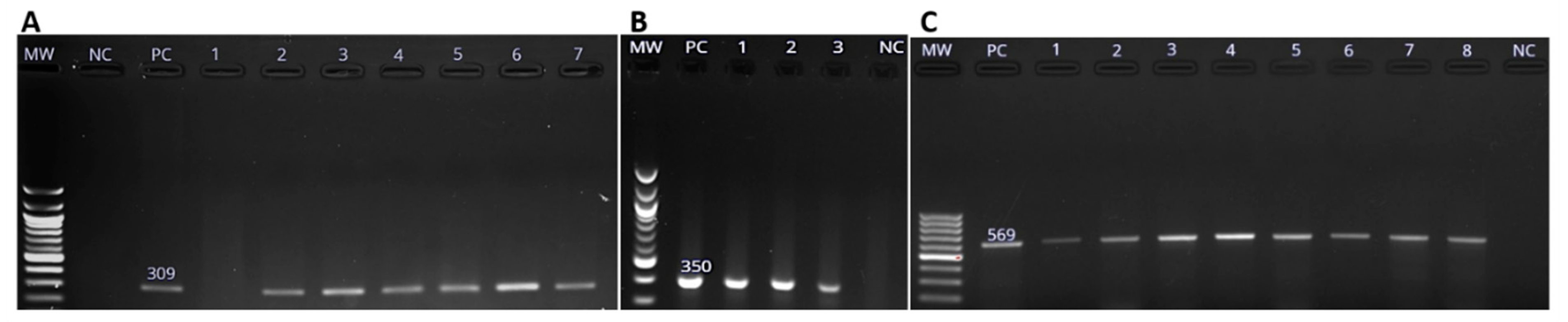
Figure 4.
The distribution of Salmonella virulence genes in animal host species in this study.
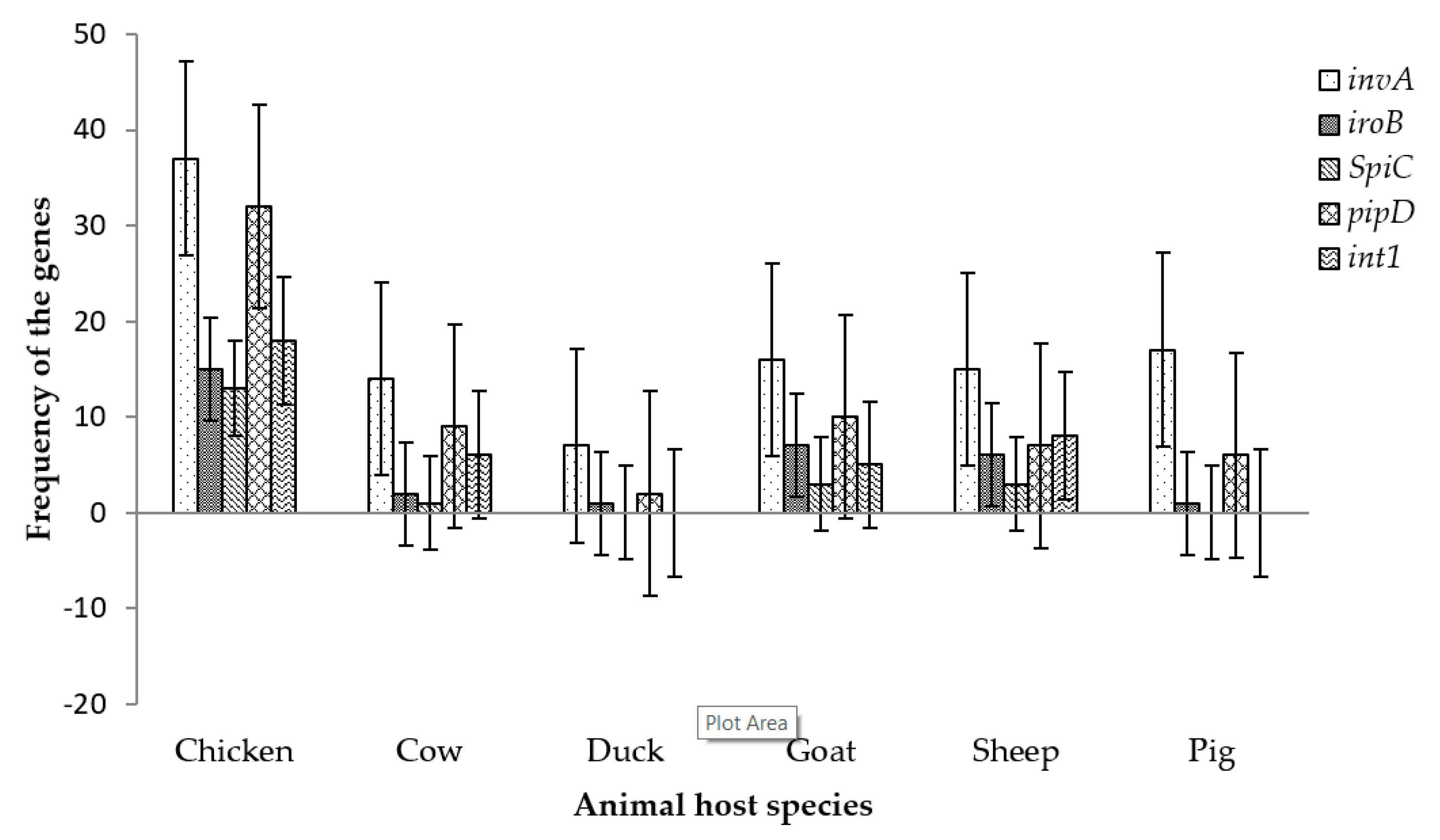
Figure 5.
Total number of samples collected and number of positive samples after culturing on Xylose-Lysine-Deoxycholate (XLD) agar and screening of virulence genes.
Figure 5.
Total number of samples collected and number of positive samples after culturing on Xylose-Lysine-Deoxycholate (XLD) agar and screening of virulence genes.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Number of samples collected from livestock farms in Flagstaff, Verulam and South Coast in South Africa in 2018.
Table 1.
Number of samples collected from livestock farms in Flagstaff, Verulam and South Coast in South Africa in 2018.
| Animal Host | Flagstaff | Verulam | South Coast | Total | Positive Samples (%) | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Oral | Fecal | Feed | Soil | Water | Oral | Fecal | Feed | Soil | Water | Oral | Fecal | Feed | Soil | Water | |||
| Chicken | 0 | 0 | 0 | 0 | 0 | 0 | 24 | 0 | 0 | 0 | 40 | 40 | 0 | 5 | 5 | 114 | 10.25 |
| Ducks | 0 | 10 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 10 | 1.94 |
| Cow | 0 | 5 | 0 | 5 | 0 | 0 | 20 | 0 | 0 | 0 | 0 | 10 | 0 | 5 | 5 | 50 | 3.88 |
| Goats | 10 | 9 | 0 | 6 | 6 | 0 | 10 | 0 | 0 | 0 | 17 | 16 | 0 | 0 | 5 | 79 | 4.43 |
| Sheep | 12 | 10 | 0 | 6 | 0 | 0 | 10 | 0 | 0 | 0 | 4 | 8 | 0 | 0 | 0 | 50 | 4.15 |
| Pigs | 17 | 17 | 9 | 9 | 6 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 58 | 4.71 |
| Total | 39 | 51 | 9 | 26 | 12 | 0 | 64 | 0 | 0 | 0 | 61 | 74 | 0 | 10 | 15 | 361 | 29.36 |
Table 2.
The effect of location as a predictor for the presence iroB, pipD and int1 as measured by the binary logistic regression.
Table 2.
The effect of location as a predictor for the presence iroB, pipD and int1 as measured by the binary logistic regression.
| p-Value | Odds Ratio | 95% C.I. for Odds Ratio | |
|---|---|---|---|
| iroB | |||
| Location | 0.005 | ||
| Verulam | 0.007 | 5.429 | (1.577, 18.686) |
| South coast | 0.881 | 0.924 | (0.330, 2.587) |
| pipD | |||
| Location | 0.007 | ||
| Verulam | 0.006 | 19.991 | (2.330, 171.530) |
| South coast | 0.994 | 0.994 | (0.237, 4.173) |
| Int1 | |||
| Location | 0.022 | ||
| Verulam | 0.042 | 8.053 | (1.801, 59.968) |
| South coast | 0.409 | 0.564 | (0.145, 2.193) |
Table 3.
Pearson’s correlation analysis measuring the strengths of the relationships between the virulence genes and between using XLD and invA for the detection Salmonella spp.
Table 3.
Pearson’s correlation analysis measuring the strengths of the relationships between the virulence genes and between using XLD and invA for the detection Salmonella spp.
| Variables | Pearson’s Correlation (p-Value) |
|---|---|
| XLD and invA | 0.402 (0.000) |
| iroB and spiC | 0.407 (0.000) |
| iroB and pipD | 0.258 (0.008) |
| iroB and int1 | 0.294 (0.002) |
| spiC and pipD | 0.357 (0.000) |
| spiC and int1 | 0.102 (0.298) * |
| pipD and int1 | 0.325 (0.001) |
* The correlation is not significant as p > 0.05.
Table 4.
The p-values obtained from Fischer’s exact test investigating the association between the variables (location, animal host, sampling season, sample material) and the virulence genes.
Table 4.
The p-values obtained from Fischer’s exact test investigating the association between the variables (location, animal host, sampling season, sample material) and the virulence genes.
| Variable | iroB | spiC | pipD | Int1 |
|---|---|---|---|---|
| Location | 0.002 | 0.000 | 0.000 | 0.000 |
| Animal host | 0.037 | 0.019 | 0.002 | 0.000 |
| Sampling season | 0.001 | 0.000 | 0.000 | 0.000 |
| Sample material | 0.345 * | 0.467 * | 0.365 * | 0.004 |
* There is no significant association as the p > 0.05.
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Mthembu, T.P.; Zishiri, O.T.; El Zowalaty, M.E. Detection and Molecular Identification of Salmonella Virulence Genes in Livestock Production Systems in South Africa. Pathogens 2019, 8, 124. https://doi.org/10.3390/pathogens8030124
AMA Style
Mthembu TP, Zishiri OT, El Zowalaty ME. Detection and Molecular Identification of Salmonella Virulence Genes in Livestock Production Systems in South Africa. Pathogens. 2019; 8(3):124. https://doi.org/10.3390/pathogens8030124
Chicago/Turabian StyleMthembu, Thobeka P., Oliver T. Zishiri, and Mohamed E. El Zowalaty. 2019. "Detection and Molecular Identification of Salmonella Virulence Genes in Livestock Production Systems in South Africa" Pathogens 8, no. 3: 124. https://doi.org/10.3390/pathogens8030124
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.